
Abstract
Neural system development is one of the most important stages of embryogenesis. Perturbations in this crucial process due to genetic and environmental risk factors cause neural tube defects and other central nervous system diseases. We investigated the effects of prenatal exposure to 900-MHz electromagnetic field (EMF) on the spinal cord. Pregnant rats were exposed to 900-MHz EMF for 1 h/day from E13.5 until birth. Six pups from the control and EMF groups were sacrificed at postnatal day 32, and the upper thoracic region of the spine was removed and processed for histological procedures. For histopathological analyses, hematoxylin&eosin staining and, for stereological analyses and the quantitation of motor neurons, cresyl violet staining was performed. H3K27me3 levels were determined via immunofluorescence staining. Histopathological analysis identified structural alterations of ependymal cells, enlarged central canals, as well as degenerated and shrunken motor neurons in the EMF group, while the control group tissues had normal appearances. We also observed enrichment of H3K27me3 in the ependymal cells and the motor neurons in the spinal cord of the control group rats, while the EMF group had low levels of H3K27me3 staining. Our results suggest that the loss of H3K27me3 signals might correlate with reduced neuronal stem cell potential in the EMF group and result in anatomical and structural differences in the spinal cord. This study provided a comprehensive histopathological analysis of the spinal cord after prenatal EMF exposure and offered an H3K27me3-dependent molecular explanation for the detrimental effects of EMF exposure on the spine.
Introduction
Modernization of everyday life with the advancements in technology has caused an unprecedented increase in the emittance of artificial electromagnetic fields (EMFs). Humans and animals are often exposed to EMFs with varying frequencies emitted by devices such as power cables for electrical supplies, computer or TV screens, as well as mobile phones and broadcasting towers (Gye and Park, 2012). Whether prolonged exposure to artificial EMFs has any effect on public health has been a major area of interest to the scientific community. In line with this, several studies have evaluated the biological changes triggered by EMF at the cellular, molecular, and organismal levels in relation to human disease. An extensive number of epidemiological and experimental studies linked EMF exposure with increased risk of cancer, neurodegenerative diseases, reproductive toxicity, and disabilities in learning behavior and motor skills (Brainard et al., 1999; İkinci et al., 2013; London et al., 1991; Odacı et al., 2013; Pourlis, 2009; Sobel et al., 1996).
Development of the neural system is one of the earliest events in the embryonic period and is completed after birth. Neurulation is the stage of embryonic development that results in the formation of neural tube in vertebrates, which will give rise to future brain and spinal cord to form the central nervous system (CNS) (Detrait et al., 2005). In human embryos, neural tube closure occurs 3–4 weeks after fertilization (Blom, 2009). Primary neurulation involves shaping and folding of the neuroepithelium, generating the entire neural tube (Copp et al., 2003). Cells of the neuroepithelium form the ventricular layer, which will then give rise to the majority of the cells in the adult CNS (Moore and Persuad, 1993). It is widely believed that undifferentiated adult neural stem cells are responsible for the maintenance of the nervous system later on in life. The ependymal region, which is largely occupied by the ependymal cells that surround the central canal, is considered as a pool of neural stem and progenitor cells (Mladinic et al., 2014). Studies have indicated that these cells can readily proliferate upon spinal damage, migrate to the site of injury, and differentiate (Hugnot and Franzen, 2011; Lacroix et al., 2014; Sabelström et al., 2014).
Neural system development is one of the most important periods during a pregnancy since it is an early developmental stage that could be influenced by external factors. For instance, lack of sufficient folate in the maternal diet results in neural tube defects (Hall and Solehdin, 1998). EMF exposure is known to affect the CNS, with possibly even more detrimental outcomes during the prenatal period; both because the embryo is more vulnerable to such exposures and neural development is a critical phase, significantly affecting the adult CNS. Therefore, in this study, we aimed to evaluate the histopathological, cellular, and epigenetic alterations in the spinal cord induced by prenatal exposure to EMF.
Materials and methods
Animal experiments and EMF exposure
In these experiments, 16 Female Sprague Dawley rats (180–250 g) that exhibited regular estrous cycles were mated with male rats. Vaginal plugs were checked and designated as day 0 of pregnancy. The pregnant female rats were randomly separated into two groups (control and EMF) and kept in separate cages. The control group received no treatment throughout their pregnancy. From E13.5 until parturition, the EMF group was exposed to 900-MHz EMF for 1 h per day, every day at the same time. Control and EMF pregnant female rats 13 and 10 female gave birth, respectively, and the experiments were carried out using 6 randomly selected pups per group (Keleş et al., 2019a).
Animals were held at a mean temperature of 22 ± 2°C and humidity of 50% ± 5 by an automated system. Also, they were held in an automatically adjusted 12-h light and dark cycle. Standard rat palette chow (Bayramoğlu Yem and Un Sanayi Tic. A.Ş., Erzurum, Turkey) and drinking water were used. The water and feed of the animals were monitored daily.
The system for EMF exposure was set up and the procedure was applied as described previously (İkinci et al., 2013; Keleş et al., 2019a; Odacı et al., 2013). EMF intensity measurements were carried out at nine distinct points at the bottom of the cage (Figure 1) with/without rats inside, using a C.A 43 isotropic electrical field intensity meter (Chauvin Arnoux Group, Paris, France). The rats were exposed to an average 10-V/m electric field (0.265 W/m2 power intensity). All animal procedures were approved by the Institutional Ethics Committee at Niğde Ömer Halisdemir University (Date: September 10, 2019, Protocol number: 2019/27) and were carried out in accordance with the principles of the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996).
Representation of the electromagnetic field cage. Locations of the nine points used to measure electromagnetic field intensity on the cage floor are marked by asterisks.
Preparation of the tissues
All pups were anesthetized via ketamine (50 mg/kg) and sacrificed by decapitation at postnatal day 32 in order to eliminate the effect of external stress factors such as weaning on neural development. The upper thoracic region of the spine was removed and fixed in 10% formaldehyde overnight. After decalcification and routine histological procedures, the tissues were embedded into paraffin blocks (Odacı et al., 2013). Tissue sections (5 µm in thickness) were prepared using a Leica Biosystems RM 2245 Microtome (Germany).
Histological evaluation
For the histopathological evaluation of the tissues, sections were stained with hematoxylin&eosin (H&E) following routine procedures. The analysis of the stained sections was performed using a light microscope (Olympus, BX53, Japan) with an attached digital camera (DP 80, Olympus, Japan) and Cellsens standard program (version 1.17).
Stereological (volumetric) analyses
In order to perform stereological analyses, five tissue sections per animal (10 µm in thickness) were cut and stained with cresyl violet (CV). The section sampling fraction was 1/10 (Table 1) (Aktürk et al., 2014). Analyses were conducted using the Stereology Analyses System available at the Stereology Laboratory of the Histology and Embryology Department, Medical Faculty, Karadeniz Technical University (Trabzon, Turkey). Volumetric evaluation of the spinal cord tissues was performed using the Cavalieri principle (Aktürk et al., 2017; Keleş, 2019; Yahyazadeh and Altunkaynak, 2019).
Stereological analysis sampling strategy used when measuring spinal cords and counting motor neuron (10 tissue sections per animal).

Quantitation of motor neurons
There are several methods for the determination of cell numbers (Keleş, 2019). In this study, 10-µ sections of the upper thoracic region of the spine (three sections per animal) stained with CV were evaluated under a light microscope, and the motor neurons were counted using Cellsens standard program.
Immunofluorescence staining
Tissue sections were deparaffinized, and heat-induced antigen retrieval in 10-mM sodium citrate buffer and permeabilization steps in 0.4% Triton X-100 in Tris-buffered saline (TBS) were performed. Sections were blocked with 4% bovine serum albumin-TBS-Triton X for 1 h at room temperature and incubated with the α-H3K27me3 primary antibody (CST #9733) O/N at 4°C. After washing in TBS, sections were incubated with fluorescein isothiocyanate-labeled secondary antibody (Proteintech #SA00003-2). The sections were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and mounted in Fluoroshield mounting medium (Abcam #ab104139) and then sealed and stored at 4°C in the dark. Microscopic analysis was performed using the same microscope and software in the “Histological evaluation” section. Scale bars were inserted using Image J.
Statistical analyses
Statistical analyses were carried out using Statistical Packege for the Social Sciences v. 22. Data distribution was assessed using the Shapiro–Wilk normality test as well as by drawing a histogram. As the volume of the white matter and the neuron count displayed normal distribution, Student’s t-test was performed. For the volume of the gray matter, the Mann–Whitney U Test was preferred as the data were not normally distributed. The values were expressed as mean ± SD. Values of p <0.05 were considered statistically significant.
Results
Histopathological alterations within the spinal cord
Our histopathological analysis revealed both anatomical and structural differences within the spinal cord tissue between the control and the EMF groups. The control group had normal appearances of the central canal and motor neurons, and the ependymal cells had a regular columnar shape (Figure 2). On the other hand, the central canal was enlarged, the motor neurons were structurally degenerated and shrunken, and the ependymal cells were flattened into a more cubical shape in the EMF group. There was no significant structural difference in the neuroglial cells of the white matter between the groups (Figure 3).
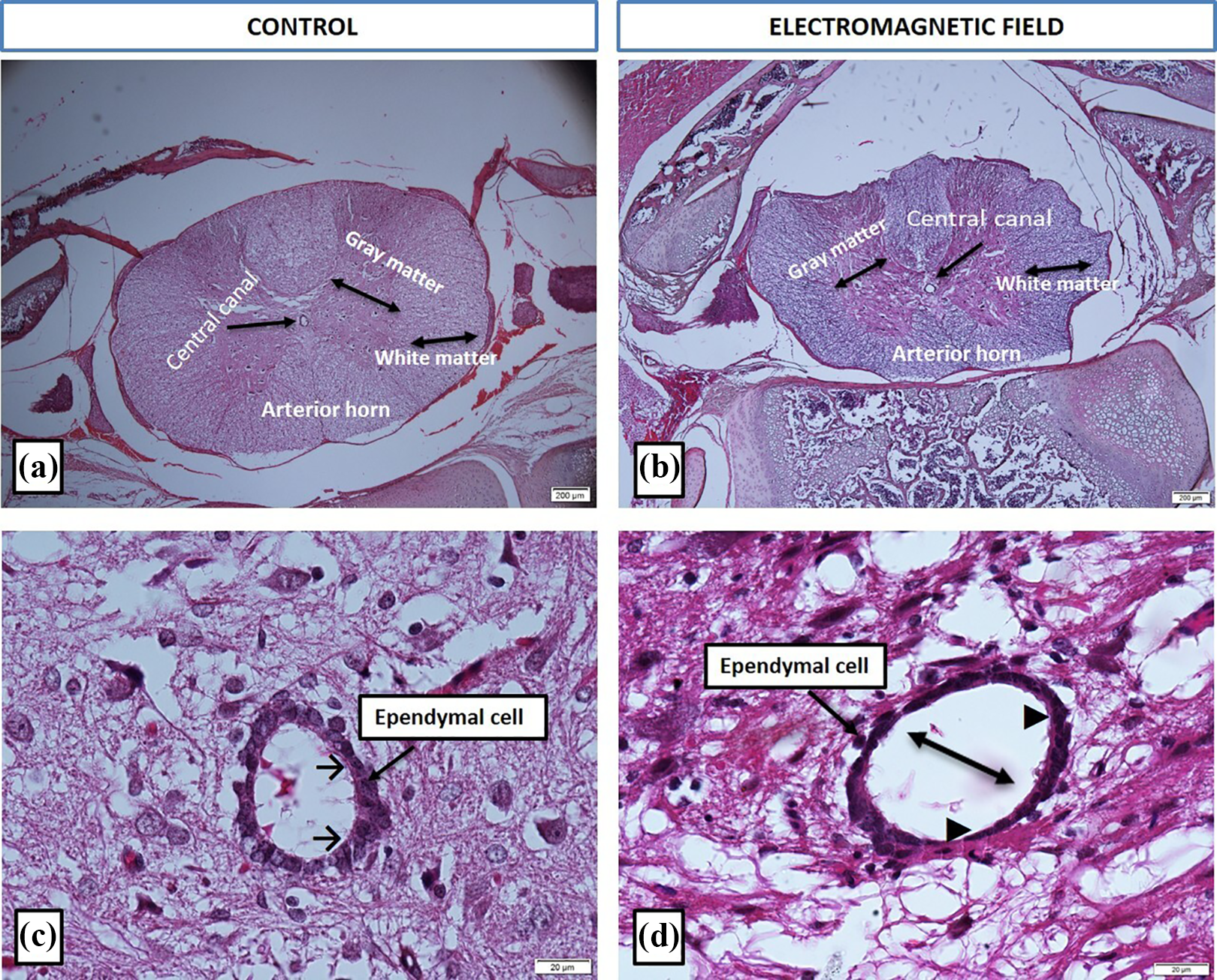
Representative micrographs of spinal cords of the control and the electromagnetic field groups (hematoxylin&eosin). General view of the spinal cord (a and b, ×4) and the ependymal cells (c and d, ×60). Headed arrow (→): normal ependymal cells; black arrow (▴): flattened ependymal cells; double headed arrow (↔): central canal expansion.
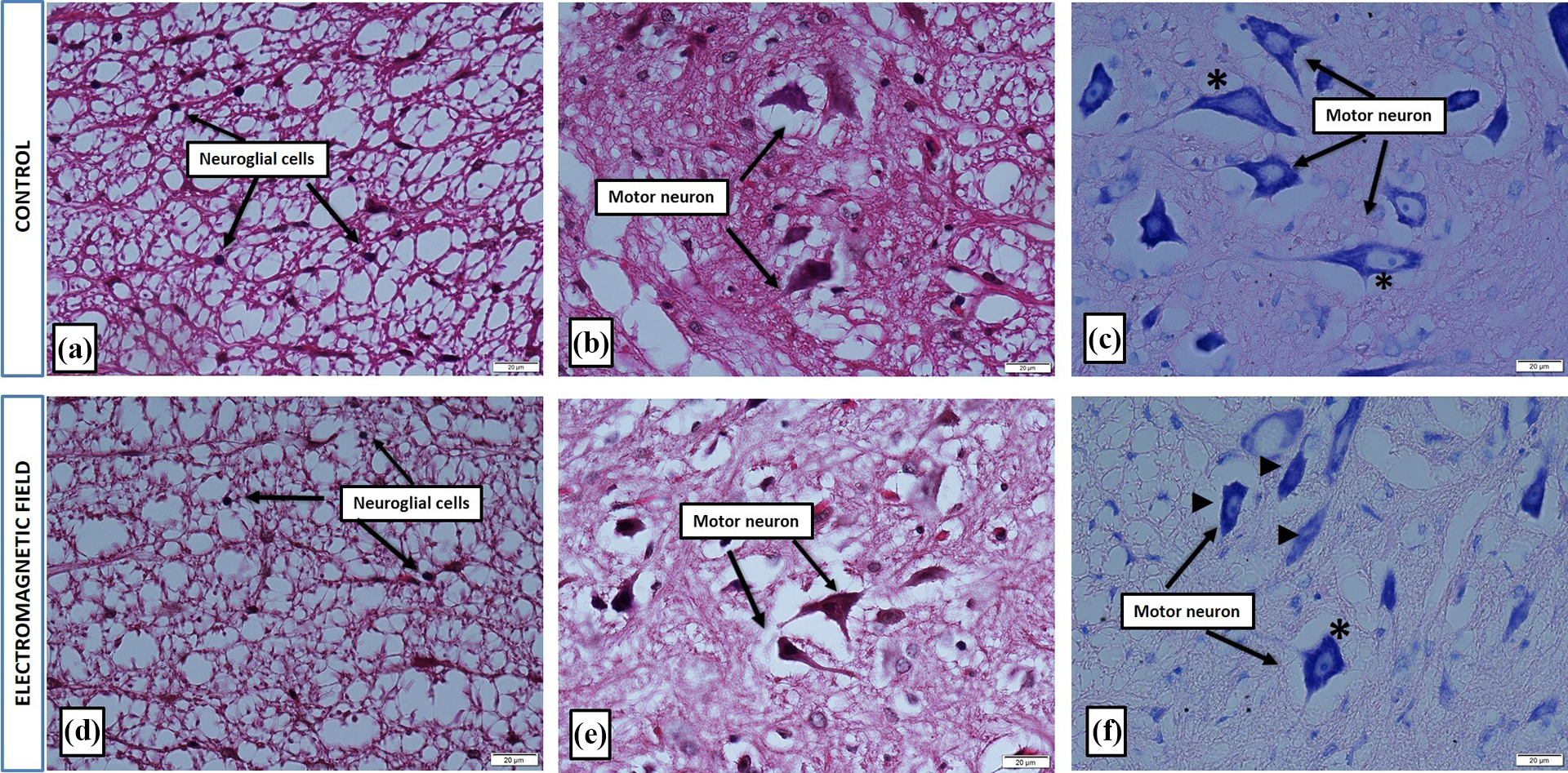
Representative micrographs of spinal cords of the control and the electromagnetic field groups ×60, stained with hematoxylin&eosin (a, b, d, and e) and cresyl violet (c and f) stained. Star (*) indicates normal motor neurons, while the black arrow (▴) marks degenerated and shrunken motor neurons.
In addition to the structural disruptions at the cellular level in the EMF group, it was also strikingly evident from the H&E-stained tissue sections that the total area of the spinal cord was smaller. In order to evaluate whether this was a statistically significant difference, we performed volumetric analysis of the spinal cord, which indicated that the volumes of both the gray and the white matter in the EMF group were significantly decreased in comparison with their respective counterparts in the control group (p = 0.002 and p = 0.001, respectively). Furthermore, we quantified the number of motor neurons in both groups via stereological approaches and found a statistically significant decrease in the EMF group (p = 0.001, Table 2).
Statistical data of the white and gray matter volume and motor neurons of control and electromagnetic field groups.a
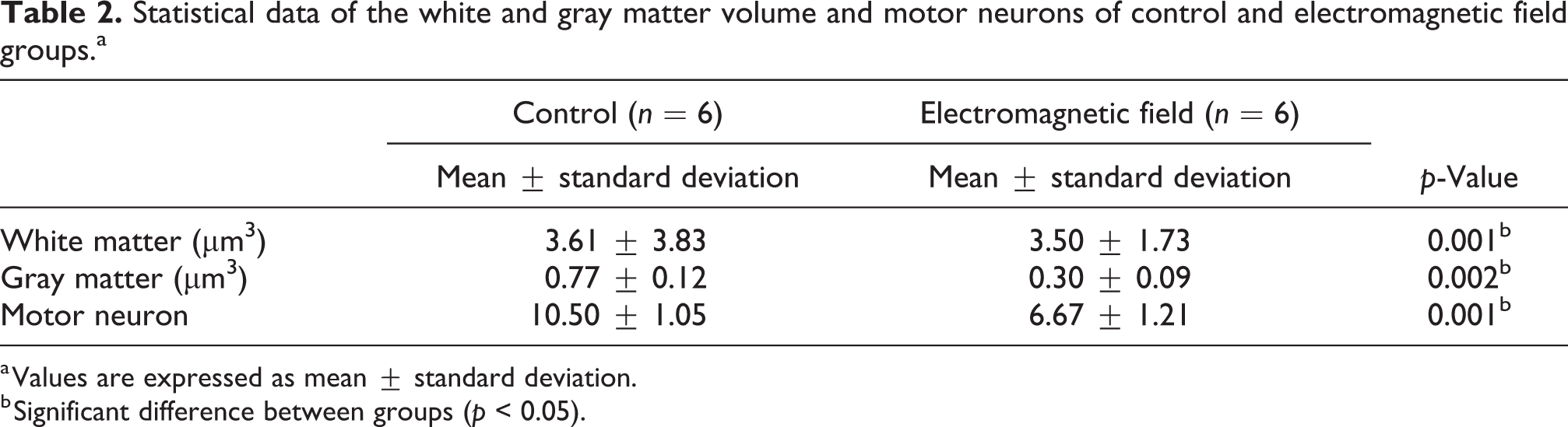
a Values are expressed as mean ± standard deviation.
b Significant difference between groups (p < 0.05).
Distribution of the H3K27me3 marker in the spinal cord
After establishing that EMF exposure caused histopathological alterations in the spinal cord in comparison with the control group, we investigated whether EMF exposure would induce epigenetic alterations as well. Tri-methylation of histone H3 at lysine 27 (H3K27me3) is an important epigenetic marker of neural commitment and differentiation of embryonic stem cells (Juan et al., 2016) and is associated with several regulatory mechanisms in the spinal cord. Therefore, we performed immunofluorescence staining of the spinal cord tissue sections using H3K27me3-specific antibodies. In the control group, we observed a widespread enrichment of H3K27me3-specific signals throughout the spinal cord (Figure 4, upper left). More interestingly, the ependymal cells around the central canal had high levels of the H3K27me3 marker as well. The signal localized to the cell nuclei as evident by DAPI staining, as expected (Figure 4, lower left). On the other hand, tissue sections of the EMF group lacked specific enrichment of H3K27me3 and rather exhibited an unspecific faint signal that resembled autofluorescence (Figure 4, right panels). We also found that the motor neurons in the control group had high levels of H3K27me3, which was missing in the EMF group (Figure 5).
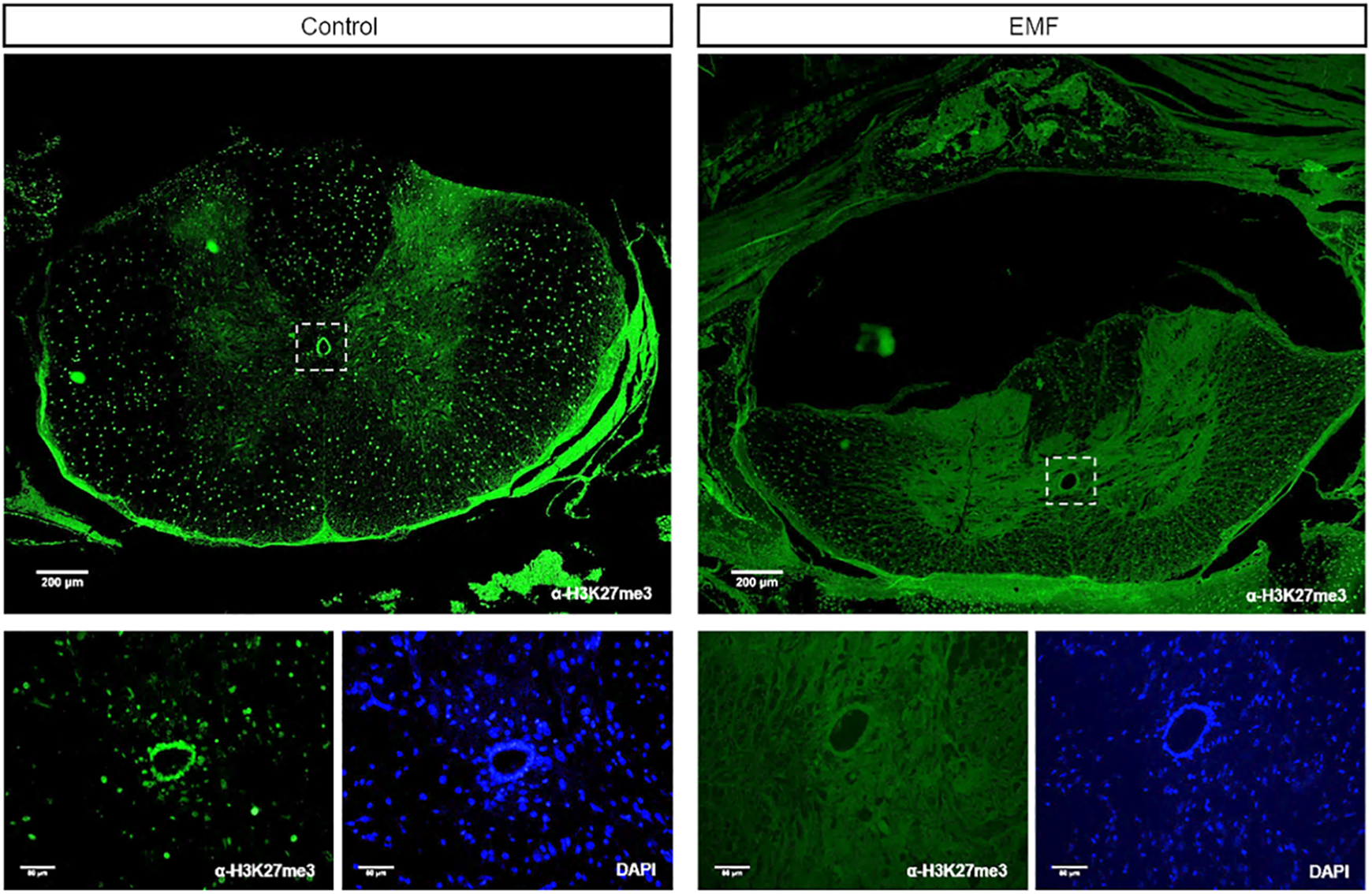
Immunofluorescence labeling of the rat spinal cord. Tissue sections were stained with H3K27me3 antibody (green) and imaged at ×4 (upper panels) and ×20 (lower panels) magnification. Zoomed-in views of the ependymal cells (marked by dashed squares) are presented in the lower panels. Control group spinal cords, especially the ependymal cells, show high levels of H3K27me3-specific signal. DNA stained with DAPI (blue) marks the nucleus. DAPI: 4′,6-diamidino-2-phenylindole; EMF: electromagnetic field.
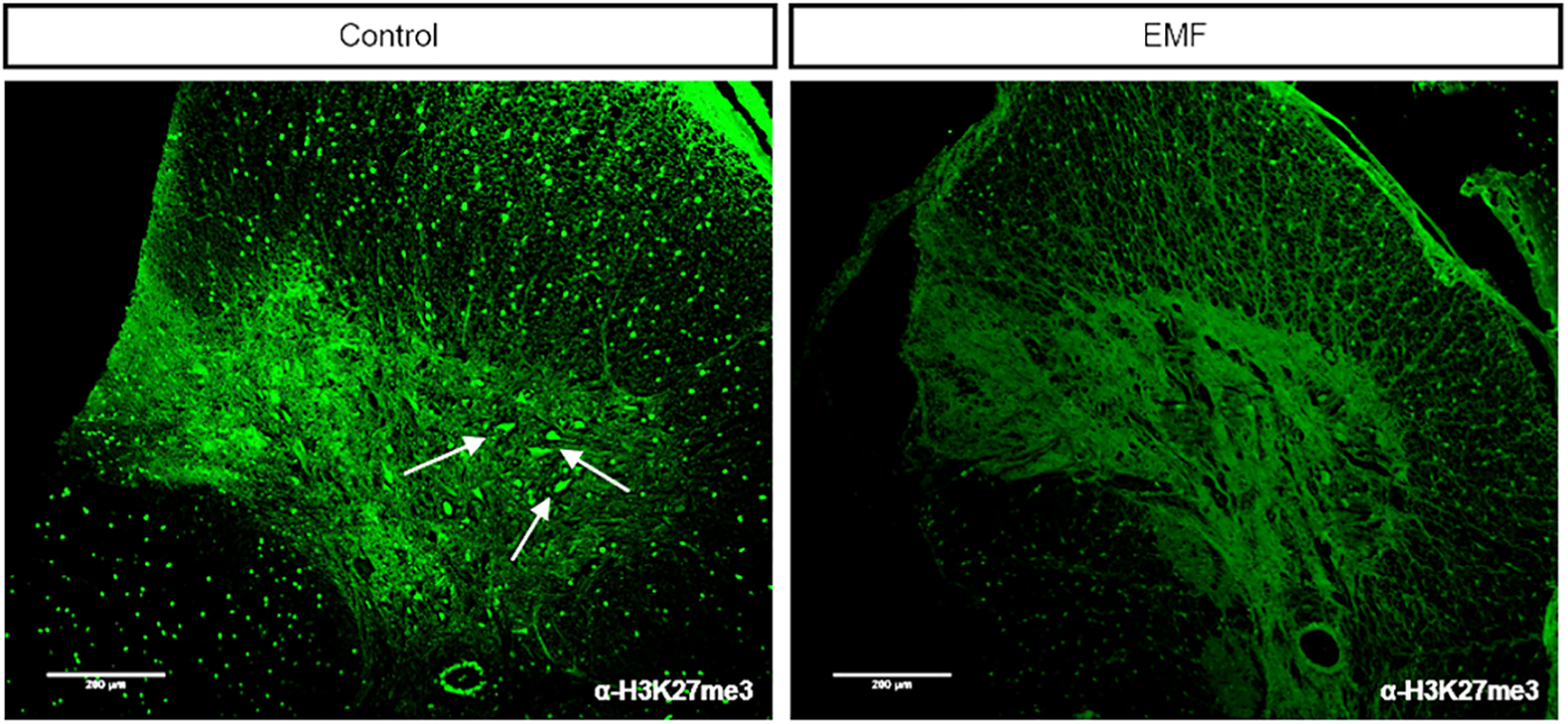
Immunofluorescent staining of the rat spinal cord tissue sections indicates specific enrichment of H3K27me3 signal in motor neurons (marked by arrows) in the control group, while the H3K27me3-specific signal was not detected in the motor neurons of the EMF group (×10). EMF: electromagnetic field.
Discussion
Neural development is an important stage of embryonic development. Failure to complete this crucial process correctly causes neural tube defects, which in most severe cases are often lethal. Several risk factors, both genetic and environmental, are associated with CNS diseases. Therefore, in this study, we investigated the effects of prenatal EMF exposure on the spinal cord.
Our results showed that EMF exposure caused structural disruptions and regression in the spinal cord in comparison with the control group. Similar to our findings, previous studies on the adult spinal cord suggested degeneration and decreased number of motor neurons (Yahyazadeh and Altunkaynak, 2019). EMF exposure in adults has also been linked with decreased oxidative stress and inflammation (Wang et al., 2019) as well as enhanced neurological function (Li et al., 2019) upon spinal cord injury. Prenatal EMF exposure is generally associated with detrimental effects on development and proposed as a teratogenic agent affecting CNS (Bas et al., 2009). Likewise, studies reported decreased heart weight and ventricle wall thickness in developing chick embryos after EMF exposure (Pawlak et al., 2018), lower birth weight (Mortazavi et al., 2013), pathological changes in the spleen (Keleş et al., 2019b) and cerebellum (Odacı et al., 2016), as well as increased motor behavior (Odacı et al., 2013) in rats. Our evaluation of the structural changes in the spinal cord showed central canal enlargement and decreased gray and white matter volume in the EMF group. Similar to our findings, in a study conducted, it was stated that there is an enlargement in the central canal as a result of EMF applied during prenatal period (Keleş, 2020). The difference in volume strongly suggests that the prenatal exposure to EMF interferes with the spinal cord development and results in the structural malformations that we observed. On the other hand, a study with contradictory findings in the adult spinal cord reported neuronal degeneration while the volume did not significantly change (Yahyazadeh and Altunkaynak, 2019).
Epigenetic mechanisms are central players of transcriptional regulation with significant roles in the development and function of the CNS, affecting cell fate decisions, neuronal differentiation, memory, and behavior (Feng et al., 2007). Posttranslational histone modifications, such as the H3K27me3, regulate critical cellular mechanisms in the CNS. Studies have shown that H3K27me3 controls dorsal interneuron generation via regulating bone morphogenetic proteins activity (Akizu et al., 2010). H3K27me3 levels increase during the differentiation of oligodendrocyte progenitors into oligodendrocytes, and H3K27me3 is required for their proper myelination (Wang et al., 2020), while reduced H3K27me3 levels are associated with neural tube defects (Yu et al., 2019). Furthermore, reduced levels of H3K27me3 are suggested as an epigenetic biomarker in ependymomas, which are tumors of the brain or any part of the spinal cord (Zhang and Kuo, 2017). Likewise, mutations resulting in the missense substitution of histone H3 lysine 27 to methionine (K27M) are often found in spinal cord gliomas (Chai et al., 2020).
In addition to these important roles of H3K27me3 in neural development, it is also associated with neural commitment and differentiation of embryonic stem cells (Juan et al., 2016). Adult CNS is comprised of cells that have differentiated from the embryonic neural stem cells during development, and undifferentiated adult neural stem cells are suggested to play a critical role for the maintenance of the nervous system. Therefore, it is possible that the structural regression in the spinal cord of EMF group rats resulted due to disruptions of their neural stem cells or the lack of neuronal differentiation. Our finding that many cells, including the cells of the ependymal region, where the adult neural stem cells are believed to form a niche around (Mladinic et al., 2014), were specifically stained with α-H3K27me3 antibody in the control group, while the spinal cord of the EMF group lacked this specific H3K27me3 staining, is in support of this. H3K27me3 is considered as an epigenetic indicator of stem cell potential (Burney et al., 2013); therefore, we could speculate that the H3K27me3-positive cells in the control group are neural progenitor cells, possibly at different developmental stages, which ensure the normal development of the spinal cord. In line with this, due to the loss of H3K27me3 signal and the correlated stem cell potential in the EMF group, either the normal development of the spinal cord or its maintenance fails. From another perspective, H3K27me3 levels could indicate an injury-like state in the EMF-exposed group, as lower H3K27me3 levels were reported in a previous study after spinal injury in a mouse model (Descalzi et al., 2015).
H3K27me3 is also enriched in a subset of developmental genes (Hox family genes) during differentiation of stem cells into motor neurons (Murre, 2015) and is a marker of motor neurons at the progenitor stage (Mazzoni et al., 2013). Our identification of H3K27me3 positive motor neurons in the control but not in the EMF group provided additional support for neuronal regeneration by the adult stem cells in the spinal cord, which seems to be impaired in the EMF group. Lastly, we observed an increased background signal for H3K27me3 in the gray matter in comparison with the white matter in both control and EMF groups, which could be explained by unspecific binding of the antibody.
In conclusion, although we have been living with increased artificial EMF exposure for over 20 years, we still do not know the full extent of its effects on living organisms yet. Therefore, here in this study, we performed the histopathological analysis of the spinal cord after prenatal EMF exposure for the first time in the literature and attempted to provide a molecular explanation to the detrimental effects of EMF exposure on the spine. We believe that our findings will be useful for future studies aimed at better understanding the effects of EMF on neural system development.
Footnotes
Acknowledgment
The authors would like to thank Dr Hatice HANCI for animal supply in the study.
Author contributions
Both authors contributed to the conceptualization and methodology of the study as well as to the writing of the manuscript.
Data availability statement
Data available on request from the authors.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All animal procedures were approved by the Institutional Ethics Committee at Niğde Ömer Halisdemir University (Date: September 10, 2019, Protocol number: 2019/27).
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.