
Abstract
In this presented study, the aim was to investigate the toxic effects of bisphenol S (BPS), one of the bisphenol A analogues, on the thyroid glands of male Wistar albino rats. Toward this aim, the rats (n = 28) were given a vehicle (control) or BPS at 3 different doses, comprising 20, 100, and 500 mg/kg of body weight (bw) via oral gavage for 28 days. According to the results, BPS led to numerous histopathological changes in the thyroid tissue. The average proliferation index values among the thyroid follicular cells (TFCs) displayed increases in all of the BPS groups, and significant differences were observed in the BPS-20 and BPS-100 groups. The average apoptotic index values in the TFCs were increased significantly in the BPS-500 group. The serum thyroid-stimulating hormone and serum free thyroxine levels did not show significant changes after exposure to BPS; however, the serum free triiodothyronine levels displayed significant decreases in all 3 of the BPS groups. BPS was determined to cause significant increases in the antioxidant enzyme activities of catalase, superoxide dismutase, glutathione-S-transferase, glutathione peroxidase, as well as a significantly decreased content of reduced glutathione. The malondialdehyde level in the thyroid tissue was elevated significantly in the BPS-500 group. The data obtained herein revealed that BPS has thyroid-disrupting potential based on structural changes, follicle cell responses, and biochemical alterations including a decreased serum free triiodothyronine level and increased oxidative stress.
Introduction
In the manufacturing industry, bisphenol A (BPA; 4,40-dihydroxy-2,2-diphenylpropane) is a well-known endocrine-disrupting chemical that is also suspected of being a carcinogen. It is an ingredient in epoxy resins and polycarbonate plastics and a wide variety of products of daily life including infants’ feeding bottles, toys, medical equipment, thermal paper receipts, food cans, beverage containers, dental sealants, and flame retardants (Huang et al., 2012). BPA exposure in humans has been linked with many disorders, including cardiovascular diseases, diabetes, reproductive disorders, birth defects, obesity, immune-related diseases, and liver-enzyme abnormalities (Caporossi and Papaleo, 2017; Rezg et al., 2014; vom Saal and Myers, 2008; Xu et al., 2016). Humans are mainly exposed to BPA via their diet, (such as through food and materials that come into contact with their food) (Andújar et al., 2019), in addition to dermal contact and dust inhalation (Manzoor et al., 2022). Owing to its noxious effects on public health, the use of BPA has been banned by several government organizations in daily items including infant’s bottles, sippy cups, and thermal receipts in the United States, Canada, and European Union (Beg and Sheikh, 2020; Liao and Kannan, 2014; Naderi et al., 2014; Rosenmai et al., 2014). Thus, regulations and increasing health concerns have motivated manufacturers to consider developing and producing BPA-free products. Consequently, manufacturers have phased out BPA usage and have begun to use chemically and functionally alternative substances including bisphenol F (BPF), bisphenol S (BPS, bis-(4-hydroxyphenyl)-sulfone), bisphenol B, and bisphenol AF instead of BPA in many applications (den Braver-Sewradj et al., 2020; Harnett et al., 2021; İyigündoğdu et al., 2020; Rochester and Bolden, 2015).
As a BPA derivative, BPS possesses higher thermal stability and resistance to sunlight than BPA and has been commonly used in industrial applications in recent years. It has been gradually replacing BPA in the production of polycarbonate resins, epoxy resins, personal care products, canned soft drinks, canned food, thermal papers, etc. Because of its increasing utilization, BPS was reported to be detected in different environments such as sewage sludge, surface water, tap water, sediment, and indoor dust, and also in daily consumer products such as personal care products and foodstuffs. Thus, humans are inevitably exposed to BPS and as a result of this, BPS was found in biological samples collected from humans including breast milk, blood, and urine at comparable concentrations with BPA (Deceuninck et al., 2015; Jin et al., 2018; Liao et al., 2012; Thayer et al., 2016; Zhang et al., 2016a).
Many in vitro as well as in vivo laboratory studies that have been conducted have shown that BPS possesses adverse effects such as cytotoxicity, genotoxicity, endocrine disruption, neurotoxicity, and immunotoxicity similar to BPA. The endocrine-disrupting potential of BPS includes estrogenic, androgenic, and anti-androgenic activities, as reported before for BPA. It also promotes oxidative stress (OS) generating of reactive oxygen species (ROS), inducing lipid peroxidation (LPO), and depressing activities of antioxidant enzymes (Chen et al., 2016; İyigündoğdu et al., 2020; Wu et al., 2018). Experimental studies in rodents conducted in the last decade showed that BPS caused structural and endocrine alterations similar to BPA. Exposure to BPS subchronically has been reported to affect liver morphology and function in mice by inducing oxidative damage (Pal et al., 2017; Zhang et al., 2018a) reported that rats exposed to BPS exhibited alterations in their blood biochemistry, and that it increased the serum total glucose, protein content, total cholesterol, triglyceride, low-density protein as well as very low-density protein levels, while it reduced the high-density lipoprotein level, and it induced liver function enzymes which are probably related to cardiac tissue damage, such as aspartate aminotransferase, alanine aminotransferase, and alkaline phosphatase activities. Thus, the authors proposed that cardiovascular risks might be promoted by BPS in rats as a consequence of impairment of blood function. Treatment of female pups with BPS resulted in a delay in the onset of puberty as well as alterations in the estrous cycle, increased final body weight, reduced gonadosomatic index and uterus weight, increased plasma testosterone and estradiol levels, decreases in the levels of plasma progesterone, luteinizing hormone, and follicle-stimulating hormone, and increased cystic and atretic follicles in the ovaries (Ahsan et al., 2018). BPS was shown to reduce plasma and intratesticular testosterone levels, and cause histopathological alterations in the testes and epididymis, and escalations in the testicular OS status (Ullah et al., 2016).
Thyroid hormones have significant involvement in regulating many important physiological processes including growth, development, metabolism, energy expenditure, reproduction, and thermoregulation in vertebrates. The synthesis, secretion, and the metabolism of thyroid hormones are controlled by the hypothalamus-pituitary-thyroid (HPT) axis. The hypothalamus is responsible for releasing thyrotropin-releasing hormone, which stimulates the anterior pituitary to produce thyroid-stimulating hormone (TSH). The production of 3,3′,5-triiodothyronine (T3) and thyroxine (T4), which are thyroid hormones, occurs in the thyroid gland under the stimulation of TSH. Synthesized thyroid hormone binds to blood proteins, such as albumin, transthyretin (TTR), or thyroxine-binding globulin (TBG), and circulates in the blood. Thyroid hormones bind to the thyroid hormone nuclear receptors (TRα or TRβ) at their corresponding target tissues and stimulate thyroid hormone signaling pathways. It was reported in recent years that myriads of endocrine-disrupting chemicals have the potential to interfere with the synthesis and control mechanism of thyroid hormones (Calsolaro et al., 2017; Leemans et al., 2019; Oliveira et al., 2018; Thambirajah et al., 2022; Zoeller, 2010). Many previous reports have indicated that BPA is capable of disrupting thyroid function by binding thyroid receptors or interfering with thyroid hormones. There is now reasonably firm evidence that the synthesis, transport, and metabolism of thyroid hormones can be influenced adversely by BPA as well as its derivatives (Kim and Park, 2019; Milczarek-Banach and Miśkiewicz, 2020; Yuan et al., 2020). In experimental studies in which zebrafish (Danio rerio) were used as a model organism, BPS was reported to have adverse effects on the HPT axis. Lu et al. (2018) reported that environmentally relevant concentrations of BPS have been shown to have antagonistic activity toward TRβ and the expression of mRNA on the TRβ gene was significantly altered by BPS in zebrafish embryos. In another study, BPS was shown to decrease the T4 and T3 levels, while it increased the TSH level in zebrafish larvae. Additionally, increments in the transcription levels of genes that are associated with thyroid development and synthesis and the upregulation of genes involved in thyroid hormone metabolism have been reported. Thus, BPS accelerates thyroid disruption by changing thyroid hormone and TSH levels, and the expression profiles of key genes related to the HPT axis in zebrafish larvae (Zhang et al., 2017). In a study involving a series of in vivo and in vitro assays, it was reported that BPS, just like BPA, interferes with the thyroid hormone signaling pathway (Zhang et al., 2018b).
Thyroid-follicular cell proliferation, which increases the total thyroid-follicular cell number, is an indicator of growth of the thyroid gland (Hood et al., 1999a; Wynford-Thomas et al., 1982). Apoptosis is a form of programmed cell death that plays a critical role in tissue homeostasis during normal development. It is also responsible for the elimination of unwanted cells in tissues, and it can be triggered by toxic stimuli (Chiappini et al., 2009). The critical balance between thyroid-follicular cell proliferation and apoptosis has been reported to play an important role in the modulation of the thyroid gland mass (Tamura et al., 1998).
BPS exerts endocrine-disrupting impacts on organisms and possesses targets in the body similar to the BPA. It is a hormonally active compound and also shows a similar manner of action to BPA in terms of affecting physiological processes. Moreover, important membrane-mediated pathways in cells, including proliferation, differentiation, and death, can be influenced by BPS, similar to what occurs with estradiol (Rochester and Bolden, 2015). Therefore, questions regarding the usage of BPS as a safe alternative instead of BPA are raised on a daily basis due to its health-related and environmentally toxic effects. Thus, there is a need for more research to learn about the negative impacts of BPS on the structure, function, and health of the body’s endocrine organs, including the thyroid gland.
Although there are studies on the thyroid-disrupting effects of BPA in rats (Alkalby, 2015; Ahmed 2016; Ibrahim and Morsy, 2019; da Silva et al., 2018; da Silva et al., 2019), as far as is known, in vivo research regarding the toxic and disrupting impacts of BPS on the thyroid have not been reported in a rat model to date. To fill this gap in the literature, the study presented herein was designed to investigate the adverse effects that BPS has on the thyroid gland. Toward this aim, male rats were exposed to BPS daily for 28 days and the histopathological changes that took place were examined. To our knowledge, there are no reports on the subject of how BPS effects thyroid follicular cell (TFC) responses. Therefore, the follicular cell proliferation was investigated using proliferating cell nuclear antigen (PCNA) immunohistochemistry, as was follicular cell apoptosis using the terminal deoxynucleotidyl transferase nick end labeling (TUNEL) technique. Moreover, to assess the biochemical effects of BPS in relation to its toxicity, the levels of serum hormones and antioxidant system indicators in the gland were evaluated.
Materials and methods
Animals
Purchased from The Experimental Medicine Research and Application Centre of Our University were 28 male Wistar albino rats, which were between 2 and 3 months old and weighed 180 to 245 g. The male rats were preferred in this study to avoid the physiological variability related to the estrous cycle of females. The experiments were performed at the same center. The animals were kept under controlled experimental conditions that consisted of a light/dark photoperiod of 12:12, an average temperature of 25°C, and humidity level of 50% to 70%. All of the animals were housed in standard cages and standard pellet feed and water were available for the rats to consume ad libitum. The rats were acclimated to the experimental laboratory conditions prior the study to minimize their stress responses.
BPS treatment and experimental design
The test chemical BPS (bis-(4-hydroxyphenyl)-sulfone; molecular formula: C12H10O4S; purity: 99.7%, molecular weight: 250.27; code: 146915000; Acros Organics, New Jersey, USA) was dissolved in ethanol-olive oil (1:4 v/v) mixture (da Silva et al., 2018; Pal et al., 2017). The doses of BPS used in this study were determined according to previous studies based on bisphenol exposure in rats (da Silva et al., 2018; Higashihara et al., 2007; Pal et al., 2017; Sencar et al., 2021; Uzunhisarcikli and Aslanturk 2019; Yamasaki et al., 2002). The animals (n = 28) were divided randomly into 4 experimental groups, each containing 7 rats. The rats were treated with BPS daily by oral gavage for 28 consecutive days. Standard pellet feed and water were available for the rats to consume ad libitum throughout the experiment. The study groups were established as follows:
Vehicle control (VC) Group: (n = 7). These animals received the vehicle (ethanol-olive oil mixture, 1:4 v/v) daily via oral gavage for consecutive 28 days.
BPS-20 Group: 20 mg of BPS/kg body weight (bw)/day (n = 7). The animals received BPS dissolved in the vehicle at a daily dose 20 mg/kg via oral gavage according to their bw during the study. Standard pellet feed and water were available for the rats to consume ad libitum throughout the experiment.
BPS-100 Group: 100 mg of BPS/kg bw/day (n = 7). The animals received BPS dissolved in the vehicle at a daily dose 100 mg/kg via oral gavage according to their bw during the study.
BPS-500 Group: 500 mg of BPS/kg bw/day (n = 7). The animals received BPS dissolved in the vehicle at a daily dose 500 mg/kg via oral gavage according to their bw during the study.
Sample collection
On the day following the final BPS treatment, the rats were anesthetized with ketamine/xylazine. Blood was withdrawn from the heart of each animal using disposable sterile syringes and transferred into biochemistry tubes. Following coagulation, centrifugation of the tubes was performed at 400 g for 10 min at a temperature of 4°C for collection of the serum and then the sera were removed and used immediately to assay the hormone levels. After collection of the blood samples, the thyroid glands were quickly excised via the use of fine scissors and forceps. Next, 10% neutral buffered formalin was used for fixation of the right lobes of the glands for 24 h and they were then used for the histological, histochemical, and immunohistochemical examinations, while the other lobes were stored at −80°C for the biochemical assays.
Histopathological evaluations
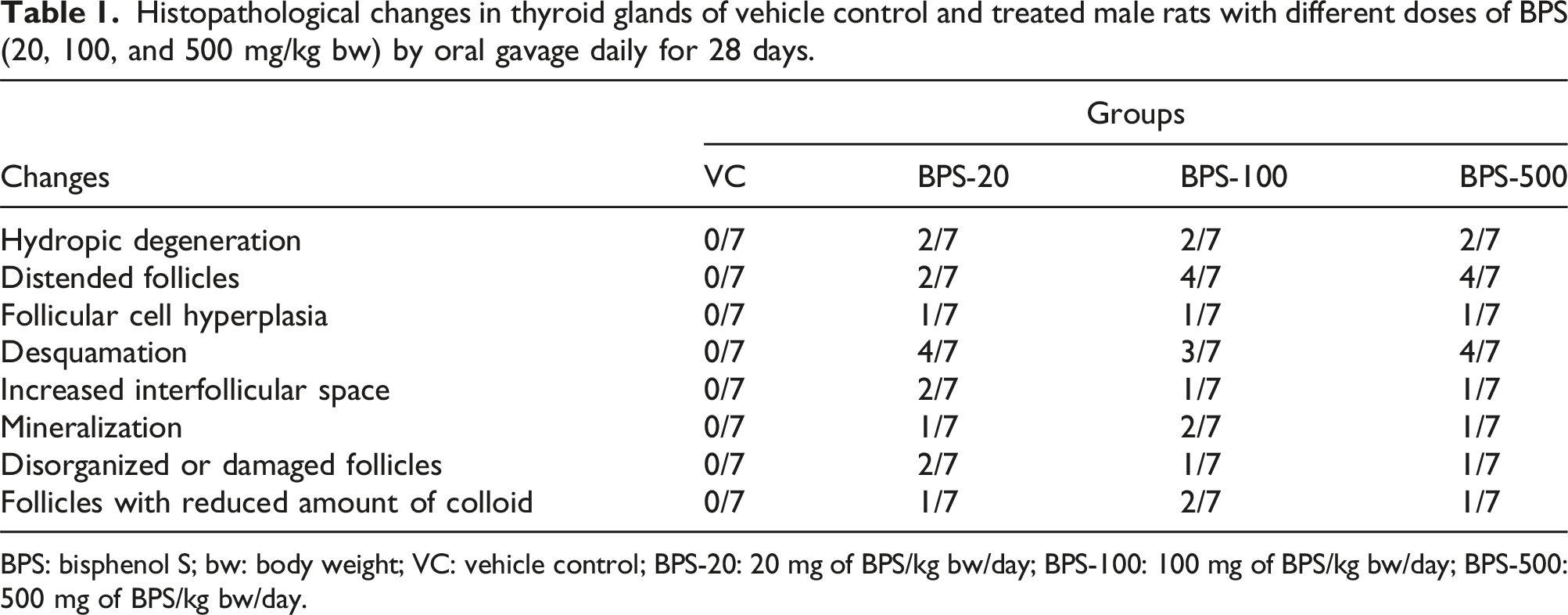
Following fixation in formalin, phosphate buffered saline (PBS) (0.01 M at pH 7.4) was used to wash the tissues. Afterward they were passed through a graded series of ethanol, comprising dilutions of 70%, 80%, 96%, and absolute, cleared using xylene, and then they were embedded in paraffin as a final step. Next, sections that measured 5 µm thick were removed from the tissue blocks by means of a manual microtome (MICROM HM 325, Waldorf, Germany). The sections were then placed onto slides that were coated with adhesive deparaffinized, rehydrated, and ultimately stained with hematoxylin and eosin (H&E), and mounted with Entellan. The preparations were then examined under a microscope (Leica DMI 6000B microscope, Wetzlar, Germany). Histopathological evaluations of the thyroid glands were performed according to the methods of Yoshizawa et al. (2010), Hadie et al. (2012), Abd El-Twab and Abdul-Hamid (2016), and Brändli-Baiocco et al. (2018). A histopathological examination of the thyroid tissues was performed to assess lesions, including hydropic degeneration, distended follicles, follicular cell hypertrophy, follicular cell hyperplasia, desquamation, increased interfollicular space, mineralization, disorganized or damaged follicles, and follicles with a reduced amount of colloid. Next, images were taken at appropriate magnifications using a Leica Digital DFC490 model camera (Leica Microsystems CMS GmbH, Wetzlar, Germany) that was attached to the microscope.
Immunohistochemical staining of the proliferating cell nuclear antigen
The proliferating activity of the thyroid-follicular cells was then evaluated via the use of immunohistochemical staining of the proliferating cell nuclear antigen (PCNA) in the thyroid sections. Toward this aim, a commercial PCNA primary antibody (PC10, cat. no.: 13–3900, Thermo-Fisher Scientific, Invitrogen) and commercial immunohistochemistry (IHC) kit (Mouse and Rabbit Specific HRP/DAB IHC Detection Kit-Micropolymer, ab236466, Abcam Cambridge, UK), which works principally via the use of a system that detects biotin-free immunoenzymatic antigens, were used. Briefly, the sections, which measured 5 µm, were first deparaffinized, and then rehydrated and also washed using tris-buffered saline (TBS) (20 mM at a pH of 7.6; 140 mM NaCl). Following this, inactivation of the tissue endogenous peroxidase activity using the H2O2 supplied in the kit was conducted. Afterward, antigen recovery was performed via the use of antigen retrieval solution (Dako, Glostrup, Denmark). After performing protein blocking to prevent nonspecific bindings, TBS was used to wash the sections, which was done several times, and then they were incubated in a diluted solution of PC10 primary antibody (1:400) for 2 h at room temperature. After incubation was completed, the slides were washed with TBS, and they were then treated using a complement, which was then followed by a horseradish peroxidase conjugate that had been supplied in the IHC kit. After a final washing step with TBS, a solution of substrate-chromogen, 3,3′-diaminobenzidine, was dropped onto the sections in order to be able to visualize the peroxidase activity indicating antibody binding. Mayer’s hematoxylin was preferred for counterstaining and the sections were then mounted using Entellan. For testing of the specificity of the primary antibody, a negative control slide was used, in which TBS took the place of the primary antibody.
Detection of apoptotic thyroid-follicular cells (TFCs)
Apoptotic TFCs possessing fragmented nuclei were labeled using the histochemical TUNEL method on the tissue sections. TUNEL staining was conducted via the use of a commercially available terminal deoxynucleotidyl transferase (TdT)-FragEL DNA Fragmentation Detection Kit (cat. no: QIA33, Calbiochem, Merck, USA) by following the instructions listed in the kit by the manufacturer. Briefly, after deparaffinization and rehydration of the sections, tissue permeabilization was performed using proteinase K for 20 min. Next, the endogenous peroxidases were inactivated by exposing the sections to 3% H2O2. As a next step, the sections were equilibrated via the use of a buffer that had been provided in the kit. They were then incubated in a TdT labeling mixture at 37°C in a humidified chamber for 90 min. Next, a stop solution was dropped onto the sections to terminate this reaction. After washing with TBS several times, the sections were treated with a blocking buffer, and then they were incubated again, but this time with a conjugate of peroxidase streptavidin at room temperature for 30 min. After a final washing step with TBS, a solution of substrate-chromogen, 3,3′-diaminobenzidine, was dropped onto the sections in order to develop a brown color in the nuclei within the apoptotic cells. Methyl green (0.3%) was preferred for counterstaining and Entellan was then used for mounting of the sections. The sections that were on the negative control slides included the substitution of bidistilled water in place of the reaction mixture, whereas the positive control slides included incubation with DNase I (AppliChem, cat no: A3778).
Counting of the proliferating and apoptotic thyroid follicular cells
At least 10 randomly chosen fields in the thyroid sections with both PCNA immunohistochemistry and TUNEL stainings pertaining to each animal were photographed at ×400 magnification. The images (.jpg) were transferred to ImageJ software (National Institutes of Health, USA, at https://rsbweb.nih.gov/ij/) for counting of the cells. Next, the positively (brown-colored) and negatively stained nuclei of the TFCs were counted via the use of the cell-counter plugin within the software (https://imagej.nih.gov/ij/plugins/cell-counter.html) in different fields for both of the stainings. Following the completion of the counting, the PCNA index (PI) and apoptotic index (AI) were calculated according to the formula: number of PCNA/TUNEL-positive nuclei × 100/total (positive + negative) number of nuclei. At least 600 nuclei of the TFCs were evaluated for each animal. The PI and AI values obtained from the different fields were averaged for each animal and they were then used in the statistical evaluations (Iglesias-Osma et al., 2019; Matsuu-Matsuyama et al., 2015). The results obtained were expressed as percentages (%) for the PI and AI.
Assay of serum hormone levels
The levels of free (unbound) triiodothyronine (FT3), free thyroxine (FT4) and thyroid-stimulating hormone (TSH) in the serum were assayed by chemiluminescent microparticle immunoassay via the use of an Architect i4000 SR autoanalyzer (Abbott Diagnostics Inc., Illinois, USA). The values were expressed as pg/mL for the FT3, ng/mL for the FT4, and mLU/mL for the TSH.
Tissue homogenization
Thyroid tissues stored in a deepfreeze (−80 C°) were thawed on ice. First, the tissues were homogenized in PBS (ice-cold at pH 7.4) using a Bandelin Sonopuls HD 2070 ultrasonic homogenizer (Bandelin Electronic GmbH & Co KG). Centrifugation of the obtained homogenates was then performed at 15,000 × g at 4°C for 20 min. Next, the supernatants were carefully removed, pipetted into different tubes, and stored at −20°C until measurement of the total protein content was conducted and parameters of the antioxidant defense system were determined.
Total protein assay
The total protein content in the supernatants was assayed spectrophotometrically at 595 nm via the use of the method reported by Bradford (1976) with bovine serum albumin being used as the standard.
Assay of antioxidant defense system parameters
A commercial kit (Ransod, cat. no: SD125, Randox Laboratories Ltd, County Antrim, UK) was used to the assay superoxide dismutase (SOD) activity in the supernatants. The kit procedures, based on the method of Suttle and McMurray (1983), were followed throughout the assays using a Shimadzu UV/VIS 1201 spectrophotometer (Shimadzu Corporation, Kyoto, Japa) at 505 nm. The results that were obtained were expressed as the number of units for each milligram of protein (U/mg of protein).
The activity of catalase (CAT) was assayed spectrophotometrically at 240 nm according to the method that was reported by Aebi (1974) which is based on measurement of the decrease in the absorbance at 240 nm as a result of the consumption of H2O2. The results that were obtained were expressed as the number of micromoles of H2O2 that are consumed each minute, for each gram of protein (µmole of H2O2 consumed/min/g of protein).
A commercial kit (Ransel, cat. no: RS504, Randox Laboratories Ltd, County Antrim, UK) was used to determine the glutathione peroxidase (GPx) activity of the supernatants. The kit procedures, which are based on the method that was reported by Paglia and Valentine (1967), were followed throughout the assays using a UV-VIS spectrophotometer at 340 nm. The results that were obtained were expressed as the U/g of protein.
Determination of the glutathione S-transferase (GST) activity was performed using the method that was reported by Habig et al. (1974). The assay was spectrophotometrically performed at 340 nm. The activity was expressed as the number of nanomoles of CDNB-glutathione conjugate each min for each mg of protein (nmol of CDNB-glutathione conjugate/min/mg of protein).
The method reported by Beutler (1984) was used to determine the reduced glutathione (GSH) content. The assay was spectrophotometrically performed at 412 nm. To calculate the concentrations in the samples, a standard curve that was derived via the use of external standards was used. The results that were obtained were expressed as the number of µmoles of GSH for each mg of protein (µmoles of GSH/mg of protein).
Malondialdehyde (MDA), which is the end product resulting from lipid peroxidation, was spectrophotometrically measured at 532 nm using the method that was described by Buege and Aust (1978). A standard curve that was derived via the use of external 1,1,3,3-tetraethoxypropane standards was used. Next, the concentration of MDA in the supernatants was calculated. The results that were obtained were expressed as the number of nmole/mg of protein.
Statistical analysis
The statistical analyses were all conducted with IBM SPSS Statistics for Windows 20.0 (IBM Corp., Armonk, NY, USA). Evaluation of the differences between the groups was conducted with 1-way analysis of variance (ANOVA) that was followed by a Duncan multiple-comparison post-hoc test. All of the values obtained were expressed as the mean ± standard deviation (SD). p values that were less than .05 were considered to have statistical significance.
Results
Histopathological lesions
The thyroid gland sections stained with H&E pertaining to the VC group displayed normal histological architecture. The control glands included numerous and different sized follicles, and the lumina of the follicles were filled with acidophilic homogenous colloid (Figure 1(A)). However, exposure to BPS was determined to have led to marked histopathological changes in the architecture of the gland. The histopathological changes described after BPS exposure included disorganized and damaged follicles (Figure 1(B) and (G)), increased interstitial space that was characterized by widening of the spaces among the follicles (Figure 1(B)–(D)), hydropic degeneration in the follicular cells that was characterized by a swollen cytoplasm with a glassy appearance (Figure 1(B), (D) and (G)), the follicles containing decreased amounts of colloid or no colloid (Figure 1(B) and (H)), desquamated follicular cells in the follicular lumen (Figure 1(B), (D) and (H)), mineralization in the colloid (Figure 1(C)), follicular cell hypertrophy that was characterized by a decreased diameter of the follicular lumen with an increased height of the follicular epithelium (Figure 1(D)), follicular cell hyperplasia that was characterized by infoldings or stratification of follicular epithelium that occurred with the former alteration (Figure 1(E)), distended follicles that were characterized by enlarged follicular lumen surrounded by simple squamous epithelia with a flat nuclei (Figure 1(F)). The histopathological changes observed in the thyroid glands in the VC and BPS-20, BPS-100, and BPS-500 groups, and the number of animals in which different types of histopathologic changes were observed in each group are summarized in Table 1. Representative photomicrographs of sections taken from the thyroids of the VC group (A) and BPS-20, BPS-100, and BPS-500 groups showing various types of histopathological changes (B−H) (H&E). (A) VC group: the thyroid gland possessing normal histological architecture displays numerous follicles in different sizes that are surrounded by simple cuboidal epithelia and the lumina of the follicles are filled with acidophilic homogenous colloid. (B, C) BPS-20 group: glands displaying loss of normal architecture with histopathological changes such as disorganized and damaged follicles (black arrows), hydropic degeneration in the follicular cells (black arrowheads), follicles with no colloid (yellow asterisk), desquamation into the follicular lumen (green arrow) (in B), and mineralization of the colloid (yellow arrowheads) (in C). (D−F) BPS-100 group: histopathological lesions including desquamated cells (green arrows), increased interfollicular space (black asterisk), hydropic degeneration in the follicular cells (black arrowhead), follicular cell hypertrophy (yellow arrows) (in D), follicular cell hyperplasia characterized by infolding or stratification of follicular epithelium (blue arrows) (in D and E), and distended follicles (in F). (G, H) BPS-500 group: damaged follicle (black arrow), hydropic degeneration in the follicular cell (black arrowhead) (in G), follicles with a reduced amount of colloid (yellow asterisk), and desquamation into the follicular lumen (green arrow) (in H). BPS: bisphenol S; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; H&E stain, H&E: hematoxylin and eosin. Histopathological changes in thyroid glands of vehicle control and treated male rats with different doses of BPS (20, 100, and 500 mg/kg bw) by oral gavage daily for 28 days. BPS: bisphenol S; bw: body weight; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day.
Effect of BPS on the PI in the TFCs
Widespread PCNA-positive TFCs with brown-colored nuclei were observed in the glands of the BPS-20, BPS-100, and BPS-500 groups in comparison with the VC group (Figure 2). In accordance with this observation, the values of the PI for the TFCs in the BPS-20, BPS-100, and BPS-500 groups were observed to be higher in comparison with those in the VC group with statistical significances (p < .05) at 20 and 100 mg/kg of BPS (Table 2). Representative photomicrographs of thyroid sections immune-stained with PCNA primary antibody in which PCNA-positive proliferating TFCs appeared as brown in their nuclei. In comparison with the VC group (A), a greater amount of positively stained TFCs can be seen in the BPS-20 (B), BPS-100 (C), and BPS-500 (D) group samples. PCNA: proliferating cell nuclear antigen; TFCs: thyroid follicular cells; BPS: bisphenol S; VC: vehicle control; bw: body weight; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; PCNA immunstaining. PI: proliferating cell nuclear antigen index; TFCs: thyroid follicular cells; BPS: bisphenol S; bw: body weight; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; SD: standard deviation. aValues are presented as the mean ± SD. bDifferent superscripts indicate statistically significant differences according to the Duncan multiple-comparison post-hoc test that were detected among the groups (p <.05).

Effect of BPS on the AI in the TFCs
Apoptotic (TUNEL-positive) follicular cells stained with brown-colored nuclei were observed scarcely in TFCs in the glands in the VC group. However, the presence of apoptotic cells was observed to increase remarkably in the BPS-20, BPS-100, and BPS-500 groups in comparison with the VC group (Figure 3). The values of the AI for the TFCs was appeared increased in the BPS-20, BPS-100, and BPS-500 groups in comparison with the VC group with statistical significance at 500 mg/kg of BPS (p < .05) (Table 3). Representative photomicrographs of thyroid sections stained with TUNEL in which apoptotic TFCs appeared as brown in their nuclei. In comparison with the VC group (A), a greater amount of TUNEL-positive stained (apoptotic) TFCs can be seen in the BPS-20 (B), BPS-100 (C), and BPS-500 (D) group samples. TUNEL: terminal deoxynucleotidyl transferase nick end labeling; TFCs: thyroid follicular cells; BPS: bisphenol S; VC: vehicle control; bw: body weight; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; TUNEL staining. AI: apoptotic index; TFCs: thyroid follicular cells; BPS: bisphenol S; bw: body weight; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; SD: standard deviation. aValues are presented as the mean ± SD. bDifferent superscripts indicate statistically significant differences according to the Duncan multiple-comparison post-hoc test that were detected among the groups (p <.05).

Serum hormone levels

FT3: free triiodothyronine; FT4: free thyroxine; TSH: thyroid-stimulating hormone; BPS: bisphenol S; bw: body weight; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; SD: standard deviation.
aValues are presented as the mean ± SD.
bDifferent superscripts indicate statistically significant differences according to the Duncan multiple-comparison post-hoc test that were detected among the groups (p <.05).
Effect of BPS on the antioxidant defense system parameters in the thyroid gland
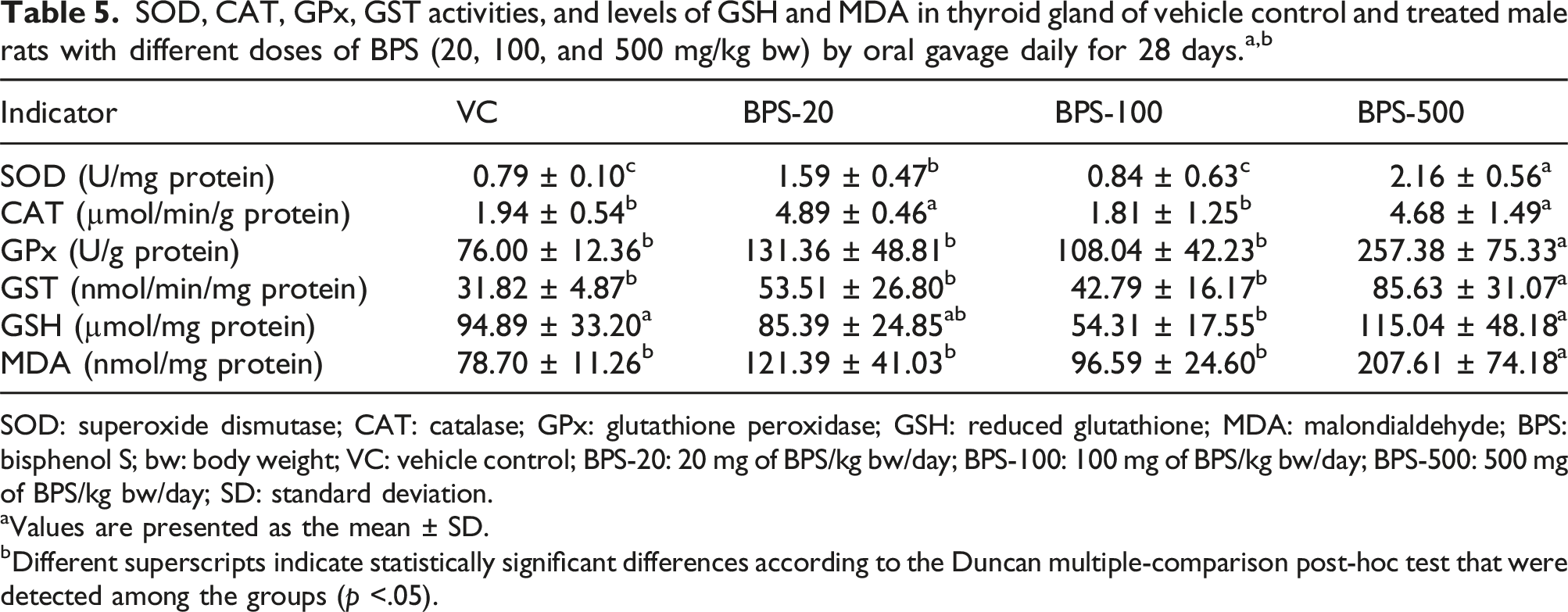
SOD: superoxide dismutase; CAT: catalase; GPx: glutathione peroxidase; GSH: reduced glutathione; MDA: malondialdehyde; BPS: bisphenol S; bw: body weight; VC: vehicle control; BPS-20: 20 mg of BPS/kg bw/day; BPS-100: 100 mg of BPS/kg bw/day; BPS-500: 500 mg of BPS/kg bw/day; SD: standard deviation.
aValues are presented as the mean ± SD.
bDifferent superscripts indicate statistically significant differences according to the Duncan multiple-comparison post-hoc test that were detected among the groups (p <.05).
Discussion
Due to its endocrine-disrupting potential, adverse toxic effects and implications with many serious health issues, including such things as cardiovascular disease, neurologic disorders, developmental anomalies, type-2 diabetes mellitus, as well as obesity (Andújar et al., 2019; Atay et al., 2019, 2020; Costa and Cairrao, 2024), BPA, which is used widely in various industrial applications, has been regulated in many countries. Those regulations have led environmental agencies and manufacturers to seek alternatives that are less harmful when compared to BPA (Erkekoğlu et al., 2022; Moon, 2019). In recent years, BPS has been suggested and introduced into industrial applications as an alternative substitute for BPA. However, it has been revealed in vitro studies that BPS can result in cytotoxicity, genotoxicity, mutagenicity, neurotoxicity, hepatotoxicity, anti-androgenic, and estrogenic activities because of its structural similarity to BPA (Eladak et al., 2015; Feng et al., 2016; Fic et al., 2013; Kitamura et al., 2005; Lee et al., 2013; Mathew et al., 2014; Molina-Molina et al., 2013; Peyre et al., 2014; Vinas and Watson, 2013). In recent experimental studies in which rats were used, BPS was reported to exhibit toxic behaviors and functional alterations on the male reproductive system (Ullah et al., 2016, 2018). As best as is known, there have been no studies investigating the negative impacts of BPS on the thyroid gland in rats. Thus, in the research that was conducted in the current paper, the aim was to evaluate and determine the effects that BPS has on the thyroid structure, function, and biochemistry. For this purpose, male rats were treated by oral gavage with BPS for 28 days and then histopathological changes, responses of TFCs using proliferation and apoptosis as indicators, serum hormone levels, and antioxidant defenses in the gland were investigated.
The doses of BPS selected in the current study were determined according to the following criteria. First; the European Commission requested European Food Safety Authority (EFSA) to assess the impact on the current authorization of BPS in plastic food contact materials of recent studies submitted by BPS registrant(s) in response to the European Chemicals Agency’s Decision on the evaluation of substances under Regulation (EC) No. 1907/2006 (European Union, 2006). Those studies included an Extended One-Generation Reproductive Toxicity Study (EOGRTS), with developmental neurotoxicity cohorts (Developmental Neurotoxicity Study) and Developmental Immunotoxicity Study (The Organisation for Economic Co-operation and Development (OECD) Test Guidelines, TG 443), and a Toxicokinetic Study (OECD TG 417) in rats. The lowest NOAEL from the EOGRTS was identified for developmental toxicity and developmental immunotoxicity at the lowest BPS dose tested of 20 mg/kg bw per day. The mid dose of 60 mg BPS/kg bw/day was the NOAEL for overall systemic toxicity, while developmental neurotoxicity, fertility and reproductive performance were not affected even with the high tested dose of 180 mg BPS/kg bw/day (EFSA, 2020). According to the low-dose hypothesis, chemical substances referred to as endocrine active substances or endocrine disruptors may cause adverse effects at low doses but not necessarily at all higher doses (EFSA, 2012). On the other hand, low-level exposures to known toxic chemicals could be “beneficial” to human health, an effect that is known as hormesis (Calabrese, 2018; Shanker, 2008). Recent studies reported that BPA or its derivatives might exhibit hormesis or hormesis-like effects in some cases (Fan et al., 2020; Liu et al., 2022). Second; previous studies in which rats were exposed to BPA or its analogues such as BPS and BPF were taken into account. For example, in studies associated with BPA-induced tissue inflammation in rats, doses of the BPA given to the animals by oral gavage can be up to 500 mg/kg/day (see for a recent review by Ricker et al. (2024)). In a study by Yamasaki et al. (2002), BPA at doses of 0, 40, 200, and 600 mg/kg per day were orally administered to SD rats. Sencar et al. (2021) preferred 0, 50, 100 and 200 mg/kg/day doses of BPA and Uzunhisarcikli and Aslanturk (2019) have been used a single dose of BPA (130 mg/kg/bw per day) in their toxicity studies in which rats were used. In an another study, bisphenol F was orally administered at doses 0, 20, 100, and 500 mg/kg per day to SD rats for at least 28 days while no clear endocrine related effect has been seen (Higashihara et al., 2007). On the other hand, adult male albino rats of Sprague-Dawley strain were given BPS at a dose level of 30, 60 and 120 mg/kg bw/day, respectively, for 30 days for determination of effects of BPS on hematological functions and cardiovascular risks (Pal et al. (2017). Due to the above-mentioned reports, we selected 20, 100, and 500 mg/kg/day doses of BPS in this study.
As a consequence of BPS exposure, histopathological alterations in the thyroid gland were seen to occur, including damaged follicles, desquamated follicular cell in the follicular lumen, increased interfollicular space, hydropic degeneration, reduced or absence of the amount of colloid in some follicles, mineralization, hypertrophy, hyperplasia, and distended follicles. Even though there have been no experimental studies reporting the histopathologic effects of BPS on the thyroid gland, similar observations were also seen to occur in the thyroid glands of rats that had been exposed to BPA in different experimental studies (Alkalby, 2015; Ahmed, 2016; da Silva et al., 2018; Ibrahim and Morsy, 2019). Alkalby (2015) reported that BPA caused histopathological lesions, such as different sized thyroid follicles with vacuolated colloid, follicles with flattened thyrocytes, and increased microfollicles in the thyroid gland of adult male rats (Rattus novergicus). Ahmed (2016) also reported histopathological changes that included fibroblast proliferation, edema, luminal obliteration, hyperplasia, and degeneration in the thyroid glands of the fetuses of pregnant rats which were exposed to BPA starting at day 1 of gestation to day 20. In other research, an increased number of hypoactive follicles with squamous epithelia that were lined with interfollicular connective tissue were seen in the thyroid glands of female Wistar rats exposed to BPA (da Silva et al., 2018). Exposure of male rats to BPA was reported to have resulted in vacuolated follicular cells, colloidal vacuolation, congested dilated capillaries in the interfollicular space, increased microfollicles, and coalescence of some thyroid follicles and exfoliated epithelial cells in the lumen in the thyroid gland (Ibrahim and Morsy, 2019). In accordance with the findings that were found in this current research, endocrine-disrupting chemicals in different classes including heavy metals (Salah et al., 2022; Wade et al., 2002), a plasticizer di (2-ethylhexyl) phthalate (Ye et al., 2017), a carbamate pesticide carbofuran (Hadie et al., 2012), 2,3,7,8-tetrachlorodibenzo-p-dioxin and dioxin-like compounds (Yoshizawa et al., 2010), an agricultural pesticide 1,1,1-trichloro-2,2-bis(4-chlorophenyl)ethane (p,p′-DDT) (Tebourbi et al., 2010), polychloriated biphenyls (Kılıç et al., 2005), and an organotin compound, tributyltin (Badr El Dine et al., 2017), have been reported to lead to similar structural changes observed in rat thyroid glands. It has been reported in many studies that the histopathological lesions that developed in rat thyroid gland after exposure to toxic chemicals, including BPA, were associated with remarkable decreases in the serum T4 and T3 levels, and significant elevations in the serum TSH levels (Abd El-Twab and Abdul-Hamid, 2016; Ahmed, 2016; Ibrahim and Morsy, 2019; Jeong et al., 2006; Salah et al., 2022; Tebourbi et al., 2010). Elevated levels of TSH could lead to follicular hypertrophy as well as hyperplasia in the gland (Badr El Dine et al., 2017; Salah et al., 2022; Yoshizawa et al., 2010). On the other hand, oxidative stress has been reported to be an important mechanism in thyroid toxicity and the histopathological changes in the thyroid were demonstrated to be related to an elevated oxidative stress status and the production of ROS (Abd El-Twab and Abdul-Hamid, 2016; da Silva et al., 2018; Salah et al., 2022). In the current study, the serum TSH and T4 levels remained unchanged, while serum the T3 levels decreased significantly. However, increased levels of MDA, which is a marker of oxidative stress, were measured in the gland after BPS treatment. Thus, it seems that the histopathological changes might have resulted from oxidative damage rather than thyroid-related hormonal effects.
Herein, the results pertaining to the serum hormone levels, at all of the BPS concentrations used, revealed that BPS could lead to significant decreases in the serum FT3 levels, while the serum FT4 and TSH levels remained unchanged. Although no experimental studies were found in the literature regarding the effect of BPS on hormone levels related with thyroid function in rats, it was reported in past studies that BPA, displaying a similar chemical structure to BPS, is capable of altering levels of those hormones in line with this study. The treatment of rats with BPA led to significant reductions in the serum T4 level; however, no significant alterations were reported in the serum TSH and T3 concentrations after BPA exposure (Alkalby, 2015). In another study, the serum T3 and T4 levels demonstrated significant decreases while the serum TSH level displayed a significant increase as a result of the BPA impact in adult male rats (Ibrahim and Morsy, 2019). da Silva et al. (2018) reported that BPA caused an increased serum T4 level, unchanged level of serum T3, and decreased level of β-TSH mRNA expression in the pituitary gland in female rats. Remarkable reductions in the levels of serum T3 and T4 of fetuses of pregnant rats exposed to BPA and an increase in the serum TSH level implicated with those reductions were reported (Ahmed, 2016). A transgenerational study performed using zebrafish has shown that BPS could lead to thyroid hormone disruption via maternal transfer that decreased the levels of plasma T4 in F0 females, while increased levels of plasma T3 in F0 males and females were seen. Additionally, the thyroid hormone content of F1 eggs that had been spawned by F0 females which had undergone BPS exposure were seen to exhibit similar trends, such as a significant decrease in the T4 level and an increase in the T3 level (Wei et al., 2018). Another study showed that BPS caused a decrease in the T4 and T3 levels, while an increase seen in the TSH level in the larvae of zebrafish and an altered expression in the profiling of key genes that are related to the HPT axis and thyroid hormone metabolism [deiodinases: dio1 and dio2, and uridinediphosphate glucoronosyltransferases (ugt1ab)], eventually induced thyroid disruption (Zhang et al., 2017). It can be concluded from the findings of the current study, which are parallel to the above-mentioned studies, that BPS, similar to BPA, has the ability to change the levels of thyroid hormones and causes a disruption in thyroid function. On the other hand, the serum TSH levels remained unchanged in this study. Similarly, although significant decreases were seen to occur in the serum T4 and T3 levels, the serum TSH level did not show any significant changes in rats exposed to di (2-ethylhexyl) phthalate, and it was expressed that an impairment in the negative feedback system of the HPT axis might lead to such an affect (Ye et al., 2017). Chiappini et al. (2009) reported also an unchanged level of TSH in rats exposed to hexachlorobenzene, and even a significant decrease was seen to occur in the levels of serum T4. Moreover, in spite of unchanged levels of serum FT4 levels, significant decreases were seen to occur in the levels of serum FT3 in this study. In an in vitro study that was conducted by Moriyama et al. (2002), it was reported that function of the thyroid hormone can be impaired by BPA via inhibiting of T3 from binding to the thyroid hormone receptor and suppressing of the transcriptional activity of T3. Thus, BPA can exert an antagonistic effect on T3 at the transcriptional level. In addition, different types of nuclear thyroid hormone receptors and their cofactors can be functionally disrupted by BPA. In a study established using Pelophylax nigromaculatus tadpoles, BPS was reported to disrupt functioning of the thyroid hormone signaling pathway, similar to BPA, and it can behave similar to thyroid hormone signaling agonists when T3 is absent, whereas when T3 is present, it could agonize or even antagonize the function of thyroid hormones under certain conditions (Zhang et al., 2018b). BPA has been reported to have inhibited type 1 and type 2 deiodinases in vitro. Moreover, prominent reductions in hepatic type 1 deiodinase activity in BPA-exposed rats along with increased serum T4 levels, but unchanged T3 levels, were found, and finally, the T3/T4 ratio was diminished. Thus, BPA has the capability to influence the metabolism of thyroid hormones (da Silva et al., 2019; Guarnotta et al., 2022). Hence, remarkable decreases in the T3 levels following BPS exposure in this study might have arisen from the inhibiting effect of BPS on deiodinases, similar to BPA, but more detailed research should be performed to determine the exact reason for such a result. On the other hand, it is understood from the above-mentioned studies, and including the current work, that both BPA and BPS can produce different effects on the T4, T3, and TSH levels. In this regard, it can be remarked that the experimental animals selected in the studies, the concentration and mode of administration of the test chemical, and the duration of the experiments might have led the researchers to obtain different results in the hormone levels related to thyroid function.
In the current study, BPS caused an increase in the PI values of TFCs with significant differences in the BPS-20 and BPS-100 groups. Antithyroid drugs (e.g., aminotriazole and propylthiouracil) or microsomal enzyme inducers (e.g., phenobarbital) induce biotransformation and the elimination of T4, and cause reduced serum T4 concentrations. The decreased T4 levels cause a reduction in the negative feedback effect in relation to TSH synthesis and secretion in the pituitary gland in rats. Next, an increased TSH level stimulates thyroid gland growth resulting in thyroid neoplasia (Hood et al., 1999a, 1999b). Microsomal enzyme inducers cause an increase in the serum TSH level (phenobarbital as well as pregnenolone-16-carbonitrile) and stimulate proliferation of TFCs, while they do not cause an increase in the serum TSH level (3-methylcholanthrene and Aroclor 1254) or in the proliferation of TFCs. Minimal increases in serum TSH due to the effects of phenobarbital as well as pregnenolone-16-carbonitrile have been reported to have caused increased TFC proliferation to a significant extent. On the other hand, although dramatic increases in TFC proliferation were observed in rats that had received treatment with those microsomal enzyme inducers, the total TFC count was reported to have remained unchanged (Hood et al., 1999b; Klaassen and Hood, 2001). It was proposed that a restricted increase in the total number of TFCs as a result of treatment with the microsomal enzyme inducers might have arisen from elevated levels of apoptosis in TFCs (Klaassen and Hood, 2001; Kolaja et al., 1999). In the current study, BPS was not seen to have caused any significant changes in the serum TSH levels, whereas in contrast to the antithyroid drugs or microsomal enzyme inducers, it increased the proliferation of TFCs. The past studies displayed presence of estrogen receptors in the thyroid tissue via immunohistochemistry or receptor binding assays (Chaudhuri et al., 1986; Clark et al., 1985; Giani et al., 1993; Inoue et al., 1993; Yane et al., 1994). Some in vivo and also in vitro research studies have been showed that estrogen and estrogenic drugs could induce thyrocyte proliferation and effect thyroid growth via estrogen receptors (Banu and Aruldhas, 2002; Banu et al., 2001, 2002a, 2002b; de Araujo et al., 2006; Manole et al., 2001). The administration of equine conjugated estrogens and tamoxifen has been reported to cause a proliferative effect on TFCs in ovariectomized rats and non-ovariectomized rats in the phases of the estrous cycle, and even significant decreases were found in the serum TSH levels (de Araujo et al., 2006). Accordingly, stimulation of proliferating TFCs by BPS without any significant increases in the serum TSH levels in this study may have occurred due to its estrogenic potential. However, further and more detailed studies should be conducted in order to uncover the reasons that underlie this effect by BPS.
Herein, apoptosis in TFCs was observed to be augmented with a dose increase after the BPS exposure. The AI value displayed significant elevation in the BPS-500 group. Tamura et al. (1998) reported that apoptosis is a basic mechanism for cell loss during the formation and involution of goiter and indicated that the Fas/FasL system is associated with increased thyroid follicle cell apoptosis. Furthermore, the researchers suggested that the critical balance that exists between apoptosis and cell proliferation might provide an important ability to control the functioning of the thyroid gland. On the other hand, microsomal enzyme inducers were reported to have induced apoptosis in the thyroid gland via an increased expression of transforming growth factor-β1 (TGF-β1), probably as a result of an increased TSH level, and such an effect may compensate for the hypertrophy and hyperplasia of the thyroid gland (Kolaja et al., 1999). Hexachlorobenzene, a microsomal enzyme inducer, upregulated TGF-β1 gene expression and induced apoptosis in rat thyroid glands without any increase in the plasma TSH levels. It was speculated that hexachlorobenzene can have an effect on those parameters via a direct effect on the gland itself instead of disrupting the homeostasis of the thyroid hormone (Chiappini et al., 2009). If the above-mentioned studies are taken into consideration, the increased apoptosis in TFCs in the present study following BPS exposure may have occurred in order to provide cellular balance in the gland or as a consequence of the direct effect that the BPS had on the TFCs. Moreover, it was reported that dysfunction or impairment of the thyroid and increased apoptosis in the gland may be related to ROS accumulation and OS (Kochman et al., 2021; Li et al., 2021). Thus, the reason for the increased number of apoptotic follicle cells following BPS treatment might have occurred also as a consequence of an elevated OS status in the gland.
Past studies have shown a close relationship between increased levels of ROS and toxicity sourced from BPA and its structural analogues, and OS in the tissues was reported to be induced in the wake of exposure to bisphenols (Li et al., 2022; Meli et al., 2020; Steffensen et al., 2020). The OS-inducing effect of BPS has been shown in the red blood cells of humans (Maćczak et al., 2017), RWPE-1 cells (Kose et al., 2020), mouse liver and kidney cells (Zhang et al., 2016b), and fish hepatocytes (Kaptaner et al., 2021) in vitro. It was also reported that BPS can induce OS in the liver (Zhang et al., 2018a) and testes (Ullah et al., 2016, 2018) in in vivo research in which mice and rats were used. However, there has not been any research conducted evaluating the adverse effects of BPS on the responses of antioxidant defenses in rat thyroid glands. It has been reported that BPA caused oxidative damage by increasing hydrogen peroxide production in PCCL3, the thyroid cell line, and also in rat thyroid glands (da Silva et al., 2018). As part of this study, increased activities of SOD and CAT, as well as GPx and GST were assayed in the thyroid gland after BPS exposure. The GSH content was reduced significantly in the BPS-100 group, while the MDA level was increased significantly in the BPS-500 group. Those findings displayed that BPS is capable of impairing the oxidant/antioxidant balance in the thyroid gland. The first ROS generated in the respiratory chain of all living aerobic organisms is the superoxide anion radical. SOD catalyzes the dismutation of the superoxide radical anion into hydrogen peroxide and water, and it constitutes the first line in the antioxidant defense (Abd El-Twab and Abdul-Hamid, 2016; Andrés et al., 2023; Salah et al., 2022). Increased SOD activity, as observed in this study, was found also in the liver of mice treated with BPA (Kabuto et al., 2003). BPF, a derivative of BPA, has been also reported to cause significant increases in the activity of SOD in hepatocytes that had been isolated in fish (Oncorhyncus mykiss), in line with the current results (Aykut and Kaptaner, 2021). The reason for such an effect seems to be due to elevated levels of the superoxide anion radical as a result of BPS exposure (Kabuto et al., 2003). Hydrogen peroxide, another oxidant that is formed by SOD activity, is then converted into water and molecular oxygen by CAT. In the case of saturation the former mechanism, GPx is activated as the second step of antioxidant defense (Piper et al., 1995; Yu et al., 2007). CAT activity was found to be increased in this study. Consistently, elevated CAT activities have been reported in the hepatocytes as well as the renal cells in mice that had been treated with BPS, and it has been shown that BPS could cause alterations in its structure and also induce the activity of the enzyme via binding to the Gly 117 residue found on its substrate channel. Moreover, it was expressed that BPS could increase the activity of CAT by means of stimulating ROS generation in those cells (Zhang et al., 2016b). The selenium-containing antioxidant enzyme GPx effectively detoxifies highly toxic HO2 and lipid peroxides using GSH as a cofactor (Łukaszewicz-Hussain, 2003). Increased activities of GPx in the thyroid tissue might have arisen from elevations of hydrogen peroxide and other hydroperoxides as the result of its neutralizing effect and preventative role in membrane damage (Sabry et al., 2022). GST is a phase-II enzyme that functions to protect cellular macromolecules from attack of reactive electrophiles. It provides elimination of xenobiotics by conjugating their electrophilic centers with GSH (Li, 2009; Townsend and Tew, 2003). In accordance with the current study, dose-dependent significant increases in the GST activity with other antioxidant enzymes, including SOD and GPx, were reported in the liver of chick embryos treated with BPA (Sravani et al., 2015). Increased GST activity was found also in the heart of rats exposed to BPA (Aboul Ezz et al., 2015). In a previous in vitro study in which primary hepatocytes from rainbow trout (Oncorhynchus mykiss) were used, the activities of GST, along with CAT and GPx, were reported to be increased significantly after exposure to BPS for a period of 24 h (Kaptaner et al., 2021). GSH is a tripeptide that is functionally related to cellular defense mechanisms and the metabolism of toxic compounds. GSH performs its role in those processes by conjugating directly with xenobiotics or serving as a substrate in detoxifying reactions that are catalyzed by some enzymes, such as GPx and GST, which need thiol-reducing equivalents. GSH is regenerated by the flavoenzyme glutathione reductase from oxidized glutathione within cells (Dringen et al., 2000; Lushchak, 2012; Sipes et al., 1986; Sznarkowska et al., 2017). GSH levels might be reduced or elevated in tissues, in the case of an increased OS status (Noeman et al., 2011; Nwogueze et al., 2021). Similar to the present study, decreased GSH levels indicating OS were reported as well in different tissues including the liver, testes, and pancreas of rats exposed to BPA (Kabuto et al., 2003; Korkmaz et al., 2010; Özaydın et al., 2018). On the other hand, BPA has been linked to significantly decreased levels of glutathione reductase activity, playing a role in the synthesis of GSH (Bindhumol et al., 2003). Even though the glutathione reductase activity was not measured, BPS might have also caused such an effect in this study, similar to BPA. Free radicals and ROS could attack the lipids such as polyunsaturated fatty acids, which possess carbon-carbon double bonds in biological membranes and cause there to be an impairment of the membrane function, ultimately leading cells to undergo apoptosis (Yin et al., 2011). MDA is highly mutagenic and preferred commonly as a useful biomarker for the determination of oxidative damage (Ayala et al., 2014). MDA levels in the thyroid gland were elevated with a dose-dependent increase in this study and showed a difference that was statistically significant at the highest dose. Similarly, Ullah et al. (2018) found that BPS could increase the level of MDA in rat sperm in vitro and the testicular tissue of rats in vivo. Moreover, in a study that was undertaken by da Silva et al. (2018), BPA was seen to have increased the ROS production and led to OS in the rat thyroid glands.
It was noted in the present study that some measured parameters (AI values, serum FT3 levels, and MDA content) showed a classic dose-related effect while some of them (i.e., activities of SOD and CAT) displayed “U”-shaped changes. The underlying mechanism for such a condition might arise from “hormesis effects.” ROS are generated by internal and external sources and balanced by antioxidant defense systems to maintain redox homeostasis. In this situation which is called oxidative eustress, ROS act as pleiotropic signaling molecules playing a role in cell growth, proliferation and differentiation. If ROS reaches to moderate levels in cells, an adaptive response known as “hormesis” is triggered to guarantee cell survival. On the other hand, elevated or supraphysiological levels of ROS could cause irreversible damage to biological macromolecules such as protein and DNA and induce lipid peroxidation (oxidative distress) (Nitti et al., 2022). Among the ROS, both hydrogen peroxide and superoxide anions have been implicated in hormesis effects and hydrogen peroxide possesses a crucial role in the induction of hormesis (Ludovico and Burhans, 2014). It was reported in recent studies that BPA or its derivatives might exhibit hormesis or hormesis-like effects (Fan et al., 2020; Liu et al., 2022).
Conclusions
This is the first to demonstrate that BPS, which is thought to be less harmful than BPA and is used an alternative in place of BPA in plastic manufacturing and industrial applications, could cause disruptions in the thyroid gland. The present study revealed that BPS induced histopathological lesions and led to follicle cell responses by increasing proliferation and inducing apoptosis in the thyroid gland. Moreover, it is capable of suppressing FT3 levels and impairing the antioxidant balance, resulting in OS. Data obtained from this study has led us to conclude that similar toxic effects reported for BPA in the thyroid gland of rats may occur as a consequence of BPS exposure and the function of the gland can be interfered with by BPS. Utilization of BPS as a safer alternative instead BPA should be approached cautiously, and the required measures should be taken considering the negative effects of BPS on the thyroid gland and its possible relation with thyroid diseases, like BPA.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Support for this work was undertaken by the Scientific Research Coordination Unit of Our University, under project number FYL-2021-9359.