
Abstract
Environmental pollutants are an important cause of depression. Dibromoacetonitrile (DBAN) is a disinfection by-product, which is neurotoxic and induces oxidative stress. This study aimed to elucidate whether and how DBAN provoked depression-like behavior. Mitogen-activated protein kinase phosphatase-1 (MKP-1) and P38 mitogen-activated protein kinase (MAPK) are key factors in depression. In mice, 8-week gavage with 20 mg/kg DBAN caused depressive symptoms, serotonin (5-HT) loss, hippocampal neuronal damage, and elevated MKP-1/P38 MAPK signaling; 80 mg/kg proved lethal. DBAN disturbed the intestinal flora of mice. Depression-related bacteria Firmicutes increased. Bacteroidota showed a decreasing trend, and the relative abundance of Bifidobacterium also showed a decreasing trend. Co-treatment with antioxidant N-acetylcysteine (NAC, 150 mg/kg) prevented mortality, restored 5-HT and norepinephrine, normalized MKP-1/P38 phosphorylation, and alleviated behavioral deficits. 10 μM DBAN raised reactive oxygen species (ROS), upregulated MKP-1, increased the phosphorylation levels of P38 and c-Jun amino-terminal kinase (JNK), and decreased the phosphorylation levels of the extracellular signal-regulated kinase (ERK1/2) in HT22 cells. Following HT22 cell MKP-1 knockdown and DBAN exposure, HT22 cells exhibited decreased MKP-1 expression along with decreased P38 and JNK phosphorylation levels. Still, the levels of ERK1/2 phosphorylation were not significantly affected. NAC exerts a prophylactic protective effect against these adverse outcomes.
Introduction
Environmental pollutants are a major contributing factor to the sharp rise in the number of patients suffering from depression recently. Epidemiological studies have demonstrated that long-term exposure to ambient air pollutants, including fine particulate matter (PM2.5), nitrogen dioxide (NO2), and black carbon, is significantly associated with increased risk of depression incidence (Fan et al., 2020; Yang et al., 2023). A large-scale prospective cohort study found consistent associations between multiple air pollutants and incident depression and anxiety disorders (Yang et al., 2023). Subsequent meta-analyses have further confirmed this robust association, with PM2.5 showing the strongest effect estimates (Bereziartua et al., 2026). Beyond air pollution, emerging evidence from heavy metal research has revealed independent associations between specific metals and depression. A recent study analyzing NHANES data from 2003 to 2020 identified urinary tungsten and antimony as independent risk factors for depression, while cadmium showed nonlinear dose-response relationships (Guo et al., 2026). Another multi-omics study confirmed that cadmium and antimony increased depression risk, with accelerated biological aging underlying this association (Kan et al., 2026). Additionally, a network toxicology and epidemiology integrated study revealed that cadmium-dominated heavy metal mixtures exhibited significant depression associations through oxidative stress pathways (Li et al., 2026). Intragastric administration of 44 mg/kg dichloroacetonitrile (DCAN) for 4 weeks caused movement disorders, inducing anxiety-like behaviors and oxidative stress in rats (Li et al., 2017). These findings supported that environmental pollutant exposure represented a significant and modifiable risk factor for depression.
DCAN is a drinking water disinfection by-product (DBP) and belongs to haloacetonitriles (HANs). With the increasingly serious global water pollution, disinfection of drinking water is essential to ensure its safety for consumption. Chlorine is widely used in drinking water disinfection, with low price and high efficiency, to prevent infectious diseases in water medium (Diana et al., 2019). It is currently the most popular method for disinfecting drinking water worldwide. Chlorinated disinfectants act by eliminating microorganisms in water. However, chlorine can also react with organic matter and pollutants present in water to form DBPs, which pose risks to the environment and human health (DeMarini, 2020). DBPs are recognized as environmental pollutants. Over 700 DBPs have been identified in water, including haloacetic acids (HAAs), trihalomethanes (THMs), and HANs, among others. THMs and HAAs are the most common DBPs (Koley et al., 2024). Many DBPs exhibited carcinogenic, cytotoxic, genotoxic, and neurotoxic. Compared to DBPs governed by the World Health Organization (WHO), some recently identified DBPs might be more harmful (Wagner and Plewa, 2017). HANs include DCAN, bromoacetonitrile (BAN), dibromoacetonitrile (DBAN), chloroacetonitrile (CAN), and bromochloroacetonitrile (BCAN) (Wei et al., 2020). In recent years, HANs have garnered considerable attention owing to their widespread presence in disinfected drinking water and recycled wastewater.
It had been discovered that following 4 weeks of giving SD rats drinking water with 20 and 100 μg/L DCAN, the body weight of rats decreased. The intestinal microflora and fecal metabolites varied (Xue et al., 2020). Additionally, the WHO, in its “Drinking Water Quality Guidelines” (Fourth Edition), stipulated a DBAN limit of 20 μg/L and a DCAN limit of 70 μg/L, indicating that DCAN below this limit may cause harm to animal health.
One target of HAN toxicity is the central nervous system. Our earlier research has shown that DBAN causes mouse hippocampus neurons (HT22 cells) to undergo autophagy, endoplasmic reticulum stress, and oxidative stress (Li et al., 2024). DBAN has stronger neurotoxicity than DCAN (Li et al., 2022). Additionally, Sayed et al. (2012) discovered that rats given 60 mg/kg DBAN for 7 days took longer to choose to leave the center area during the open field test, remained immobile for longer periods, walked and stood less, and spent less time on the revolving rod. Therefore, we hypothesize that DBAN mediates animal depression-like behavior.
The mitogen-activated protein kinase (MAPK) signaling pathway plays a pivotal role in transducing extracellular stimuli into intracellular signals, thereby orchestrating key cellular processes including mitosis, metabolism, gene expression, migration, and apoptosis (Chakraborty et al., 2023). This pathway comprises three subtypes, c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase (ERK1/2), and P38 MAPK (P38). Among them, P38 is a central regulator of diverse cellular events, such as apoptosis, cell proliferation, and stress responses. Activation of P38 is tightly coupled to environmental perturbations, including heat shock, oxidative stress, inflammatory cytokines, and various DNA-damaging agent (Martínez-Limón et al., 2020). Excessive reactive oxygen species (ROS) in vivo activates JNK and P38 protein expression. Depression is a well-recognized inducer of oxidative stress (Bhatt et al., 2020). Consistent with this, Liu et al. (2024) induced a mouse depression model with lipopolysaccharide (LPS). They found that the JNK/P38 pathway was turned on in the hippocampus of depressed mice due to elevated JNK and P38 phosphorylation levels and a decrease in the number of nerve cells in the hippocampus. A negative regulator of the MAPK signaling pathway is mitogen-activated protein kinase phosphatase-1 (MKP-1). MKP-1 imbalance can also lead to diseases such as depression and Alzheimer’s disease (Geng et al., 2024). Following the establishment of a depression model using chronic unpredictable mild stress (CUMS), Feng et al. (2024) discovered that autophagy was improved and MKP-1 expression was upregulated in the rat hippocampus.
Considering that HANs induce oxidative stress in nerve cells and DBAN causes neurobehavioral damage in rats. Given also that depression is associated with MKP-1/P38 MAPK signaling pathway disruption, we hypothesized that DBAN would affect this pathway and thereby mediate depression-like behavior in animals. HT22 cells and ICR mice were widely used as models for the study of depression (Jin et al., 2023).
To investigate whether DBAN-induced depression-like behavior in animals and the role the MKP-1/P38 MAPK signaling pathway plays in this process, ICR mice were employed as an in vivo model and HT22 cells as an in vitro model in this work.
Water treatment facility workers exposed to high concentrations of disinfection by-products (DBPs) during drinking water disinfection processes, chemical production personnel involved in the synthesis, packaging, and transportation of DBAN, and laboratory personnel handling reagents or standard solutions containing this chemical are all identified as high-risk populations for occupational exposure (U.S. Environmental Protection Agency, 2006; IARC, 2012). DBAN, as an industrial chemical, is widely used as an antimicrobial additive in metalworking fluids (U.S. Environmental Protection Agency, 2006). This results in occupational exposure risks through inhalation and dermal contact for workers engaged in the production, formulation, and application of metalworking fluids (U.S. Environmental Protection Agency, 2006; NIOSH, 2017).
Therefore, this study held significance not only in environmental toxicology but also bore direct relevance to the field of industrial health, providing neurotoxicity data to support the establishment of occupational exposure limits and worker health surveillance strategies.
Materials and methods
Experimental reagents and antibodies
DBAN was purchased from Adamas-beta (Shanghai, China). N-acetyl-L-cysteine (NAC) was obtained from Sigma-Aldrich (St. Louis, USA). Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), and reduced-serum medium (Opti-MEM) were purchased from Gibco (NY, USA). Bicinchoninic acid (BCA) protein assay kit, RIPA lysis buffer, polyacrylamide gel electrophoresis (SDS-PAGE) sample loading buffer, primary antibody dilution buffer, and protease inhibitor were obtained from Beyotime (Shanghai, China). Phosphoprotease inhibitors were purchased from Servicebio (Wuhan, China). Cell Counting Kit-8 (CCK-8) reagent was purchased from Vazyme (Nanjing, China). The siRNA oligo for MKP-1 was purchased from GenePharma (Shanghai, China). ROS generation and Kits for determining glutathione (GSH) content were obtained from Jiancheng Bioengineering Institute (Nanjing, China). Saline was obtained from Sinopharm Chemical Reagent (Shanghai, China). PBS was obtained from MeilunBi (Liaoning, China). Lipofectamine 2000 was obtained from Invitrogen (Carlsbad, USA). A sodium dodecyl sulfate-polyacrylamide gel kit was purchased from Vazyme (Nanjing, China). Polyvinylidene fluoride (PVDF film) was purchased from Sigma-Aldrich (St. Louis, USA). Skimmed milk powder and 4% paraformaldehyde were purchased from Biosharp (Anhui, China). MKP-1 and Goat Anti-Rabbit IgG antibodies were obtained from Abways (Shanghai, China). Antibodies of Phosphorylated-P38 (P-P38) and Phosphorylated-JNK (P-JNK) were obtained from Cell Signaling Technology (Boston, USA). Antibodies of P38, ERK1/2, and Phosphorylated-ERK1/2 (P-ERK1/2) were obtained from Abclonal (Wuhan, China). An antibody of JNK was obtained from Beyotime (Shanghai, China). An antibody of GAPDH was obtained from Engibody (Milwaukee, WI, USA). 5-HT and norepinephrine (NE) ELISA kits were obtained from Shanghai Enzyme-linked Biotechnology (Shanghai, China).
Experimental animals and treatment
Acute experiment: We designed a 7-day toxicology experiment to calculate the LD50 value and then estimate the dose of the later subchronic experiment. Mice were randomly divided into three groups: 240 mg/kg, 360 mg/kg, and 480 mg/kg, with 10 mice in each group. DBAN was dissolved in corn oil. At 9:00 am on the first day, 240, 360, and 480 mg/kg DBAN were administered by gavage, respectively. The body weight, status, and survival data were observed and recorded at 9:00 am on the next 6 days.
Grouping information and treatment protocols of the subchronic experiment.
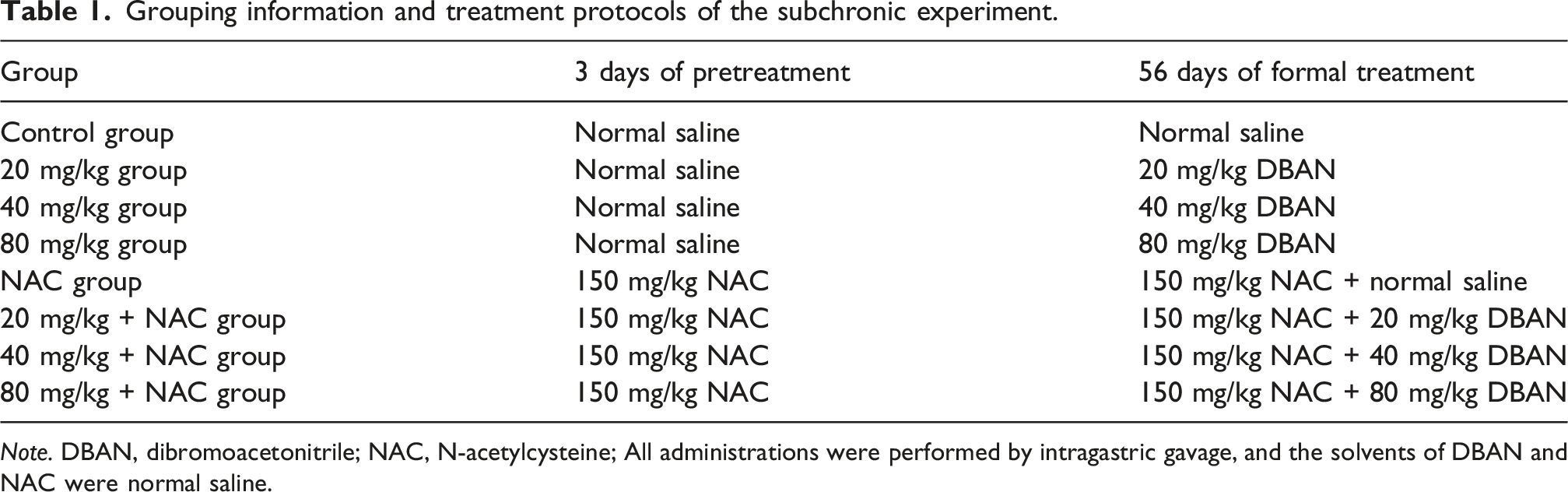
Note. DBAN, dibromoacetonitrile; NAC, N-acetylcysteine; All administrations were performed by intragastric gavage, and the solvents of DBAN and NAC were normal saline.
Behavioral test
(1) Open field test: An open field composed of opaque steel plates was situated in a dark room. The mice were positioned in the middle of the open field during the experiment. The mice were first adapted for 1 min, then were photographed with a camera for 5 min. MCS ActiMot-OF software recorded and analyzed the behavior and the total distance of movement within the next 5 min. After each experiment, the open field was cleaned, feces were removed, and the field was gently wiped with 75% alcohol to remove the remaining odor. (2) Grip test: TSE (Model: 303500-M/E-1) was used to test. The tail of the mouse was pinched. The forelimb of the mouse was grabbed by the same metal rod and pulled backward. Then the grasping force data were displayed by the instrument. (3) Forced swimming test: The mice spent 6 minutes swimming in an open barrel container with a height of 32 cm and a diameter of 22 cm. For 6 min, the water temperature was kept at 25°C while the container was filled with water that was deeper than the mouse’s length. The mice’s immobility time within the next 4 min was captured by a camera, which was based on the fact that the mice did not struggle on the water surface. (4) Tail suspension test: Each mouse was gently suspended by attaching adhesive tape to the distal portion of its tail, at a height of approximately 50 cm, for a total of 5 minutes. During the subsequent 4 minutes, the duration of aggressive struggling behavior was recorded using TSE software (Model: 303590-M).
Tar violet staining
After being preserved in 4% paraformaldehyde for 24 h, the mouse brain tissue was dehydrated, made translucent, dipped in wax, embedded in paraffin, cut, and stained with tar violet for pathological examination. The hippocampal dentate gyrus (DG) and Ammon’s horn (CA1) regions were observed and photographed with Caseviewer software.
ELISA
The hippocampus of mice was weighed, and normal saline was added at a ratio of 1:9 (1 g tissue +9 mL normal saline). After ultrasonic lysis, the contents of 5-HT and NE were measured according to the steps of ELISA kits. The steps of the ELISA assay were provided by the supplier of the ELISA kits, which were obtained from Shanghai Enzyme-linked Biotechnology (Shanghai, China). The specific steps were as follows: 50 µL of standard solution was added to the standard wells and 50 µL of the sample to be tested to the sample wells. Then, 100 µL of horseradish peroxidase-labeled antigen was added to each sample well and standard well, the plate was sealed, and incubated at 37°C for 1 h. The liquid in the wells was discarded, and the plate was patted dry, each well was filled with washing solution and allowed to stand for 20. The washing solution was discarded and the plate was patted dry. This step was repeated 5 times. Substrate A and substrate B were mixed at a ratio of 1:1, 100 µL of the mixed solution was added to each well, the plate was sealed, and incubated at 37°C for 15 min 50 µL of stop solution was added to each well, and the absorbance was measured at 450 nm with a microplate reader.
Immunohistochemistry
Mouse brain tissue was taken and paraffin sections were made and stained by Servicebio (Wuhan, China). Following the dewaxing and antigen repair, the sections were placed in PBS on a decolorization shaker and shaken for 5 min each. They were then incubated for 25 min in the dark in a 3% hydrogen peroxide solution at room temperature, then placed in PBS in the decolorization shaker shake and washed 3 times for 5 min each. 3% BSA was added to ensure that the tissue was evenly covered, and the tissue was sealed for 30 min at room temperature. The sections were incubated at 4°C for the entire night after the primary antibody was added. The slides were incubated with a secondary antibody at room temperature for 50 min after being submerged in PBS, shaken, and cleaned 3 times for 5 min each on a decolorization shaker. The slides were washed 3 times for 5 min each on a decolorization shaker after being submerged in PBS and shaken. The DAB chromogenic solution was added dropwise. The brownish-yellow color was observed under the microscope as positive, and the color was terminated by washing with tap water. After hematoxylin re-staining, the sections were rinsed with running water, and finally were dehydrated and sealed. ImageJ was used to analyze the proportion of brown positive particles.
Measurement of protein concentration
The hippocampus of mice was weighed, and the lysate was added at a ratio of 1:9 (1 g tissue +9 mL lysate). The sample was ultrasonically broken, and placed in a pre-cooled high-speed centrifuge at 18,800
Western blotting
To prepare the extracted protein samples for electrophoresis, a loading buffer was added. The 10% or 12% sodium dodecyl sulfate-polyacrylamide gel was prepared. The sample was added for electrophoresis. Following the transfer of the protein to the PVDF membrane, the membrane was blocked for 2 h using 5% skim milk in a shaker. Tris-buffered saline and Tween20 (TBST) were used to wash the membrane for 5 min, and then the solution was changed and repeated 3 times. After overnight incubation with the primary antibody at 4°C, the membrane was washed 3 times with TBST and incubated for 1 h with the secondary antibody at room temperature. Lastly, the MiniChemi Mini Size chemiluminescence imaging system (Sage Creation Science, Beijing, China) was used to detect the protein bands after they had been washed 3 times with TBST.
16S rDNA sequencing
After 8 weeks of administration, fresh feces of mice in control, 20, 40, and 80 mg/kg groups were collected for 16S rDNA sequencing. The samples were extracted, sequenced, and analyzed by Biotree (Shanghai, China).
Experimental cells and treatment methods
HT22 cells were purchased from Tongpai Biotechnology (Shanghai, China). Cells were cultivated in an incubator at 37°C, 5% CO2, and 95% humidity in high glucose DMEM media with 10% FBS. HT22 cells were treated with 0–10 μM DBAN for 24 h; after being pretreated with 2.5 mM NAC for 2 h, HT22 cells were co-treated with 10 μM DBAN for 24 h.
CCK-8
HT22 cells were inoculated in 96-well plates and treated with 0–10 μM DBAN for 24 h when the cells grew to about 60%; after being pretreated with 2.5 mM NAC for 2 h, HT22 cells were co-treated with 10 μM DBAN for 24 h. CCK-8 reagent was added and incubated at 37°C for 45 min. A microplate reader was used to measure the absorbance at 450 nm.
Measurement of protein concentration
HT22 cells were inoculated in a 6-well plate, and were treated according to the steps described in section “Experimental cells and treatment methods.” The cells were washed 3 times with PBS, and then the lysate as added. The cells were lysed for 15 min at 4°C. After that, the cell extract was scraped off. A pre-cooled high-speed centrifuge was used to centrifuge the cell extract for 15 min at 18,800
Determination of ROS generation
HT22 cells were inoculated in 6-well plates, and were treated according to the steps described in section “Experimental cells and treatment methods.” Blank tubes and positive control tubes were set up. After that, the cells were gathered and centrifuged at 2000
Measurement of intracellular GSH content
HT22 cells were inoculated in 6-well plates and treated according to the steps described in section “Experimental cells and treatment methods.” Following collection, the cells were washed twice with PBS. The GSH kit’s instructions were followed to measure the GSH content.
siRNA transfection
HT22 cells were inoculated in 6-well plates and washed with PBS when the cells had grown to about 60%. Opti-MEM medium was added to each well of the transfection group. The 20 μM MKP-1 siRNA and negative control were prepared and diluted with Opti-MEM medium with Lipofectamine 2000 transfection reagent to make transfection complexes. The concentration of negative control and MKP-1 siRNA was 80 nM, and each hole was added according to the group. After incubation at 37°C for 6 h, the transfection solution was sucked out and incubated with high glucose DMEM medium for 24 h. Then the post-treatment group was treated with 10 μM DBAN for 24 h, and Western blotting was performed. The sequence of the MKP-1 siRNA was 5'-GCCAAUUGUCCUAACCACUTT-3'.
Histopathological analysis
Following euthanasia, mouse brains were fixed in 4% paraformaldehyde, dehydrated, embedded in paraffin, and serially sectioned at 5 μm. Hematoxylin and eosin (HE) staining was performed according to standard protocols to evaluate general hippocampal morphology.
Statistical analysis
GraphPad Prism 9.0.0 was used to analyze the data, which are presented as mean ± standard deviation. After comparing the groups using one-way analysis of variance (ANOVA), the groups were statistically compared using Šídák’s multiple comparison test. The Log-rank test was used to compare the survival curve data. The difference was considered significant when p < 0.05.
Results
DBAN mediates depression-like behavior in mice and the antagonistic effect of NAC on it
Acute toxicity test for determination of subchronic toxicity dose
To determine the LD50 dose of DBAN in mice, we first performed a 7-d acute DBAN administration experiment. Mice in the 480 mg/kg group began to die at 6 h post-administration, whereas those in the 360 mg/kg group began to die at 8 h post-administration (Figures 1(a) and (d)). 6 h after treatment, the hair of mice in the 360 mg/kg group was rough and sweaty, and the mice in the 480 mg/kg group began to sleep (Figures 1(e) and (f)). 1 day after administration, the survival rate of the 360 mg/kg group decreased to 80%, and that of the 480 mg/kg group decreased to 25%. The survival rate of the 360 mg/kg group was reduced to 60% at 2 days after administration, while the survival rate of the 480 mg/kg group was only 10%. There was no death in the 240 mg/kg group 6 day after administration (Figure 1(b)). The average body weight of mice in the 240 mg/kg group decreased continuously from day 3 after administration, and the average body weight of mice in the 480 mg/kg group decreased continuously throughout the post-administration period (Figure 1(c)). The LD50 dose of DBAN in mice was calculated to be 398 mg/kg by SPSS software. The above results indicated that acute exposure to 360 and 480 mg/kg DBAN for 7 days can affect the body weight of mice and cause death in mice. Acute toxicity of DBAN exposure in mice. (a) Survival curve of mice within 8 h after administration. (b) Survival curve of mice within 6 days after administration. (c) Body weight changes of mice within 6 days after administration. (d) Representative moribund and dead mice during the observation period. (e) At 6 h after administration, mice in the 360 mg/kg group showed rough fur and piloerection. (f) At 6 h after administration, mice in the 480 mg/kg group exhibited lethargy and drowsiness. Mice were intragastrically administered DBAN and divided into 240 mg/kg, 360 mg/kg, and 480 mg/kg groups (n = 10 per group). Data are presented as the mean ± standard deviation (SD). *p < 0.05 vs. 480 mg/kg group; # p < 0.05 vs. 240 mg/kg group; DBAN: dibromoacetonitrile.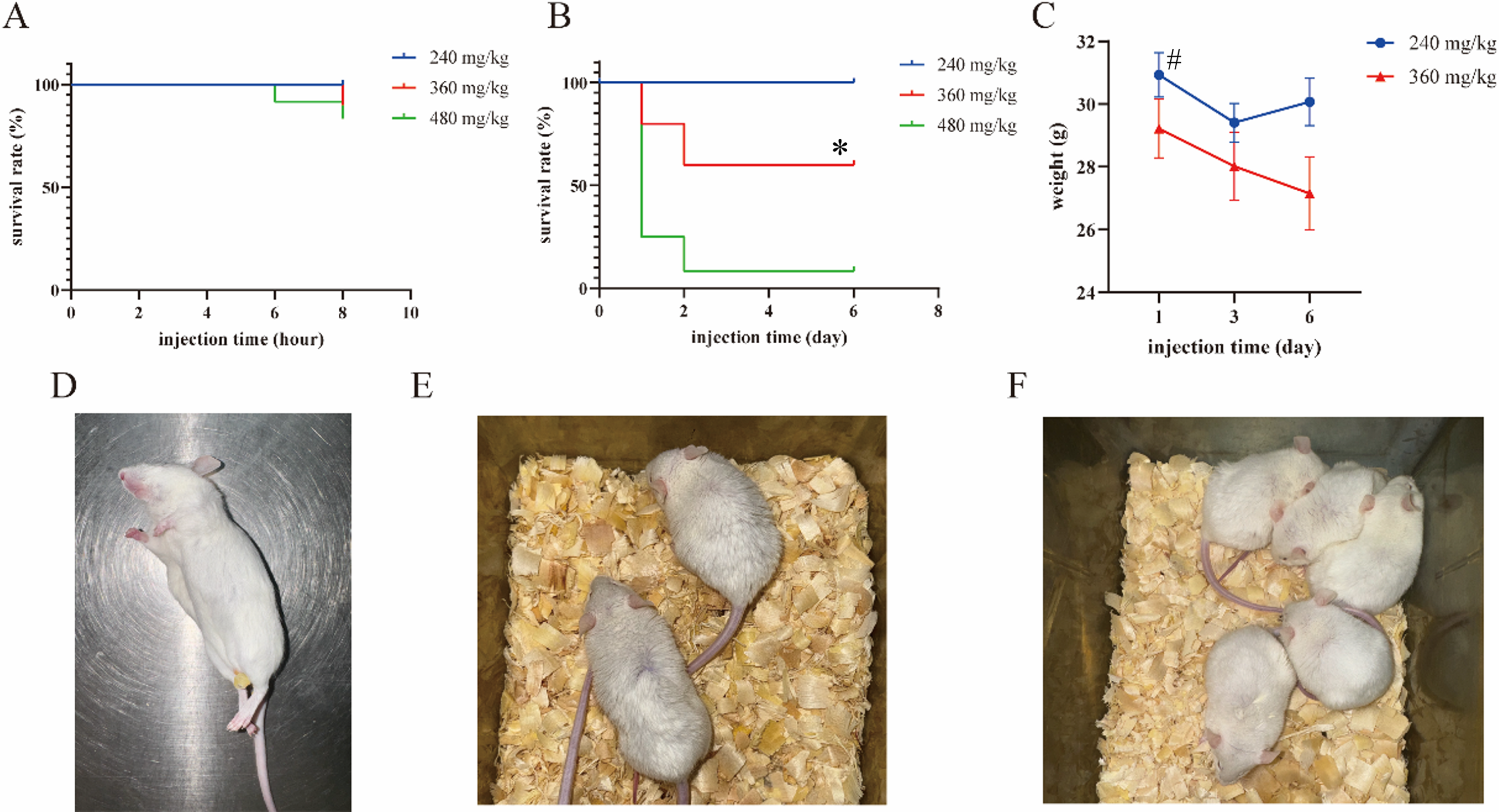
DBAN disrupts intestinal flora in mice
Emerging evidence suggests that intestinal flora abnormalities are linked to depression. To further study whether DBAN causes intestinal flora disorders in mice and changes in bacterial abundance such as Firmicutes, and Bacteroidota related to depression, an 8 weeks of subchronic toxic exposure was conducted. 16S rDNA sequencing was used to examine the intestinal flora of fresh feces of mice in control, 20, 40, and 80 mg/kg groups after administration. First, DATA2 was used to screen out amplicon sequence variants (ASVs). The numbers of general and specific ASVs in each group was obtained and presented in the Venn diagram (Figures 2(a)–(d)). The results revealed that, compared with the control group, the 40 and 80 mg/kg groups exhibited more unique ASVs than the 20 mg/kg group. Effects of 8-week DBAN exposure on intestinal flora in mice. (a–d) Venn diagrams of ASVs among different groups. (e) Violin plot of the Chao1 index for alpha diversity analysis. (f) Violin plot of the Shannon index for alpha diversity analysis. (g) Two-dimensional PCoA plot for beta diversity analysis based on unweighted UniFrac distance. (h) ANOSIM box plot for beta diversity analysis based on unweighted UniFrac distance. Mice were intragastrically administered DBAN and divided into control, 20 mg/kg, 40 mg/kg, and 80 mg/kg groups (n = 10 per group). Intestinal flora was analyzed using mouse fecal samples. ASV: amplicon sequence variant; PCoA: principal coordinate analysis; ANOSIM: analysis of similarities.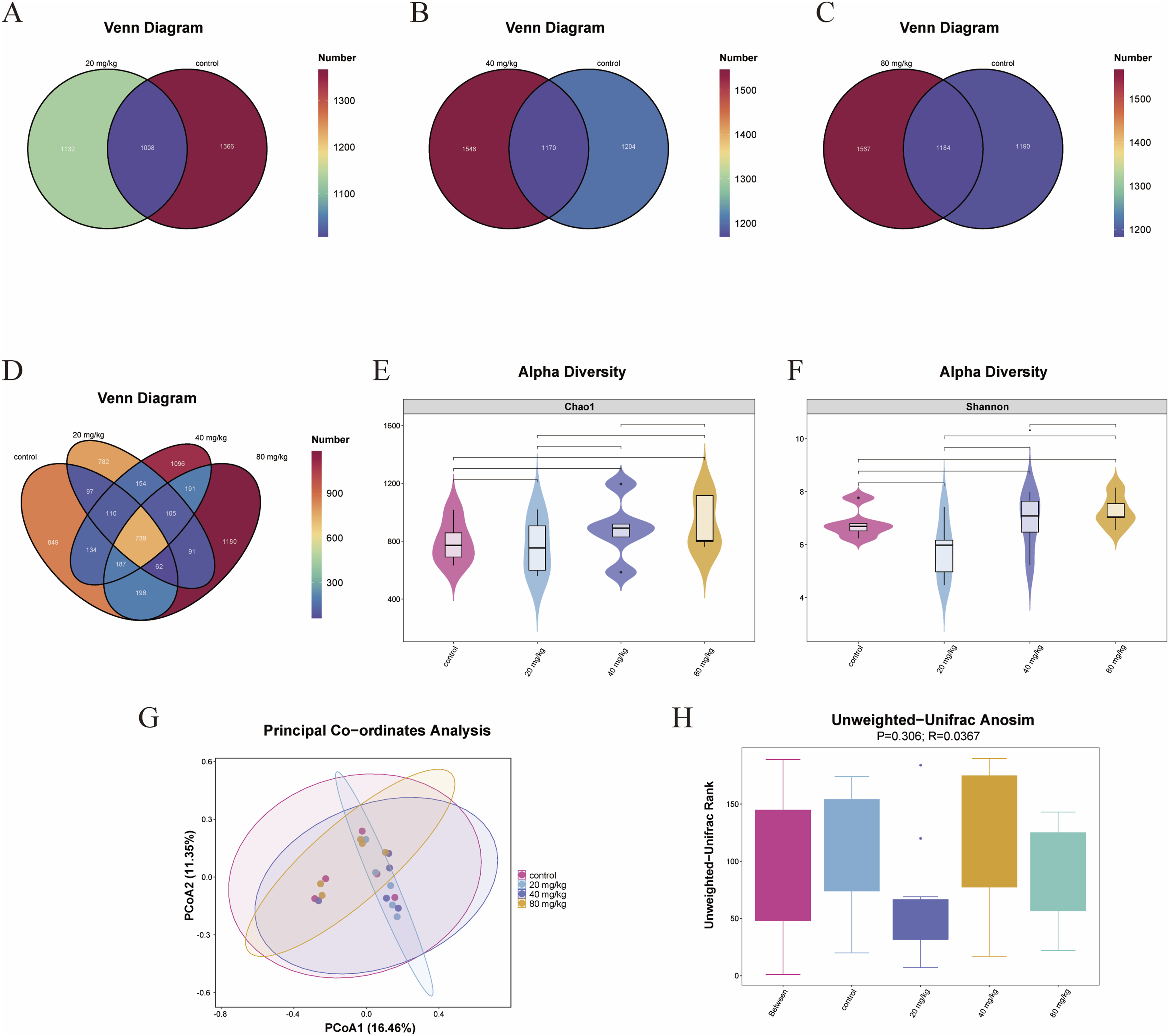
The variety and abundance of bacteria in environmental populations can be reflected in alpha diversity. Violin plots were used to display each group’s community richness Chao1 index and community diversity Shannon index (Figures 2(e) and (f)). Beta diversity reflects the difference of bacteria, and the difference between samples was evaluated by principal coordinate analysis (PCoA). The PCoA 2D diagram was shown in Figure 2(g). The farther the distance is, the greater the difference in microbial composition between the samples. To determine if the difference between groups was significantly larger than the difference within the group, the analysis of similarities was employed. The unweighted unifrac was used to evaluate the difference between the sample group and the group. The distribution characteristics of sample distance within and between groups were shown in detail by the ANOSIM box plot (Figure 2(h)), with larger R-values indicating greater differences in distance distribution between samples. These results indicated that DBAN altered the composition of gut flora in mice and induced gut flora disturbance.
Next, bacterial composition analysis was performed. The TOP30 most abundant bacteria at the phylum and genus levels were selected to show relative abundance. The relative abundance of Firmicutes in the three DBAN treatment groups rose at the phylum level as compared to the control group (control: 34.36%; 20 mg/kg: 15.9%; 40 mg/kg: 25.07%; 80 mg/kg: 30.96%), with a significant difference in the 20 mg/kg group. The relative abundance of Bacteroidota in the three DBAN administration groups showed a decreasing trend (control: 34.36%; 20 mg/kg: 15.9%; 40 mg/kg: 25.07%; 80 mg/kg: 30.96%), with no significant difference (Figure 3(a)). The relative abundance of Bifidobacterium in the 20 and 80 mg/kg groups exhibited a declining trend at the genus level when compared to the control group (control: 34.36%; 20 mg/kg: 15.9%; 40 mg/kg: 25.07%; 80 mg/kg: 30.96%), with no significant difference (Figure 3(b)). Heat maps of the top 30 bacteria at the phylum and genus levels were shown in Figures 3(c) and 3(d), where colors closer to red indicated higher bacterial abundance (Figures 3(c) and (d)). Effects of 8-week DBAN exposure on the relative abundance of intestinal microbiota in mice. (a) Relative abundance of the top 30 bacterial taxa at the phylum level in each group. (b) Relative abundance of the top 30 bacterial taxa at the genus level in each group. (c) Heatmap of the top 30 bacterial taxa at the phylum level in each group. (d) Heatmap of the top 30 bacterial taxa at the genus level in each group. Mice were intragastrically administered DBAN and divided into control, 20 mg/kg, 40 mg/kg, and 80 mg/kg groups (n = 10 per group). Intestinal microbiota analysis was performed using fecal samples. The top 30 most abundant taxa were selected for visualization and comparison.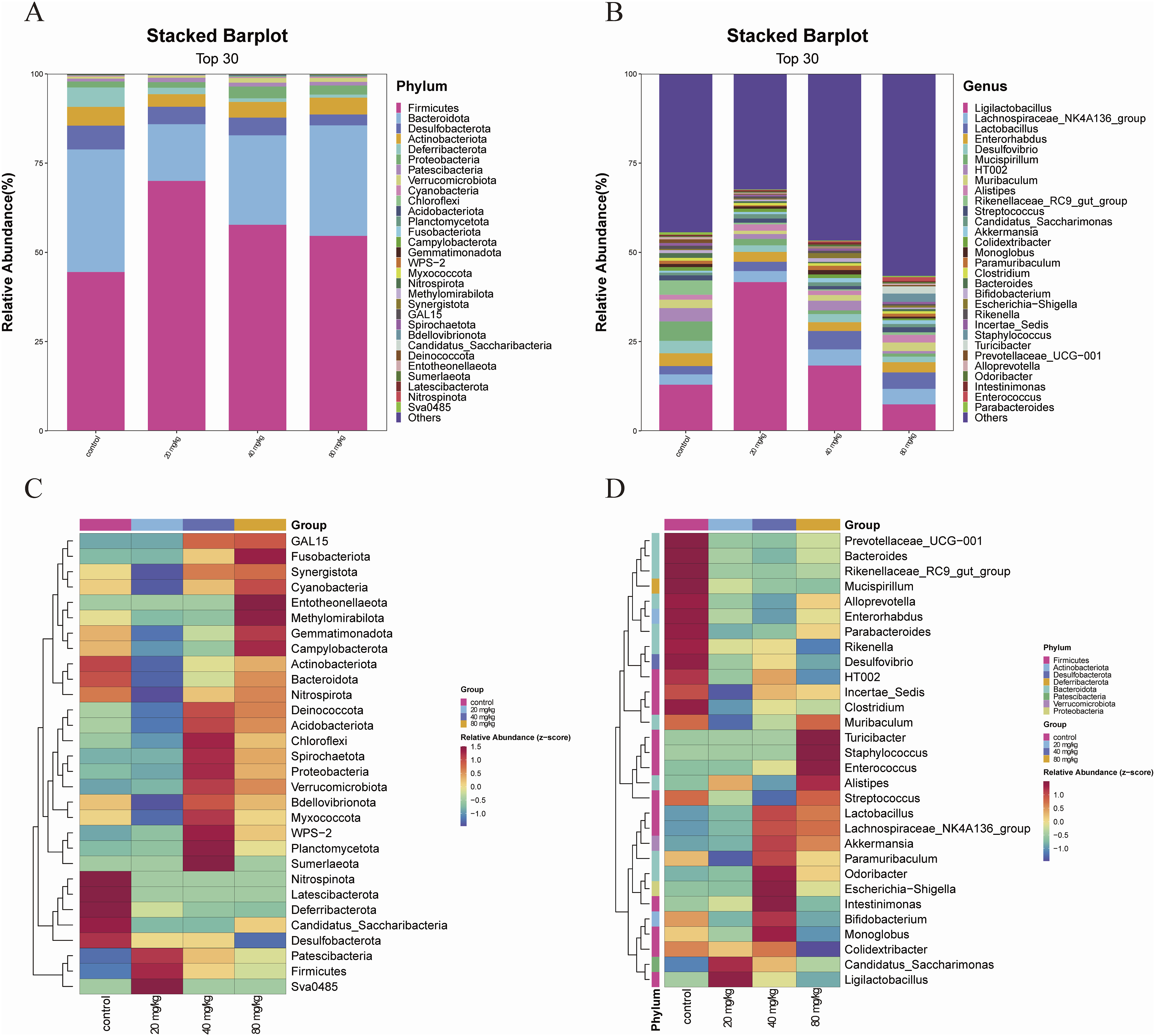
Effects of subchronic toxic DBAN administration on survival rate and body weight of mice
Mice in the 80 mg/kg group began to die on the 7th day of administration of DBAN, and the survival rate decreased to 50% on the 54th day of administration of DBAN, while the mice in the 80 mg/kg + NAC group all survived (Figure 4(a)). The survival rate of the 80 mg/kg + NAC group was significantly higher than that in the 80 mg/kg group. After the administration of DBAN, the average body weight of mice in the 80 mg/kg group decreased until 22nd day, then increased until 40th day, and subsequently decreased again. The average body weight of mice in the 80 mg/kg + NAC group decreased until the 16th day of administration. The average body weight decreased more slowly than that in the 80 mg/kg group and finally continued to increase (Figure 4(b)). These results indicated that 8 weeks of 80 mg/kg DBAN administration affected body weight and caused death in mice, whereas NAC pretreatment attenuated the rapid weight loss and delayed mortality. Effects of 8-week DBAN exposure on body weight, survival rate and behavioral performance in mice. Mice were divided into eight groups: control group, 20 mg/kg DBAN group, 40 mg/kg DBAN group, 80 mg/kg DBAN group, NAC group, 20 mg/kg DBAN + NAC group, 40 mg/kg DBAN + NAC group, and 80 mg/kg DBAN + NAC group. DBAN and NAC were administered by intragastric gavage, and the dose of NAC was 150 mg/kg. (a) Survival curve of mice in the 80 mg/kg DBAN group and 80 mg/kg DBAN + NAC group. (b) Changes in body weight of mice in different groups. (c, d) Total moving distance and time spent in the central zone in the open field test. (e) Grip strength of mice in the grip strength test. (f) Immobility time in the forced swimming test. (g) Struggling time in the tail suspension test. Data are presented as mean ± standard deviation (SD), with n = 10 mice per group. Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparison, followed by Šídák’s multiple comparison test. *p < 0.05, **p < 0.01, ****p < 0.0001 vs. the control group; bP< 0.05, bbbbP <0.0001 vs. 40 mg/kg group; cP< 0.05, ccP <0.01, ccccP <0.0001 vs. 80 mg/kg group. DBAN, dibromoacetonitrile; NAC, N-acetylcysteine.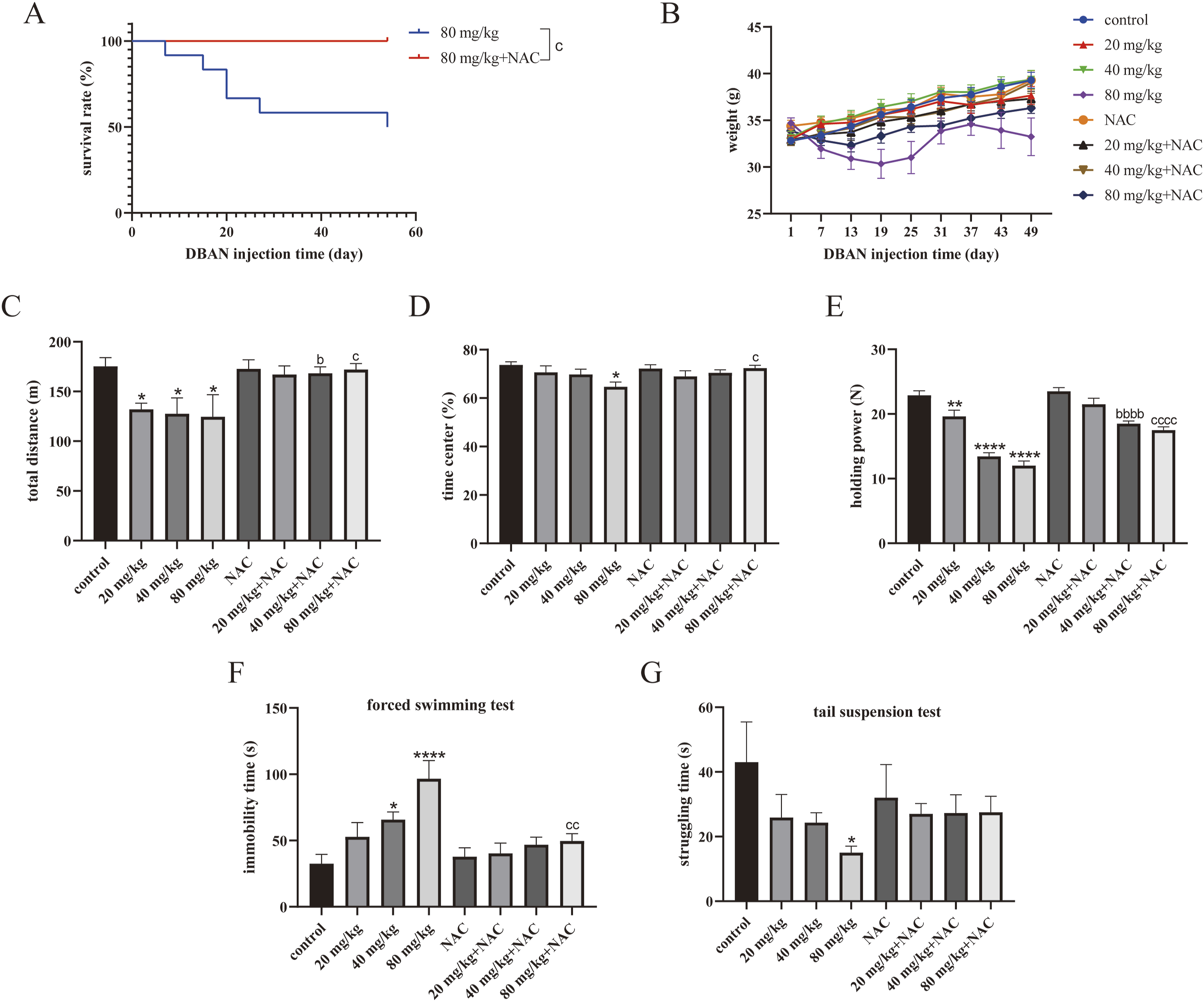
DBAN mediates depression-like behavior in mice
Open field test, tail suspension test, grip strength test, and forced swimming test were performed on mice. They are the main indicators for detecting depression-like behavior. (1) Open field test: Compared with the control group, the total distance of movement in the open field experiment of mice in the 20, 40, and 80 mg/kg groups was lower than that in the control group. The total distance of movement in the 40 mg/kg + NAC group and the 80 mg/kg + NAC group was more than that in their respective DBAN control groups (Figure 4(c)). The time in the central range of 80 mg/kg group was lower than that of the control group. The time of the 80 mg/kg + NAC group was higher than that of the 80 mg/kg group (Figure 4(d)). (2) Grip strength test: The mice in the 20, 40, and 80 mg/kg groups had weaker grips than the mice in the control group, according to the results. The grip strength of mice in the 40 mg/kg + NAC group and the 80 mg/kg + NAC group was higher than that in their administration group (Figure 4(e)). The results indicated that DBAN mediates depression-like behavior in mice, while NAC treatment alleviated depression-like behavior. (3) Forced swimming test: The mice in the 40 and 80 mg/kg groups had considerably longer immobility times than those in the control group, and the immobility time of mice in the 80 mg/kg + NAC group was less than that in 80 mg/kg group (Figure 4(f)). (4) Tail suspension test: The findings demonstrated that mice in the 80 mg/kg group had a shorter strenuous struggle time than those in the control group (Figure 4(g)).
DBAN affects the morphology of the hippocampus in mice
DBAN affects the morphology of the hippocampus in mice Hematoxylin and eosin (HE) staining was performed on mouse brain tissue sections to assess hippocampal morphological changes. The findings demonstrated that the control group’s hippocampal tissue cells exhibited normal morphology, with intact and neatly arranged structures... Effects of 8-week DBAN exposure on hippocampal morphology and neurotransmitter levels in mice. Mice were divided into eight groups: control group, 20 mg/kg DBAN group, 40 mg/kg DBAN group, 80 mg/kg DBAN group, NAC group, 20 mg/kg DBAN + NAC group, 40 mg/kg DBAN + NAC group, and 80 mg/kg DBAN + NAC group. DBAN and NAC were administered by intragastric gavage, and the dose of NAC was 150 mg/kg. (a) HE staining of the DG region in the mouse hippocampus. (b) HE staining of the CA1 region in the mouse hippocampus. (c, d) Levels of neurotransmitters 5-HT and NE in mouse brain tissue. Scale bar = 50 μm. Red arrows indicate normal neuronal cells. Data are presented as mean ± standard deviation (SD), with n = 10 mice per group. Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparison, followed by Šídák’s multiple comparison test. *p < 0.05 vs. the control group; aaaaP <0.0001 vs. the 20 mg/kg group; bbbP <0.001 vs. the 40 mg/kg group; cP< 0.05, ccP <0.01 vs. the 80 mg/kg group. Scale: 50 μm. Red arrow: normal nerve cells. DBAN, dibromoacetonitrile; NAC, N-acetylcysteine. HE, hematoxylin eosin; DG, dentate gyrus; 5 HT, 5 hydroxytryptamine; NE, norepinephrine.
DBAN affects neurotransmitters in mouse brain tissue
5-HT and NE levels in brain tissue were measured. Mice in the 80 mg/kg group had less 5-HT in their brain tissue than mice in the control group. The content of 5-HT in the brain tissue of mice in the 80 mg/kg + NAC group was higher than that in the 80 mg/kg group (Figure 5(c)). The NE content in the brain tissue of the NAC treatment group increased significantly in comparison to their DBAN group, but there was no significant difference between the DBAN group and the control group (Figure 5(d)). The above results indicated that DBAN affects the level of neurotransmitter 5-HT in the brain of mice, while NAC treatment reversed the decrease of 5-HT level and induced NE.
DBAN affected the detection of MKP-1 and P-P38 in the hippocampus of mice
Sections of mouse brain tissue from the 80 mg/kg and 80 mg/kg + NAC groups were immunohistochemically stained. The findings demonstrated that in contrast to the control group, 80 mg/kg DBAN administration increased the detection of MKP-1 and P-P38 in the hippocampus of mice. After NAC treatment, the detection of P-P38 in the brain tissue was lower than that in the 80 mg/kg group, but the decrease of MKP-1 was not significant (Figures 6(a)–(d)). Effects of 8 week DBAN exposure on the expression of depression-related factors MKP-1 and P-P38 in mouse brain tissue. Mice were divided into eight groups: control group, 20 mg/kg DBAN group, 40 mg/kg DBAN group, 80 mg/kg DBAN group, NAC group, 20 mg/kg DBAN + NAC group, 40 mg/kg DBAN + NAC group, and 80 mg/kg DBAN + NAC group. DBAN and NAC were administered by intragastric gavage, and the dose of NAC was 150 mg/kg. (a) Immunohistochemical staining of MKP-1 in the DG region of the mouse hippocampus. (b) Immunohistochemical staining of P-P38 in the DG region of the mouse hippocampus. (c) Higher magnification immunohistochemical staining of MKP-1 in the DG region of the mouse hippocampus. (d) Higher magnification immunohistochemical staining of P-P38 in the DG region of the mouse hippocampus. (e, f) Western blot analysis of MKP-1 protein expression. Scale bar = 50 μm. Red arrows indicate positive cells. Data are presented as mean ± standard deviation (SD), with n = 10 mice per group. Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparisons, followed by Šídák’s multiple comparison test. *p < 0.05, **p < 0.01 vs. the control group; cP< 0.05 vs. 80 mg/kg group. DBAN, dibromoacetonitrile; NAC, N-acetylcysteine; MKP-1, mitogen-activated protein kinase phosphatase-1; P-P38, phosphorylated-P38 mitogen-activated protein kinase; IHC, immunohistochemical staining; DG, dentate gyrus; WB, Western blot.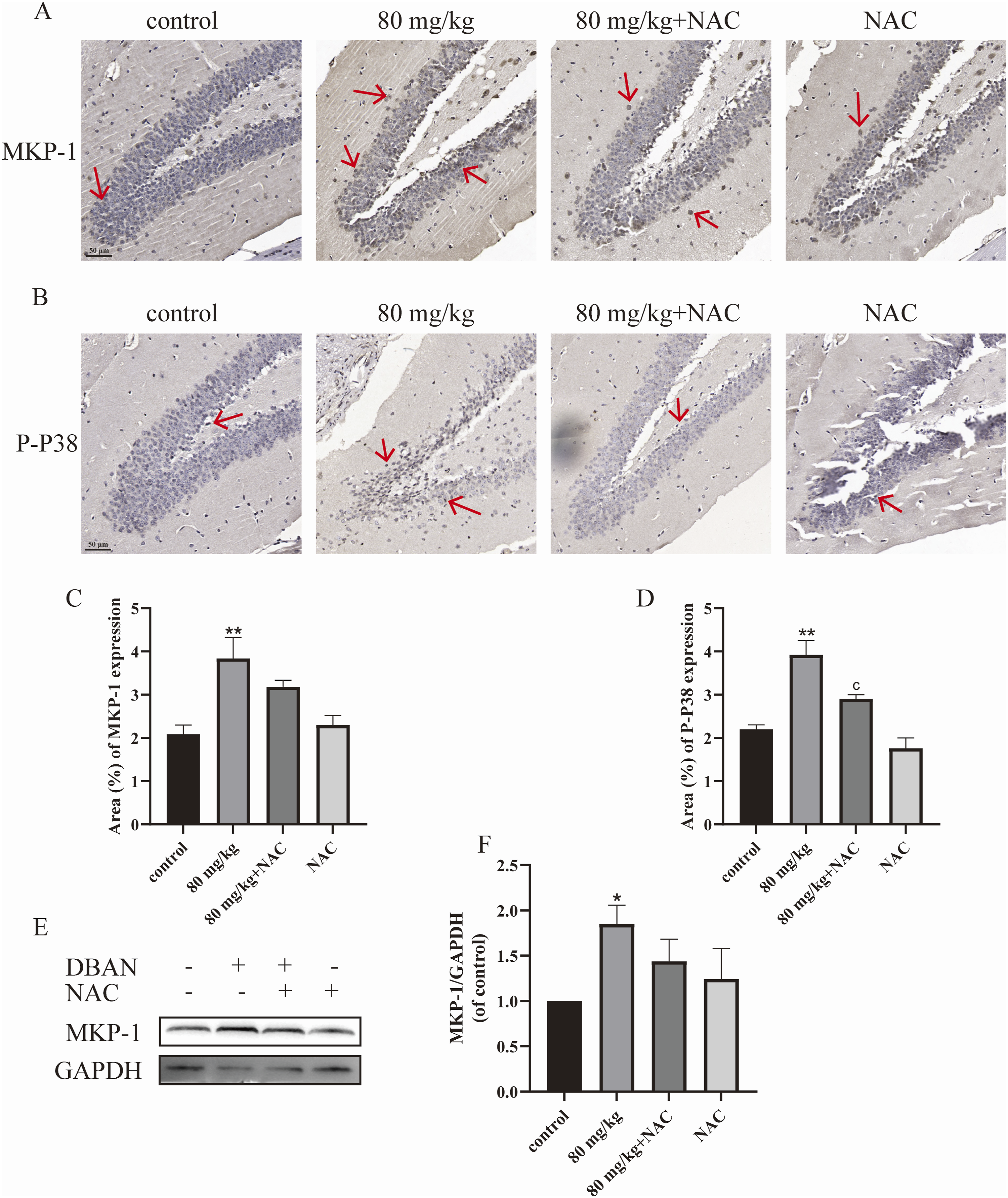
DBAN affects MKP-1 protein level
Western blotting was used to identify the MKP-1 protein expression in the hippocampal regions of the 80 mg/kg and 80 mg/kg + NAC groups. The findings demonstrated that the mice in the 80 mg/kg group had a considerably higher amount of MKP-1 protein in their hippocampal regions as compared to the control group. After NAC treatment, the MKP-1 level was alleviated compared with the 80 mg/kg group, but there was no significant difference (Figures 6(e) and (f)).
DBAN-induced cytotoxicity in HT22 cells
Following a 24 h treatment with several concentrations of DBAN (0, 2.5, 5, and 10 μM), the oxidative stress level of HT22 cells was determined by flow cytometry, and the viability of HT22 cells was assessed using the CCK-8 reagent. According to the findings, HT22 cell viability declined in a way that was dependent on concentration (Figure 7(a)). DBAN increased the ROS level of HT22 cells (Figures 7(b) and (c)). The above results indicate that DBAN can cause HT22 cell damage, and produce oxidative stress. Toxicity of DBAN to HT22 cells. HT22 cells were treated with different concentrations of DBAN (0, 2.5, 5, 10 μM) for 24 h. (a) Cell viability was determined using the CCK-8 assay. (b, c) Intracellular ROS levels were measured using the DCFH-DA fluorescent probe. (d) Protein expression levels related to the MAPK signaling pathway were detected by Western blotting, showing representative bands of MKP-1, P38, JNK, ERK1/2, and their phosphorylated proteins. (e) The protein expression of MKP-1 was quantified and normalized to GAPDH. (f–h) The protein expression of P-P38, P-JNK, and P-ERK1/2 was quantified and normalized to total P38, JNK, and ERK1/2, respectively. Data are presented as mean ± standard deviation (SD). Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparisons, followed by Šídák’s multiple comparison test. *p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001 vs. the control group. DBAN, dibromoacetonitrile; CCK-8, Cell Counting Kit-8; ROS, reactive oxygen species; DCFH-DA, 2',7'-dichlorodihydrofluorescein diacetate; MAPK, mitogen-activated protein kinase; MKP-1, mitogen-activated protein kinase phosphatase-1; P38, P38 mitogen-activated protein kinase; JNK, c-Jun N-terminal kinase; ERK1/2, extracellular signal-regulated kinase 1/2; P-P38, phosphorylated-P38 mitogen-activated protein kinase; P-JNK, phosphorylated c-Jun N-terminal kinase; P-ERK1/2, phosphorylated extracellular signal-regulated kinase 1/2; GAPDH, glyceraldehyde-3 phosphate dehydrogenase.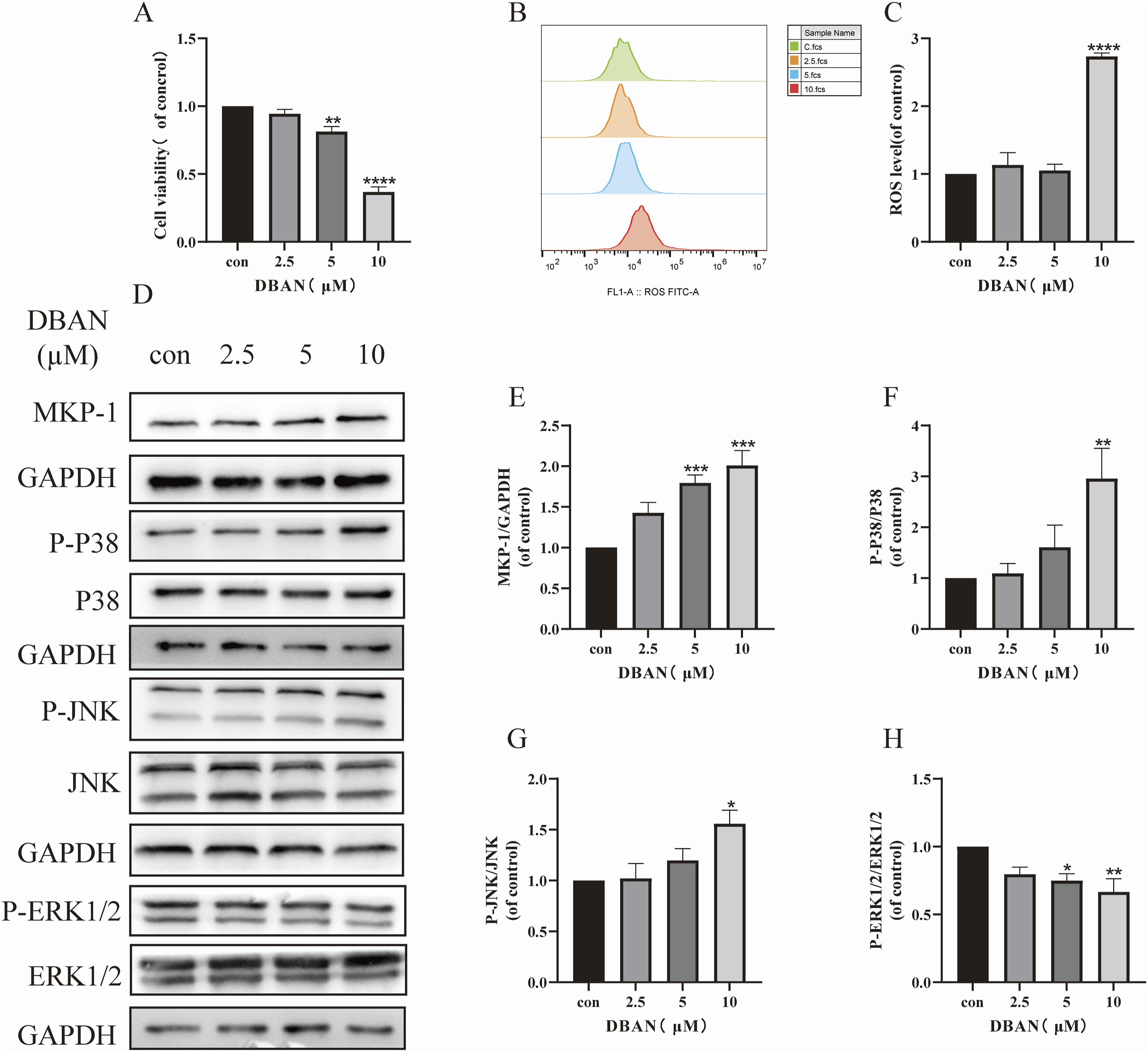
DBAN activated MKP-1 and MAPK signaling pathways in HT22 cells
Different DBAN concentrations (0, 2.5, 5, 10 μM) were applied to HT22 cells for 24 h. The MKP-1 protein and the phosphorylation levels of JNK, P38, and ERK1/2 proteins—the four primary proteins of the MAPK pathway—were detected in HT22 cells using Western blot. It was discovered that DBAN may decrease the expression of P-ERK1/2 protein, activate the expression of P-P38 and P-JNK protein, and raise the expression of MKP-1 protein in comparison to the control group (Figures 7(d)–(h)). The above results indicated that DBAN activated MKP-1 and MAPK signaling pathways, which were cytotoxic.
The effect of DBAN on MKP-1 and MAPK signaling pathways after MKP-1 knockdown
MKP-1 serves as a key regulator of the MAPK pathway and is critically implicated in depression. To elucidate its role in DBAN-induced neuronal injury, MKP-1 knockdown was performed using specific siRNA. The optimal ratio of MKP-1 siRNA to Lipofectamine 2000 was determined to yield a transfection efficiency of approximately 80% (Figure 8(a)). Among four candidate siRNAs, the sequence producing the most significant reduction in MKP-1 protein expression was selected for subsequent assays (Figures 8(b) and (c)). MKP-1 siRNA effectively suppressed MKP-1 expression in HT22 cells. Notably, DBAN-induced upregulation of MKP-1, P-JNK, and P-P38 was markedly reversed by MKP-1 knockdown, whereas P-ERK1/2 levels remained unchanged (Figures 8(d)–(h)). These results demonstrate that MKP-1 mediates DBAN-stimulated hyperphosphorylation of P38 and JNK, and that DBAN exerts cytotoxic affected on HT22 cells via regulating MKP-1 and the MAPK signaling components P38 and JNK. Effects of DBAN on MAPK signaling pathway-related proteins in HT22 cells after MKP-1 knockdown. HT22 cells were transfected with negative control (NC) or MKP-1 siRNA (small interfering RNA), and then treated with DBAN (0 or 10 μM) for 24 h where indicated. (a) FAM (Fluorescein Amidite) fluorescence negative control was transfected into HT22 cells, and the transfection efficiency was observed under a fluorescence microscope. (b, c) NC and MKP-1 siRNA were transfected into HT22 cells, and the expression of MKP-1 (Mitogen-Activated Protein Kinase Phosphatase-1) protein was detected by Western blotting. (d–h) NC and MKP-1 siRNA were transfected into HT22 cells and treated with DBAN (dibromoacetonitrile, 0 or 10 μM) for 24 h; Western blotting was used to detect the expression of MKP-1, P38, JNK, ERK1/2, and their phosphorylated proteins. Scale bar = 100 μm. Data are presented as mean ± standard deviation (SD). Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparisons, followed by Šídák’s multiple comparison test. * p < 0.05 vs. the NC group; ## p < 0.01, ### p < 0.001 vs. NC + DBAN group. DBAN, dibromoacetonitrile; MAPK, mitogen-activated protein kinase; MKP-1, mitogen-activated protein kinase phosphatase-1; siRNA, small interfering RNA; NC, negative control; FAM, fluorescein amidite; P38, P38 mitogen-activated protein kinase; JNK, c-Jun N-terminal kinase; ERK1/2, extracellular signal-regulated kinase 1/2.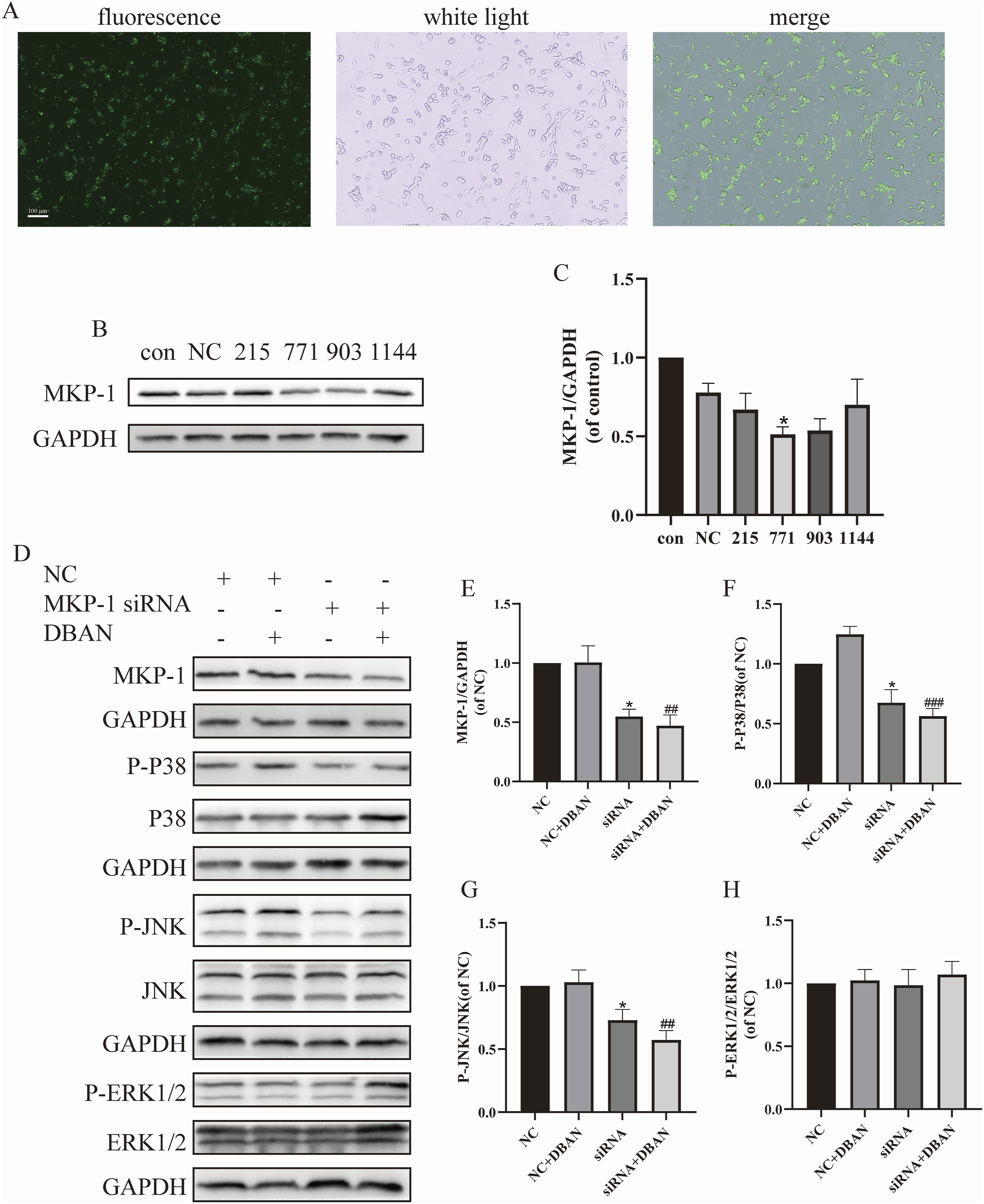
NAC regulates DBAN-induced MKP-1/P38 MAPK pathway dysregulation and oxidative stress
NAC is an antioxidant with neuroprotective effects (Arakawa and Ito, 2007). After 2 h of pretreatment with 2.5 mM NAC, HT22 cells were co-treated for 24 h with 10 μM DBAN. Variations in cell viability were detected using CCK-8. Changes in MKP-1 protein expression and phosphorylation levels of JNK, P38, and ERK1/2 proteins following NAC administration were identified by Western blotting. The results showed that compared with the DBAN treatment group, NAC pretreatment alleviated DBAN-induced HT22 cell damage (Figure 9(a)), significantly inhibited the expression of MKP-1, P-P38, and P-JNK proteins, and increased the expression of P-ERK1/2 (Figures 9(e)–(I)). HT22 cells were pretreated with NAC, and their ROS levels were measured by flow cytometry, it was found that NAC alleviated the increase of ROS level induced by DBAN (Figures 9(b) and (c)). The GSH content of HT22 cells after NAC pretreatment was detected by the GSH kit, and it was found that NAC alleviated the decrease of GSH content induced by DBAN (Figure 9(d)). The above results indicated that NAC can reverse DBAN-induced HT22 cell damage and regulate MKP-1/MAPK signaling pathway. Regulation of NAC on MKP-1/MAPK pathway and oxidative stress. HT22 cells were pretreated with 2.5 mM NAC for 2 h, and then co-treated with 10 μM DBAN for 24 h. (a) Cell viability was detected using the CCK-8 assay. (b, c) Intracellular ROS levels were detected by the DCFH-DA fluorescent probe. (d) Intracellular GSH content was detected by the GSH kit. (e–i) Western blotting was used to detect the expression of MKP-1, P38, JNK, ERK1/2, and their phosphorylated proteins. Data are presented as mean ± standard deviation (SD). Statistical analysis was performed using GraphPad Prism 9.0.0. One-way analysis of variance (ANOVA) was used for group comparisons, followed by Šídák’s multiple comparison test. *p < 0.05, **p < 0.01, ***p < 0.001 vs. the control group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. the DBAN treatment group. NAC, N-acetylcysteine; DBAN, dibromoacetonitrile; MKP-1, mitogen-activated protein kinase phosphatase-1; MAPK, mitogen-activated protein kinase; CCK-8, Cell Counting Kit-8; ROS, reactive oxygen species; DCFH-DA, 2',7'-dichlorodihydrofluorescein diacetate; GSH, glutathione; P38, P38 mitogen-activated protein kinase; JNK, c-Jun N-terminal kinase; ERK1/2, extracellular signal-regulated kinase 1/2.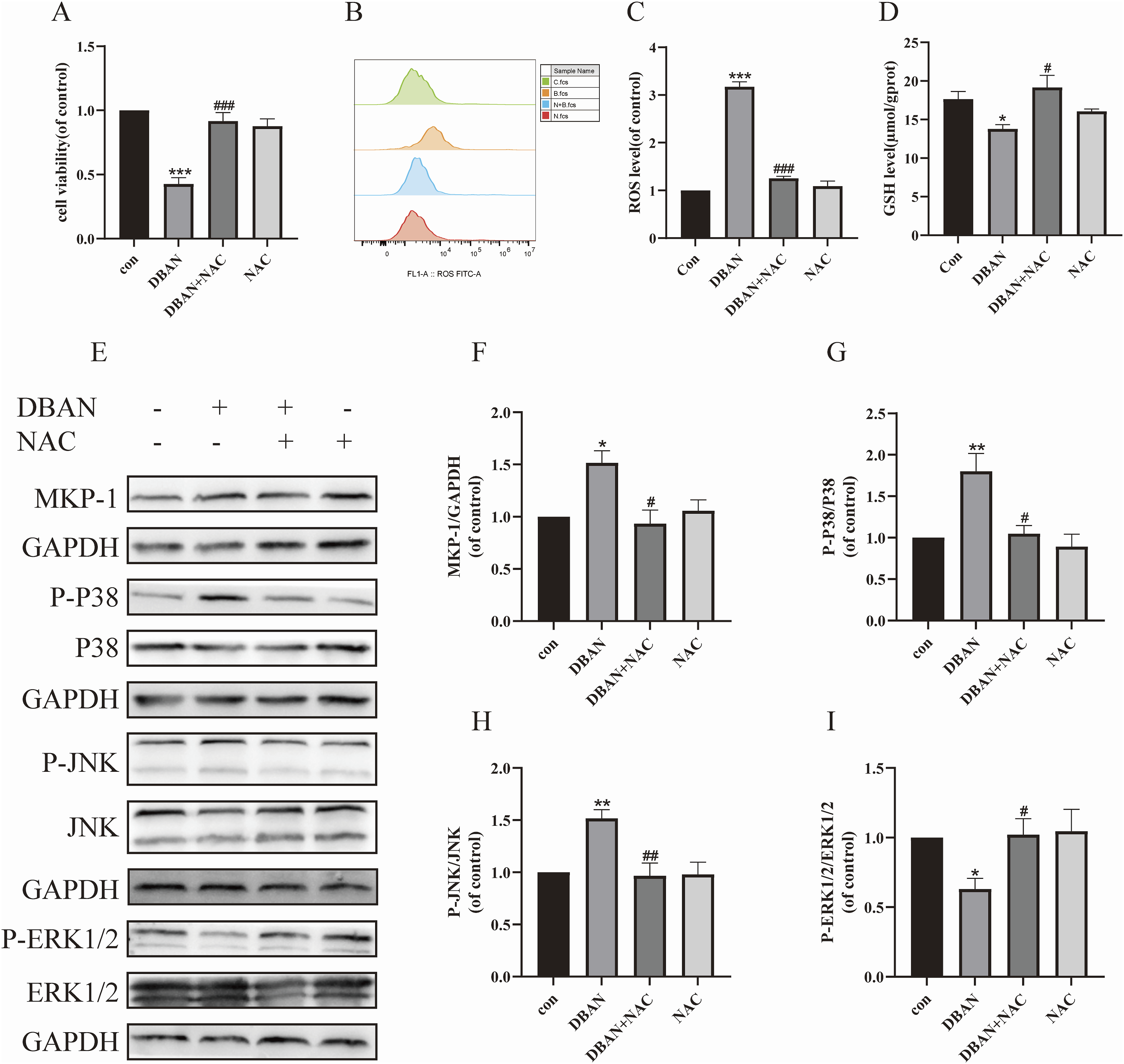
Discussion
In this study, DBAN was administered to HT22 cells and ICR mice. DBAN was found to mediate depression-like behavior in animals and affect to the MKP-1/P38 MAPK signaling pathway in HT22 cells. This study further evaluated NAC as an antidepressant agent capable of preventing and alleviating DBAN-induced depressive-like behavior. Depression is a complex disorder that may arise from a combination of psychological, biological, and environmental factors (Johns et al., 2025). In recent years, numerous environmental contaminants have been implicated in the onset and progression of depression. Consistent with this, our study demonstrated that DBAN exposure induced depression-like behavior in animals.
In mice with successfully established CUMS modeling, exploratory behavior was significantly reduced in the open field test, as evidenced by a shorter total distance traveled and a decreased number of rearing episodes. The sucrose preference test showed a reduced sucrose preference index, while the forced swimming and tail suspension tests showed longer periods of immobility (Zhou et al., 2024). According to these findings, our study showed that DBAN administration included depressive-like behavior in mice in a dose-dependent manner. Both the tail suspension test and the forced swimming test had longer depressed immobility times. Gradually, the grip strength weakened. Consequently, as the study showed, DBAN caused mice to exhibited depressive-like behavior. Depression is closely associated with structural alterations in key brain regions. The hippocampus, critical for learning and memory, exhibited atrophy and morphological abnormalities in depressed individuals, accompanied by impaired memory and cognitive function, along with reduced functional connectivity among the nucleus accumbens, frontal lobe, and parietal lobe (Tian et al., 2025). Our investigation revealed that the DG and CA1 regions of the hippocampus had increasingly loosening nerve cell arrangements, a profoundly stained nucleus, and the morphology was changed. The lack of neurotransmitters such as NE, 5-HT, and DA is also an important cause of clinical depression (Cui et al., 2024). Consistently, we found that 80 mg/kg DBAN administration reduced 5-HT content in mouse brain tissue.
Recent research has revealed a possible link between gut flora abnormalities and depression. Some intestinal bacteria, such as Bifidobacterium, can create metabolites called short-chain fatty acids (SCFAs), which can pass across the blood–brain barrier and disrupt the microbiota–gut–brain axis. At this time, the abundance of Bifidobacterium may decrease (Kim et al., 2023). Chen et al. identified four significantly altered bacterial phyla in patients with severe depression, characterized by markedly decreased abundance of Bacteroidetes and Proteobacteria and increased abundance of Firmicutes and Actinobacteria (Chen et al., 2018). Consistent with these observations, our study revealed that DBAN administration elevated the relative abundance of Firmicutes and tended to reduce Bacteroidota in mouse gut microbiota. Meanwhile, Bifidobacterium abundance was decreased in the 20 and 80 mg/kg DBAN groups. This provides more proof that DBAN treatment led to depression-like behaviors and abnormalities in the gut flora in mice.
According to related research, MKP-1 has a significant role in depression. Duric et al. (2010) evaluated the brain tissue of 21 patients with depression after death. They discovered that patients’ hippocampus DG and CA1 areas had far higher levels of the MKP-1 gene than those of healthy individuals. They also injected the mice with MKP-1 overexpression virus in the hippocampal DG region and found that the mice after MKP-1 overexpression had depression-like behavior similar to the CUMS model: reduced sucrose loss rate, extended immobility during the forced swimming test, and inability to flee during the active avoidance test. By giving rats chronic corticosterone, Langreck et al. caused depression-like behavior in them, and the hippocampal protein level of MKP-1 rose noticeably. Conversely, knockdown of MKP-1 via CRISPR/Cas9 attenuated the depression-like effect induced by chronic stress, further highlighting the critical role of MKP-1 in the onset and progression of depression (Langreck et al., 2021). In the present study, 80 mg/kg DBAN treatment for 8 weeks significantly upregulated MKP-1 protein expression in mouse hippocampus, and exposure to 5–10 μM DBAN for 24 h enhanced MKP-1 levels in HT22 cells. These results support that MKP-1 contributes critically to DBAN-elicited depressive-like behaviors.
MKP-1 functions as a negative regulator of the MAPK signaling pathway, among which p38 MAPK represents a key branch involved in neuroinflammation, depressive-like behaviors, and cytokine-mediated synaptic dysfunction. As reported by Liu et al., mice exposed to 40 mg/L NaAsO2 in drinking water for 12 weeks exhibited markedly elevated hippocampal and prefrontal cortical p38 phosphorylation accompanied by depressive-like behaviors, while 8 μM NaAsO2 treatment for 24 h increased p38 phosphorylation and ROS levels in HT22 cells (Liu et al., 2023a). Similarly, CUMS exposure for 6 weeks enhanced hippocampal p38 and JNK phosphorylation in rats, along with astrocytic ROS overproduction and inflammatory activation. These effects were reversed by antidepressant agents such as total ginsenosides (Qin et al., 2023). At present, new P38 inhibitors such as MW150 have been developed to treat depression, neuroinflammation, and other neurological diseases in clinical trials (Frazier et al., 2024). In the present study, DBAN administration upregulated MKP-1 and phosphorylated p38 levels in mouse hippocampus, and increased ROS production and phosphorylation of p38 and JNK in HT22 cells. These observations suggest that DBAN may trigger neuronal injury via the p38 MAPK and JNK pathways, and mediate depressive-like behaviors through the MKP-1/p38 MAPK signaling axis. Previous studies reported that morphine-dependent mice (a depressive model) displayed enhanced hippocampal MKP-1 expression and reduced ERK1/2 phosphorylation, which were restored by the MKP-1 inhibitor sanguinarine chloride (Jia et al., 2013). Consistent with this, DBAN treatment suppressed ERK1/2 phosphorylation in HT22 cells, whereas MKP-1 knockdown did not significantly alter p-ERK1/2 levels, implying that DBAN-induced depressive-like behaviors were less dependent on the ERK1/2 branch. Given that DBAN exposure also induced oxidative stress in HT22 cells, we hypothesized that DBAN exposure activates ROS and subsequently affected the MKP-1/P38 MAPK signaling axis.
According to recent reports, NAC may be the perfect chlorine quencher for organic DBPs, preventing both the deconstruction of DBPs brought on by residual chlorine and the ongoing production of DBPs during disinfection (Li et al., 2023). NAC is a common reducing agent and a precursor to GSH. It has antioxidant properties and can penetrate the blood–brain barrier (Mulè et al., 2024; Yifu, 2024;). Its antidepressant potential has also been investigated (Peng et al., 2024). NAC can alleviate the depressive symptoms that methotrexate causes in rats (Aslanlar et al., 2024). Patients with major depression also improved depressive symptoms after 12 weeks of NAC treatment with 2000 mg/d (Russell et al., 2023). We found that NAC alleviated DBAN-induced depression-like behavior in mice, protecting the morphology of nerve cells, and reversing the mice’s hippocampal overexpression of MKP-1 and P-P38. Consistent with our findings, Fan et al. (2020) pretreated CUMS rats with 300 mg/kg NAC for 5 weeks and found that NAC effectively prevented oxidative stress damage and depressive-like behavior in the CA1 region of the hippocampus. NAC also reversed the decrease of ERK phosphorylation and increased P38 and JNK phosphorylation. This was similar to the results of our study. At the same time, the 2-h pretreatment of HT22 cells with 2.5 mM NAC reduced the rise in P-P38 and P-JNK expression and ROS levels brought on by DBAN exposure. It also reversed the decrease of P-ERK1/2. So we believe that NAC alleviates nerve injury and animal depression-like symptoms by anti-oxidation and stabilizing the MKP-1/P38 MAPK signaling pathway. Furthermore, our results demonstrated that NAC inhibited the DBAN-induced increase in MKP-1 protein levels in both HT22 cells and mouse hippocampal tissue, further supporting the critical role of MKP-1 in depression. The fact that NAC is an antioxidant implies that DBAN exposure induced ROS activation in HT22 cells, ultimately disrupting the MKP-1/P38 MAPK signaling pathway. Although there are few studies that have connected MKP-1 to oxidative stress, it has also been shown that MKP-1 can cause oxidative stress (Xu et al., 2004).
Notably, despite MKP-1 acting as a canonical negative regulator of the MAPK pathway, MKP-1 knockdown in DBAN-treated HT22 cells reduced phosphorylation of p38 and JNK compared with the negative control group, whereas ERK1/2 phosphorylation remained unchanged. Similarly, Liu et al. used 10 Multiplicity of Infection (MOI) Mycobacterium bovis BCG to infect induced THP-1 macrophages. BCG caused apoptosis and inflammation, increased MKP-1 protein expression, and activated the MAPK pathway (JNK, P38 MAPK, ERK1/2). Knockdown of MKP-1 reversed the increased phosphorylation of JNK, P38, and ERK (Liu et al., 2023b). In the present study, MKP-1 appears to function as a positive regulator of the p38 MAPK pathway in DBAN-exposed HT22 cells. We hypothesize that DBAN triggered inflammatory response in HT22 cells, which in turn activate MKP-1 and p38 signaling, while MKP-1 knockdown may attenuate inflammation and thereby reduce p38 phosphorylation. This regulatory mechanism warranted further experimental validation.
Conclusion
In mice, DBAN triggered the hippocampus MKP-1/P38 MAPK signaling pathway and caused depression-like behavior. DBAN also induces neurotoxicity in HT22 cells by activating oxidative stress, thereby affecting MKP-1 and the three major branches of the MAPK signaling pathway, namely P38 MAPK, JNK, and ERK1/2. As an antioxidant and an emerging antidepressant, NAC could alleviate DBAN-induced depression-like behavior and hippocampal nerve injury in mice.
Footnotes
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by Social Development Guiding Science and Technology Plan Project of Zhenjiang (FZ2022026) and Social Development Guiding Science and Technology Plan Project of Jurong (ZA42307).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.