
Abstract
Immunological homeostasis relies on a sophisticated bidirectional dialogue between the immune and nervous systems, primarily coordinated by cytokine signaling. These peripheral signals access the central nervous system (CNS) via circumventricular organs and are integrated within the hypothalamus to mount appropriate homeostatic responses. Given that the immune system is under rigorous circadian control, circadian desynchrony—such as constant light exposure—often compromises this coordination, thereby increasing disease vulnerability. This study investigates the hypothesis that interleukin-6 (IL-6) is a pivotal mediator in this neuroimmune dialogue, specifically examining how astrocytic IL-6 trans-signaling modulates hypothalamic activity following an immune challenge. Utilizing male wild-type (WT) and GFAP-sgp130Fc (TG) mice—the latter genetically engineered to express a soluble gp130 fusion protein that selectively binds the IL-6/sIL-6R complex, thereby acting as a specific decoy receptor to inhibit astrocytic IL-6 trans-signaling without altering classical membrane-bound signaling—we assessed physiological and molecular responses under standard LD cycles and constant light (LL) conditions, the latter serving as a model for circadian desynchronization. Our results reveal that TG mice exhibit compromised circadian patterns and significantly altered thermoregulatory responses to lipopolysaccharide (LPS) compared to WT controls. Notably, TG mice under LL conditions showed a complete abolition of the thermoregulatory response to LPS, identifying a critical failure in homeostatic integration when both astrocytic signaling and circadian organization are impaired. Furthermore, c-Fos immunoreactivity within the suprachiasmatic nucleus and paraventricular nucleus indicated a profound suppression of hypothalamic activation in TG animals. Peripheral analysis of hepatic mRNA (IL-1β, IL-6, TLR4) confirmed genotype- and photoperiod-dependent variations, with TG mice displaying a markedly blunted inflammatory profile. These findings underscore the essential role of astrocytic IL-6 trans-signaling in neuroimmune communication and demonstrate that its absence, coupled with circadian disruption, induces a state of “neuronal deafness” that severely compromises the ability of the CNS to respond to inflammatory challenges.
Keywords
Introduction
Cytokines are multifunctional pleiotropic proteins essential for cell-to-cell communication and cellular activation. Among these, interleukin-6 (IL-6), initially identified as a B-cell differentiation factor, plays crucial roles not only in the systemic immune response but also in physiological and pathological processes within the peripheral and central nervous systems (CNS) (Lehrskov and Christensen, 2019). The neuroimmune dialogue is fundamentally bidirectional: immune cell products modulate neuronal activity and survival, while the CNS exerts tonic control over cytokine production. This interaction is strictly regulated within a temporal framework to maintain physiological homeostasis.
A primary integrator of these temporally organized neuroimmune signals is the hypothalamus, which acts as a bridge between metabolic and immune demands (Kamimura et al., 2020). Within the hypothalamus, the suprachiasmatic nucleus (SCN) within the hypothalamus serves as the master circadian pacemaker, orchestrating daily temporal organization by entraining peripheral circadian clocks throughout the body. Thus, the SCN provides the necessary rhythmic background for the timely coordination of immune responses and CNS signaling (Lu and Kim, 2022). Central to this coordinated response is the regulation of specific molecular mediators, where IL-6 stands out as a primary link between systemic timing and cellular action. The central actions of this cytokine are supported by the early characterization of high-affinity IL-6 binding sites within the Hypothalamus (Cornfield and Sills, 1991). The functional impact of IL-6 is determined by its specific signaling modality (Ding et al., 2024).
At the molecular level, IL-6 signals through 2 distinct pathways: classical and trans-signaling. Classical signaling requires the binding of IL-6 to the membrane-bound IL-6 receptor (mIL-6R), which subsequently associates with the signal-transducing receptor gp130 (Taga et al., 1989; Hibi et al., 1990; Saito et al., 1992). This process is restricted to specific cell types, such as hepatocytes and certain leukocytes. In contrast, IL-6 trans-signaling occurs when IL-6 binds to the soluble form of the receptor (sIL-6R), enabling the activation of gp130 on virtually all cell types (Schöbitz et al., 1995). While classical signaling is generally associated with homeostatic and regenerative functions, trans-signaling is primarily linked to pro-inflammatory responses (Rose-John and Heinrich, 1994). This distinction is particularly relevant in the CNS, where IL-6-dependent pathways contribute to neuroimmune communication and circadian regulation; however, the efficiency of these pathways is heavily dependent on the timing of their activation (Ding et al., 2024).
Aligned with this temporal regulation, the immune system operates under robust circadian control. Virtually all immunological variables, including cytokine production, exhibit marked daily cycles. This rhythmicity is exemplified by the release of IL-6 induced by lipopolysaccharide (LPS), which peaks during the subjective day (Adams et al., 2013), a process driven by functional circadian clocks within immune cells such as macrophages (Keller et al., 2009). Beyond its immune functions, IL-6 contributes to normal brain physiology, including the regulation of body weight, food intake, and energy expenditure (López-Ferreras et al., 2021). It also acts as a principal endogenous pyrogen, signaling to the hypothalamus to induce fever (Klir et al., 1993; Tanaka et al., 2014; Blomqvist and Engblom, 2018; Herrmann et al 2003). Furthermore, IL-6 is involved in complex neurobehavioral processes, such as sleep-wake cycles, emotional reactivity, sickness behavior and memory (Morrow and Opp, 2005; Baier et al., 2009; Armario et al., 1998; Bluthé et al., 2000). This multifaceted role is supported by the widespread expression of IL-6 and its receptors across the CNS—involving microglia, astrocytes, neurons, and endothelial cells—and its critical role in activating the hypothalamic-pituitary-adrenal (HPA) axis (Schöbitz et al., 1992; Mastorakos et al., 1993; Girotti et al., 2013), including the adrenal medulla (Gadient et al., 1995) as well as sympathetic ans sensory ganglia (Gadient and Otten, 1996).
The relationship between circadian desynchrony and immune pathology is well established. Chronic disruption—stemming from shift work, social jet lag, or environmental light interference—elevates LPS-induced IL-6 release by increasing the responsiveness of immune cells rather than altering their absolute number (Adams et al., 2013; Cermakian et al., 2013; Castanon-Cervantes et al., 2010; Guerrero-Vargas et al., 2015). This heightened sensitivity is mirrored at the genetic level; for instance, mice with disrupted clock genes, such as Cry1−/−/Cry2−/−, exhibit significantly enhanced pro-inflammatory cytokine levels (Hergenhan et al., 2020). Similarly, the nuclear receptor REV-ERBα plays a critical role in the circadian gating of innate immunity; its deficiency leads to elevated, non-oscillating IL-6 levels following an immune challenge (Gibbs et al., 2012).
To investigate the central mechanisms underlying this neuroimmune interaction, selective inhibition of IL-6 trans-signaling can be achieved using a soluble gp130 fusion protein (sgp130Fc). This protein acts as a specific decoy receptor that traps the IL-6/soluble IL-6 receptor (sIL-6R) complex completely preventing it from binding to and dimerizing membrane-bound gp130. Crucially, this mechanism blocks the pro-inflammatory trans-signaling pathway while preserving classical IL-6 signaling via cell-surface receptors transgenic (TG) mice expressing sgp130Fc under the control of the glial fibrillary acidic protein (GFAP) promoter provide a robust to interfere with this signaling cascade predominantly within astrocytes. This model facilitates a precise exploration of how altered cerebral IL-6 signaling impacts circadian regulation and neuroimmune integration, particularly when an immune challenge coincides with circadian disruption.
Based on this framework, we hypothesized that astrocytic IL-6 trans-signaling is a mechanical necessity for the CNS to integrate peripheral immune signals with the circadian system. Specifically, we proposed that blocking this pathway prevents the hypothalamus from coordinating a normal neuroimmune response, a failure that becomes particularly acute under the physiological stress of circadian desynchrony induced by constant light (LL).
Consequently, this work examines the circadian regulation of the immune system, emphasizing the mechanisms by which the SCN coordinates immune responses and integrates neuroimmune signals during inflammatory events. By elucidating the role of IL-6 in temporal regulation, this study contributes to a broader understanding of chronobiology and its clinical implications for immunological health.
Materials and Methods
Animals
This study was conducted using 8- to 10-week-old male mice from 2 distinct strains: wild-type (WT; C57BL/6) and GFAP-sgp130Fc transgenic mice (TG). The TG strain was originally generated by the research group of Prof. Stefan Rose-John (Kiel University, Germany) and subsequently provided for this study by Dr. Marco Atzori (Faculty of Sciences, UASLP). These TG mice are genetically engineered to express the sgp130Fc protein—a fusion protein consisting of the extracellular domain of gp130 and the Fc portion of human IgG1—under the transcriptional control of the GFAP promoter. This construct ensures that the decoy receptor, which selectively inhibits IL-6 trans-signaling, is predominantly synthesized, and secreted by astrocytes within the CNS.
Animals were housed individually in acrylic cages (27 × 31 × 15 cm) within sound-attenuated, environmentally controlled lockers. The ambient temperature was maintained at 22 °C ± 2 °C, with continuous airflow. Food (Purina Lab Chow 5001) and water were provided ad libitum. During a 3-week acclimatization period, mice were synchronized to a 12-h light/12-h dark cycle (12L:12D). In this photoperiod, lights-on was defined as zeitgeber time 0 (ZT0) and lights-off as ZT12. Following successful entrainment, animals were randomly assigned to their respective experimental protocols. All experimental procedures were approved by the Bioethics Committee for Research and Teaching of the Faculty of Chemical Sciences at the Autonomous University of San Luis Potosí (approval CEID2014030) and followed the Mexican standard for the care and use of laboratory animals (NOM-062-Z00-1999).
Experimental Design
Two experimental protocols were used: one to evaluate physiological responses after 24 h post-inflammatory challenge and another to assess neuronal activity 90 min post-challenge. Each protocol included both WT (n = 16) and TG mice (n = 16), further subdivided into synchronized (12L:12D) and desynchronized (constant light; LL) groups (n = 8 per subgroup). To desynchronize animals, these were exposed to constant light for 4 weeks. Each subgroup was divided into a control group and an experimental group. The experimental groups received a non-lethal intraperitoneal injection of LPS (500 μg/kg in 0.1 mL; Sigma-Aldrich), while control animals received an equal volume of saline.
Locomotor Activity Monitoring
General locomotor activity was continuously monitored using a combination of pressure and infrared sensors positioned beneath and above each cage. These behavioral data were digitized and automatically logged at 1-min intervals by a dedicated acquisition system for subsequent analysis. Data processing was conducted using SPAD9 software (version 1.1.2; Omnialva SA de CV). To visualize the circadian distribution of activity, double-plotted actograms were generated for each subject (n = 8 per group), with activity levels binned into 15-min intervals throughout the experimental duration.
Intra-Abdominal Temperature Sensors
Mice were anesthetized with a cocktail of ketamine and xylazine (90 mg/kg and 10 mg/kg, respectively; PiSa, Hidalgo, Mexico). Prior to sterilization and surgical implantation, iButtons (Maxim Integrated, San Jose, CA, USA) were pre-programmed via a “delayed start mission” to record core body temperature (Tb) at 15-min intervals. This configuration allowed the sensors to remain dormant during the initial postoperative phase and automatically initiate data logging exactly 7 days after surgery. This approach ensured that data collection only commenced once the animals had achieved physiological stability and recovered from surgical stress, without the need for additional handling or remote activation.
The surgical procedure involved a small midline laparotomy to facilitate the insertion of a sterilized iButton into the peritoneal cavity. The abdominal musculature and skin were subsequently sutured using black braided silk (Tramat, Mexico). For the analysis of the febrile response, Tb values were averaged into 15-min bins for each experimental group. The magnitude of the response to LPS was determined by calculating the temperature change (ΔTb) (Supplemental Figure S2) and the absolute core body temperature profiles (Figure 2) and, defined as the difference between the mean temperature of each 15-min post-injection interval and the baseline mean temperature recorded during the 1-h period immediately preceding the challenge.
Treatment Solutions and Reagents
For all experiments, systemic inflammation was induced using a single intraperitoneal (i.p.) injection of LPS from Escherichia coli (serotype 0127: B8; Lot No. 051M4004; Sigma-Aldrich, St. Louis, MO, USA). The lyophilized LPS was reconstituted with sterile, pyrogen-free 0.9% saline (Baxter, México) to prepare a 1 mg/mL stock solution, which was then aliquoted and stored at −20 °C. On the day of the experiment, aliquots were thawed and appropriately diluted with sterile 0.9% saline to achieve the final administration concentration of 500 µg/kg. At the designated time point (ZT3), mice in the control groups received an i.p. injection of the vehicle (1 mL/kg of 0.9% sterile, pyrogen-free saline).
Sample Collection
After completion of the 28-day LD or LL protocol, animals received an infusion of LPS or vehicle. Ninety minutes later, mice were deeply anesthetized with an overdose of sodium pentobarbital (Pisabental, PiSA, México; 65 mg/mL), and blood samples (≈250 μL per animal) were collected from all groups using Microvette/500 tubes (Sarstedt, Nümbrecht, Germany) (n = 6 per group). Samples were centrifuged at 7000 rpm for 7 min at 4 °C, and plasma was collected and stored in 60 μL aliquots at −80 °C until analysis. A 0.2 g segment from the left hepatic lobe was rapidly excised and immediately frozen at −80 °C. Subsequently, animals were transcardially perfused with 0.9% sterile saline followed by 4% paraformaldehyde (PFA; Sigma-Aldrich, St. Louis, MO) in phosphate buffer (0.1 M, pH 7.4). Brains were removed, post-fixed for 24 h in 4% PFA, and then cryoprotected in a 30% sucrose and 0.04% NaN3 solution (Amresco LLC) in phosphate-buffered saline (PBS, 0.1 M, pH 7.4) at 4 °C until further processing.
Cytokine Level Determination
Plasma concentrations of IL-6, IL-1β, and TNF-α were quantified using enzyme-linked immunosorbent assay (ELISA) kits (Abcam, USA), following the manufacturer’s instructions. Standard curves were generated for each cytokine, and concentrations were expressed as picograms per milliliter (pg/mL).
Semi-Quantitative RT-PCR
Total RNA was extracted from liver tissue using Trizol Reagent (Invitrogen) following the manufacturer’s instructions. Equal RNA amounts from at least 3 mice per group were used to generate complementary DNA by using the reverse transcriptase cDNA synthesis kit (Thermo, USA). For RT-PCR analysis of cytokines (IL-1β, IL-6, TNFα, CRP, INFγ and TLR4), 500 ng/μL of RNA per sample was used. Primer sequences are listed in Table 1. All data were normalized to the GAPDH expression level. The expression of mRNA was quantified using a Bio-Rad Personal Molecular Imager FX, and quantification was performed using ImageJ (RRID: SCR_003070).
Primer Sequences for Quantitative QRT-PCR.
Immunohistochemistry
Mice were deeply anesthetized with an overdose of sodium pentobarbital (Pisabental, PiSa, México; 65 mg/mL) and perfused transcardially with 150 mL of 0.9% sterile, pyrogen-free saline, followed by 150 mL 4% PFA diluted in phosphate buffer (PBS, 0.1 M, pH 7.2). Brains were removed, post fixed and kept in fixative for 24 h, at 4 °C, and cryoprotected in 30% sucrose for 3 to 4 days. Brains were frozen and cut into coronal sections of 30 μm at −20 °C. Sections were serially collected and processed for c-Fos immunohistochemistry. Free-floating SCN sections were incubated for 72 h under constant shaking at 4 °C with rabbit c-Fos antibody (1:1000; Santa Cruz). Subsequently, sections were rinsed and incubated (1 h) at room temperature with biotinylated donkey-antirabbit serum (1:200, Jackson, ImmunoResearch Laboratories Inc., Baltimore, USA), rinsed and incubated in avidin-biotin complex (1:500 Vector Laboratories) for 2 h; product visualization was obtained with 0.01% diaminobenzidine, 0.05% nickel ammonium sulfate, and 0.01% hydrogen peroxide for 6 min. Brain sections were mounted, dehydrated, and cover-slipped with microscopy Entellan New (Merck, Whitehouse Station, NJ).
Cell Count
To quantify immunoreactivity of c-Fos (Fos-IR) in the SCN, paraventricular nucleus (PVN), and arcuate nucleus (ARC), 3 representative sections in the antero-posterior axis were selected in accordance with the stereotaxic atlas (for SCN −0.34, −0.46, and −0.70, for PVN −0.58, −0.82, −1.04, and for ARC −1.34, −1.70, −2.30 anterior/posterior level to bregma) as is indicated in the stereotactic atlas (Franklin and Paxinos, 2008). Images were examined under a light microscope and captured using a 20× objective lens. Immunoreactive-positive nuclei in the SCN, PVN, and ARC were counted bilaterally using a computerized image analysis system (Image J, 1.42q, National Institutes of Health, Bethesda, MD) by determining the area by free hand for each side of each hypothalamic nuclei. The background was subtracted, and threshold was determined; particle analysis was set for particles of 1.0-2.0 circularity and 500-800 pixels. Background optic density was established in a nearby region lacking Fos-IR. Immunohistochemistry results were analyzed under light microscopy at 20× using ZEN blue (Carl Zeiss., 2011). Micrographs were taken from rostral, medial, and caudal sections of target brain regions (SCN, PVN, ARC) and c-Fos-positive cells were counted bilaterally using ImageJ (v1.53e).
Data Analysis
All data are expressed as the mean ± standard error of the mean (SEM). Core body temperature (Tb) dynamics were evaluated using a 2-way repeated-measures analysis of variance (ANOVA) with Treatment and Time as the primary factors. Plasma cytokine concentrations following LPS or vehicle administration were analyzed via a 2-way repeated-measures ANOVA, utilizing zeitgeber time (ZT) and Time post-injection as factors. For the analysis of hepatic cytokine mRNA expression, a 3-way ANOVA was employed to assess the main effects and interactions of Genotype, Lighting Condition, and Treatment. Neuronal activation, quantified as c-Fos immunoreactivity (c-Fos-IR) within the SCN, was analyzed using a 2-way ANOVA with zeitgeber time and treatment as independent variables. In all cases, significant interactions were further explored using Bonferroni’s post hoc test for multiple comparisons to identify specific differences between groups. All statistical analyses were performed using GraphPad Prism (version 8.0.2; GraphPad Software, San Diego, CA, USA). The threshold for statistical significance was set at α = 0.05.
Results
Circadian Evaluation of Locomotor Activity
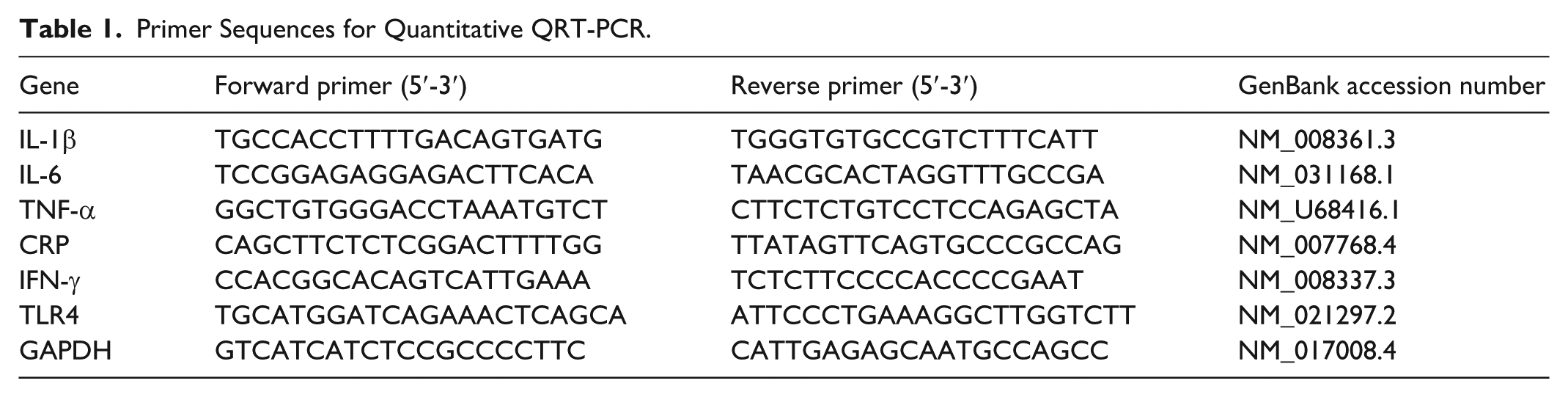
Under a 12:12 h light/dark photoperiod, control WT animals demonstrated robust and well-synchronized circadian rhythmicity (Figure 1a and c). This rhythm was characterized by increased activity during the dark phase, which represented 85% of total activity, and a marked reduction during the light phase (Figure 1b). In contrast, constant light (LL) exposure abolished this rhythmicity, resulting in evenly distributed activity across the 24-h period (Figure 1g-i).
Locomotor activity actograms in WT and TG mice under light/dark (LD) and constant light (LL) cycles. The actograms compare the activity patterns of WT and TG animals. Shading at the top of each panel represents the lighting schedule (black = dark, white = light). WT animals show a strong, synchronized rhythm under LD, with nocturnal activity (a-c). This rhythm is abolished under LL, resulting in disorganized, evenly distributed activity (g-i). TG animals display an abnormal LD pattern, notably with reduced activity late in the dark phase (d-f). Under LL, the TG mice exhibit a distinct free-running rhythm (j-l) characterized by an initial phase advance followed by a phase delay, demonstrating that the genetically modified IL-6 signaling alters the expected behavioral response to circadian desynchronization (n = 8 per group).
Transgenic (TG) animals displayed atypical circadian patterns under 12:12 h light/dark cycle (LD). Although their activity was primarily nocturnal (81% of total), it was notably reduced toward the end of the dark phase (ZT20) indicating reduced and less consolidated activity compared with WT controls (Figure 1d-f). When exposed to LL, TG animals remained rhythmic even after 3 weeks, unlike WT mice, which became arrhythmic (Figure 1j). Instead, TG mice maintained a free-running rhythm characterized by an initial period shortening (~23.5 h) followed by a gradual lengthening (26 h) (Figure 1k and l).
Thermoregulatory Response to LPS Administration in Light/Dark Conditions
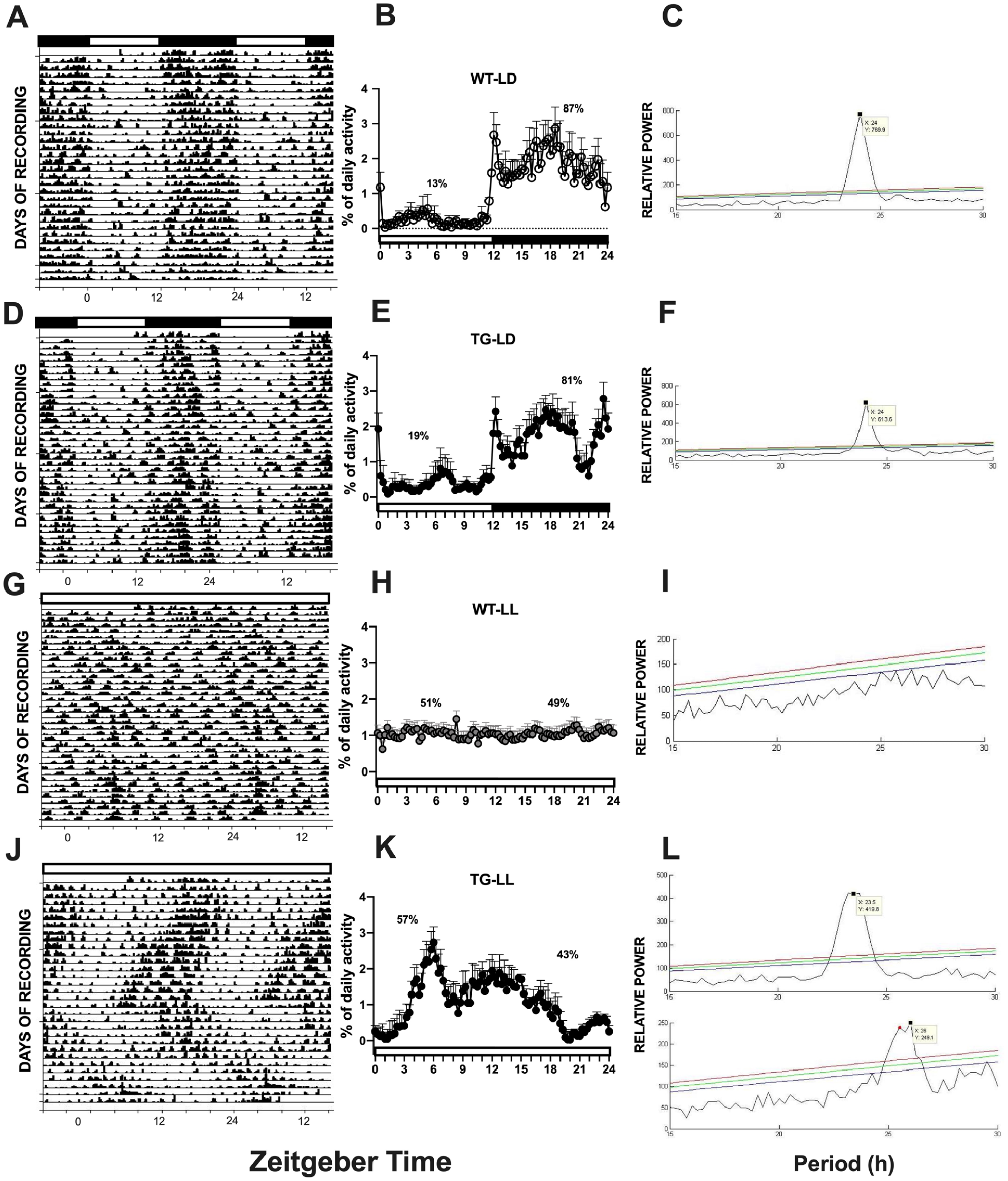
Control WT animals receiving saline (WT-SAL) showed no significant changes from their baseline temperature rhythm. In contrast, WT animals treated with LPS displayed a biphasic temperature response. An initial hyperthermic peak exceeding +2 °C was followed by a prolonged hypothermic period, with temperatures dropping to a maximum of −3 °C around 9 h after LPS treatment. Normal body temperature was restored by the 15-h mark post-injection (Figure 2a; n = 6-7, p < 0.05).
Core body temperature (Tb) dynamics in WT and TG mice during LPS challenge under LD and LL conditions. The traces illustrate absolute hourly temperature recordings across basal (black), saline (blue), and LPS (red) conditions. (a, c) Under LD cycles, both genotypes exhibit robust endogenous circadian oscillations. Following LPS administration, WT mice (a) display the characteristic biphasic febrile response, whereas TG mice (c) exhibit a sustained monophasic fever with an absent hypothermic phase. (b, d) Under constant light (LL), basal recordings reveal that the circadian temperature rhythm is severely compromised in WT mice (b) but remains remarkably preserved in TG mice (d). Notably, while WT mice exhibit an exacerbated acute fever under LL (b), TG mice are almost entirely unresponsive to LPS challenge (d), indicating a profound failure of central thermoregulatory integration under conditions of environmental desynchrony.
Saline-treated TG mice (TG-SAL) showed no significant changes from baseline core temperature. In contrast, LPS administration induced a monophasic fever, with a temperature increase of up to 2 °C. This hyperthermia was maintained for approximately 7 h, with temperatures starting to return to baseline just prior to the animals’ active phase (Figure 2c).
Thermoregulatory Response to LPS Administration in Constant Light
Under constant light (LL) condition, saline administration did not alter the body temperature of WT animals. However, LPS administration induced a temperature increase, with temperatures rising by up to 3 °C and normalizing within 4 h, notably without the secondary hypothermia observed in WT animals under a light-dark cycle (Figure 2b). Remarkably, TG mice exposed to LL conditions were completely unresponsive, showing no significant temperature changes after either saline or LPS administration (Figure 2d).
Relative mRNA Expression of IL-1β, IL-6, TNF-α, CRP, INF-γ, and TLR4 Genes in the Liver
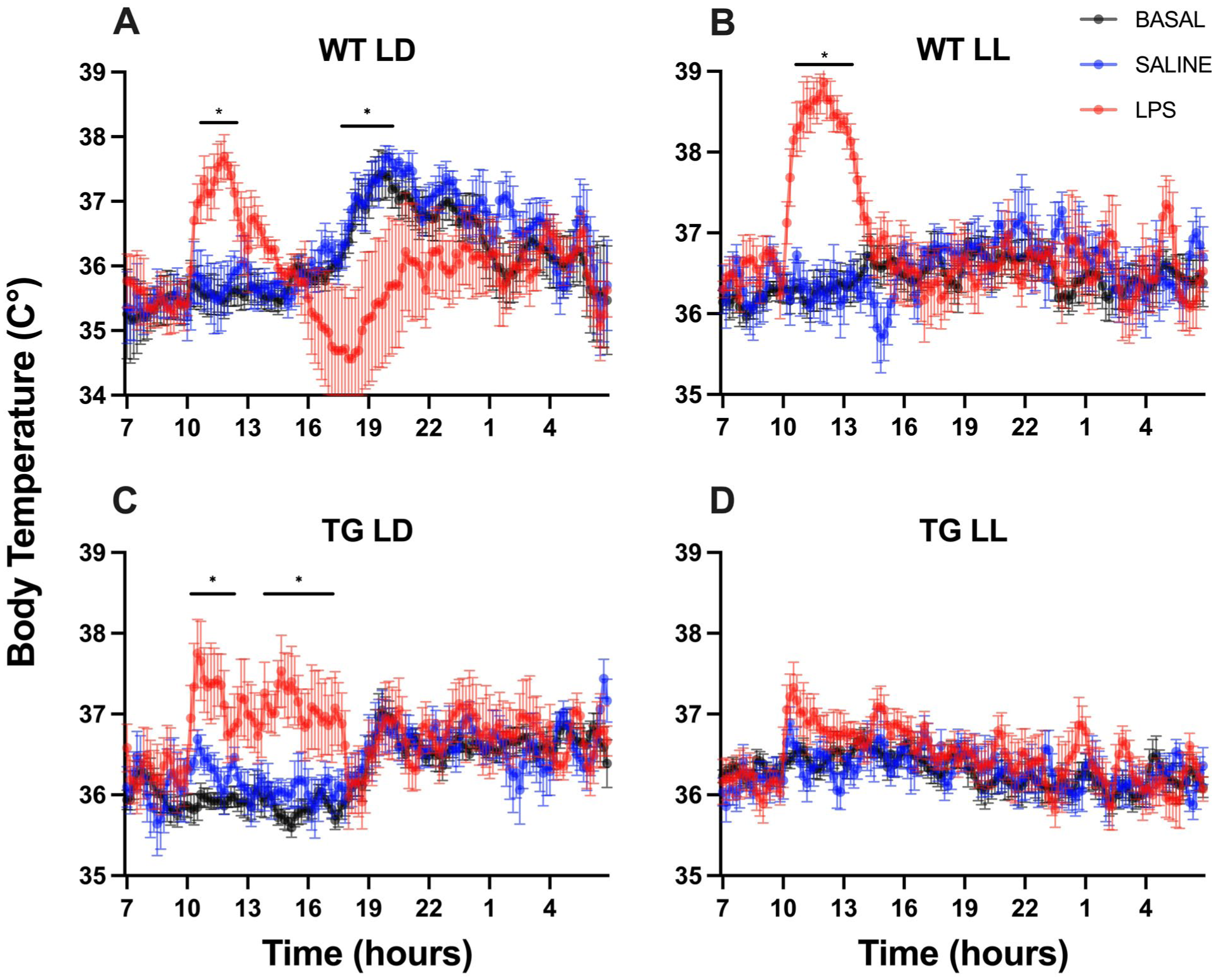
Under LD conditions, baseline IL-1β expression was low in the WT SAL group. Upon LPS administration, IL-1β expression significantly increased in WT LPS mice. The TG-LPS mice responded similarly to WT; however, though their baseline expression (TG SAL) was higher than in WT SAL mice (Figure 3a). Under constant light condition (LL), LPS-induced IL-1β expression was lower compared to LD mice. The TG SAL group in constant light showed a higher baseline IL-1β expression compared to WT SAL (Figure 3b).
Relative mRNA expression of immune-related genes in the liver under LD (ZT3) and LL (CT3) conditions. The images present the mRNA expression levels of multiple cytokines and receptors—IL-1β (a and b), IL-6 (c and d), TNF-α (e and f), CRP (g and h), IFN-γ (i and j), and TLR4 (k and l)—across 4 experimental groups: WT-Saline, WT-LPS, TG-Saline, and TG-LPS.
In LD conditions, WT LPS mice exhibited a significant increase in IL-6 expression compared to WT SAL mice. In contrast, TG LPS mice showed lower IL-6 expression levels than WT LPS mice, indicating that the LPS-induced increase in IL-6 was largely blunted or absent in the TG group. Baseline IL-6 levels in TG SAL mice remained low, like WT SAL (Figure 3c). Under LL conditions, IL-6 expression in both genotypes remained unchanged after either saline or LPS administration. However, the baseline expression levels in LL were higher than those observed in WT LD mice. Similar to TG-LD group, TG mice in LL do not respond to the LPS stimulus (Figure 3d).
Under LD conditions, basal TNF-α expression was low in both genotypes. TNF-α expression did not show a significant LPS-induced increase in either group (Figure 3e). When exposed to LL conditions, TNF-α exhibited a similar pattern, with no significant changes observed after LPS administration in both groups. Interestingly, the basal levels of TG animals were greater than those of WT mice (Figure 3f).
C-reactive protein expression was moderately, but not significantly, induced by LPS in WT animals exposed to LD conditions in both WT and TG genotypes (Figure 3g). When exposed to LL conditions, CRP expression was similar across all groups, with negligible changes in WT LPS mice (Figure 3h).
At both LD and LL conditions, WT LPS treatment led to a significant increase in INF-γ compared to WT SAL. Interestingly, in TG LPS mice under LD conditions, LPS treatment results in INF-γ levels comparable to those in TG SAL (Figure 3i). However, under LL conditions, INF-γ expression remained largely unchanged across all groups (Figure 3j).
Under an LD photoperiod, TLR4 was upregulated upon LPS stimulation in WT genotype, while TG mice showed non-significant changes after LPS treatment (Figure 3k). Under LL conditions, LPS induced increasing TLR4 levels in WT mice to approximately 0.6, while TLR4 expression remained largely unchanged after LPS in the TG LPS group (Figure 3l).
Cytokine Levels (ELISA)
In both LD and LL conditions, WT LPS consistently showed the highest concentrations of all 3 cytokines (IL-6, TNF-alpha, and IL-1β) compared to WT SAL group, indicating a strong inflammatory response (Figure 4). In TG LPS, the LD group exhibited a significant increase in IL-1β levels compared to their respective saline controls (TG SAL), though generally lower than those observed in WT LPS, suggesting a potentially modulated inflammatory response in transgenic animals when stimulated with LPS (Figure 4e). The WT SAL and TG SAL groups maintained high basal cytokine levels under constant lighting conditions.
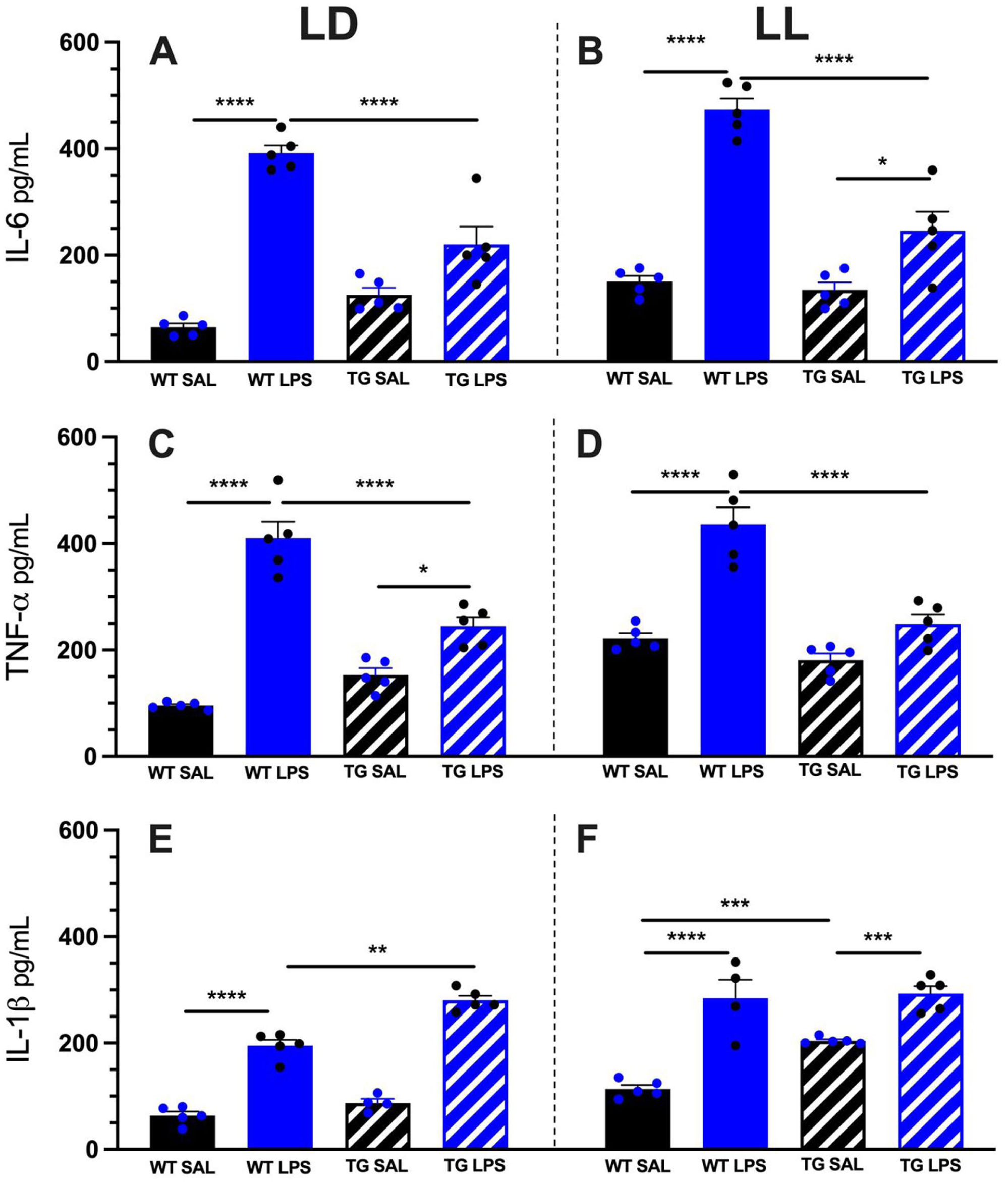
Serum cytokine concentrations (IL-6, TNF-α, and IL-1β) 3 h after LPS challenge in WT and TG mice. Serum levels of IL-6 (a and b), TNF-α (c and d), and IL-1β (e and f) were quantified by ELISA following i.p. injection of saline (SAL) or LPS in mice maintained under light/dark (LD) or constant light (LL).
Immunohistochemistry for c-Fos
Analysis of c-Fos immunoreactivity in the SCN revealed similar patterns in both WT and TG mice. For both genotypes, LL conditions significantly reduced c-Fos expression compared to a standard LD cycle. Within each lighting condition, LPS administration induced higher c-Fos expression than saline. However, the overall immunoreactivity in TG mice under LL was markedly lower than in all other groups, remaining suppressed regardless of LPS treatment (Figures 5a and 6a).
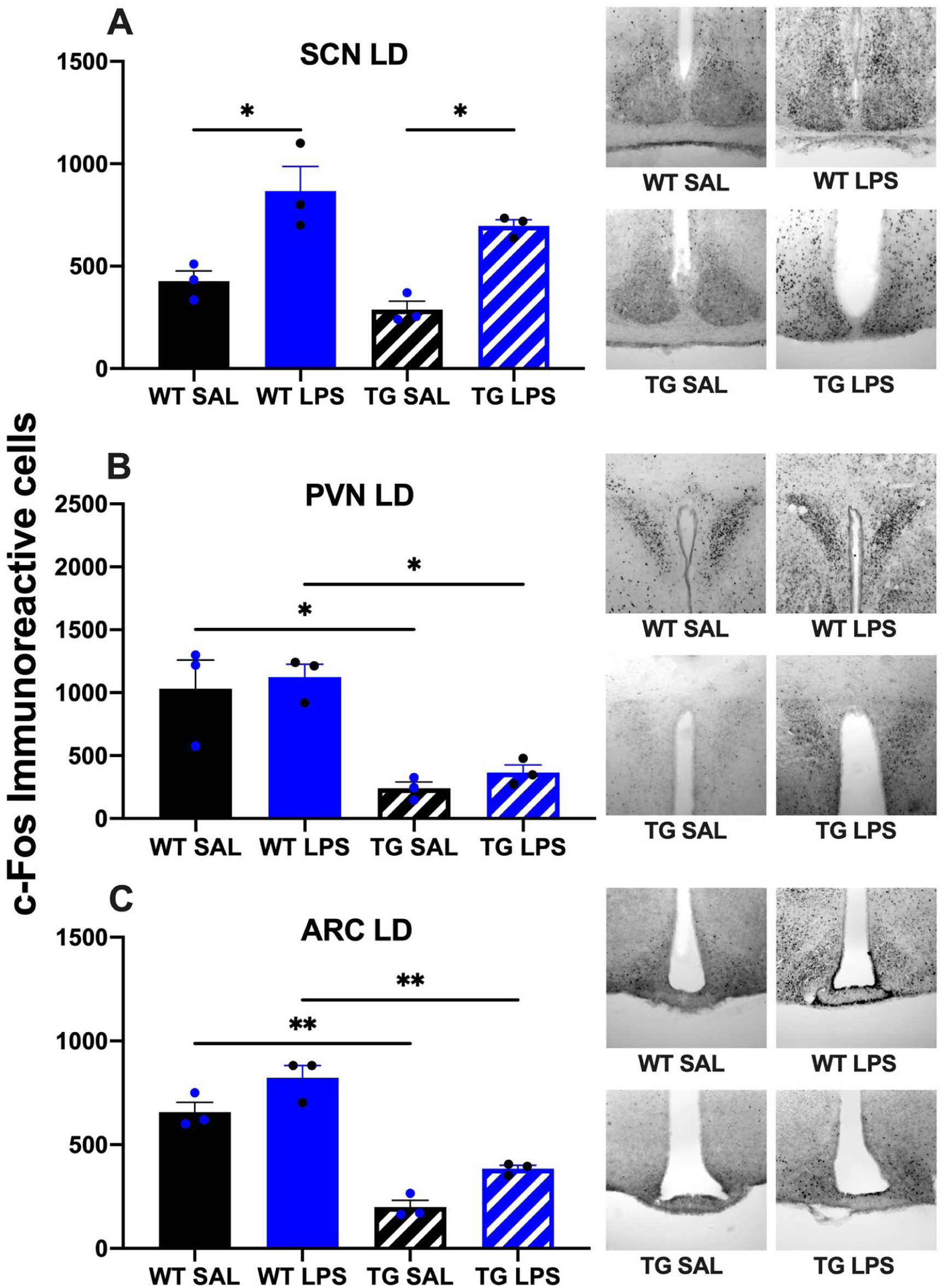
Neuronal activation (c-Fos immunoreactivity) in hypothalamic nuclei of WT and TG mice following LPS challenge under LD conditions. Quantification of c-Fos-positive neurons is shown for the SCN (a), PVN (b), and ARC (c) following i.p. injection of SAL or LPS in mice maintained under a 12:12 h LD cycle.
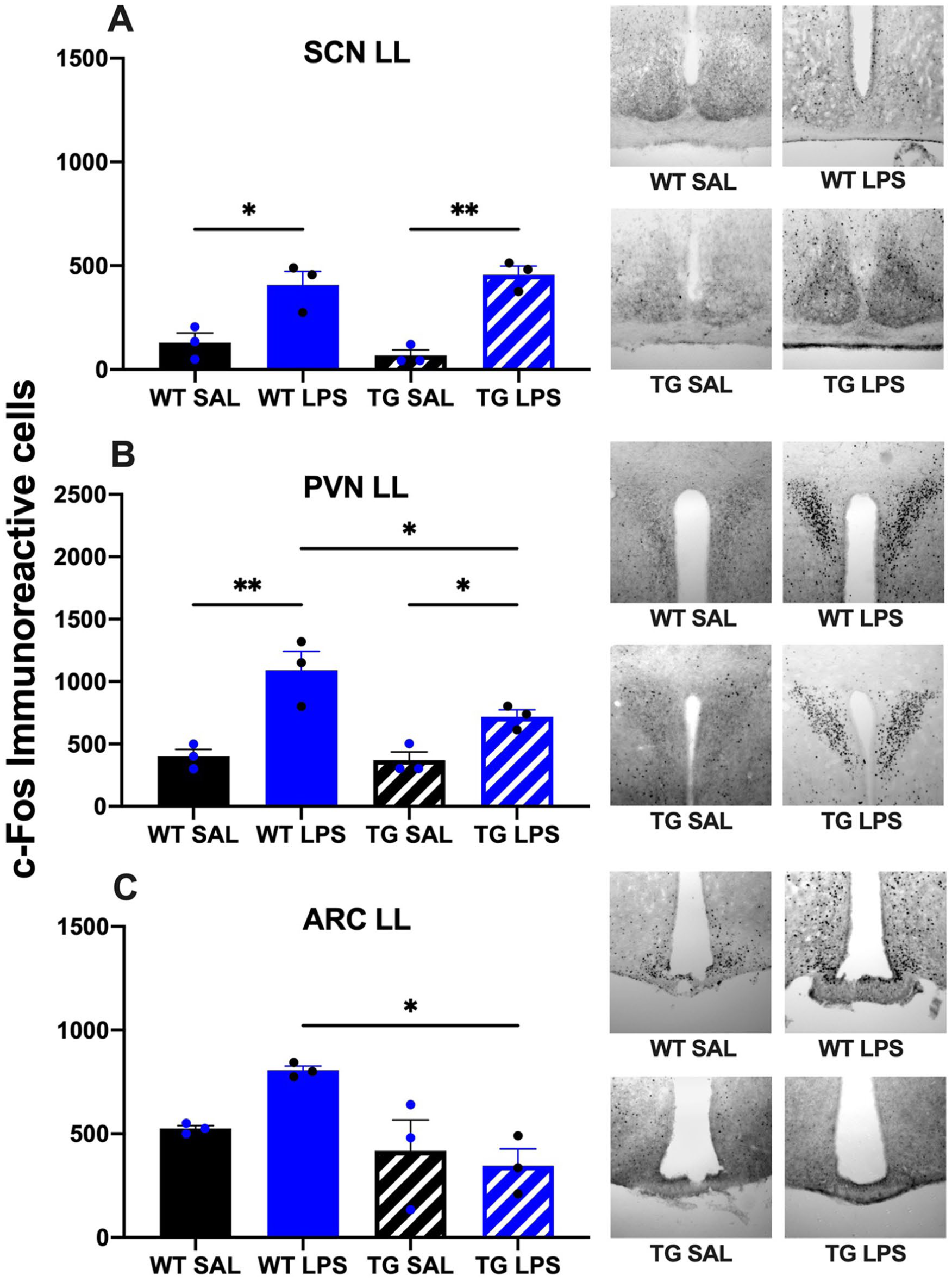
Neuronal activation (c-Fos immunoreactivity) in hypothalamic nuclei of WT and TG mice following LPS challenge under constant light (LL). Quantification of c-Fos-positive neurons is shown for the SCN (a), PVN (b), and ARC (c) following i.p. injection of SAL or LPS in mice maintained under LL conditions.
The regulation of c-Fos expression in the PVN differed strikingly between genotypes and lighting conditions. In WT animals, LL tended to decrease basal c-Fos expression compared to a LD cycle. Furthermore, WT mice in LD were unresponsive to LPS, whereas those in LL exhibited a significant, LPS-driven increase in c-Fos. In stark contrast, TG animals displayed significantly elevated c-Fos expression under LL conditions compared to LD. This elevated baseline in the TG-LL group was further amplified by LPS administration (Figures 5b and 6b).
In both lighting conditions, the ARC displayed a marked increase in the number of c-Fos-positive cells in the WT LPS-treated group compared to the WT saline-treated controls. Notably, the increase is more clearly observed in wild-type animals maintained under constant light conditions. In TG animals, no significant differences were observed between treatment groups in mice maintained under LL conditions; both saline- and LPS-treated animals exhibited comparable levels of c-Fos expression in this region. However, TG mice under LD conditions showed a clear LPS-driven increase in c-Fos expression (Figures 5c and 6c).
Discussion
This research delves into the complex interplay between circadian rhythms and neuroimmune responses, focusing specifically on the role of astrocytic interleukin-6 (IL-6) trans-signaling in the CNS. Our findings reveal that this pathway is not merely a cytokine relay but a critical “gating factor” for hypothalamic plasticity and systemic homeostasis under environmental stress. The GFAP-sgp130Fc transgene operates through the astrocytic synthesis and secretion of a soluble gp130-Fc fusion protein into the interstitial space. This protein binds exclusively to the circulating or local complexes formed by IL-6 and soluble IL-6 receptor (sIL-6R), acting as a highly specific decoy inhibitor of IL-6 trans-signaling. Crucially, classical signaling via the membrane-bound receptor (mIL-6R) remains entirely functional. Thus, given that the IL-6/sIL-6R complex plays a well-established role in modulating astrocytic differentiation and regional expression profiles within the brainn matrix (Marz et al., 1999), the observed changes in circadian behavioral resilience and blunted neuroimmune integration reflect an extracellular blockade of trans-signaling that intercepts neuroinflammation before it alters target cellular networks throughout the brain. In WT animals, observations confirm a robust and synchronized circadian rhythm under a 12:12 h light/dark (LD) cycle, characterized by typical nocturnal activity. This rhythmicity is, as expected, abolished under constant light (LL) conditions, leading to an aperiodic distribution of activity and a state of light-induced circadian chaos. Remarkably, transgenic (TG) animals (GFAP-sgp130Fc) displayed a compromised circadian pattern even under the entraining LD cycle. Although they maintained largely nocturnal activity (81% of total), the significant reduction in locomotor activity late in the dark phase (ZT20) suggests that intact astrocytic IL-6 signaling is fundamental for the clock’s sensitivity and plasticity to environmental cues (Figure 1d).
The role of astrocytes in the central regulation of circadian rhythms has advanced dramatically through genetic models with distinct regional and molecular targets. Initially, Prolo et al. (2005) utilized real-time bioluminescence imaging to reveal that isolated, cultured cortical astrocytes possess autonomous molecular clocks capable of sustaining robust circadian oscillations of Period1 expression independently of neuronal inputs. This established that astrocytic rhythmicity is an intrinsic property. Subsequently, to understand the systemic impact of these glial clocks, Barca-Mayo et al. (2017) generated a global conditional knockout by deleting Bmal1 across all astrocytes throughout the brain using a Gfap-Cre driver. These authors found that disrupting the astrocytic clock led to a disrupted global circadian architecture, an altered free-running period in constant darkness, and cognitive vulnerabilities, confirming that astrocytes actively gate behavioral output and synaptic plasticity.
However, these global glia-directed manipulations could not isolate whether the behavioral changes were driven specifically by the master pacemaker or by a distributed network. To answer this, Tso et al. (2017) utilized a highly restricted temporal Aldh1l1-CreERT2 model to delete Bmal1 or Clock exclusively within the cell populations of the SCN. They demonstrated that SCN astrocytic clocks alone are sufficient to lengthen the free-running period of locomotor activity, establishing a direct hierarchical control over the master pacemaker. Mechanistically, Brancaccio et al. (2017) further elucidated that SCN astrocytes actively dictate circadian behavioral pace by driving an anti-phase intracellular calcium (Ca2+) oscillation and controlling extracellular glutamate clearance through GLAST/GLT-1 transporters, thereby altering the excitability of SCN neurons.
In contrast to these pivotal studies that directly manipulated cell-autonomous core clock genes (Bmal1, Clock) or intracellular signaling (Ca2+) inside astrocytes, our study targeted an extracellular neuroinflammatory cascade via the global expression of the GFAP-sgp130Fc decoy receptor. Because the GFAP promoter drives transgene secretion pan-neuronally across multiple brain regions, our studies do not manipulate or test the role of astrocytes within any specific, isolated brain region. Therefore, it is inappropriate to assume that the SCN was exclusively or directly responsible for the altered behavioral or thermoregulatory phenotypes seen in our data.
The fascinating behavioral resilience of TG mice under constant light (LL)—where they maintain a distinct free-running rhythm instead of entering the state of circadian arhythmicity observed in WT mice—must be interpreted cautiously. Rather than an intra-SCN clock alteration, this phenotype likely reflects a broader disruption in neuroimmune-neuronal coupling across a distributed homeostatic network. This includes the anterior hypothalamic area orchestrating core temperature responses, the PVN involved in neuroendocrine output, and the SCN. In WT mice, constant light promotes an integrated, pro-inflammatory glial tone that drives internal desynchronization and “circadian chaos.” By intercepting IL-6 trans-signaling globally within the CNS, the TG brain becomes functionally disconnected or “deaf” to these disruptive extrinsic signaling cascades. This insulation prevents the master pacemaker and its systemic circuits from processing photic and inflammatory stress, explaining the rigid preservation of behavioral rhythms under constant light at the cost of failing to clear or respond to peripheral challenges.
The distinct thermoregulatory responses to LPS provided crucial evidence for a functional neuroimmune-circadian axis that requires intact astrocytic signaling for proper execution. LPS typically induces inflammatory cytokines, including IL-6, in both astrocytes and microglia, although species differences exist, with human astrocytes requiring IL-1β for IL-6 induction (Sawada et al., 1992; Lee et al., 1993). Under standard LD conditions, WT animals exhibited the canonical biphasic temperature response to LPS—initial hyperthermia followed by secondary hypothermia. This complex physiological reaction is orchestrated by the CNS, where IL-6 acts as a principal endogenous pyrogen signaling to thermoregulatory neurons in the anterior hypothalamic area, likely via prostaglandin pathways (Szelényi, 2001; Erta et al., 2012). In contrast, the blunted, monophasic fever and absent hypothermic phase observed in TG mice indicate that while the initial pyrogenic trigger remains partially active, the astrocytic IL-6 trans-signaling pathway is essential for orchestrating the full kinetic spectrum of the febrile response (Rose-John et al., 2023).
The interaction between genotype and photoperiod became most apparent under constant light (LL). While circadian desynchrony alone attenuated the WT response, TG mice exposed to LL were completely unresponsive to LPS, aligning with the known suppressive effect of constant light on SCN activity (Tapia-Osorio et al., 2013). This “thermal deafness” suggests that the combined stress of circadian disruption and impaired IL-6 trans-signaling induces a state of “homeostatic paralysis.” Our c-Fos analysis further elucidates this central failure. The markedly lower immunoreactivity in the PVN and ARC of TG mice—regardless of LPS treatment—reveals a profound inability of the hypothalamus to integrate systemic neuroimmune cues (Guerrero-Vargas et al., 2014). We acknowledge that the preoptic area (POA) is the canonical site for fever initiation via prostaglandin E2 (PGE2) signaling. However, while the POA may issue the primary “fever command,” the PVN and ARC serve as critical nodes for the autonomic, metabolic, and neuroendocrine translation of that signal. The complete absence of a febrile response and suppressed c-Fos activation across these regions in the TG-LL group suggests a broader failure in the relay and execution of the thermoregulatory response rather than an isolated POA defect.
Given that the ARC and PVN are essential for coordinating systemic output, their suppressed activity suggests that astrocytic IL-6 trans-signaling acts as a molecular gateway that allows the wider hypothalamus to “sense” and respond to peripheral inflammation. In this model, astrocytes within the SCN-PVN-ARC axis likely function as vital relays; without their contribution, the thermostatic set-point shift initiated in the POA may fail to engage the downstream effector pathways. While our findings highlight this systemic integration failure, future studies utilizing PGE2 microinjections directly into the POA will be necessary to further isolate the “sensing” versus “execution” components of this defect.
Importantly, the total abolition of fever and the severe suppression of PVN c-Fos in TG mice—particularly under the stress of LL—cannot be explained solely by lower peripheral cytokine titers. Instead, our data suggest that astrocytic IL-6 trans-signaling acts as a necessary gate for the CNS to integrate systemic inflammatory cues. Without this central “sensing” mechanism, the hypothalamus fails to coordinate a systemic response, regardless of the cytokine levels present in the periphery.
Furthermore, the peripheral inflammatory gene expression in the liver provides a mirror to this central dysfunction. The blunted profile of IL-6 and TNF-α in TG mice directly reflects the targeted inhibition by the sgp130Fc construct, which appears to truncate the inflammatory cascade without preventing upstream mediators like IL-1β. However, the most striking finding was the pervasive suppression of nearly all inflammatory markers in both genotypes under LL conditions. This supports the hypothesis that constant light severely attenuates the inflammatory response, likely by desynchronizing the circadian clocks that gate immune signaling pathways. While the sample size utilized for these molecular quantifications (n = 3 per group) represents a limitation of the current study, the high degree of phenotypic consistency observed—coupled with the robust effect sizes and the alignment with our continuous telemetric data—supports the validity of these inflammatory profiles.
The elevated baseline of TNF-α in TG mice under LL, even in the absence of LPS, points toward a pre-existing state of dysregulation. This aligns with the concept that chronic circadian disruption leads to uncoordinated and potentially exacerbated inflammatory states (Castanon-Cervantes et al., 2010), but our data suggests that without the astrocytic IL-6 “bridge,” this inflammation remains sequestered or fails to trigger the appropriate central homeostatic corrections. While the GFAP-sgp130Fc model is well established, its application in the current study reveals a previously unknown role for astrocytic IL-6 in regulating the SCN’s vulnerability to light-induced desynchrony. We acknowledge the potential peripheral distribution of the secreted sgp130Fc protein; consequently, future studies should aim to quantify its levels in both the CNS and peripheral compartments to further refine the relative neuroimmune versus systemic contributions.
In summary, our results highlight a critical vulnerability: the neuroimmune system requires both a synchronized circadian background and functional astrocytic IL-6 trans-signaling to mount a coordinated defense. The loss of either component impairs the response, but the loss of both leads to a complete failure of the host to react to an immunological challenge, consolidating astrocytic IL-6 as a master integrator of environmental and inflammatory inputs.
A limitation of the current study is that direct biochemical quantification of central sgp130Fc protein expression or downstream STAT3 phosphorylation (p-STAT3) was not performed on the brain tissue of this specific cohort. However, the functional impairment of IL-6 trans-signaling in this specific transgenic line has been comprehensively validated previously, demonstrating a complete blunting of trans-signaling-dependent cellular signaling within the brain parenchymal matrix without disrupting classical IL-6 pathways (Garbers et al., 2013; Campbell et al., 2014). Furthermore, our physiological data provide strong indirect proof of functional impairment: TG mice displayed a significantly blunted and modified kinetic profile in their thermoregulatory response to systemic LPS (Figure 2) and a truncated downstream expression profile of inflammatory markers in the liver and serum (Figures 3 and 4), consistent with a compromised neuroimmune signaling cascade.
Conclusion
These findings collectively highlight the critical and context-dependent role of IL-6 trans-signaling in mediating neuroimmune communication, particularly within the framework of chronobiology. The differential responses observed between WT and TG mice under synchronized (LD) and desynchronized (LL) conditions underscore a complex interplay between the immune and nervous systems, demonstrating that their communication is profoundly governed by circadian organization.
The pervasive impairment of immune and behavioral responses in TG mice—most notably under circadian desynchrony—is evidenced by a striking unresponsiveness to LPS in thermoregulation and severe suppression of hypothalamic c-Fos activation. These results strongly suggest that astrocytic IL-6 trans-signaling is a mechanical necessity for maintaining immunological homeostasis and enabling an effective response to systemic challenges. Specifically, the blunted IL-6 and IFN-γ cytokine profiles, coupled with dysregulated TLR4 expression in TG mice, indicate a broader disruption of innate immune pathways when this specific signaling route is inhibited. In summary, this body of evidence identifies astrocytic IL-6 trans-signaling as a master molecular integrator that bridges systemic inflammatory cues with the central circadian clock. By characterizing this axis, our results pave the way for novel therapeutic strategies targeting the IL-6 trans-signaling pathway to treat inflammatory and chronobiological disorders where neuroimmune integration is fundamentally compromised.
Supplemental Material
sj-docx-1-jbr-10.1177_07487304261459132 – Supplemental material for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice
Supplemental material, sj-docx-1-jbr-10.1177_07487304261459132 for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice by Josué S. Ambríz-Zárate, Adrián Báez-Ruiz, Nadia Saderi and Roberto C. Salgado-Delgado in Journal of Biological Rhythms
Supplemental Material
sj-jpg-1-jbr-10.1177_07487304261459132 – Supplemental material for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice
Supplemental material, sj-jpg-1-jbr-10.1177_07487304261459132 for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice by Josué S. Ambríz-Zárate, Adrián Báez-Ruiz, Nadia Saderi and Roberto C. Salgado-Delgado in Journal of Biological Rhythms
Supplemental Material
sj-jpg-2-jbr-10.1177_07487304261459132 – Supplemental material for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice
Supplemental material, sj-jpg-2-jbr-10.1177_07487304261459132 for IL-6 Trans-Signaling Is Critical for Integrating Circadian Rhythms and Neuroimmune Responses to LPS Challenge in Mice by Josué S. Ambríz-Zárate, Adrián Báez-Ruiz, Nadia Saderi and Roberto C. Salgado-Delgado in Journal of Biological Rhythms
Footnotes
Acknowledgements
We thank Dr. Marco Atzori for the donation of the TG animals.
Author Contributions
R.C.S.-D. directed the study. J.A.-Z., A.B.-R., and N.S. developed the methodology, conducted the acquisition, and performed the data analysis. R.C.S.-D., N.S., and J.A.-Z. contributed to interpretation of data, writing, and manuscript review. All authors contributed to the discussion of the data and editing of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Secretaría de Ciencia, Humanidades, Tecnología e Innovación, México (SECIHTI, grant CBF-2025-I-3783) for R.C.S.-D. (COPOCYT, FIDEICOMISO 23871; FME/2024/SE-04/27).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Supplemental Material
Supplementary material is available for this article online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.