
Abstract
The use of genetically introduced optical reporters has improved understanding of circadian rhythms at the cellular and systems levels, but the use of this approach in mammals has been limited to mice and rats. The Syrian hamster (Mesocricetus auratus) has been a long-standing circadian model due to its rhythmic precision and physiological responses to photoperiod. Here we report the development of the Per2LUC hamster, which harbors a knock-in construct inserted using the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) technology. We test circadian and photoperiodic function at the behavioral level in the Per2LUC hamster and evaluate ex vivo Per2-driven bioluminescence rhythms across a variety of tissues. Our results indicate modest effects of transgene insertion on circadian function in vivo and provide evidence that ex vivo Per2-driven bioluminescence rhythms are robust to dissection for many of the 10 tissues examined here. Properties of bioluminescence rhythms varied by tissue type in the Per2LUC hamster, which was overall similar to those displayed by tissues collected from the Per2LUC mouse. Collectively, these results indicate that this new animal model may be beneficial in both circadian and non-circadian applications in future studies.
Introduction
Circadian rhythms support homeostasis by aligning physiology with predictable changes in the 24 h environment (Pittendrigh, 1960). At the cellular level, the transcription and translation of clock genes provide a molecular mechanism for circadian timekeeping (Campbell-Galland et al., 2025). In mammals, CLOCK and BMAL1 activate Period (Per1 and Per2) and Cryptochrome (Cry1 and Cry2) transcription. Once translated, PER and CRY proteins form complexes that inhibit their own transcription. Many genetic networks are regulated by this core molecular clock, with physiological consequences that vary by cell and tissue type (Koike et al., 2012; Noya et al., 2019; Zhang et al., 2014). The circadian system is hierarchical; clock tissues are entrained and adjusted to the solar cycle by the suprachiasmatic nucleus (SCN) in the anterior hypothalamus (Mohawk et al., 2012). The SCN is a network of interconnected neurons and glia that regulate circadian timekeeping in tissue clocks throughout the body (Smyllie et al., 2025a). Understanding circadian control mechanisms across the molecular to systems level remains critical given the widespread influence of daily rhythms on health and disease (Evans and Davidson, 2013; Shen et al., 2023).
Optical reporters that monitor clock gene expression in real time have advanced insight into circadian control mechanisms in many organisms (c.f., Smith et al., 2022). In mouse and rat reporter models, the use of this approach has demonstrated that molecular rhythms are generated by many different cell types (Balsalobre et al., 2000; Yamazaki et al., 2000; Yoo et al., 2004). In addition, these findings highlight that the molecular clock function varies across tissues and cell types (Schibler et al., 2015). For instance, many peripheral clocks damp over time ex vivo, but the SCN and several other tissues display long-lasting, self-sustained molecular rhythms (Abe et al., 2002; Leise et al., 2012; Yamaguchi et al., 2003; Yoo et al., 2004). Real-time imaging approaches have also been used to understand population-level dynamics in the SCN (reviewed in Evans and Gorman, 2016; Evans and Schwartz, 2024). The conservation of findings across circadian reporter models is notable given differences in genetic design, signal dynamics, and modulation by native control mechanisms. For example, in mPer1-luciferase reporter models (mPer1-luc), the transgene is driven by a Per1 promoter fragment (e.g., 3-7.2 kb across models) inserted into non-native location(s) in the genome (e.g., 1-12 copies across models), which produces a transcriptional reporter not modulated by the native genetic and epigenetic landscape (Kuhlman et al., 2000; Wilsbacher et al., 2002; Yamaguchi et al., 2000; Yamazaki et al., 2000). In contrast, the Per2LUC knock-in mouse harbors a transgene inserted into the native Per2 locus (Yoo et al., 2004), with signal dynamics dictated by post-translational modulation of the PER2::LUC fusion protein. Although this technique has limitations (e.g., dissection-induced resetting [Davidson et al., 2009; Yoshikawa et al., 2005]; interference with molecular clock function [Ralph et al., 2021]), the use of bioluminescent reporter models of molecular clock function has advanced our understanding of cellular and systems-level clocks in both mouse and rat models.
Here we report the development of a bioluminescent reporter model for the study of circadian rhythms in the Syrian hamster. This species has been widely used due to its circadian precision (Pittendrigh and Daan, 1976a), circadian control of the estrous cycle (Fitzgerald and Zucker, 1976; Russo et al., 2015), and circadian control of photoperiodic responses (Pittendrigh and Daan, 1976b; Pittendrigh et al., 1984). Per2LUC hamsters were generated using the CRISPR technology with a knock-in strategy incorporating the T2A sequence, which is designed to produce independent translation of the resulting protein products. Per2LUC hamsters displayed typical behavioral responses across a battery of circadian assays (e.g., photoentrainment, constant darkness [DD], simulated jetlag, short photoperiods), suggesting the transgene does not interfere with the clock function. Furthermore, many tissues from the Per2LUC hamster exhibited bioluminescence rhythms that persist for at least 3 cycles ex vivo. By varying the time of dissection, we identified tissues for which the phase of ex vivo rhythms was affected by dissection and/or cell culture. Using these data, we selected tissues to define system-level timekeeping in the Per2LUC hamster and compared these results to the Per2LUC mouse (Yoo et al., 2004). Furthermore, we performed real-time imaging for Per2LUC hamster SCN slices, which reveals species similarities and differences in central clock organization when compared to previous work conducted in the Per2LUC mouse. Collectively, these results provide detailed insights into the circadian organization of the Syrian hamster and support the use of this model for understanding circadian function in this species.
Materials and Methods
Animals and Husbandry Conditions
Hamsters were bred and raised under a 24 h light:dark cycle with 14 h of light and 10 h of darkness (LD14:10, lights on defined as zeitgeber time 1 [ZT1]). Syrian hamsters used for generating Per2LUC hamsters were produced from a wildtype breeding colony derived from Charles River stock (LVG Golden Syrian Hamster, Strain Code: 049). After generation, Per2LUC hamsters were competent breeders, with estrous cycles and aggression starting in females as young as 2-3 months. For breeding, Per2LUC female hamsters were staged to determine estrous state using vaginal smears and/or lordosis testing, then left with males for a short period of time (2-8 h) prior to female separation for gestation and lactation. The Per2LUC mice used here were originally derived from the stock produced by Yoo et al. (2004), bred from monogamous pairs, and raised in a separate colony room programmed to LD12:12 (lights off: 1800 CST). At weaning, animals of both species were group-housed in same-sex cages without running wheels. Throughout life, ambient temperature in all animal rooms was maintained at 22 ± 2 °C, with ad libitum access to water and food. All procedures were conducted according to the NIH Guide for the Care and Use of Animals and approved by the Institutional Animal Care and Use Committee at each respective site (Utah State University IACUC protocol 2091, 13758; Marquette University IACUC protocol 3924; University Massachusetts, Amherst IACUC protocol 3537).
sgRNA Selection and Donor Plasmid Construction
Given the potential for single nucleotide polymorphisms between the published sequence of the hamster Per2 locus (Gene ID: 101834497) and the sequences from hamsters utilized here, isogenic genomic DNA approximately 2 kb upstream and 2 kb downstream of the stop codon of Per2 was polymerase chain reaction (PCR) amplified from genomic DNA isolated from hamster breeders, subcloned into TA cloning vectors and sequenced. Using Benchling software (San Francisco, CA), 3 sgRNAs were designed (Figure 1a, sgRNA1: 5’-AGATTGCTGGCTGGGGGACA-3’; sgRNA2: 5’-TAGATTGCTGGCTGGGGGAC-3’; sgRNA3: 5’-TAATGTAGATTGCTGGCTGG-3’), synthesized by Integrated DNA Technologies (IDT; Coralville, IA), and tested individually for on-target activity. Ribonucleoprotein complexes (RNPs) comprising sgRNAs and Cas9 protein (sgRNAs/Cas9) were assembled by mixing equal concentrations of sgRNAs and Cas9 protein to a final concentration of 1 μg/μL each in TE buffer (10 mM Tris, 0.1 mM ethylenediaminetetraacetic acid [EDTA] solution, pH 7.0). BHK-21 cells were nucleofected with sgRNAs/Cas9 RNP (3 μg/3 μg) using a P3 Primary Cell 4D-Nucleofector X kit (Lonza, Basel, Switzerland). Cells were harvested 72 h after nucleofection for gDNA extraction. Whether sgRNAs introduced modifications (indels) into the hamster genome was assessed by PCR restriction fragment length polymorphism (PCR-RFLP). PCR was performed with Ex Taq (Takara, San Jose, CA): initial denaturation (94 °C, 5 min), followed by 32 cycles of 30-sec denaturation (94 °C), 30-sec annealing (58 °C), and 30-sec extension (72 °C), with a final extension step for 5 min (72 °C). Primers for amplifying the region targeted by the sgRNA were Per 2F: 5’-TCCCTTGGGACTCGCTGATA-3’ and R: 5’-ACGACAAATCTCACCCCGTC-3’. The restriction enzyme BseYI (New England Biolabs, NEB, Ipswich, MA) was used for PCR-RFLP. The donor plasmid pKO2.1-Per2-Luc (Figure 1a) used for pronuclear injection was constructed by using a NEBuilder HiFi DNA Assembly Cloning Kit (NEB, Ipswich, MA). Briefly, left homologous arm (~1 kb), T2A (5’-GAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGCCCA-3’, as a synthetic DNA oligo), the firefly luciferase gene (from pGL4.20 [luc2], Promega #E6751), and right homologous arm (~1 kb) were PCR amplified with a 25-bp overlap on each side and assembled with plasmid pKO2.1 (Addgene plasmid #22674) linearized with MscI. The resultant plasmid was confirmed by restriction enzyme digestion mapping and Sanger sequencing (Figure S1).
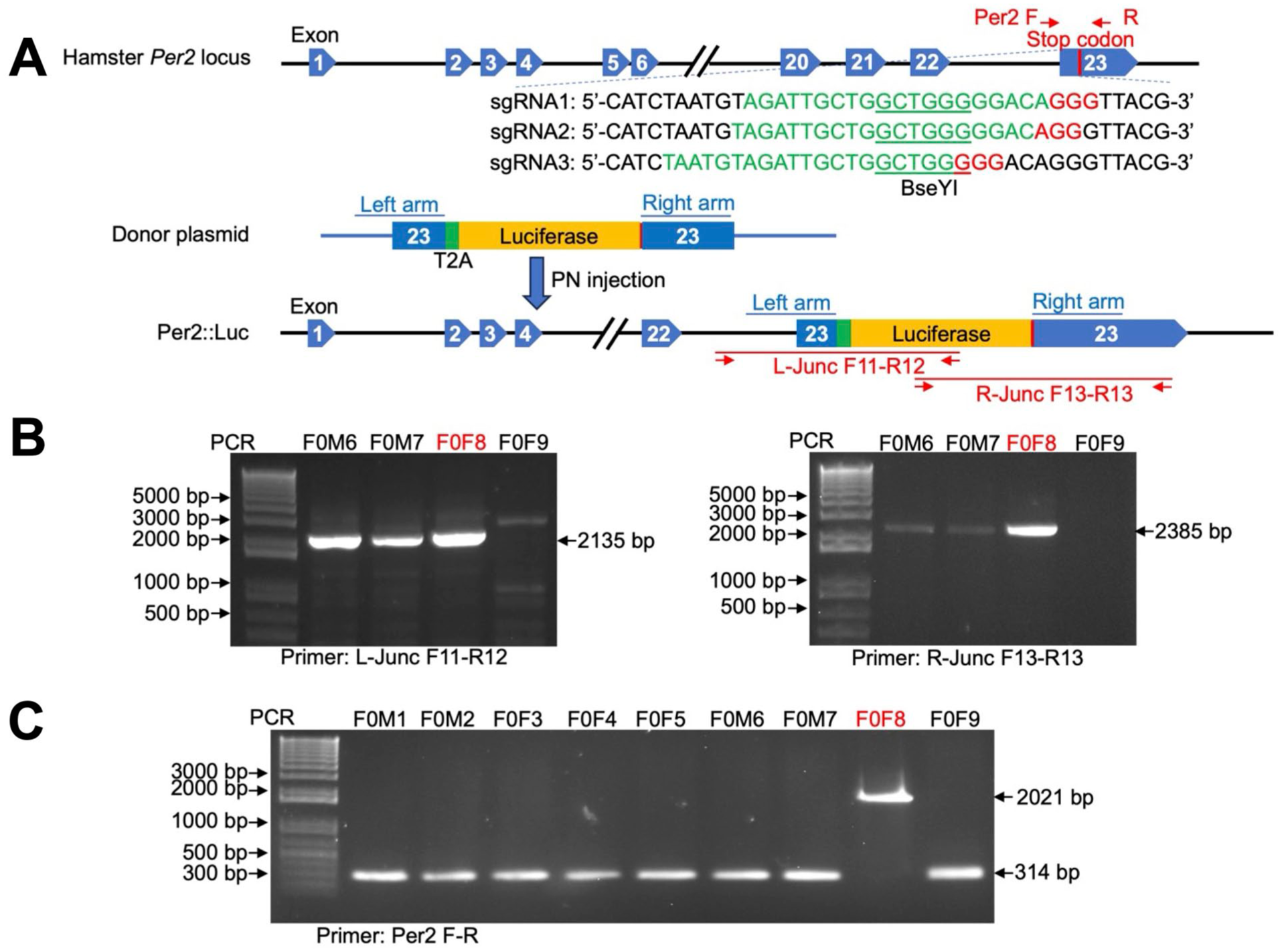
Generation of Per2LUC Syrian hamsters using the CRISPR/Cas9-mediated gene knock-in strategy. (a) Schematic diagram illustrating the Syrian hamster Per2 gene (Gene ID: 101834497), the sgRNA sequences (in green, with the Protospacer Adjacent Motif, PAM, highlighted in red), and PCR primers (Per2F-R) and the restriction enzyme (BseYI, green underlined) used for detecting DNA cleavage efficiency. Also depicted are the donor plasmid carrying the T2A-Luciferase cassette, the Per2-Luc allele after the intended T2A-luciferase cassette integration immediately before the stop codon of Per2, and the junction PCR primers, F11-R12 and F13-R13, that were used to detect the genomic junctions generated by transgene cassette integration. Diagrams are not to scale. See also Figure S1. (b) Genotyping results of F0 hamsters by junction PCR to detect the Per2LUC allele. (c) Genotyping results of F0 hamsters using Per2F-R to distinguish Per2LUC/+ and Per2LUC/LUC knock-in alleles.
Generation of Per2LUC Hamsters
As in previous work (Fan et al., 2014), female Syrian hamsters were superovulated by an intraperitoneal injection of 10-25 IU of pregnant mare serum gonadotropin (BioVendor, Ashville, NC) based on body weight on the morning of Day 1 of the estrous cycle (ZT1-4). These females were mated to fertile males on Day 4 of the estrous cycle (ZT13) and were euthanized approximately 18 h after mating for zygote isolation. Pronuclear injections were performed in a dark room, and red filters were used for all microscope light sources. Zygotes were flushed from oviducts with warmed and equilibrated HECM-9 medium supplemented with 0.5 mg/ml human serum albumin, then washed twice with HECM-9, transferred into 20-µL drops of HECM-9 covered by mineral oil, and cultured at 37.5 ℃ under 10% CO2, 5% O2, and 85% N2 for pronuclear injection. The sgRNA/Cas9 ribonucleoprotein and donor plasmid were diluted to 60 ng/µL:60 ng/µL using Tris/EDTA buffer (pH 7.0). For pronuclear injections, a group of 15-20 hamster zygotes were transferred to a 200-µL HECM-9 drop in the microinjection dish and covered with mineral oil. The ribonucleoprotein/donor was then injected into the male pronucleus. After injection, embryos were washed twice with equilibrated HECM-9 and cultured at 37.5 ℃ under 10% CO2, 5% O2, and 85% N2. Embryos with normal morphology were transferred into the oviducts of pseudopregnant recipients that had been mated with vasectomized males on the prior day. Embryos were transferred bilaterally with 10-15 embryos per oviduct. Pups were genotyped at 2 weeks of age, with genomic DNA isolated from toe-clipping using a Qiagen Blood and Tissue Kit (Qiagen; Hilden, Germany). PCR was performed to screen T2A-luciferase cassette insertion with Ex Taq and the following parameters: initial denaturation at 94 °C for 1 min, followed by 32 cycles of 10-sec denaturation at 98 °C, 20-sec annealing at 56 °C, and 2-min extension at 68 °C, with a final extension step of 72 °C for 10 min. Hamsters were genotyped to identify animals carrying Per2LUC by genomic PCR using primers specific for the new genomic junctions formed by the T2A-luciferase cassette insertion. Primers for amplifying the left junction are F11: 5’-CAGCGACTGTGTCCTGTTAC-3’ and R12: 5’-AAGTCCACCACCTTAGCCTC-3’. Primers for the right junction are F13: 5’-ACGTGCAAAAGAAGCTACCG-3’ and R13: 5’-GAGGTTCCCATCTCCCCAAA-3’ (Figure 1a and b). In addition, PCR with primers, Per2F and R, which were used in the DNA cleavage efficiency assays mentioned earlier, was also performed to further confirm genotyping results (Figure 1a and c). PCR was performed with Ex Taq using the following parameters: initial denaturation at 94 °C for 1 min followed by 32 cycles of 10-sec denaturation at 98 °C, 20-sec annealing at 56 °C, and 2 min extension at 68 °C, with a final extension step of 72 °C for 10 min.
Behavioral Analyses
Adult Per2LUC and wildtype hamsters of both sexes were singly housed in cages equipped with running wheels. Locomotor rhythms were visualized and analyzed with Clocklab software (Actimetrics, Evanston, IL). To test effects of the Per2LUC transgene on photoentrainment, hamsters were entrained to LD14:10 for at least 10 days (Per2LUC/ LUC n = 11, wildtype n = 17). To evaluate effects on the free-running period, hamsters were released into DD for 9-12 days (Per2LUC LUC n = 20, wildtype n = 14). Free-running period was measured with linear regression fit to activity onsets. To determine effects on photic re-entrainment, hamsters were exposed to an 8 h shift of the LD14:10 cycle in each direction (i.e., advance, then delay). Only hamsters with uninterrupted data collection were used in analyses for the advance (Per2LUC/LUC n = 18, wildtype n = 6) and delay directions (Per2LUC/LUC n = 15, wildtype n = 9). Rate of re-entrainment was quantified by visually determining the number of days required to realign the onset of locomotor activity to the new time of lights off in each hamster for each shift direction. Last, Per2LUC hamsters were transferred from LD14:10 to LD6:18 for 8-12 weeks to track photoperiodic changes in behavior and physiology. Estrous cycles were monitored by daily vaginal smears taken at ZT0-2 under both LD14:10 and LD6:18, as in Kent et al. (1968). Reproductive tissue weight was collected after 9 weeks of LD6:18 for each sex (Per2LUC/LUC female n = 4, Per2LUC/LUC male n = 6).
Bioluminescent Tissue Collection
Per2LUC hamsters were deeply anesthetized with pentobarbital (80 mg/kg, University of Massachusetts) or isoflurane anesthesia (Marquette), then euthanized by decapitation. Dissections were performed near dawn (ZT0-4), midday (ZT5-8), afternoon (ZT9-12), or night (ZT13-20, n = 5-16/tissue/ZT, with only 1 sample/tissue collected from each animal). Tissues collected were the SCN, pineal (PIN), pituitary (PIT), adrenal (ADR), lung, spleen (SPLN), white peri-adrenal adipose (ADI), liver, ovary (OVR), and testes. As in previous work (Evans et al., 2015), the brain and other tissues were removed and placed in chilled Hank’s Balanced Salt Solution supplemented with HEPES, NaHCO3, and penicillin-streptomycin. SCN slices were collected in the coronal plane with a motorized vibratome (Leica Biosystems, VT1200S), and other tissue samples were collected manually with a scalpel. After collection, samples were cultured on a Millicell membrane (Millipore, Cat# PICM01250) with 1.2 ml of serum-free, air-buffered Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco 12100-046) supplemented with HEPES, NaHCO3, penicillin-streptomycin, and D-luciferin (<0.2 mM, potassium salt, Gold Biotechnologies). For a subset of hamster tissues, dexamethasone (100 nM, Sigma, D1756) or forskolin (50 uM, Sigma, F6886) was applied for 1 h after a minimum of 7 days of recording to test if Per2-driven bioluminescence rhythms could be restored after damping, as in the works by Balsalobre et al. (2000) and Yamazaki et al. (2000). Tissues were collected at ZT6-10 from Per2LUC mice at Marquette with isoflurane and other procedures described earlier, then compared to hamster tissues collected at the same ZT using identical procedures.
Bioluminescent Recording
Per2-driven bioluminescence rhythms were measured at the whole dish level for at least 6 days using a luminometer (Hamamatsu LM2400, Hamamatsu City, Japan; or Actimetrics) housed inside a light-tight incubator set to 36 °C. For SCN imaging, slices were imaged using a Stanford Photonics XR Mega-10Z cooled intensified CCD camera on a Zeiss AxioObserver Z1 microscope with a 10x Fluar objective lens. Images were collected with Piper Software (Stanford Photonics, Palo Alto, CA) at 15 frames/sec, filtered in real time to eliminate single-image noise events (i.e., cosmic rays), and stored as 90-sec images collected once every 10 min. A 2-h moving average was then applied, images were converted to 8-bit, pixel dimensions were reduced in half, and 3 consecutive images were summed to produce a series of 30-min images. Time in vitro was normalized across samples by expressing the start time of recording relative to ZT (e.g., if a recording started 1 h after lights off, the ZT start would be ZT13).
Luminometry Analyses
Per2-driven bioluminescence rhythms were analyzed using the Lumicycle analysis software (Actimetrics). Each time series was detrended by subtracting the 24-h running average from the raw data before fitting a damped sine wave to the first 5 cycles in vitro, starting with the time of the first trough. Period, damping rate (the number of days required for the rhythm amplitude to decrease to 1/e of the initial value, ca. 37%), and the goodness of fit of the cosine wave (% variance accounted by the fitted model) were recorded for each sample. Tissue samples were deemed rhythmic when goodness of fit exceeded 65%, which was selected to detect less-precise oscillations observed across mouse strains (Kim et al., 2021). Finally, the time and value for peak and trough bioluminescence levels were recorded for each sample over each cycle in vitro, from which amplitude (i.e., difference between peak and trough values) and peak-to-peak period length (i.e., difference between 2 consecutive peak times) were quantified for each cycle in vitro. For each sample, we calculated the average period and the precision of period (i.e., 1/standard deviation of peak-to-peak period length) over the first 5 cycles in vitro.
Imaging Analyses
Per2-driven bioluminescence rhythms were extracted and analyzed from imaging data using Matlab-based scripts, as in the study by Evans et al. (2011). For phase maps, a time series was generated for each 12-pixel-diameter region-of-interest (ROI) on a uniform grid with 2-pixel spacing. The time series for each ROI was judged circadian if the autocorrelation coefficient with a lag of 24 h was significant at α = 0.05, a local maximum occurred in the autocorrelation corresponding to a lag between 18 h and 30 h, and the signal-to-noise ratio of the time series was at least 0.75. The linear trend was eliminated, and a Butterworth filter was applied once forward and once backward to remove high- and low-frequency interference. Phase was calculated as the number of hours between the start of the recording and the time of peak bioluminescence expression on the first cycle in vitro. Because time in vitro is corrected for the ZT start time of recording, this measure is referred to as “ZT Peak Time.” To examine phase relationships across the SCN network in average phase maps, regional ZT peak time was expressed relative to the peak time of the whole anterior slice on the first cycle in vitro and referred to as “Relative Peak Time.”
Statistical Analyses
Statistical analyses were performed with JMP software (SAS Institute, Cary, NC). Data are represented in figures and tables as mean ± standard error of mean. Where appropriate, full factorial analysis of variance (FF ANOVA) was performed with the following factors: genotype, sex, tissue, and/or time of day, along with the corresponding interactions. The criterion for statistical significance was set at α = 0.05.
Results
Generation of Per2LUC Hamsters
The Per2LUC construct was knocked into the endogenous Per2 locus using CRISPR/Cas9-mediated genetic engineering, as previously established for the Syrian hamster (Fan et al., 2014). The reporter gene luciferase (Luc) was inserted between the last codon and the stop codon of the Per2 gene with a viral T2A peptide linker between the coding sequences of Per2 and Luc to produce independent proteins from a single mRNA transcript (Figure 1a, Figure S1). Three sgRNAs were designed to target the stop codon region of the hamster Per2 locus (Figure 1a) and were 20 bp in length. After synthesis of sgRNA and assembly with Cas9 into the RNP complex, a PCR-RFLP assay was performed on transfected hamster BHK-21 cells to evaluate DNA cleavage (measured as indels). sgRNA1 exhibited the highest cleavage activity, resulting in 35% of indels in the transfected cells at its targeting site (data not shown). Pronuclear injection was then performed in fertilized hamster zygotes with sgRNA1/Cas9 RNP and the donor plasmid.
Pronuclear injected zygotes with normal morphology (363 total) were transferred into pseudopregnant female hamsters (14 total), resulting in 4 litters with 26 pups. Pups were genotyped by PCR using genomic DNA isolated from toe-clipping using primers to amplify the left genomic junction (primers F11-R12) and right genomic junction (primers F13-R13), created by integrating the T2A-luciferase cassette into the hamster Per2 locus (Figure 1a and b). Among the 26 pups screened, 3 pups (F0M6, F0M7, and F0F8) were positive for both left and right junctions. However, the right junctions in pups F0M6 and F0M7 were only weakly amplified (Figure 1b). Sanger sequencing of the junction PCR products indicated that the T2A-luciferase cassette was successfully integrated before the Per2 stop codon in all three pups, but indels were introduced into the right homologous arm regions in pups F0M6 and F0M7 (data not shown). These indels were most likely introduced by the non-homologous end joining (NHEJ) process, which are DNA mutations commonly observed with CRISPR-mediated gene knock-in (Merkle et al., 2015). Further genotyping demonstrated that F0F8 was homozygous for the T2A-luciferase cassette, while F0M6 and F0M7 were heterozygous (Figure 1c). Based on these results, we selected F0F8 to breed with a wildtype male, producing F1 Per2LUC/+ hamsters used for breeding to establish the Per2LUC/LUC colony. All the experiments described in this manuscript utilized Per2LUC hamsters produced from this line.
To further evaluate the inserted sequence, we conducted detailed sequence analysis on the Per2LUC genomic locus in the F0F8 progeny (n = 9). Genomic DNA was PCR amplified with primers F11 and R13 (Figure 1a) and used for Sanger sequencing (Supplemental Figure S1). Consistent with genotyping results using junction primers, DNA sequencing confirmed that the T2A-Luc cassette was integrated immediately before the Per2 stop codon as intended (Supplemental Figure S1). However, a C nucleotide insertion was identified 7 nucleotides downstream of the stop codon of the inserted Luc gene in each hamster (Supplemental Figure S1), which was likely introduced into the F0F8 founder DNA during CRISPR-mediated transgenic cassette integration.
Behavior and Physiology of Per2LUC/LUC Hamsters
Photic entrainment was examined in wildtype and Per2LUC/LUC Syrian hamsters of both sexes held under a standard photocycle (Figure 2a). Under LD14:10, wildtype and Per2LUC/LUC hamsters did not differ in the phase of locomotor onset relative to lights off or the phasing of locomotor offset relative to lights on (Figure 2b). Similarly, the duration of the active phase did not differ by genotype (Figure 2c), but overall wheel revolutions were higher in Per2LUC/LUC Syrian hamsters (data not shown, FF ANOVA omnibus: F(3,24) = 2.8, p = 0.06; GT: F(1,24) < 7.9, p < 0.01; sex: F(1,24) < 0.1, p = 0.8; GT*Sex: F(1,24) = 1.5, p = 0.2).
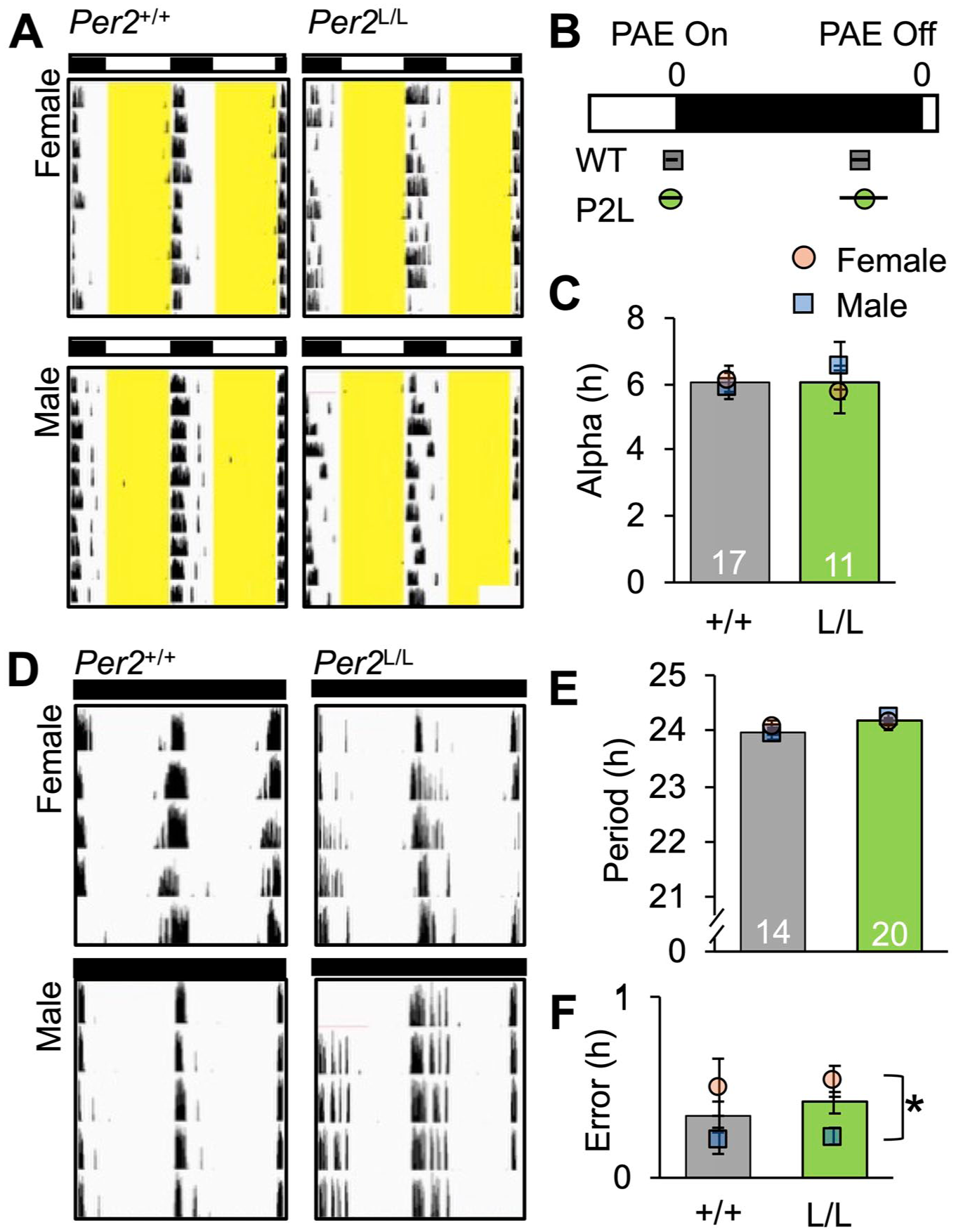
Photoentrained and free-running locomotor activity rhythms in Per2LUC/LUC (L/L) and Per2+/+ hamsters (+/+). (a) Representative, doubled-plotted wheel-running actograms illustrating entrained locomotor rhythms in both genotypes. (b) Phase angle of entrainment for activity onset and offset in each genotype. PAEon FF ANOVA omnibus: F(3,24) = 0.2, p = 0.8; GT: F(1,24) < 0.1, p = 0.8; sex: F(1,24) < 0.1, p = 0.8; GT*Sex: F(1,24) = 0.5, p = 0.5. PAEoff FF ANOVA omnibus: F(3,24) = 0.2, p = 0.8; GT: F(1,24) < 0.1, p = 0.9; sex: F(1,24) = 0.4, p = 0.5; GT*Sex: F(1,24) = 0.4, p = 0.5. (c) Activity duration did not differ by genotype. Alpha FF ANOVA omnibus: F(3,24) = 0.3, p = 0.8; GT: F(1,24) < 0.1, p = 0.8; sex: F(1,24) = 0.3, p = 0.5; GT*Sex: F(1,24) = 0.9, p = 0.3. (d) Representative, doubled-plotted wheel-running actograms illustrating free-running locomotor rhythms in both genotypes. (e and f) Period and error of period in each genotype. Note: Period was measured with a linear regression fit to activity onsets, and error of period was calculated from the deviation of daily onsets around the linear fit for each hamster. Period FF ANOVA omnibus: F(3,30) = 1.2, p = 0.3; GT: F(1,30) = 2.5, p = 0.12; sex: F(1,30) < 0.1, p = 0.9; GT*Sex: F(1,30) = 1.1, p = 0.3. Error FF ANOVA omnibus: F(3,30) = 3.8, p < 0.05; GT: F(1,30) = 0.1, p = 0.7; sex: F(1,30) = 10.2, p < 0.003; GT*Sex: F(1,30) < 0.1, p = 0.9.
Next, free-running rhythms were examined under DD (Figure 2d). Wildtype and Per2LUC/LUC Syrian hamsters of both sexes were entrained to LD14:10, then released into DD for 9-12 days. The period of free-running rhythms did not differ between Per2LUC/LUC and wildtype hamsters (Figure 2e). Variance in period did not differ by genotype, but this measure of circadian clock function did differ by sex (Figure 2f). Specifically, female Syrian hamsters of both genotypes displayed greater variance in free-running period than their male counterparts.
Re-entrainment to simulated jetlag was examined first by advancing the LD14:10 photocycle by 8 h, and then by delaying light offset by 8 h after a stable phase angle had been achieved (Figure 3a). Non-transgenic and Per2LUC/LUC Syrian hamsters re-entrained within 2-3 weeks (Figure 3b and c), consistent with previous work in wildtype hamsters (Sisson et al., 2022). Following the 8-h LD advance, the number of days to re-entrainment did not differ by genotype or sex (Figure 3b). Similarly, re-entrainment to the 8-h LD delay did not differ by genotype or sex (Figure 3c).
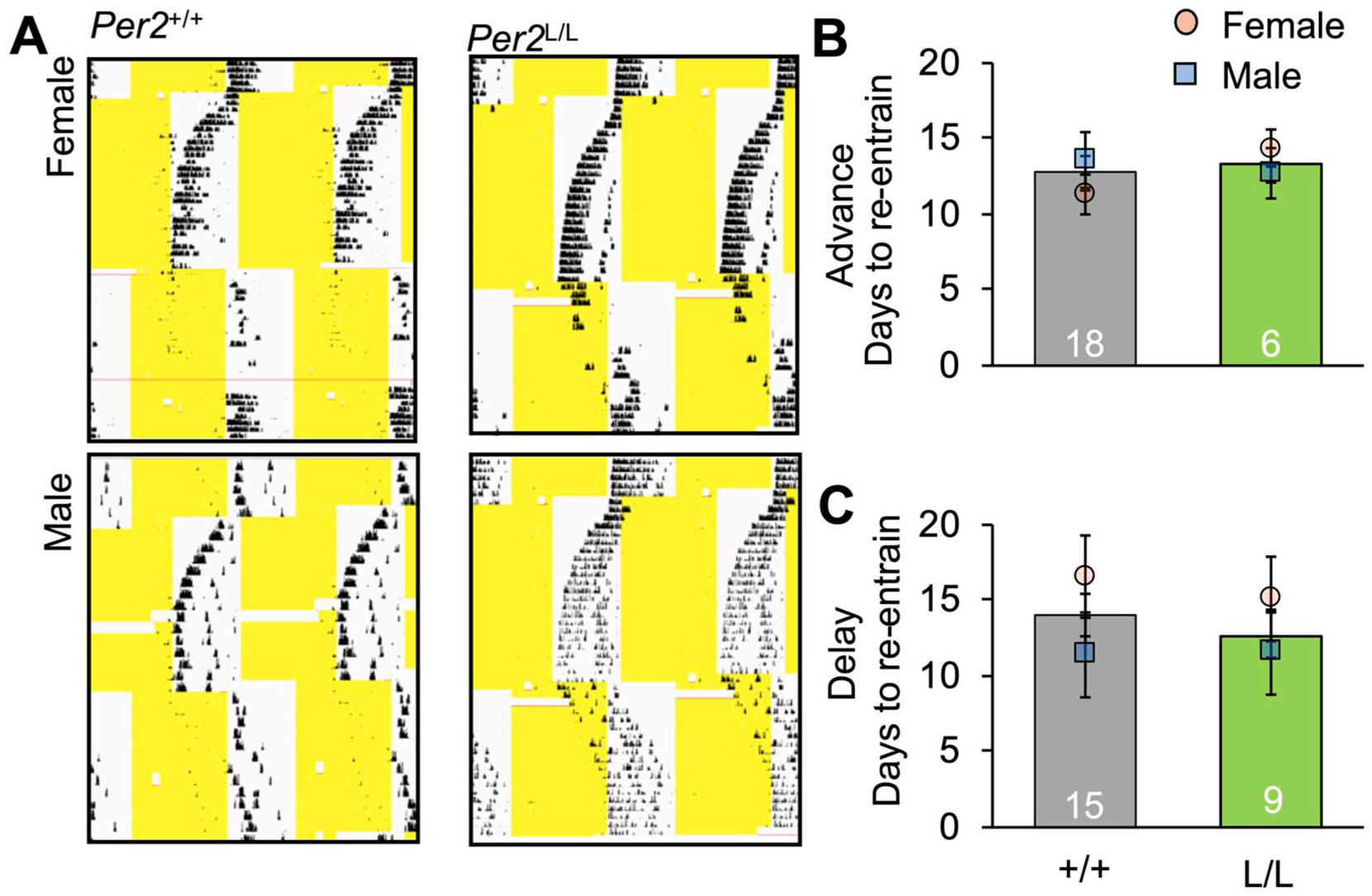
Re-entrainment to simulated jetlag in Per2LUC/LUC (L/L) and Per2+/+ hamsters (+/+). (a) Representative, doubled-plotted wheel-running actograms illustrating re-entrainment of locomotor rhythms during simulated jetlag in both genotypes. (b and c) Days to re-entrain in the eastward (8 h advance, FF ANOVA omnibus: F(3,20) = 0.4, p = 0.7; GT: F(1,20) = 0.3, p = 0.6; sex: F(1,20) < 0.1, p = 0.9; GT*Sex: F(1,30) = 0.9, p = 0.3) and westward directions (8 h delay, FF ANOVA omnibus: F(3,20) = 1.4, p = 0.2; GT: F(1,20) < 0.1, p = 0.8; sex: F(1,20) = 2.22, p = 0.15; GT*Sex: F(1,30) < 0.1, p = 0.7).
Last, Per2LUC/LUC hamsters were transferred to short-day photocycles to monitor photoperiodic changes in daily rhythms and reproduction (Figure 4). Female Per2LUC/LUC hamsters re-entrained to LD6:18 by delaying the phasing of activity onset (Figure 4a), consistent with a DD period >24 h (Figure 2e). Regular 4-day estrous cycles in behavioral and physiological indices were observed in females entrained to LD14:10 (Figure 4a, n = 7). After transfer to LD6:18, Per2LUC/LUC female hamsters displayed a positive smear every 4 days for 47.9 ± 1.6 days (Figure 4a, n = 8), after which estrous discharges ceased and the corresponding fluctuations in locomotor activity diminished (Figure 4a). Postmortem organ weights from a separate group of LD6:18 hamsters also indicated that Per2LUC/LUC hamsters were capable of regressing gonadal function after 9 weeks of short photoperiod entrainment (Figure 4b).
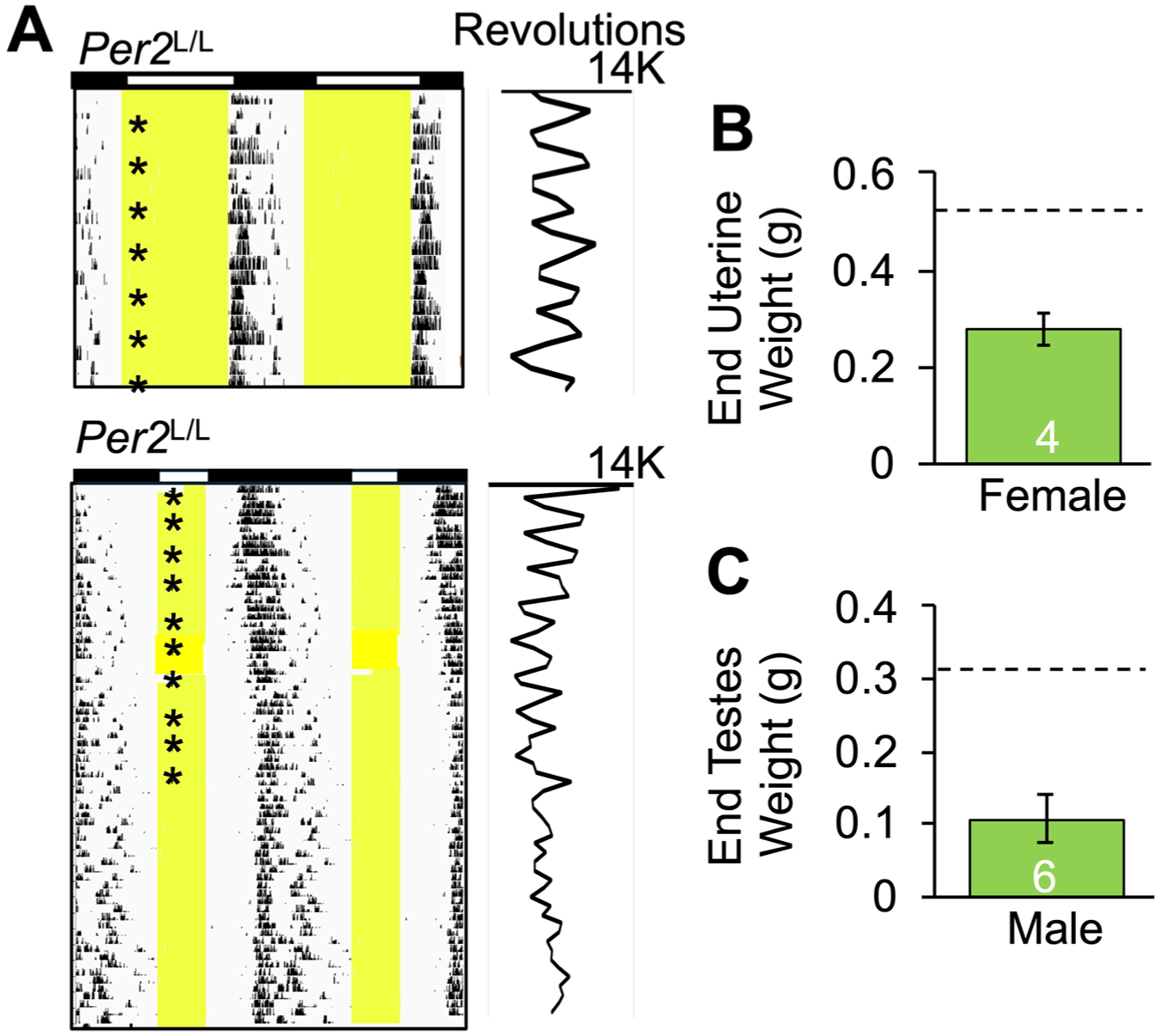
Photoperiodic modulation of reproduction in Per2LUC/LUC (L/L) hamsters. (a) Representative, doubled-plotted wheel-running actograms illustrating locomotor rhythms in female Syrian hamsters housed under LD14:10 (top) and LD6:18 (bottom). Plotted to the right of each actogram is daily total wheel revolutions, and asterisks indicate the day of a positive smear. (b and c) Paired testis and uterine weights after 9 weeks of LD6:18 entrainment in males and females, respectively. The dashed line on each panel indicates previously reported tissue weights in non-regressed hamsters of each sex maintained in long photoperiods.
Bioluminescence Luminometry and Imaging
Per2LUC/LUC and Per2LUC/+ hamsters were entrained to LD14:10 prior to PIT collection to test the degree to which Per2LUC copy number influences the properties of bioluminescence rhythms measured ex vivo (Figure 5a). The PIT was selected for initial examination due to the ease of collecting this tissue, with previous work indicating that would oscillate ex vivo, and the physiological significance of this organ for hamster physiology controlled by the circadian system (e.g., photoperiodism). Tissue samples were extracted during the middle of the photophase based on previous work, suggesting this time of tissue collection minimally resets PER2::LUC rhythms in the Per2LUC/LUC mouse (Davidson et al., 2009). Bioluminescence rhythms were monitored for at least 5 cycles, and measures of phase, period, amplitude, variability, and damping were extracted. As expected, Per2LUC copy number influenced the amplitude of bioluminescence rhythms (Figure 5b), with lower signal detected in Per2LUC/+ samples. Damping of bioluminescence rhythms also differed by Per2LUC copy number (Figure 5c), but the percentage of variance accounted for by a fitted cosine curve did not differ by genotype (data not shown, t(23) = 0.7, p = 0.48). The period length of bioluminescence rhythms did not differ by Per2LUC copy number, but cycle-to-cycle period length varied more in Per2LUC/+ samples than in Per2LUC/LUC samples (Figure 5d). Although bioluminescence rhythms damped in Per2LUC/LUC PIT samples over time in culture, higher-amplitude expression could be restored by application of dexamethasone or forskolin (Figure 5e), consistent with previous work (Balsalobre et al., 2000; Yamazaki et al., 2000). Given the impact of Per2LUC copy number on signal strength and precision without effects on period length, we used Per2LUC/LUC hamsters for further study.
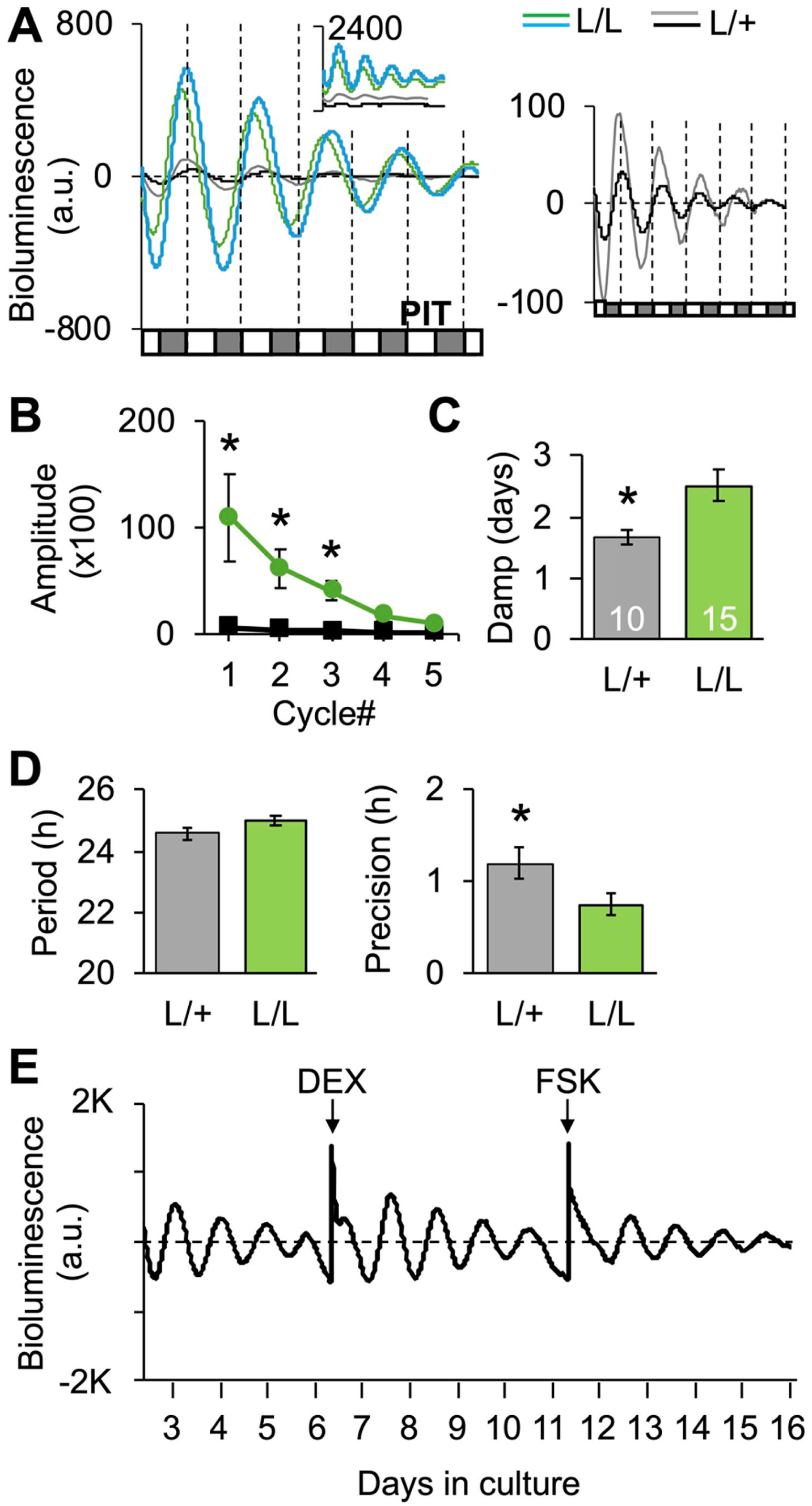
Per2-driven bioluminescence rhythms of pituitary samples collected from Per2LUC/LUC (L/L) and Per2LUC/+ hamsters (L/+). (a) Representative times series illustrating bioluminescence levels varied by transgene copy number. Left: Baseline-subtracted time series; inset illustrates raw bioluminescence with the upper value of the y-axis indicated (i.e., 2400). Two samples are illustrated for each genotype with different line colors, as represented in the symbol legend. Right: Baseline-subtracted times series for Per2LUC/+ pituitary samples, with y-axis adjusted to account for lower level of expression. Along the x-axis of each time series, white and gray boxes represent ex vivo projected times of light and darkness, respectively. (b and c) Amplitude and damping rate of bioluminescence rhythms varied by transgene copy number. Amplitude: Mixed Linear RM Model Copy#: F(1,86) = 5.4, p < 0.05; Cycle#: F(4,86) = 3.8, p < 0.01; Copy#*Cycle#: F(4,86) = 3.0, p < 0.05. Damping: t(23) = 2.9, p < 0.01; *Least Squared Means Contrasts post hoc comparisons, p < 0.05. (d) Period of bioluminescence rhythms did not differ by genotype (t(23) = 1.6, p = 0.12), but there was larger cycle-to-cycle deviation in period in Per2LUC/+ pituitary samples (t(23) = −2.3, p < 0.05). (e) Damped bioluminescence rhythms could be reinitiated with 1-h administration of 100-nM dexamethasone or 50-µM forskolin.
A variety of tissues were collected from Per2LUC/LUC hamsters of both sexes at different times of day to test whether bioluminescence rhythms monitored ex vivo are altered by dissection (Figures 6-8). Most hamster tissues examined here displayed at least 3 cycles of fluctuating bioluminescence levels (Figure 6). The exception was the testes, which failed to consistently oscillate (data not shown) despite favorable results from ovarian samples (Figure 7a). Overall, at least 50% of samples of SCN, PIN, PIT, ADR, lung, SPLN, white adipose (ADI), OVR, and liver were classified as rhythmic (Figure 7a). The percentage of tissues deemed rhythmic differed by tissue type and by time of dissection (Figure 7a). Overall, the percentage of rhythmic SCN and PIN was lower when collected at night. In addition, the goodness of fit, damping rate, and amplitude on the first cycle in vitro were affected by tissue type and/or time of dissection (goodness [tissue: F(9,364) = 6.53, p < 0.0001; time: F(3,364) = 4.75, p < 0.005]; damping [tissue: F(9,333) = 9.12, p < 0.0001; time: F(3,333) = 1.01, p > 0.3]; amplitude [tissue: (9,368) = 8.43, p < 0.0001; time: (3,368) = 0.53, p > 0.6]). Last, the period length of Per2-driven bioluminescence rhythms differed across organ types, but not by time of dissection (Figure 7c).
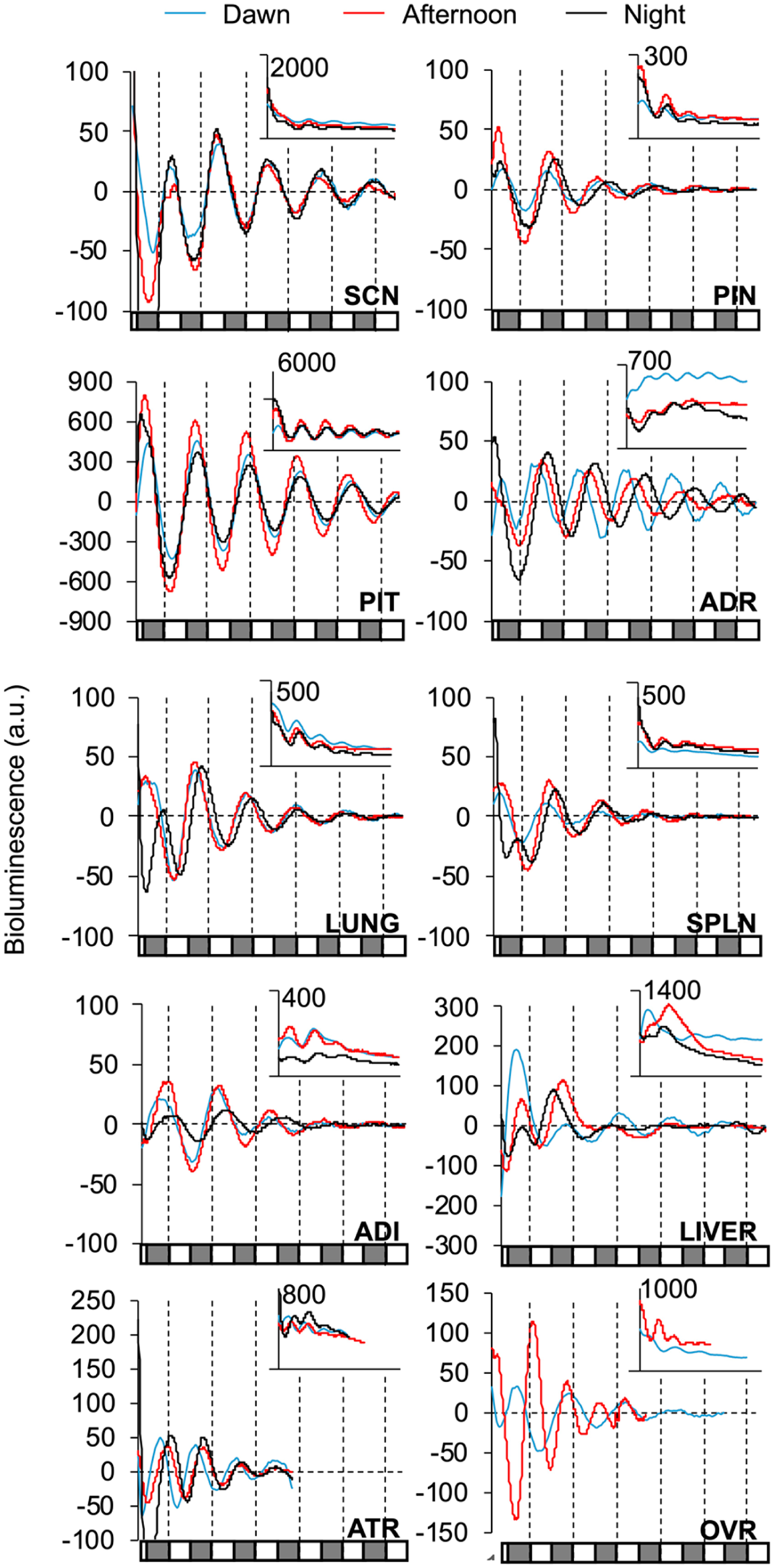
Per2-driven bioluminescence rhythms across tissues collected from Per2LUC/LUC (L/L) hamsters. Representative times series illustrating bioluminescence rhythms from each tissue when collected at different times of day. Conventions as in Figure 5.
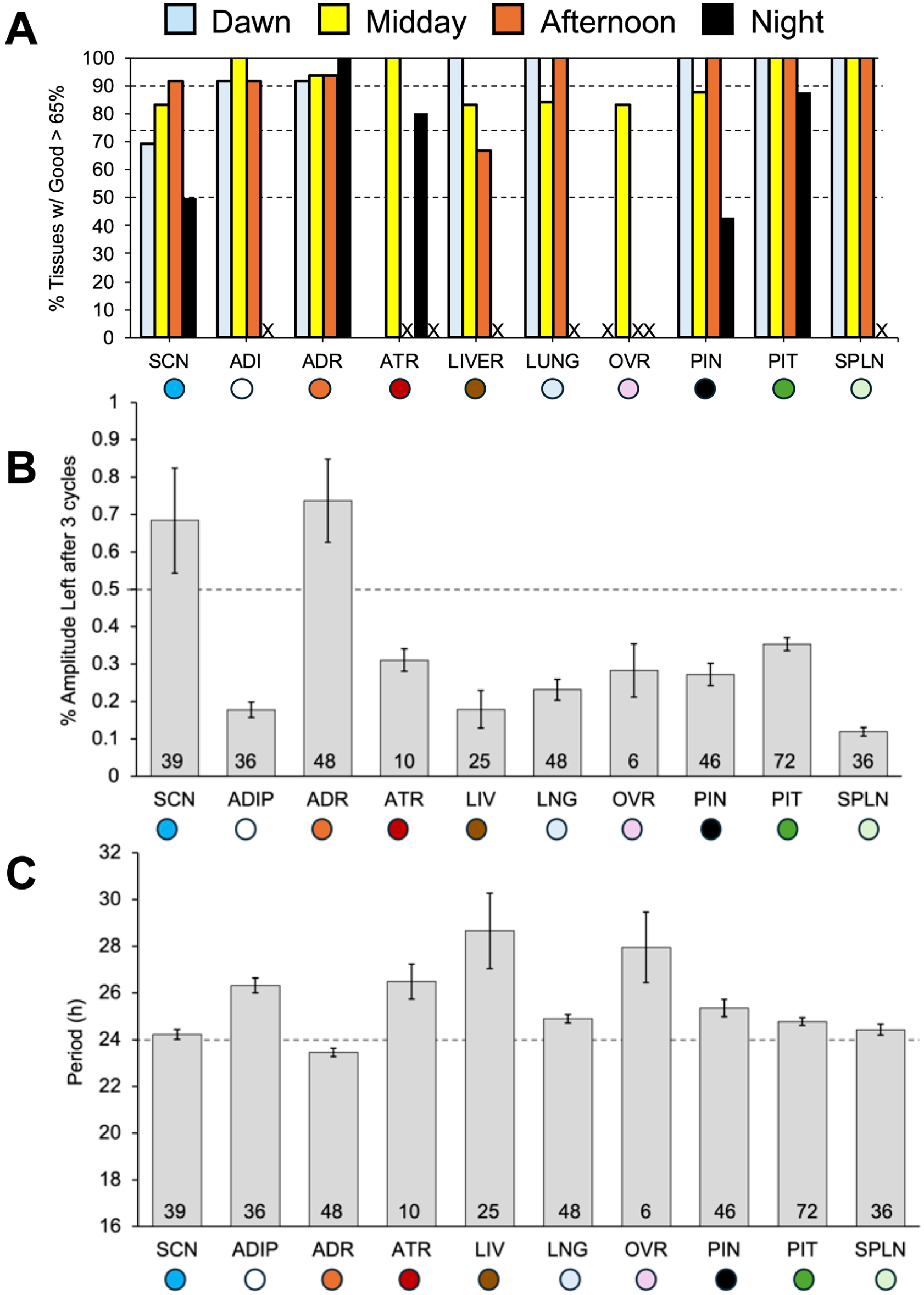
Percentage of rhythmic samples, damping, and period of Per2-driven bioluminescence rhythms over time in vitro. (a) Percentage of samples deemed to be rhythmic based on goodness of fit > 60% for each tissue type and time of dissection assessed. (a) “X” along the x-axis indicates no samples were collected for that tissue type at that time of day. FF ANOVA tissue: F(9,370) = 27.3, p < 0.005; time: F(3,370) = 16.9, p < 0.001. (b) Damping rate varied across tissue type, but not the time of dissection. FF ANOVA tissue: F(9,333) = 9.12, p < 0.0001; time: F(3,333) = 1.01, p > 0.3). (c) Period length differed by tissue type, but not time of dissection (FF ANOVA tissue: F(9,357) = 9.73, p < 0.0001; time: F(3,357) = 1.74, p > 0.15. For group sample sizes, c.f., Supplemental Table S1).
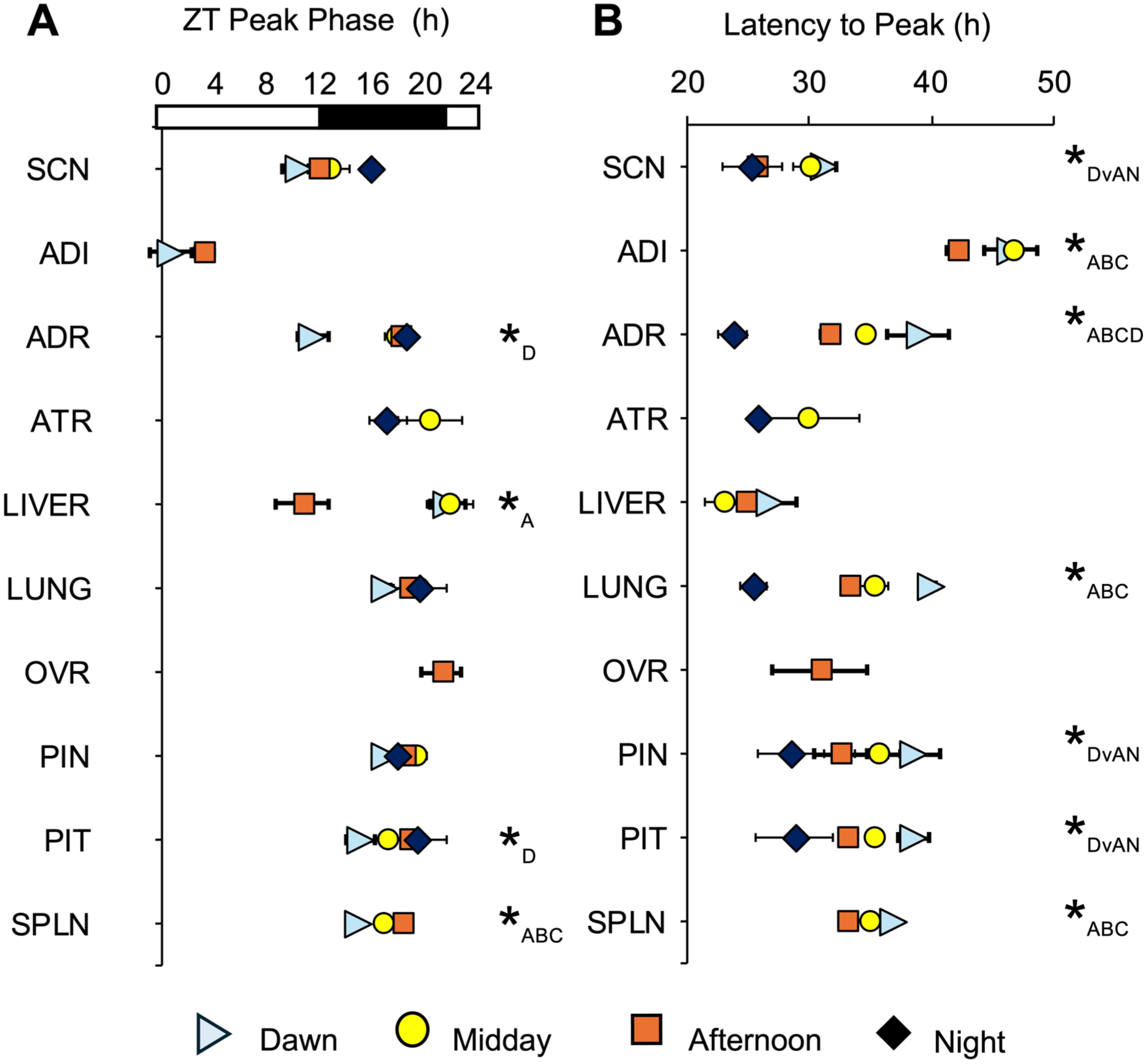
Phasing of Per2-driven bioluminescence rhythms ex vivo is affected by time of dissection in specific tissues. (a) Zeitgeber time of peak bioluminescence expression divided by tissue type and time of dissection. FF ANOVA tissue: F(9,354) = 60.4, p < 0.0001; time: F(3,354) = 8.4, p < 0.0001. (b) Time of peak bioluminescence expression ex vivo relative to time of dissection (latency to peak). FF ANOVA tissue: F(9,354) = 38.7, p < 0.0001; time: F(3,354) = 35.1, p < 0.0001. *Tukey’s HSD post hoc, p < 0.05, with subscript indicating group differences (i.e., *D = Dawn group differs from others; *DvAN = Dawn group differs from afternoon and night groups; *ABC/ABCD = all groups differ; for group sample sizes, c.f., Supplemental Table S1).
Potential resetting effects of dissection on the phase of ex vivo bioluminescence rhythms were examined using a few approaches. First, we calculated the ZT of peak bioluminescence expression on the first cycle measured ex vivo (Figure 8). Second, we examined the latency between time of dissection and peak bioluminescence expression (Figure 8). Third, we evaluated the correlation between ZT peak time and time of dissection for each tissue (Figure 9). Tissues reset by dissection are expected to differ in the ZT peak time, but not latency to peak, which would be associated with a weak correlation between ZT peak time and the time of dissection. In contrast, tissues not reset by dissection would be expected to differ in latency but not ZT peak, with a strong correlation between ZT peak time and time of dissection. Time of dissection influenced the ZT of peak bioluminescence expression in a tissue-specific manner (Figure 8a). Specifically, morning dissections appeared to reset the ADR and PIT, afternoon dissections caused the liver to peak at a ZT different from other collection times, and all dissection times differed in the ZT of peak bioluminescence expression for the SPLN (Figure 8a). Complementing these results, latency to peak bioluminescence expression was affected by tissue type and time of dissection (Figure 8b). Most tissues displayed systematic differences in the latency to the time of peak bioluminescence expression consistent with the time interval between collection time points, which suggests that these tissues are resistant to dissection-induced resetting. Latency to peak bioluminescence expression varied for each tissue, except the liver and ATR (Figure 8b), indicating that these latter two tissues were each reset by dissection. Last, we assessed the relationship between ZT peak time and the ZT of dissection through correlational analyses (Figure 9, Supplemental Table S1). Given the time-specific nature of resetting in some tissues described earlier, we fit both a linear and polynomial model to the dataset for each tissue (Supplemental Table S1). The ADR, PIT, SPLN, and liver displayed a significant linear correlation between ZT peak time and the time of dissection, with the ADR and PIT dawn samples driving this trend. Non-significant trends were also detected for the lung and SCN (Supplemental Table S1), driven by samples collected during night and dawn. Overall, these results suggest that most tissues in the Per2LUC/LUC hamster are robust to the resetting effects of dissection when collected during the afternoon (except the liver).
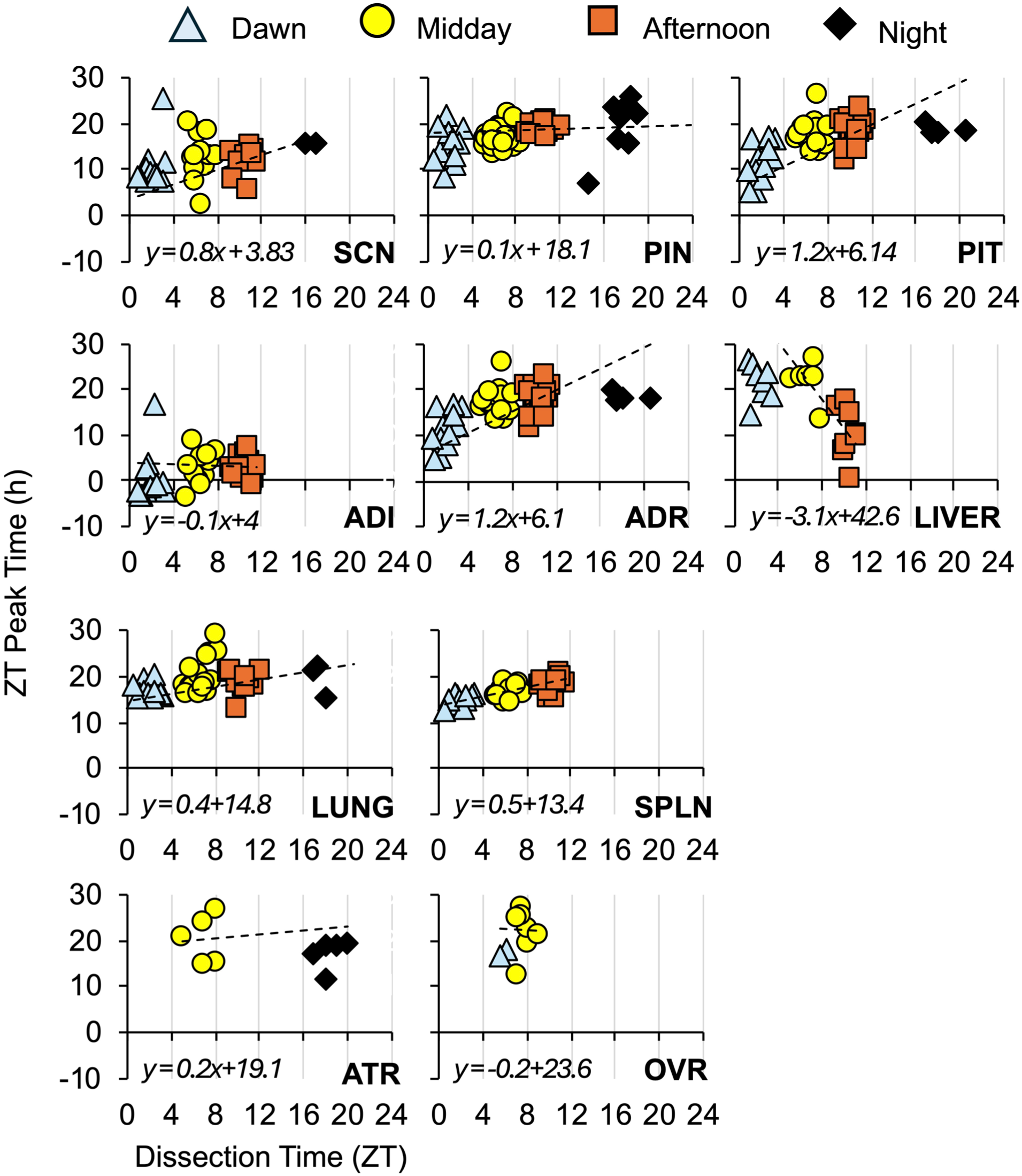
Correlational analyses of the relationship between the dissection time and the ZT peak time of Per2-driven bioluminescence in each tissue type. Each individual datapoint represents a tissue sample, with the linear regression illustrated by the dashed line fit to datapoints of each tissue type. The linear fit is represented below the x-axis for each tissue. See also Supplemental Table S1.
Next, we compared bioluminescence rhythms from the SCN, ADR, PIN, PIT, SPLN, and lung of Per2LUC/LUC hamsters to those of Per2LUC/LUC mice collected during the afternoon using identical procedures. Overall, the period length of bioluminescence rhythms was similar across species, with the exception that the Per2LUC/LUC mouse SCN displayed longer period (Figure 10a) and the Per2LUC/LUC mouse PIN displayed shorter period (Figure 10A) than observed in Per2LUC/LUC hamster samples. Phasing of peak bioluminescence expression across tissues was also comparable across species (Figure 10b and c).
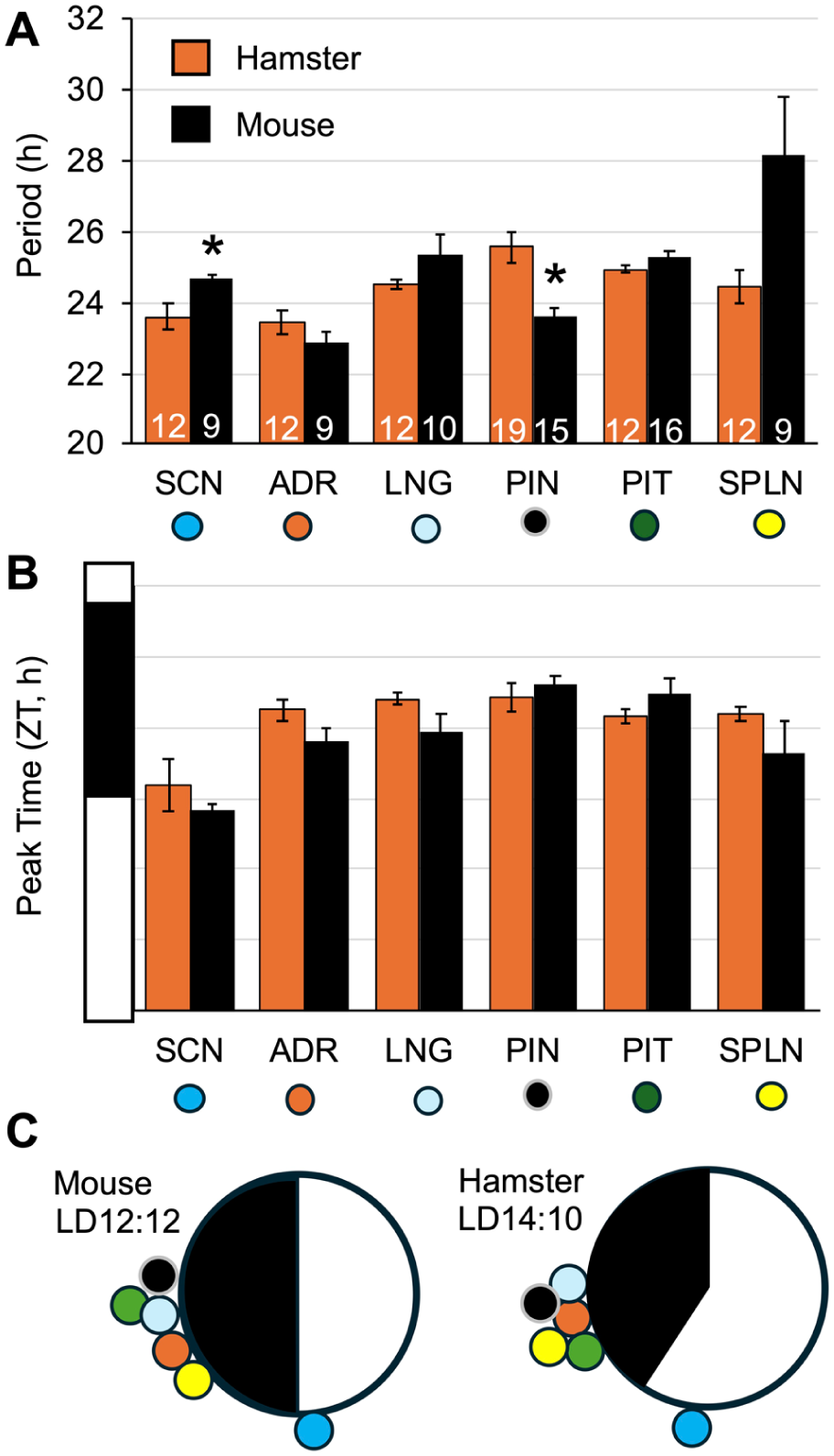
Comparison of Per2-driven bioluminescence rhythms in Per2LUC/LUC hamster and mouse tissues. (a and b) Period and ZT peak of bioluminescence rhythms in each species. (SCN period: t(19) = 2.8, p < 0.05, PIN period: t(24) = -5.9, p < 0.0001). (c) Radial plots representing phasing of bioluminescence rhythms at the systems level in each species. Note: Hamster bioluminescence data from tissues collected at midday are replotted from Figure 8.
Last, we examined the spatiotemporal organization of the hamster SCN using bioluminescence imaging (Figure 11). Four consecutive slices were taken in the coronal plane across the anteroposterior SCN of 6 hamsters (4 females, 2 males) and imaged for at least 5 cycles ex vivo. When the cumulative bioluminescence signal was evaluated relative to brightfield images taken of each slice, the dominant signal observed was derived from the dorsal SCN (Figure 11a). Lower signal could be detected in ventral SCN near the optic chiasm (e.g., Slice3), with fewer bioluminescent cell-like objects in this region. More signal was detected from middle SCN slices than in other planes (Figure 11b, left). As observed in the mouse Per2LUC/LUC SCN (Evans et al., 2011), spatiotemporal gradients in bioluminescence were observed in the hamster Per2LUC/LUC SCN across both the anteroposterior and dorsoventral axes (Figure 11b, right and c). Specifically, bioluminescence rhythms peaked earlier in the posterior SCN and dorsal SCN relative to anterior and ventral SCN (Figure 11b and c). These results suggest that this pattern of spatiotemporal organization is a conserved feature of SCN timekeeping in both hamsters and mice.
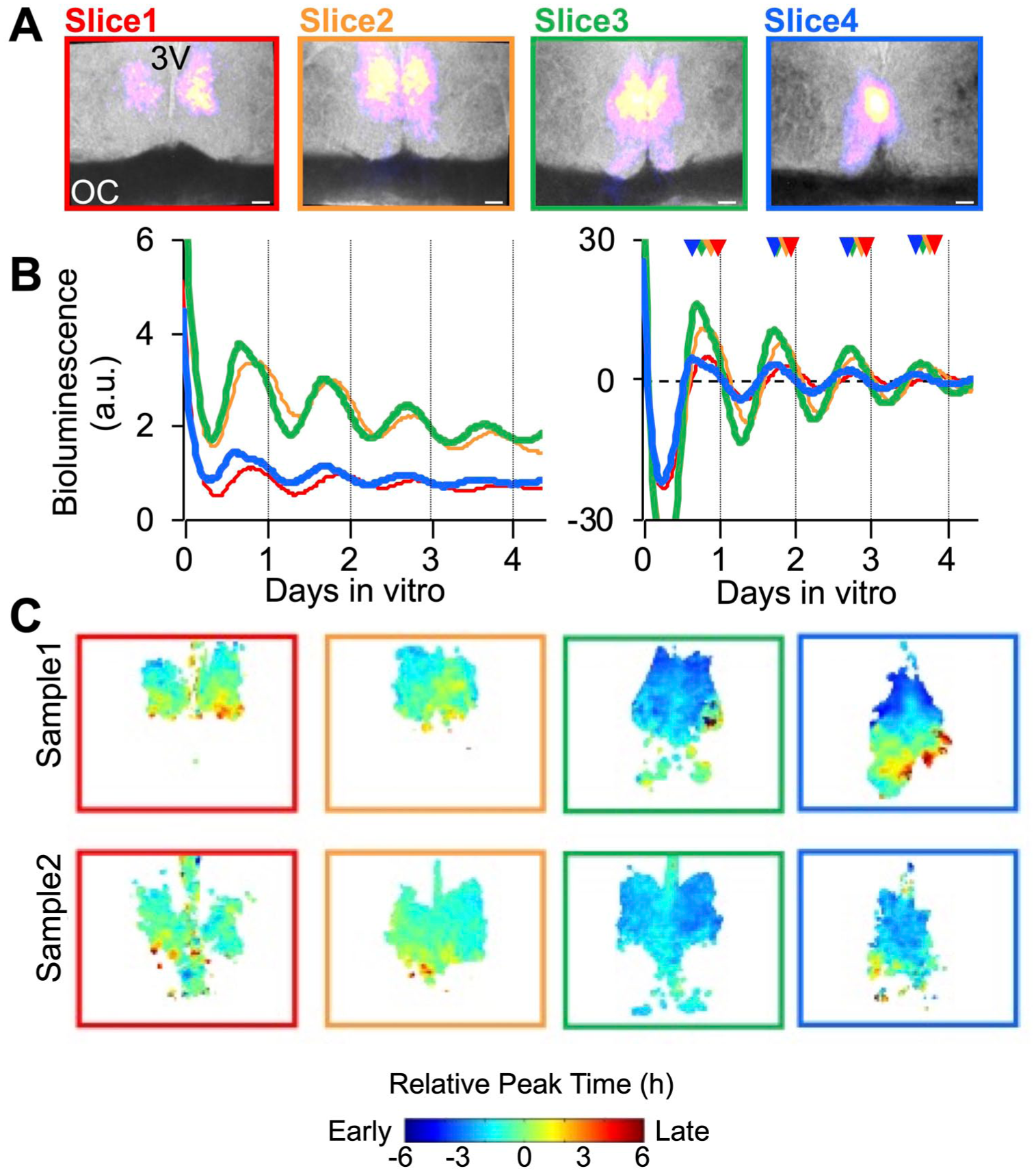
Spatiotemporal organization of Per2-driven bioluminescence rhythms in the hamster SCN. (a) Representative images of coronal SCN slices (150 μm) collected across the anteroposterior axis, with total bioluminescence over the first 24 h pseudocolored and superimposed onto bright field images. 3V: third ventricle, OC: optic chiasm, Scale bar: 100 μm. (b) Representative time series of bioluminescence rhythms measured from the SCN at each anteroposterior position. Left: raw time series. Right: baseline-subtracted time series. Color-coded arrowheads on the right plot indicate peak time for each slice. (c) Color-coded phase maps represent regional peak Per2-driven bioluminescence on the first cycle in vitro. Warm and cool colors represent late and early peaking regions, respectively. Individual phase maps are representative of n = 6 (4 females and 2 males).
Discussion
The Syrian hamster is highly valued in circadian studies for its rhythmic precision and physiological responses to photoperiod. Due to the lack of genetic engineering techniques in this species, a bioluminescent clock reporter model had not yet been established. Here, we report development of the Per2LUC hamster using the CRISPR/Cas9 technology, along with behavioral and molecular evaluation of circadian function in this new model. We find that behavioral indices of circadian and photoperiodic function appear normal in Per2LUC/LUC hamsters. Furthermore, we demonstrate that Per2-driven bioluminescence rhythms in many collected tissues are robust to the resetting effects of dissection, especially when this procedure is performed during the afternoon. Building on these results, we provide a system-level atlas for molecular clock function in the hamster, which complements previous work in mice and rats (Abe et al., 2002; Evans et al., 2015; Yamazaki et al., 2002; Yoo et al., 2004). These results provide new information into similarities and differences of molecular clock function across mammalian species. Syrian hamsters are used in several fields due to advantageous features that help with modeling human disease (Hirose and Ogura, 2019; Valentine et al., 2011; Warner et al., 2017). Given the utility of the Syrian hamster model, it may be used to advance insights into how circadian clock function interacts with a range of biological systems.
The Per2LUC hamster was generated using CRISPR-mediated approaches that have been optimized for the hamster by one of our labs (Fan et al., 2014; Li et al., 2018). We used CRISPR after first trying piggyBac-mediated transgenesis to establish mPer2-luc and mBmal1-luc models in which the expression of the luciferase reporter was under the control of mouse Per2 and Bmal1 promoters, respectively. However, results of this approach did not faithfully recapitulate the known function of the promoters in that the signal did not oscillate (data not shown), which may reflect the minimal promoter sequence used, species differences in genetic function, and/or the importance of the integration site. Therefore, we employed CRISPR to knock in a T2A-Luc cassette into the hamster Per2 locus to render expression under the control of the endogenous hamster promoter and its other regulatory sequences. The T2A linker is a common element designed to generate bicistronic expression without adversely altering the function or homeostasis of either protein (Wang and Marchisio, 2021). CRISPR was also adopted to minimize the inclusion of foreign sequences into the hamster Per2 locus, as observed in other approaches (e.g., conventional homologous recombination in embryonic stem cells or plasmid random integration). Thus, the resulting model differed from common transcriptional reporters in that the transgene is regulated by native genetic and epigenetic mechanisms. Furthermore, our model differs from the Per2LUC mouse in that the resulting signal is not generated by a fusion protein, but instead it is expected that T2A allows for the translation of 2 separate proteins differentially regulated by post-translational mechanisms. Although this was not examined here, the T2A sequence is expected to dissociate spatiotemporal kinetics of PER2 and LUC, with the former translocating to the nucleus after post-translational modification and latter being retained in the cytoplasm. The half-life of endogenous PER2 protein, mouse PER2::LUC, and luciferase are all short (i.e., 2-3 h, Etchegaray et al., 2009; Maywood et al., 2011; Meng et al., 2008; Smyllie et al., 2025b) relative to the period of circadian cycles (i.e., ca. 24 h), which suggests that transcriptional activity likely plays a major role in controlling bioluminescence rhythms generated by Per2LUC hamster tissue.
One of the major challenges faced in creating transgenic hamsters is that embryos are extremely sensitive to handling. For instance, total arrest of hamster zygote development is induced by exposure to light (Takenaka et al., 2007), which requires that hamster embryo handling be performed in a dark room under red light. Our success in conducting genetic engineering in hamster embryos helped to identify the optimal timing of embryo isolation and pronuclear injection (i.e., 18 h after mating), minimized the time required for pronuclear injections, and utilized the optimal parameters for delivering sgRNA/Cas9 vectors identified previously (Fan et al., 2014). The efficiency of Per2LUC insertion was lower than that reported in our previous work (i.e., 3/26 pups were positive for the transgene), suggesting that CRISPR-mediated insertion can vary in the hamster by genomic loci and/or length of inserted transgene. Sequencing results indicated that the Per2 and transgene cassette is not altered in the Per2LUC Syrian hamster, but that one indel was incurred downstream of the Luc stop codon. As discussed in greater detail below, our collective results indicate that this produced minimal adverse effects on molecular clock function and circadian physiology.
Using classic behavioral indices, we observed that basic circadian and photoperiodic responses are intact in the Per2LUC Syrian hamster. First, we found that photoentrainment in Per2LUC/LUC Syrian hamsters did not differ from that in the wildtype phenotype. Furthermore, the behavioral free-running period in DD did not differ statistically between wildtype and Per2LUC/LUC Syrian hamsters, although the period was 0.19 h longer in the latter. In the Per2LUC/LUC mouse, transgene insertion also lengthens free-running period of locomotor rhythms, with an increase of 0.1 h (Pastrick et al., 2024; Yoo et al., 2004) to 0.9 h (Ralph et al., 2021) relative to Per2+/+ mice. Given that previous work indicates lengthened period is displayed by the Per2LUC/LUC mouse after 4-5 weeks in DD, it is possible that a statistically significant effect of genotype might emerge in Per2LUC/LUC hamsters with longer durations of DD exposure. Next, we evaluated whether circadian responses to light were altered by transgene insertion using a simulated jetlag assay. Wildtype and Per2LUC/LUC Syrian hamsters did not differ in the number of days to re-entrainment following LD shifts in either the advance or delay direction. Likewise, wildtype and Per2LUC/LUC mice do not differ in the rate of re-entrainment following an advance of the LD cycle (Pastrick et al., 2024), but Per2LUC/LUC mice display less-robust rhythms under constant light (Ralph et al., 2021). Although constant light was not included in the battery of behavioral assays employed here, future studies modulating the duration of daily light exposure should consider whether this phenotype is more likely to manifest in Syrian hamsters carrying the Per2LUC transgene. Last, we evaluated reproductive quiescence induced by short photoperiods in Per2LUC/LUC Syrian hamsters. We found that female Per2LUC/LUC Syrian hamsters displayed regular, 4-day estrous cycles under LD14:10, with corresponding fluctuations in total wheel revolutions. Both physiological and behavioral measures indicated that the transfer to LD6:18 caused estrous cycles to cease in female Per2LUC/LUC Syrian hamsters within the typical latency displayed by this species when housed under short days or DD conditions at room temperature (Larkin et al., 2002; Manoogian et al., 2024; Pieper et al., 1986; Seegal and Goldman, 1975). Consistent with these overt indices, end-organ weights indicated that transfer to short days caused gonadal regression in both sexes, as shown previously for wildtype Syrian hamsters (Carr et al., 2003; Manoogian et al., 2024; Tong et al., 2004). Overall, these results indicate that the Per2LUC insertion does not eliminate circadian or photoperiodic responses to light in the Syrian hamster, although the free-running period of behavioral rhythms is lengthened slightly.
In addition, we evaluated the potential impact of transgene insertion by investigating the properties of bioluminescence rhythms in tissues collected from Per2LUC Syrian hamsters. Although we did not observe a statistically significant effect of Per2LUC copy number on the period of bioluminescence rhythms from the PIT gland, Per2LUC/LUC samples displayed a period 0.4 h longer than Per2LUC/+ samples. The Per2LUC copy number significantly influenced amplitude and damping of bioluminescence rhythms in the PIT gland, but bioluminescence rhythms damped over time in samples regardless of transgene copy number, consistent with previous work in rats and mice (Abe et al., 2002; Evans et al., 2015; Yamazaki et al., 2002; Yoo et al., 2004). Application of dexamethasone or forskolin was sufficient to restore higher-amplitude oscillations in Per2LUC/LUC hamster PIT gland explants, as in previous work using other rodent models (Abe et al., 2002; Balsalobre et al., 2000; Kiessling et al., 2017). Together with overt indices described earlier, these results suggest that the function of Per2 was not adversely altered by transgene insertion. Having established that molecular clock function was intact, we next collected a wider range of tissues to evaluate the extent to which time of dissection affected properties of bioluminescence rhythms. Time of dissection influenced the fraction of rhythmic samples, but only tissue type affected the amplitude and damping rate of bioluminescence rhythms. Period, amplitude, and phase also varied by tissue type, which is consistent with previous work in the Per1-luc rat (Abe et al., 2002; Yamazaki et al., 2000) and Per2LUC mouse (Davidson et al., 2009; Evans et al., 2015; Yoo et al., 2004). Notably, the period of bioluminescence rhythms was in the circadian range for all tissues collected at each dissection time. In addition, we used multiple measures to test dissection-induced phase resetting by dissection. Together, these three complementary measures indicate that most tissues examined here (except liver) were robust to the resetting effects of dissection when the tissue is collected during the afternoon, as suggested previously for Per2LUC mouse tissues (Davidson et al., 2009; Evans et al., 2015).
These data provide a comprehensive screen of molecular clock function in a range of tissues collected from both central and peripheral organs in the hamster. This provides a system-level atlas for molecular timekeeping in a species in which plasticity in SCN encoding causes adaptations at the behavioral, physiological, and molecular levels (e.g., melatonin-induced photoperiodic regulation; Carr et al., 2003; Manoogian et al., 2024; Tong et al., 2004). Although we did not compare ex vivo results with in vivo phases, comparison to published work suggests that the ZT phases of tissues examined here resemble those expected for the hamster. In particular, the SCN exhibited peak Per2-driven bioluminescence expression near ZT12, which is consistent with the reported timing of peak PER2 expression in vivo (Dey et al., 2005; LeSauter et al., 2012). Although limited protein-level analyses have been performed on a full range of tissues in the hamster, the phasing of non-SCN tissues also appears comparable if one considers the typical 4- to 6-h lag between mRNA and protein expression typical of Per2/PER2 (Lee et al., 2001). Specifically, Per2 mRNA peaks in vivo at ZT15 for the ADR, liver, heart, and SPLN (Guo et al., 2006), which is expected to correspond to peak PER2 expression at ZT19-21. The ex vivo phasing of bioluminescence rhythms in these tissues ranged from ZT18 to ZT21, suggesting that the system-level atlas provided here for the hamster can be used to infer the in vivo state of the circadian system. Likewise, there appears to be good correspondence between phase measures for the limited number of tissues where this has been examined directly in other circadian clock gene reporter models (Abe et al., 2002). Thus, our results suggest that the SCN phase leads most peripheral tissues by 3-9 h in the Per2LUC hamster, which is similar to system-level organization in the Per2LUC mouse (Evans et al., 2015; Yoo et al., 2004).
Last, we find that bioluminescence rhythms were largely similar between Per2LUC hamster and mouse tissues, which suggests that important features of circadian organization are conserved across these mammalian species. Nevertheless, some variance was observed in specific tissues despite being collected at the same time of day using the same procedures at the same institution. The main differences observed were the period of bioluminescence rhythms from the SCN and PIN gland. Unexpectedly, the mouse SCN displayed a longer period of bioluminescence rhythms than the hamster SCN, which differs from the species difference in free-running locomotor rhythms (i.e., hamsters having longer behavioral period than mouse). This may represent differential effects of transgene insertion in these two distinctly engineered models given the previous one suggests that transgene insertion influences behavioral rhythms in the Per2LUC mouse (Ralph et al., 2021). For the species difference detected in the period length of bioluminescence rhythms from the PIN gland, it is tempting to speculate that this is related to species differences in melatonin synthesis since this hormone has been noted to modulate molecular rhythms in other tissues (Baba et al., 2015; Shimomura et al., 2010). However, one limitation in the current work is that the photocycle for our hamster and mouse colonies was different (i.e., LD14:10 versus 12:12). Although difficult to discount, long photoperiods do not appear to be sufficient to modulate the period of either the SCN or PIN gland in the Per2LUC mouse (Evans et al., 2013; Evans et al., 2015). Other studies have similarly found that SCN period does not differ between mice housed in LD12:12 and those housed in LD16:8 (Ciarleglio et al., 2011; Myung et al., 2015; Tackenberg and McMahon, 2018). Last, we observed that the hamster displays SCN spatiotemporal organization similar to the Per2LUC mouse (Evans et al., 2011), with the posterior and dorsal SCN displaying an earlier peak phase of bioluminescence rhythms relative to the anterior and ventral SCN. The bioluminescence signal itself was not as uniformly expressed across the Per2LUC hamster SCN as has been found typically in SCN slices collected and acutely imaged from adult Per2LUC mouse. This may reflect species differences in the oscillatory function of the SCN network since the hamster SCN contains a calbindin + subnucleus devoid of electrical and molecular rhythms (Antle et al., 2003). In previous work, PER2 protein expression in the hamster SCN appears to be more widespread across the ventral SCN than that observed here for Per2-driven bioluminescence (Dey et al., 2005; LeSauter et al., 2012; Schwartz et al., 2011), so it remains possible that the lack of signal in the ventral SCN reflects failure of the transgene to be properly expressed in all SCN cells. Future studies are needed to build on these results and to address this issue directly.
The Per2LUC hamster is a model with many potential applications across a range of fields. First, the Syrian hamster is a highly valued model in the circadian field due to the precision and plasticity of daily rhythms in this species (Evans and Gorman, 2016; Pittendrigh and Daan, 1976a). Studies using the Syrian hamster have provided key insights into mechanisms of circadian timekeeping, including evidence supporting the SCN as the location of the central clock (Lehman et al., 1987), its multi-oscillator structure (Elliott and Tamarkin, 1994), and the genetic basis of circadian timekeeping (Ralph et al., 1990; Ralph and Menaker, 1988). In addition, the Syrian hamster has been an excellent model for understanding interactions between circadian timekeeping and reproduction (Darrow et al., 1980; Fitzgerald and Zucker, 1976; Hazlerigg et al., 2024; Zucker et al., 1980). Furthermore, the Syrian hamster produces melatonin, a hormone that regulates a range of biological processes, including reproduction, growth, and metabolism (Zhao et al., 2019). Melatonin is not the only mechanism regulating photoperiodic changes in metabolism (Bartness and Wade, 1985), and the other signals regulating this process may be explored using the Per2LUC hamster. In addition, the Syrian hamster is a facultative hibernator in which torpor is triggered by short photoperiods, cold, and food shortage (Ueda and Ibuka, 1995). Thus, the Per2LUC hamster may be useful for investigating how these factors modulate molecular clock function in tissues involved in hibernation and other seasonal adaptations. Last, hamsters are valuable research models in several fields because they more closely recapitulate human disease than mice or rats. For example, hamsters are valuable in drug studies on lipoprotein metabolism (Jove et al., 2013), respiratory disease (Safronetz et al., 2012), bacterial infections (Best et al., 2012), inflammatory myopathies (Paciello et al., 2010), and cancer biology (Takahashi et al., 2011; Yapijakis et al., 2019). Thus, we hope that the Per2LUC hamster will prove to be a valuable model both inside and outside the circadian field.
Supplemental Material
sj-docx-1-jbr-10.1177_07487304261459487 – Supplemental material for Circadian Behavioral and Bioluminescence Rhythms in the Per2LUC Syrian Hamster
Supplemental material, sj-docx-1-jbr-10.1177_07487304261459487 for Circadian Behavioral and Bioluminescence Rhythms in the Per2LUC Syrian Hamster by Yanan Liu, Elizabeth Roszkowski, Rong Li, Chip Sisson, Ella Landman, David R. Weaver, Zhongde Wang, Jennifer A. Evans and Eric L. Bittman in Journal of Biological Rhythms
Supplemental Material
sj-pdf-1-jbr-10.1177_07487304261459487 – Supplemental material for Circadian Behavioral and Bioluminescence Rhythms in the Per2LUC Syrian Hamster
Supplemental material, sj-pdf-1-jbr-10.1177_07487304261459487 for Circadian Behavioral and Bioluminescence Rhythms in the Per2LUC Syrian Hamster by Yanan Liu, Elizabeth Roszkowski, Rong Li, Chip Sisson, Ella Landman, David R. Weaver, Zhongde Wang, Jennifer A. Evans and Eric L. Bittman in Journal of Biological Rhythms
Footnotes
Acknowledgements
We thank the animal care staff at each institution for their assistance in carrying out these studies. We also thank Alec Huber, Claire Wulf, Maureen Moore, Davis Xhaxho, Armando Perea, Sydney McGraw, Jonathan Abon, and Alphonse DeVita for their contributions.
Author Contributions
YL: Resources, investigation, formal analysis, validation, visualization, writing; ER: Investigation, formal analysis, data curation; RL: Investigation; CS: Investigation; EL: Investigation, formal analysis; DRW: Methodology, resources, investigation, formal analysis, writing; ZW: Conceptualization, methodology, resources, supervision, funding acquisition, writing; JAE: Conceptualization, methodology, investigation, formal analysis, visualization, supervision, funding acquisition, writing; ELB: Conceptualization, methodology, resources, formal analysis, supervision, investigation, funding acquisition, writing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health awards R21NS099473, R01NHLBI 138551, and R21NS135308. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Jennifer A. Evans is a member of the Editorial Board of the Journal of Biological Rhythms. The author did not take part in the peer review or decision-making process of this submission and has no further conflicts to declare.
Supplemental Material
Supplementary material is available for this article online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.