
Abstract
Background:
The ongoing spillover of highly pathogenic avian influenza H5N1 virus into mammalian species, including recent outbreaks in dairy cattle, underscores an urgent need for broadly effective therapeutics. Nanobody, the variable domain of heavy chain-only antibody, offers unique advantages for viral neutralization due to its small size, stability, and accessibility to cryptic epitopes.
Methods:
In this study, we isolated a human nanobody, nAb35, targeting the hemagglutinin of the dairy cattle H5N1 virus A/Texas/37/2024 (clade 2.3.4.4b) via phage display of a human nanobody library. After three rounds of panning against the recombinant H5 ectodomain, a dominant clone (nAb35) was highly enriched. Then nAb35 was fused with a human IgG1 Fc fragment and expressed as a bivalent dimer.
Results:
Biochemical characterization demonstrated that nAb35 specifically binds to H5 under both denaturing and native conditions and recognizes a linear epitope accessible on H5 expressed on the cell surface. In vitro neutralization assays using H5 pseudovirus showed that nAb35 inhibited viral entry with a half-maximal effective concentration of approximately 10 μg/mL, as determined by both luciferase reporter and immunofluorescence assays.
Conclusion:
These findings identify nAb35 as a promising candidate for further therapeutic development against emerging H5N1 virus.
Introduction
Highly pathogenic avian influenza (HPAI) H5N1 virus poses a persistent and escalating threat to global public health. Since its emergence in 1996, the virus has evolved into multiple phylogenetic clades, with clades 2.3.2.1 and 2.3.4.4 currently dominating circulating strains and causing sporadic fatal human infection (Xie et al., 2023). A watershed event occurred in March 2024, when HPAI H5N1 clade 2.3.4.4b was detected in dairy cattle in Texas, United States, marking the first confirmed spillover of this virus into dairy cattle (Caserta et al., 2024). At least 13 human cases are linked to dairy farm exposures as of late 2024 (Pulit-Penaloza et al., 2024). Structural analysis of cattle-derived H5N1 isolates has identified mutations that enhance binding to mammalian sialic acid receptors, raising concerns about potential adaptation for efficient human-to-human transmission (Hao et al., 2025). These spillovers underscore the urgent need for broadly effective therapeutics against emerging H5N1 strains. In this context, antibody-based therapeutics have gained increasing interest as both prophylaxis and treatment of influenza virus infection. Mouse or human IgGs have specific neutralization of H5N1 and even cross-neutralization of H1N1 (Khurana et al., 2024; Li et al., 2022; Xu et al., 2025).
Nanobody—the variable domain of heavy chain antibody derived from llama—possess unique properties that distinguish it from conventional IgG antibody. With a molecular weight of approximately 15 kDa, nanobody exhibits superior tissue penetration, including access to cryptic epitopes within viral glycoproteins that are sterically inaccessible to conventional antibody, making nanobody particularly valuable for targeting conserved functional domains on viral surface proteins (Huo et al., 2020; Isaacs et al., 2025). Furthermore, nanobody demonstrates exceptional thermal stability, solubility, and ease of production in microbial expression systems and, most importantly, enable aerosol delivery and stimulate mucosal immunity in respiratory mucosa, which is or particular significance for airborne respiratory viruses (Chow et al., 2023; Yufei et al., 2020).
The hemagglutinin (HA) of the influenza virus mediates viral entry through receptor binding and membrane fusion and comprises a globular head domain (HA1) containing the receptor binding site and a stem region critical for membrane fusion; therefore, HA is the main protective and immunodominant antigen (Hao et al., 2025). Recent studies have highlighted the potential of nanobodies targeting HA. Nanobodies have shown both therapeutic and prophylactic activities for H1N1, H3N2, and H7N9 in vitro and in vivo in a mouse challenge model (Barbieri et al., 2024; Chen et al., 2025; Hwang et al., 2024; Shcheblyakov et al., 2024). Lorena et al. immunized llamas and screened nanobodies that decreased H5N1 virus replication in lung tissues and protected mice from death after a lethal challenge with H5N1 virus (Lorena et al., 2011). More recently, Nb10 was identified through yeast two-hybrid screening and targeted the conserved site in the HA1 domain of H5. Nb10 demonstrated broad-spectrum neutralization capacity against H5N1 clade 2.3.2.1 and clade 2.3.4.4 in both in vitro and in vivo tests (Xu et al., 2025). A llama-derived nanobody–Fc fusion protein, VHH10-hFc, could recognize a conformational epitope comprising residues Q187, K189, L190, Y191, N193, T215, S217, and N220 on the globular head of H5, demonstrating broad reactivity across H5N1 clades 0, 1, 2, and 4 (Liu et al., 2025).
In this study, we isolated nAb35 targeting the emerging dairy cattle H5N1 influenza virus from a human nanobody phage display library. We demonstrated that nAb35 bound H5 specifically under both denaturing and native conditions, recognized a linear epitope accessible on the H5 expressed on the cell surface, and exhibited neutralizing activity against the H5 pseudovirus in vitro. The nAb35 may be a promising therapeutic and prophylactic antibody for HPAI H5N1.
Methods and Materials
Phage display library, cells, and virus
The human nAb library was purchased from the Wellcome Trust Sanger Institute (Cambridge, UK). HEK293F cells were grown in serum-free medium in a horizontal shaking incubator at 37°C, 5% CO2. Both HEK293T cells and Vero E6 cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum at 37°C, 5% CO2. H5 pseudovirus was constructed as previously described using the HA gene from A/Texas/37/2024 H5N1. Briefly, the H5 gene was codon-optimized and cloned into the pLP/VSVG plasmid with a firefly luciferase expression cassette, and a Flag tag was fused to the N-terminal of the H5 gene. Then, pLP1, pLP2, and pLP/H5 plasmids were co-transfected into HEK293T cells to rescue the H5 pseudovirus.
Construction, expression, and purification of the recombinant ectodomain of H5 and nanobody–Fc fusion protein
The gene of the ectodomain of H5 of A/Texas/37/2024 H5N1 was synthesized by Genewiz (Suzhou, China) and fused with a 6×His tag in the C-terminal for purification and then inserted into the eukaryotic expression vector pcDNA3.1(+). The resultant expression construct was transiently transfected into HEK293F cells. After 5-day incubation, the medium supernatant was collected and loaded into a Ni2+ affinity chromatography column (Beyotime, China). The H5 protein was eluted by 200 mM imidazole and further separated with a Superdex-200 gel filtration chromatography column (GE Healthcare, US). The purified H5 was dialyzed against PBS (pH 7.4) and stored at −80°C until use. In order to construct the nanobody–Fc fusion protein, the gene of the specific nanobody was fused to the coding gene of the human IgG1 Fc fragment and inserted into the expression vector AbVec2.0. The fusion protein nanobody–Fc was expressed in HEK293F cells as above and purified in a Protein A+G affinity chromatography column (Beyotime, China).
Screening of anti-H5 nanobody by phage display
We screened a specific anti-H5 nanobody from a human nanobody library. Briefly, an immunosorbent well was coated with H5 protein at 4°C overnight. For phage panning, the phage library was added for binding to the coated antigen H5. Unbound phages were removed by washing with 0.05% PBST. Specifically bound phages were eluted by trypsin digestion, then rescued in E. coli TG1 using KM13 helper phage and re-screened in the next round of panning. After three rounds of panning, H5-specific phages were identified by ELISA, and the coding genes of specific nanobodies were sequenced.
Specific binding of nanobody to H5
Western blot
A Western blot was performed to test the specific binding of anti-H5 nanobody to H5 in a denaturing condition, that is, whether the nanobody could recognize the linear epitope of denatured H5. After SDS-PAGE of H5, the protein band was transferred to a nitrocellulose membrane, which was then blocked by 2% BSA. Anti-H5 nanobody (1 μg/mL) was used as the primary antibody, and HRP-labeled goat anti-human IgG (H+L; 1:1,000) (Beyotime, China) was used as the secondary antibody. DAB substrate was added for color development and visualization.
Flow cytometry
The binding ability of the anti-H5 nanobody to H5 was analyzed by flow cytometry. HEK293T cells were transfected by a plasmid incorporating full-length H5 (including the transmembrane domain). At 48 h after transfection, HEK293T cells were collected and stained with anti-H5 nanobody (1 μg/mL), then stained with Alexa Fluor 488-conjugated goat anti-human IgG (Sangon, China). An irrelevant nAb was used as negative control (NC). Cell samples were analyzed by a BD FACSCalibur. Flow cytometry data were analyzed by FlowJo v10.0 (BD Biosciences).
ELISA
ELISA was performed to analyze whether the nanobody could recognize the H5 antigen specifically. Microtiter 96-well plates were coated with H5 protein (0.1 μg/well) at 4°C overnight. After blocking with 4% skim milk, 100 μL of 2-fold serially diluted nanobody (0.0625, 0.125, 0.25, 0.5, 1, 2, 4, 8, 16, and 32 μg/mL) was incubated at room temperature for 1 h. Specific binding was detected with HRP-labeled goat anti-human IgG (H + L; 1:250; Beyotime, China). Finally, color development was stopped by H2SO4, and fresh TMB substrate was added for visualization. The optical absorbance was measured at 450 nm by a microplate spectrophotometer (Thermo Scientific).
Neutralization assay
HEK-293T cells (1 × 104/well) were plated in a 96-well plate or on glass slides in a 12-well plate 12 h before infection. The Anti-H5 nanobody was serially diluted (0.001, 0.01, 0.1, 1, 5, 10, 25, 50, and 100 μg/mL and 100 μL) and incubated with H5 pseudovirus (1,000 U/well, 100 μL) at 37°C for 1 h. The HEK-293T cells were incubated with the pseudovirus-nanobody mixture at 37°C for 12 h, then in fresh complete DMEM medium for 72 h.
Luciferase-based neutralization assay
The neutralization ability of the nanobody was evaluated in a luciferase assay. Pseudovirus-infected HEK293T cells were lysed in lysis buffer, and luciferin substrate was added and incubated at room temperature for 10 min. The fluorescence intensity was detected by a Luminoskan Microplate Luminometer (Thermo Scientific).
Immunofluorescence-based neutralization assay
Then, 72 h after infection, HEK293T cells were fixed in 4% paraformaldehyde (in PBS) for 20 min at room temperature and stained for nuclei with 4,6-diamidino-2-phenylindole (DAPI). Cells were subsequently incubated with recombinant flag tag mouse mAb (Beyotime, China) at room temperature for 30 min, followed by incubation with AF594-labeled goat antimouse IgG (H+L Beyotime, China) at room temperature for 30 min. The fluorescence images were recorded using a laser confocal microscope (LSM 980 Airyscan 2, Carl Zeiss). For quantification of the neutralization ability, the number of HEK293T cells with red fluorescence was recorded by the Cytation 1 Cell Imaging Multi-Mode Reader (BioTek). The neutralization was calculated by the function: Neutralization (%) = [1- (test well - cell control)/(pseudovirus control - cell control)] × 100%.
Results
Screening nanobodies against H5
As shown in Fig. 1, a human nanobody phage display library was used to screen specific nanobodies against the HA of HPAI H5N1 virus (H5). After 3 rounds of panning and sequencing, a dominant nanobody was highly enriched, that is, 35 clones containing the same nanobody sequence were isolated from 43 positive clones. Otherwise, the remaining eight clones have one unique sequence each and are not enriched significantly. Thus, this dominant nanobody was named nAb35. An unbound nanobody (nAb1) was used as NC in the following experiments. Fig. 2 shows the alignment of nine nanobody sequences.
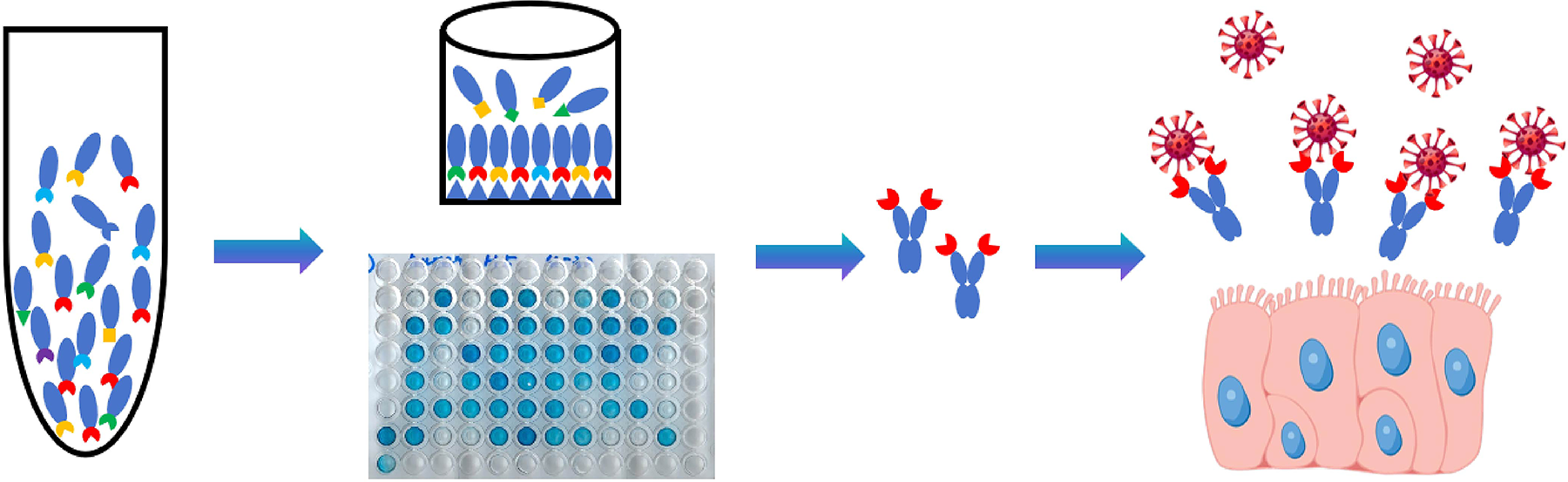
Schematic diagram of screening nanobodies against the hemagglutinin of H5N1.
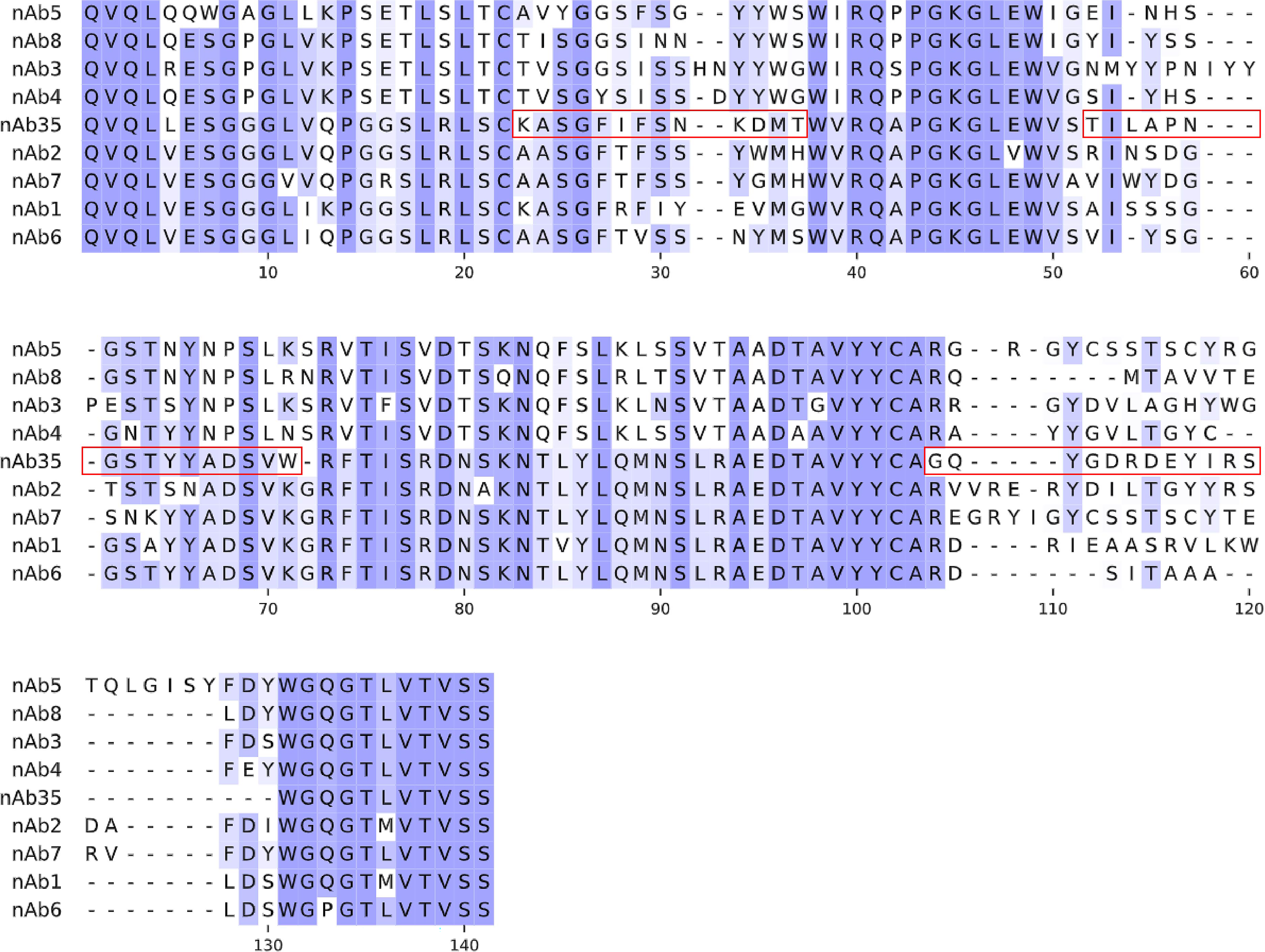
Sequence alignment of H5-bound nanobodies. Red boxes indicate CDRs.
Expression and characterization of nanobodies
The nanobody was fused with the Fc fragment of human IgG1; due to the interchain disulfide bonds, a dimer formed under native conditions. Fig. 3a and Fig. 3b showed that both the H5 ectodomain and nanobodies were successfully expressed in HEK293F cells in a suspension culture system. Both nAb35 and nAb1 formed a dimer under nonreducing conditions, while under reducing conditions, the interchain disulfide bonds were broken, and both nanobodies were in a monomer format. Dimerization could increase the affinity of the nanobody to the antigen significantly in comparison to the monomer format.
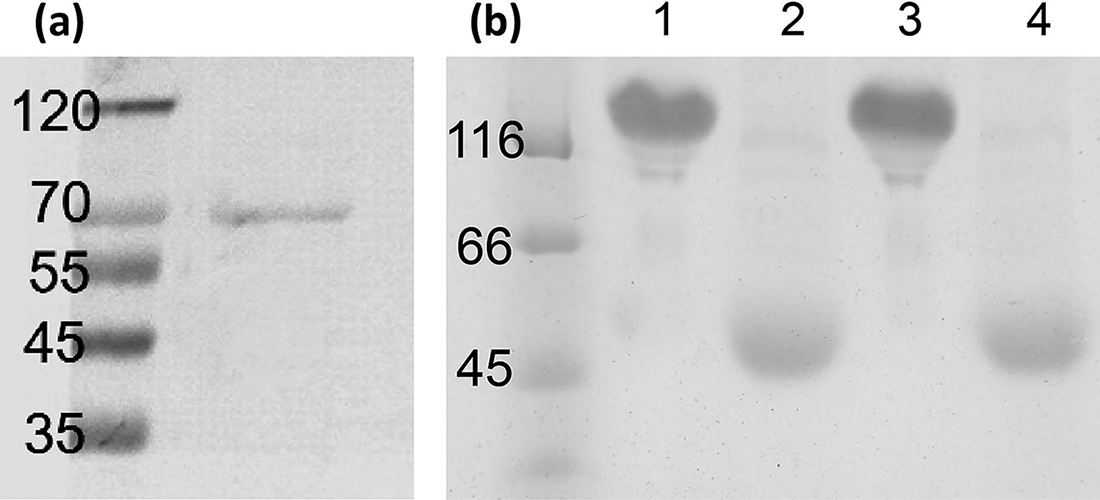
Eukaryotic expression and purification of H5 and anti-H5 nAbs. Both proteins were detected by SDS-PAGE.
As shown in Fig. 4a and 4c, both Western blot and ELISA demonstrated that nAb35 could bind to H5 specifically. As the H5 protein denatured and was in a linear format in the Western blot, nAb35 could bind to a linear epitope on H5. Flow cytometry (Fig. 4b) demonstrated that nAb35 could bind to H5 on the cell surface, which was similar to the steric structure of the influenza virus envelope; no steric hindrance was observed.
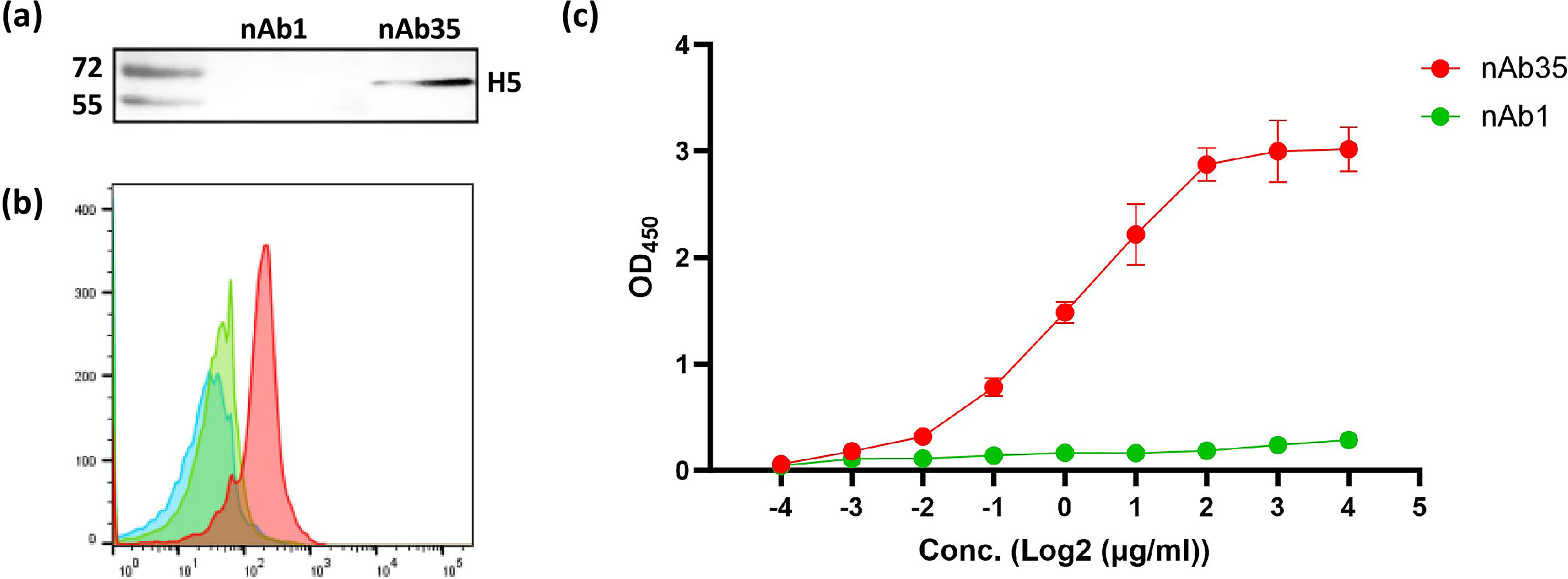
Specific binding of anti-H5 nAb to H5.
Neutralization ability of nanobodies
As H5N1 is an HPAI virus and can be manipulated in a biosafety level 3 (BSL-3) laboratory only, we tested the neutralization ability of nanobodies with an H5-based pseudovirus in our laboratory. The pseudovirus utilized a luciferase reporter system to detect the infection efficiency, and we added immunofluorescence with a Flag-tag primary antibody. As shown in Fig. 5a and 5b, the neutralization curves were subject to a sigmoidal, four-parameter logistic nonlinear fit analysis, and the half-maximal effective concentration (EC50) was calculated to be 9.8 μg/mL (pseudovirus) and 10.1 μg/mL (immunofluorescence).
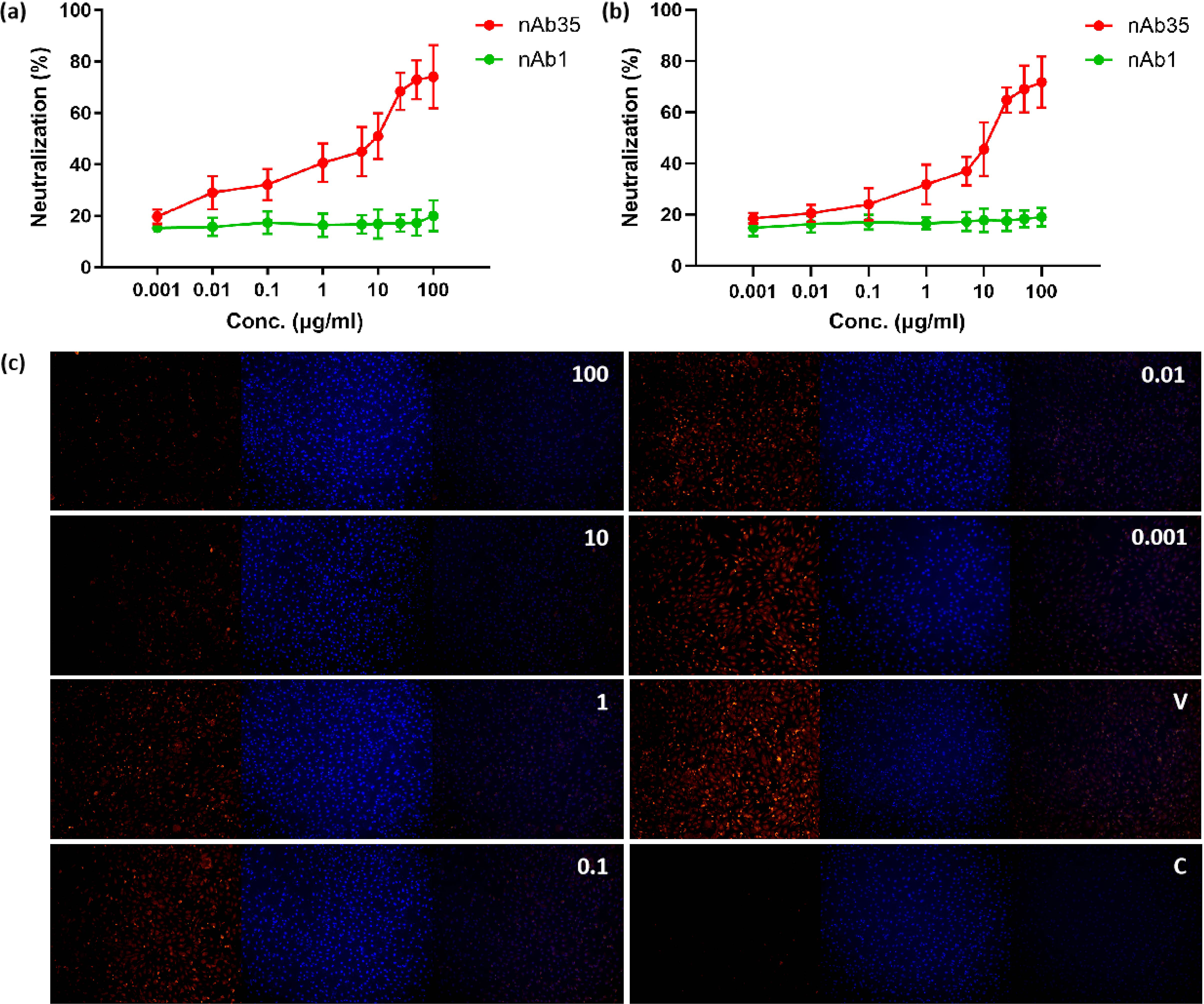
Neutralization assay of H5-based pseudovirus by anti-H5 nAb.
Specificity of nAb35 against H5
In order to determine the specificity of nAb35 to H5, the ectodomain of the H1 protein from a seasonal influenza virus A (H1N1) pdm09 was used as the coating antigen in ELISA, and full-length H1-transfected HEK293T cells were used in flow cytometry. A nanobody against H1 (nAb-H1) was used as a positive control. As shown in Fig. 6a and 6b, both ELISA and flow cytometry results confirmed no binding to H1 by nAb35, indicating the specificity of nAb35 against H5. As shown in Fig. 6c, a luciferase neutralization assay confirmed no neutralization of H1-based pseudovirus by nAb35. nAb-H1 was used as a positive control, though its neutralization potency was not high.
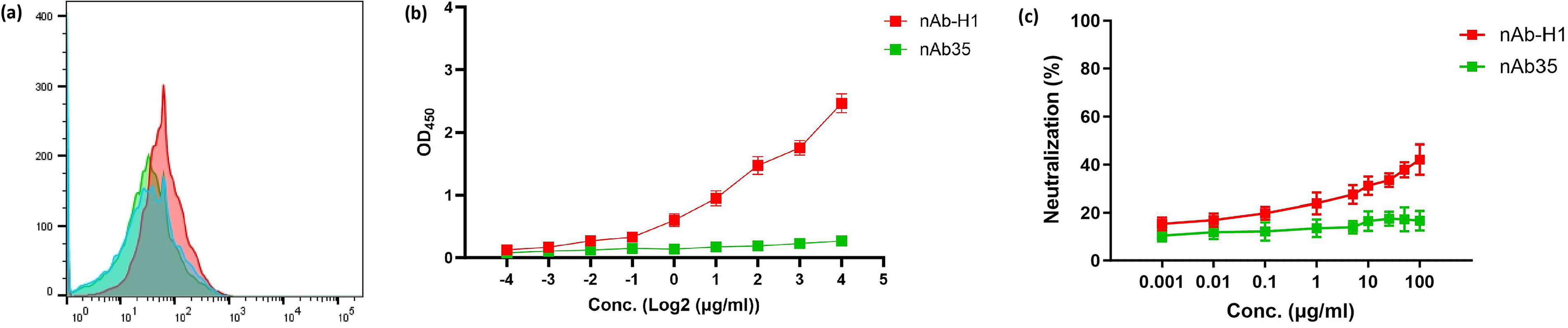
Binding to H1 protein and neutralization of H1-based pseudovirus by nAb35.
Discussion
In this study, we successfully isolated a human nanobody, nAb35, targeting the HA of the dairy cattle H5N1 virus through phage display. This nanobody demonstrated specific binding to H5, yet not H1, in multiple biochemical and cellular assays and exhibited neutralizing activity against H5-pseudovirus, rather than H1-pseudovirus, in vitro. The enrichment of nAb35 after three rounds of panning against the recombinant H5 ectodomain underscored the efficiency of the phage display platform in selecting high-affinity binders from a diverse naive library. The convergence of the selected clones towards a single sequence suggested a strong immunodominance of the epitope recognized by nAb35 within the recombinant H5 ectodomain used. This phenomenon is frequently observed in phage display, where the combination of antigen structure and elution conditions often favors a particular epitope with optimal binding kinetics under the given selective pressure (Huang et al., 2020).
Biochemical characterization revealed that nAb35 recognized a linear epitope on H5, as evidenced by its reactivity in Western blot under denatured conditions (Fig. 4a). This property distinguishes nAb35 from many conformation-dependent antibodies that fail to recognize denatured antigens. The recognition of a linear epitope may confer certain advantages, particularly in terms of stability and resistance to conformational changes in the target antigen. However, it is equally important to verify that this linear epitope is accessible on the native, membrane-bound form of H5 on the viral envelope. Flow cytometry (Fig. 4b) demonstrated clear binding of nAb35 to full-length H5 on the cell surface and provided critical validation that the epitope is indeed surface-exposed and not occluded by steric hindrance or trimerization of the HA. The red shift of nAb35, contrasted with the lack of binding by the control nAb1, confirmed that the interaction was specific and relevant to the physiological context of viral entry.
The neutralizing capacity of nAb35, with an EC50 of approximately 10 μg/mL against H5 pseudovirus, places it within a moderate potency range compared to other reported anti-influenza nanobodies and monoclonal antibodies. While some exceptionally potent antibodies achieve EC50 in the nanogram per milliliter range, the observed microgram-range activity is not uncommon for monovalent nanobodies or those requiring further affinity maturation (Beukenhorst et al., 2024; Xu et al., 2025). The format of nAb fused to human IgG1 Fc results in a bivalent molecule. Dimerization is known to enhance functional affinity through avidity effects, which likely contributes to the neutralizing activity observed compared to what a monovalent format might achieve (Li et al., 2022; Yufei et al., 2020). The neutralization curves obtained from both luciferase-based and immunofluorescence-based assays were highly concordant (Fig. 5a and 5b), mutually reinforcing the validity of the neutralization data. The immunofluorescence assay provided a valuable visualization of the protective effect, directly showing the reduction in H5-positive cells after nAb35 treatment (Fig. 4c).
Several limitations of the present study need to be addressed in the future. First, the neutralization assays were conducted using pseudovirus rather than the live, replication-competent H5N1 virus. While pseudovirus is a standard and biosafe surrogate for assessing entry inhibition, it does not fully recapitulate the complexity of live virus infection, which involves multiple rounds of replication, potential cell-to-cell spread, and interactions with other viral proteins such as neuraminidase. Therefore, validation of nAb35′s neutralizing activity against authentic H5N1 virus in a BSL-3 facility is a crucial step. Second, the EC50 of ∼10 μg/mL suggests that while nAb35 possesses intrinsic neutralizing activity, its potency may need to be enhanced for therapeutic or prophylactic applications. Strategies such as affinity maturation through directed evolution or the construction of multimer could be explored to improve its affinity or efficacy (Yufei et al., 2024; Zupancic et al., 2021). Third, the breadth of nAb35 against different H5N1 clades and other influenza subtypes remains unexplored. Given the high mutation rate of influenza viruses, cross-reactivity and the identification of the precise linear epitope are essential for assessing its potential for broad-spectrum protection and the risk of viral escape.
Conclusion
In conclusion, the present study reports the identification and initial characterization of nAb35, a human nanobody that binds to H5 and neutralizes H5 pseudovirus specifically. The binding of nAb35 to a linear, surface-exposed epitope that is accessible on both denatured protein and membrane-bound HA makes it a promising candidate for further development. Future efforts should focus on elucidating the precise molecular mechanism of neutralization, mapping its epitope at high resolution, evaluating its efficacy in animal models, and exploring strategies to enhance its potency and breadth. Continued identification and characterization of such nanobodies are vital for pandemic preparedness against evolving avian influenza threats. These findings position nAb35 as a promising candidate for further prophylactic and therapeutic development against the threat of the dairy cattle H5N1 virus. Future studies will focus on evaluating its broad-spectrum protective efficacy in vivo.
Authors’ Contributions
F.L. conducted the main experiments and wrote the article. J.C., J.Z., Y.Z., and Z.H. conducted phage display and protein expression and purification. M.Q. reviewed the article.
Authors’ Confirmation
F.L. is from Anhui Institute of Medicine (Hefei, China). M.Q. is from Anhui Medical University (Hefei, China). Education and research are the primary functions for both.
Footnotes
Acknowledgment
The authors thank M.Q. for designing this study and reviewing the article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work is supported by the