
Abstract
This review summarizes the experimental evidence that supports the role of dopamine in the regulation of ocular axial growth. The most important functions attributed to dopamine are light adaptation and regulation of the retinal circadian rhythm. An increase of the retinal levels of dopamine activates D1 and D2 dopaminergic receptors present throughout the retina, generating a signal that inhibits axial growth once the eye has reached emmetropization. Researchers induced form-deprivation myopia in animal models in order to assess the different changes of ocular axial growth. Other studies have shown that phenylethylamine is an endogenous precursor-neurotransmitter capable of modulating the activity of dopamine. Considering the role of the dopaminergic system in the development of myopia (in children and adolescents) and the fact that phenylethylamine improves the consequences of a dopamine deficit, it would be interesting to study the effect of phenylethylamine on the regulation of axial growth, which represents the genesis of myopia.
Physiopathology of Ocular Axial Growth
Myopia is a condition in which the eye is too long for the focal length of the cornea and lens, and the plane of sharp focus ends up in front of the retina, causing a refractive error. Consequently, this ametropia changes the quality of life and can be a major risk factor for cataract, glaucoma, retinal detachment, and myopic retinopathy. 1 –4 There is growing evidence that the prevalence of myopia is increasing in many parts of the world, in particular in Taiwan, Japan, Hong Kong, Singapore, and the United States. 2 Unfortunately, myopia has become an important public health issue, evidenced by the fact that it is one of the 5 ocular conditions listed as immediate priorities by the World Health Organization’s Global Initiative for the Elimination of Avoidable Blindness. 2
Many animal species, including humans, are most often hyperopic at birth; in other words, the axial length of the eye, which in adolescents is about 16.8 mm, is too small in relation to the dioptric power and this causes the focus to fall behind the retina. During development, the eye tends to grow in an anteroposterior direction through a vision-dependent process called emmetropization; this allows a correct balance between refractive power and axial length, so that the focus can fall on the retina. Most hyperopic eyes become emmetropic, ranging from +1D to –1D between the first and the second year of life. 1 –3
However, the eye, like the rest of the body, does continue growing until puberty. Thus, once emmetropization is reached, the eye remains vulnerable to the development of myopia for as long as the body grows. 5 When an increase in axial length is not accompanied by a corresponding reduction of refractive power of the ocular diopter, the subject needs to use negative lenses to bring the focus back on the retina.
In humans, certain genes associated with myopia and hyperopia have been identified. 2,6 –8 Various studies underlined the role of excessive near vision (computer, reading, videogames, etc), higher education level, urbanized environments, and diets rich in sugars and cereals 9 –11 —the latter increasing axial length by interacting with the insulin-like growth factor. 11 Outdoor activities seems instead to play a protective role. 12,13 An Australian study, conducted in 51 schools of Sydney, demonstrated that the reduction in the development of myopia is directly correlated to the number of hours spent outdoors. The authors concluded, in fact, that sunlight stimulates the release of dopamine, which inhibits axial growth and, therefore, the development of myopia in children. 12
However, blurred vision brings a signaling cascade that from the retina, through the retinal pigment epithelium, reaches the sclera, altering its growth. Various studies on animal models show that different molecules are involved in the regulation of ocular axial growth: the dopaminergic, muscarinic, and glucagonergic systems. 14 –18
In addition, the existence of go and stop signals that regulate axial growth has been proposed. 19 Stop signals include dopamine, glucagon, and the fibroblast growth factor. 14,18,19 Go signals include instead acetylcholine,17 transforming growth factor β, 19 nitric oxide, 20 and retinoic acid. 21,22 The last 2 mentioned have also been found to have an inhibitory role on axial growth. 23 This double effect is a further confirmation of the hypothesis that there is a very complicated multifactorial mechanism guided by a genetic and environmental interaction. The dopaminergic system plays an important role in the development of myopia although its mechanism is not yet completely clear. This thesis has been confirmed by various studies on animal models in which the authors influenced ocular axial growth by acting on the dopaminergic system.
This review summarizes the experimental evidence that supports the role of dopamine in the regulation of axial growth.
Role of Dopamine
Studies on animal models and on children with neonatal ptosis, cataract, or keratitis associated with myopia 24 –28 allowed us to study the possible causes and the molecular, biological, and cellular mechanisms responsible for an increase in the axial length of the eye.
Researchers induced form-deprivation myopia in animal models in order to study the different changes of axial growth. Indeed, they studied the variations in axial length after removing the eyelid closure and used negative and positive lenses. 29
Observations after unilateral visual deprivation indicate that the quality of the retinal image influences postnatal growth. Vision deprivation in young monkeys, chicks, rabbits, or in children alters the normal growth of the eyeball, leading to an excessive axial length and the eyes becoming myopic. 24 –27,30,31 The relationship between visual quality and eye dimension underlines the role of the central nervous system in growth control. Nevertheless, recent observations suggested that this control is mostly local. In fact, form-deprivation myopia in both monkeys and chickens occurred also after having damaged the optic nerve that connects the retina to the brain. 32 In chicks, a partial occlusion, used to restrict the vision of either the nasal or temporal visual fields, induced an overgrowth of the corresponding retinal portion. For example, the occlusion of the nasal visual field caused an excessive growth of the temporal part of the ocular globe. 33,34
In 1995, Guo 35 demonstrated that after applying negative lenses to the eyes of chickens, the levels of dopamine and of its major metabolite, 3,4-dihydroxyphenylacetic acid, decreased while the axial length increased (Figure 1). Likewise, the application of positive lenses had an opposite effect: dopamine and 3,4-dihydroxyphenylacetic acid levels increased with a subsequent reduction of axial length (Figure 1A, B). This experiment demonstrated how both image quality and focus location, through dopamine, regulate the growth and development of the axial portion of the eye, eventually causing growth to stop once emmetropization is reached.
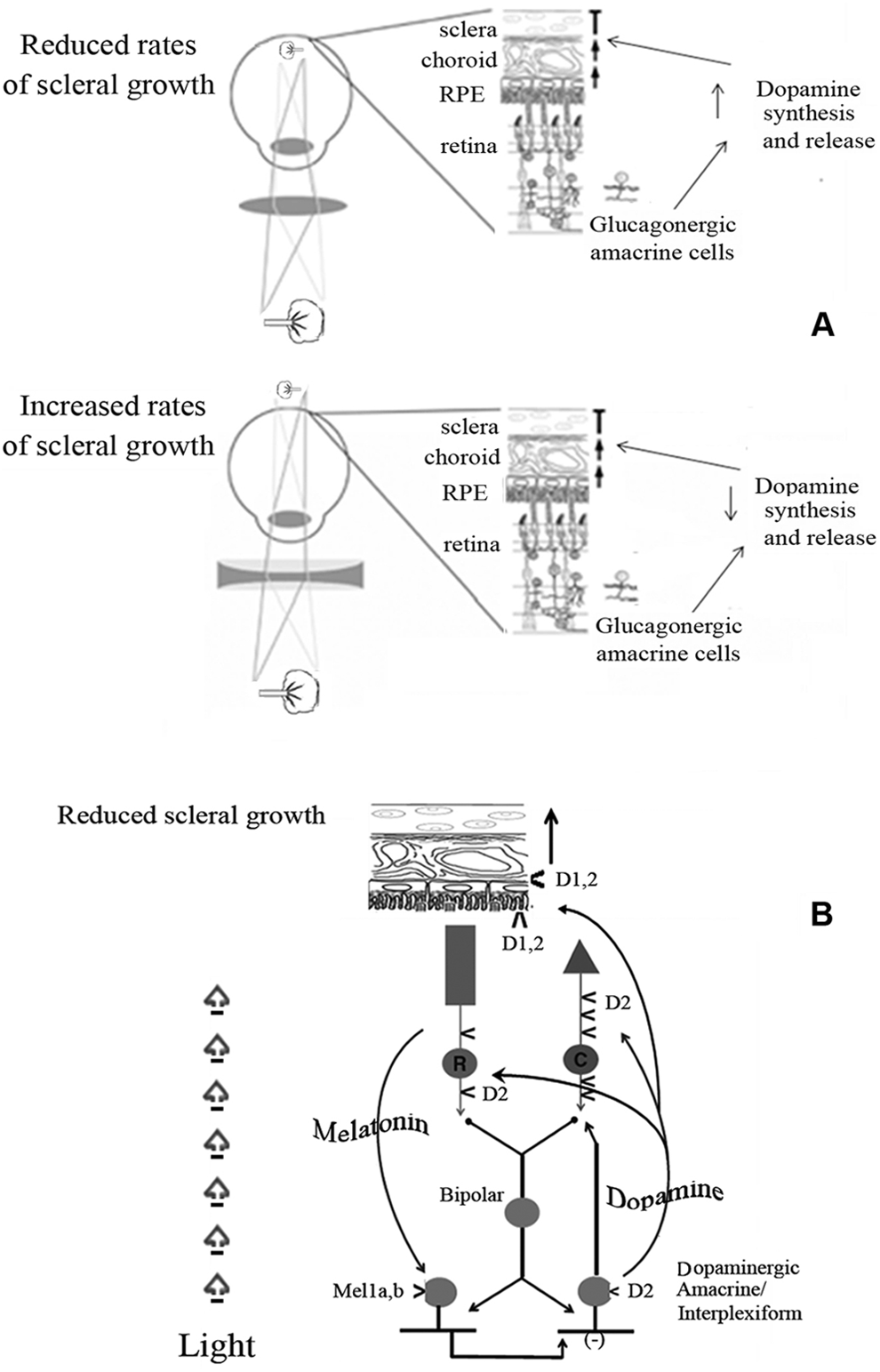
(A) Regulation of ocular axial growth. The application of a positive lens renders the eye hyperopic through an increase of dopamine that inhibits axial growth. The application of a negative lens causes myopia through decrease of dopamine levels and subsequent stimulation of axial growth. (B) Overview of the interactions among light, melatonin, dopamine, and ocular axial growth. Abbreviations: RPE, retinal pigment epithelium; Mel1a,b, melatonin receptor 1a,b; D2, D2-like dopamine receptor; D1, D1-like dopamine receptor.
Other studies, conducted on chickens and monkeys, demonstrated that by inducing form-deprivation myopia, dopamine levels in the retina start to decrease with a consequent increase of the speed of axial growth. 14,15 In contrast, an increase of dopamine concentration leads to a reduction of axial growth, once the occlusion is removed. 35
Dopamine (Figure 2) is synthesized and released by dopaminergic amacrine cells and by interplexiform retinal neuron. 36 Namely, the light-activated neuromodulators dopamine and nitric oxide, which are released by different amacrine cell subtypes, activate a number of intracellular pathways involving cyclic adenosine monophosphate– and cyclic guanosine monophosphate–dependent protein kinases. This results in the phosphorylation or dephosphorylation of gap junction connexins, altering the conductance of the gap junctions to ionic currents. This modulation varies across the population of retinal interneurons such that the conductance of some gap junctions is increased by light whereas that of others is decreased. In fact, light can increase or decrease the conductance of gap junctions in the same neuron, depending on the level of brightness. 37
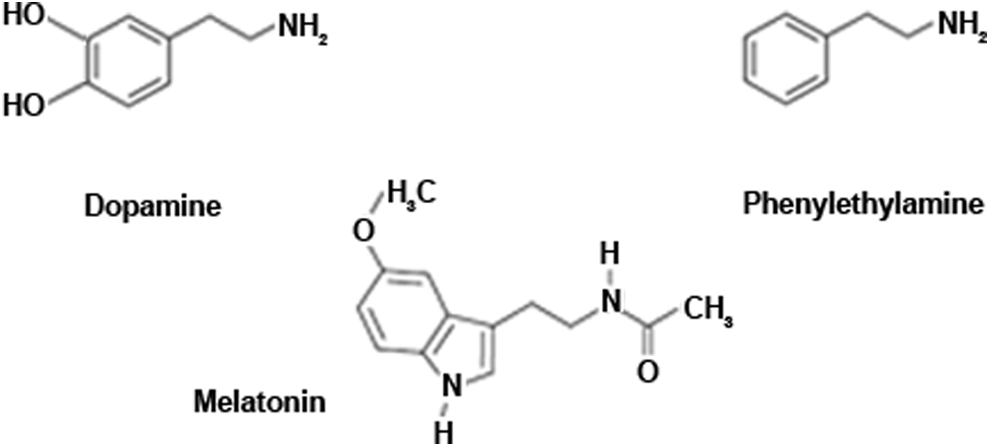
Chemical structure of dopamine, melatonin, phenylethylamine.
Furthermore, as mentioned above, several studies underlined how dopamine may affect the regulation of axial growth but before analyzing the metabolic pathways that regulate ocular axial growth, we need to mention dopamine receptors.
Dopamine Receptors
Dopamine binds to 5 different kinds of receptors (D1-D5) that belong to the class of G protein–coupled receptors and are composed of 7 transmembrane domains. 38
Dopamine receptors are grouped into 2 different families: D1-like receptors, which include D1 and D5, and D2-like receptors, which include D2, D3, and D4. 39 The main difference between these 2 families is the action of the G protein: in the D1-like family, we find the Gs protein, whose activation leads to an increase of cyclic adenosine monophosphate mediated by the action of adenylate cyclase. On the other hand, the activation of the D2-like receptors connected to a Gi protein causes inhibition of the adenylate cyclase and reduction of the cyclic adenosine monophosphate. 38,39
The use of autoradiography, immunocytochemistry, and specific antibodies has allowed us to identify the dopamine receptors present in the retina and their location. 40 These studies discovered that D5 and D2 41,42 receptors are located in the retinal pigment epithelium. Besides D2-like receptors similar to D4 found in mice are located both in rods and cones, although they have not been sufficiently characterized in other animals. 43,44 Bipolar neurons and horizontal, amacrine, and ganglion cells have D1 receptors. 45 Furthermore, the dopaminergic neuron presents the D2 receptor that, acting as an autoreceptor, inhibits the release of dopamine. 46 Dopamine also acts on the retinal glial cells of some vertebrates and on the pericytes of the retinal blood vessels of mammals. 47,48
The retinal dopamine concentration is never uniform because dopamine is produced according to a circadian rhythm, and its release from the synaptic vesicles increases during the daytime and decreases during the night. 49 The different degree of local diffusion and the variable distance of the dopaminergic cell receptors from the dopaminergic neurons influences the extracellular concentration of dopamine in the retina, and this correlates with the different sensibility of the various dopamine receptors. The D5 receptors have 10 or 20 times more affinity for dopamine than the D1 receptors, 50 which is needed because the D5 receptors are located in the retinal pigment epithelium, which is the retinal portion that is furthest from the dopaminergic neurons and therefore has a lower concentration of dopamine. Likewise, the greater sensitivity of D2 compared to D1 receptors allows dopamine to bind to them during the night, when its concentration decreases because of the inhibiting effect of melatonin (Figure 2). Various studies have shown that dopamine receptors are down- or upregulated in relation to the increase or decrease of dopamine concentration. 51,52
Biosynthesis of Dopamine
The daily rhythm of synthesis and release of dopamine depends on the interaction between retinal photoreceptors and dopaminergic neurons. Dopamine itself inhibits the synthesis of melatonin by binding with the photoreceptor D2-like receptors, whereas melatonin inhibits the synthesis and release of dopamine. 53,39 The result is represented by 2 shifted rhythms: dopamine reaches its peak during the day, whereas melatonin reaches it during the night.
In the retina, melatonin acts locally and plays an important role in the control of circadian rhythms. 49 Furthermore, it demonstrated that melatonin is a strong inhibitor of dopamine release and metabolism. Melatonin affects the eye in 2 different ways, as an antioxidant and through the G protein–coupled receptors. 54 –56 The antioxidant role has not been thoroughly studied although Marchiafava and Longoni 57 reported the protective role of prevention against the oxidation of a dye injected in the photoreceptors of a frog, and therefore assumed a protective action also against the oxidative effects of light. There are 3 different forms of the melatonin receptor Mel1, and in the retina of mammals, the melatonin receptor Mel1a,b subtypes are found. 54,56 –59 Fujieda et al 58 have identified the presence of the Mel1 receptor on GABAergic and dopaminergic amacrine cells.
In general, when melatonin binds to Mel1 Gi protein–coupled receptors, the concentration of cyclic adenosine monophosphate decreases because of the inhibition of adenylate cyclase. In the retina and mainly in the inner plexiform layer, melatonin binds to the Mel1b Gi protein–coupled receptor, 58 but their exact function in the retina is still unknown.
Dubocovich et al 59 have reported a high sensitivity of these receptors for melatonin; in their study, a melatonin concentration of 20 pM reduced by 50% the release of [3H] dopamine, and this is tightly correlated with the high affinity of Mel1b receptor for the 2-[I125] iodomelatonin.
The biosynthesis of melatonin is regulated by a light and dark cycle. In conditions of light, the acetylating enzyme that converts serotonin into N-acetyl-serotonin, precursor of melatonin, is inactivated and the concentrations of melatonin decrease. 60 A corresponding increase in the concentration of dopamine has been observed, suggesting the possibility to assume that the increase or reduction of these 2 hormones represents the base of the ocular circadian rhythms: darkness disables the dopaminergic transmission and determines the activation of melatonin synthesis.
Interactions between Dopamine and Retina
Ions and metabolites need to transit between the subretinal space and the choroid through a transport mechanism, because cells are sealed together by tight junctions and zonulae adherentes. For some retinal neurons, dopamine binds to the D1 receptors, activating adenylate cyclase and increasing the concentration of cyclic adenosine monophosphate. This in turn activates cyclic adenosine monophosphate–dependent protein kinase, phosphorylation of connexins, and a reduction in the conductance of gap junctions. D1 receptor mechanism occurs at gap junctions between horizontal cells, between AII amacrine cells (subtype of retinal amacrine cell with a small dendritic field that conveys the rod signal to cone bipolar cells), between other amacrine cell subtypes and at the amacrine cell hemichannel of amacrine cell–ganglion cell gap junctions. Dopamine also binds to D2/4 receptors, which reduces the activity of adenylate cyclase, resulting in a reduction of cyclic adenosine monophosphate levels and thereby reducing the activity of protein kinase, resulting in increased gap junction conductance. This mechanism occurs at gap junctions between rods and cones, between ganglion cells, and at the ganglion cell hemichannel of ganglion cell–amacrine cell gap junctions. 37 The specific effects of dopamine on the retinal pigment epithelium are not completely clear, but they seem to be related with the cyclic adenosine monophosphate concentration and the anion conductance across the retinal pigment epithelium. 61,62
Furthermore, it still is not clear whether the onset of myopia depends on a decrease in dopamine or if dopamine itself may generate, through its receptors, a molecular mechanism that leads to alterations of scleral growth or, even, if both factors may act together. The first evidence in favor of the receptor hypothesis is given by the fact that the administration of apomorphine, a nonspecific dopamine agonist, contrasts form-deprivation myopia. 63 Some authors stated that the ocular growth inhibition effected by the nonspecific dopamine agonist apomorphine and the D2 receptor agonist quinpirole is associated with transient increases in choroid thickness, whereas the relatively ineffective D1 agonist does not elicit the choroidal response. 64
Although a number of studies have shown that the inhibitory effects of dopamine agonists on form-deprivation myopia and lens-induced myopia are mediated through stimulation of the D2 receptor, there is also recent evidence that the balance of D2- and D1-receptor activation is important. Inhibition of D2 receptors can also slow the development of spontaneous myopia in albino guinea pigs. 40 Moreover, Seko et al 65 have demonstrated that retinal pigment epithelial cells of chicks significantly increase their mitotic rate when they are added to a culture with scleral chondrocytes. Adding apomorphine to the culture system causes a decrease of the retinal pigment epithelium–dependent stimulation of scleral chondrocytes, whereas apomorphine alone, without retinal pigment epithelial cells, has less effect on the mitosis of scleral cells.
Different Molecular Mechanisms for Various Forms of Myopia
Despite what has been said above, there are still many doubts about the role of dopamine in ocular growth, because some studies do not support this hypothesis. The antagonist of dopamine receptors should cause an increase in ocular axial growth, which would lead to myopia. Nevertheless, certain authors have found an absent or weak effect on ocular growth and on the refractive error.
63
The destruction of dopaminergic neurons with 6-hydroxydopamine prevents form-deprivation myopia instead of causing it.
66
The depletion of retinal dopamine caused by the assumption of reserpine, a blocker of the vesicular monoamine transporter, does not cause myopia.
67
These statements must be interpreted and analyzed in light of a different dopaminergic mechanism that is at the base of form-deprivation myopia and negative lens–induced myopia. In fact, 6-hydroxydopamine does not alter the response of axial length to negative lenses, but as mentioned above suppresses form-deprivation myopia. 68
In 2004, Schmid reported that atropine on its own has, in form-deprivation myopia but not in negative lens–induced myopia, an inhibitory effect that is greater than the effect obtained by the combination of atropine and apomorphine. 69 This suggests that in form-deprivation myopia, the muscarinic antagonists and dopaminergic agonists act on the same metabolic pathway and if coadministered tend to have competitive effects. Constant illumination, which alters the levels of dopamine, inhibits form-deprivation myopia but does not have effects on the axial growth of the eye induced by negative lenses. 70 The section of the optic nerve prevents myopia induced by negative lenses, but not form-deprivation myopia. 71 Furthermore, another interesting observation is that the speed of ocular axial growth is greater in negative lens–induced myopia than in form-deprivation myopia. 72 Therefore, the results of these studies support the hypothesis of different dopaminergic mechanisms at the base of form-deprivation myopia and of negative lens–induced myopia.
Nickla et al 64 suggested that the D2 receptor has an important role in form-deprivation myopia whereas both D1 and D2 receptors are responsible for the negative lens–induced myopia.
Discussion
Therefore, the dopaminergic system plays an important role in the development of myopia in children and adolescents, although its mechanism is not yet completely clear. This view has been confirmed by various studies on animal models in which the authors influenced ocular axial growth by acting on the dopaminergic system.
Schmid and Wildsoet showed how intravitreal injections of apomorphine prevent form-deprivation myopia in chickens and monkeys and the negative lens–induced myopia in chickens. 69
Recent studies demonstrated that exogenous dopamine and its precursor levodopa have similar effects in preventing form-deprivation myopia in rabbits and guinea pigs, respectively. 73,74
Finally, myopia induced by image alteration is directly proportional to the reduction of dopamine in chickens. 75 All these studies could let us assume that an increase of dopamine, especially in the age of emmetropization, may prevent myopia by affecting axial growth.
It has been shown that phenylethylamine (Figure 2) is an endogenous precursor-neurotransmitter capable of modulating the activity of dopamine and other neurotransmitters. 76,77
In brain capillary endothelium and catecholaminergic terminals, a single decarboxylation step effected by aromatic amino acid decarboxylase converts phenylalanine to phenylethylamine, at a rate comparable to that of the central synthesis of dopamine. Phenylethylamine, however, is not stored in intraneuronal vesicles and has difficulty maintaining high concentrations in the human body because of its rapidly oxidative deamination to phenylacetic acid by the enzyme B monoamine oxidase. 78,79
The molecular mechanism of the trace amines involves binding to novel G protein–coupled receptors, called TA1 and TA2 (trace amine-associated receptor family).
TA1 binds to both β-phenylethylamine and tyramine with high affinity and produces cyclic adenosine monophosphate in response to this binding, whereas the related TA2 receptor appears to be specific for β-phenylethylamine and tryptamine. Both of these G protein–coupled receptor families possess many of the structural hallmarks of the β-adrenergic/rhodopsin receptor superfamily, found mainly in the rod cells of the human retina. 79 –82
The molecular mechanism of phenylethylamine is not yet clear, but it is believed that binding of phenylethylamine to the TA1 receptor results in an alteration of the monoamine transporter functions, which leads to inhibition of the reuptake of dopamine, serotonin, and norepinephrine. 79 This results in an increase in dopamine freely circulating in the presynaptic terminals and a greater concentration of diffuse dopamine in synaptic gaps, thus producing an enhancement of dopaminergic transmission. Substandard phenylethylamine levels have been linked to disorders such as attention deficit and depression. 77,80 –82 In myopia, a dysfunction of the dopaminergic system has been demonstrated. 83,84
Considering the results of the Australian study conducted on approximately 4000 children and according to the studies on experimental animal models presented in this review, we can say that the exposure to sun and light induces the retina to produce more dopamine and therefore reduce the development of myopia in children. 12
Taking into account the role of the dopaminergic system in the development of myopia and the fact that phenylethylamine improves the consequences of dopamine deficit in other diseases, it would be interesting to study the effect of phenylethylamine on the regulation of axial growth, which represents the genesis of myopia.
Trace amines, including phenylethylamine, are constituents of many foods, including chocolate, blue-green algae, cheeses, and wines and are generated by so-called friendly bacteria such as Lactobacillus, Lactococcus, and Enterococcus species, which are found in probiotics. The best known is cocoa but, becausefor an effective quantity of phenylethylamine, people will need to eat a lot of chocolate, that is not considered an easily exploitable source. 84 –86 Phenylethylamine can be found in members of the family Leguminoseae, which is the second-largest family of seed plants and is composed of trees, shrubs, vines, herbs (such as clover), and vegetables (such as beans and peas). The various species found within this family have been used as food, green manure, and for medicinal purposes. 84 –87
Dietary supplementation of phenylethylamine is instead more effective with food such as the Klamath algae. The Klamath algae are also the only food known to date that contains significant amounts of phenylethylamine. It contributes directly to increasing the quantity of freely circulating dopamine, thus increasing dopaminergic transmission. 86 –88
Klamath algae and the family of blue-green microalgae are cyanobacteria that contain large amounts of various substances. Cyanobacteria were born about 4 billion years ago and are considered a compact food from which life emerged on earth. 86 –88
Unlike other microalgae, which are grown in artificial ponds with the addition of fertilizers, the Klamath algae are the only ones to arise spontaneously in a pristine environment and are rich in minerals. Thanks to ideal conditions in which they grow wild, the Klamath algae have a unique endowment of minerals, vitamins, amino acids, antioxidants, phycocyanin, phenylethylamine, and essential fatty acids. The nutritional profile of Klamath algae is superior to traditional vitamin supplements because the wall of the cyanobacteria is constituted by a simple layer of peptidoglycan that allows the passage of all the essential nutrients, making them absorbable by the body. Phycocyanin is the blue pigment present in all the blue-green microalgae and has an extraordinary nutraceutical power. In fact, phycocyanin has a significant antioxidant, anti-inflammatory, hepatoprotective, and radical-scavenging properties. 87
Some studies confirm the benefits of the use of Klamath algae in children. In a Nicaraguan study involving 1482 children, the administration of 1 g/d for 12 months to school-age children gave interesting results on attention and academic performance without showing any side effects. 88 In this review, we have documented that a dysfunction of the dopaminergic system plays a role in the pathogenesis of myopia (83-85). In addition, WellmixAFA (not commercial name, produced by EyePharma Company, Italy) is a substance designed specifically to take advantage of the dopaminergic characteristics of natural compounds that mimic the dopaminergic function induced by the sun helping to counter the progression of myopia.
Conclusion
In summary, further studies will be required to investigate the complicated biochemical and molecular mechanisms related to genetic and nongenetic myopia in eyes predisposed to this refractive defect. Moreover, we should not underestimate the multifactorial aspects that may be involved in the development of this abnormal condition in children and adolescents.
In the meantime, it would be useful to assure to children and adolescents a proper nutritional approach. At a time when consumption of junk food devoid of nutrients is spreading, it may be important to use natural supplements that are able to give full support.
Footnotes
Acknowledgment
The authors express their deep gratitude to Dr. F. Lenarduzzi for his help with the English.
Author Contributions
MN and NP were responsible for the design, conceptualization, and interpretation of the data. AMP and BP drafted and revised the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.