
Abstract
Engineered anisotropic tissue constructs containing aligned cell and extracellular matrix structures are useful as in vitro models and for regenerative medicine. They are of particular interest for nervous system modelling and regeneration, where tracts of aligned neurons and glia are required. The self-alignment of cells and matrix due to tension within tethered collagen gels is a useful tool for generating anisotropic tissues, but requires an optimal balance between cell density, matrix concentration and time to be achieved for each specific cell type. The aim of this study was to develop an assay system based on contraction of free-floating cellular gels in 96-well plates that could be used to investigate cell–matrix interactions and to establish optimal parameters for subsequent self-alignment of cells in tethered gels. Using C6 glioma cells, the relationship between contraction and alignment was established, with 60–80% contraction in the 96-well plate assay corresponding to alignment throughout tethered gels made using the same parameters. The assay system was used to investigate the effect of C6 cell density, collagen concentration and time. It was also used to show that blocking α1 integrin reduced the contraction and self-alignment of these cells, whereas blocking α2 integrin had little effect. The approach was validated by using primary astrocytes in the assay system under culture conditions that modified their ability to contract collagen gels. This detailed investigation describes a robust assay for optimising cellular self-alignment and provides a useful reference framework for future development of self-aligned artificial tissue.
Introduction
Designing engineered tissues frequently requires the organisation of cells and extracellular matrix (ECM) into stable anisotropic structures. Cell-populated collagen hydrogels are widely used in tissue engineering since their composition recapitulates many of the key features of natural tissues; additionally, their mechanical compliance and dynamic cell–matrix interactions can be advantageous in the construction of robust organised structures.1,2 Research largely focussed on the study of fibroblast-populated collagen lattices has revealed the close association between the ECM and cellular behaviour within collagen gels (reviewed by Brown 3 ). Harnessing the forces generated by cells within such systems provides a potential means of creating engineered tissues that can be used as model systems4–7 or within regenerative medicine where anisotropic constructs are required.4,8 Furthermore, our recent studies have shown that subsequent stabilisation of organised cellular collagen gels produces robust artificial tissues with highly aligned cell and ECM structures.9–12
The process of cellular self-alignment within collagen gels involves cells exerting forces that contract and deform the surrounding matrix. By casting the cellular gel within a mould that tethers each end to a fixed point, 2 cell-mediated contraction of the gel is restricted, and tension builds up along the longitudinal axis of the gel. Subsequent analysis shows that both cells and ECM fibrils align parallel to this axis of tension, and where there are regions with no single axis of tension the cells are orientated randomly.4,5
In order to exploit fully this phenomenon to construct anisotropic cellular gels for various applications in research and regeneration, it is important to establish a better understanding of the parameters that influence cellular self-alignment in collagen gels. Of particular interest is the further exploration of the behaviour of nervous system cells within an aligned stabilised model, since the great majority of previous work in this area has focussed on musculoskeletal cells (predominantly fibroblasts).3,6,7,13,14 Organising neural cells into aligned 3D tissues has obvious applications in regenerative medicine, where aligned tracts of glia can potentially be used to support and guide neurons to bridge gaps in peripheral nerves or replace tracts of CNS tissue. Furthermore, organising 3D cultures of neural cells into aligned constructs provides considerable advantages in terms of ease of monitoring and control of complexity when co-cultures are used as in vitro models in research. Additionally, this model system is also likely to be applicable to other areas such as musculoskeletal, cardiac tissue and engineering of the cornea. The initial aim here therefore was to explore the key parameters that influence self-alignment in collagen gels of a rat CNS glial cell line (C6) that is widely used in neuroscience research. Further to this, because different cell types vary in their ability to contract collagen gels, 15 we aimed to establish an assay system that could be applied more widely to determine optimal conditions required for self-alignment.
For successful generation of an anisotropic tract of cellular collagen, sufficient tension is required, which is dependent on factors such as the amount of force generated by the cellular component, the compliance of the ECM component, and the extent and nature of interactions between cells and ECM. Initial experiments confirmed the integrin-mediated nature of the glial cell–ECM interaction that is central to the process; then the influence of cell seeding density, collagen concentration and time on contraction of unconstrained gels and alignment of glial cells within tethered gels were explored in detail.
Materials and methods
Fabrication of collagen gels
C6 rat glioma cells (ATCC®CCL-107™), were maintained in culture medium (Ham’s F12, Gibco) supplemented with penicillin and streptomycin (100 U/ml and 100 mg/ml, respectively; Sigma) and 10% v/v foetal calf serum (Sigma) in standard cell culture flasks. Primary astrocytes were obtained from the cortices of P2 rat pups using previously reported protocols, 16 and maintained in culture medium (DMEM, Sigma) also supplemented with penicillin and streptomycin and foetal calf serum in poly-D-lysine coated cell culture flasks. All gels were prepared using 80% v/v Type I rat tail collagen (2 mg/ml in 0.6% acetic acid; First Link, UK) unless otherwise stated, mixed with 10% v/v 10 × minimum essential medium (Sigma) and the mixture neutralised using 5.8% v/v neutralising solution (TAP Biosystems) before addition to 4.2% v/v cell suspension (culture medium containing cell number appropriate for achieving the required final cell density). For multi-well plate assays, 75 µl or 400 µl of the mixture was added to individual wells of 96- or 24-well plates, respectively. For aligned gels, 1 ml of this mixture was added to a rectangular stainless steel mould and integrated with tethering mesh at opposite ends as described previously,2,17 and all gels were allowed to set at 37℃ for 10 min. Cellular gels were immersed in culture medium and incubated at 37℃ for a further 15 min before detachment from well plates where appropriate (termed free-floating gels) and incubated at 37℃ in a humidified incubator with 5% CO2/95% air for up to 24 h. In some experiments, the initial collagen concentration was altered. In those cases, Type I rat tail collagen (5 mg/ml in 0.6% acetic acid; First Link, UK) was diluted in 0.6% acetic acid to the required concentration prior to neutralisation.
To investigate the role of specific integrins in the cell–ECM interaction, anti-CD49a antibody, anti-CD49b antibody (BD Biosciences) or IgG control was diluted in Ham’s F12 and added to both the cell suspension used to make the gel and to the culture medium used to cover the gel, to give the desired final concentration (0.01–10 µg/ml).
Measurement of contraction of cellular gels
At the end of the defined period in culture, media was removed from gels in the multi-well plates and images captured of each gel. Percentage contraction was calculated by comparing the area of the well occupied by each gel to the area of the well (Image J).
Measurement of cellular alignment within gels
To assess alignment of C6 cells, haematoxylin and eosin (H & E) were used to stain cellular gels from the tethered aligned system. Following overnight fixation in 4% paraformaldehyde, the fixative was aspirated and samples washed with water, and then stained in Harris’s haematoxylin for 5 min. Gels were washed in water for 2 min and differentiated in 0.5% acid alcohol for 1 min. Gels were washed again in water for a further 2 min, and then stained in 1% Eosin-Pink for 1 min. The stained gels were rinsed once more in water. Replicate images were captured from three areas within defined regions of each gel under bright-field illumination (20 × lens, Olympus BX61 microscope). Cell processes were traced using OpenLab software (Perkin Elmer) to assess the angle of deviation of each cell process compared to the long axis of the gel.
Statistical analysis
Effects of blocking specific integrin subunits on cellular gel contraction were analysed using one-way ANOVA with Dunnett’s post test to compare each concentration to the 0 µg/ml control. The effect of blocking integrin α1 subunit on cellular alignment was analysed using a Mann-Whitney test to compare equivalent regions in the presence or absence of anti-CD49a antibody. Assessment of cellular alignment was conducted using a Kruskal-Wallis test with Dunn’s post-test to compare middle and side populations to delta zones.
Results
The effect of blocking α1 and α2 integrin subunits on contraction of cellular collagen gels
In order to investigate the cell–ECM interactions involved in the contraction of collagen gels by C6 cells, the effect of using blocking antibodies selective for α1 integrin (CD49a) and α2 integrin (CD49b) were examined separately in 96-well plate contraction assays (Figure 1). Following 24 h incubation, the presence of anti-CD49a antibody reduced the contraction in a dose-dependent manner with significantly lower contraction compared to untreated control (51.9 ± 5.9%) in the presence of antibody at concentrations of 0.1 µg/ml and above (Figure 1). Maximal effect was observed with anti-CD49a antibody concentrations of 1 and 10 µg/ml (12.0 ± 1.6% and 11.2 ± 1.2%, respectively) suggesting saturation. The presence of a rabbit IgG control (1 µg/ml) had no significant effect on cell-mediated contraction. Incubation with anti-CD49b antibody had little effect with concentrations of 0.01–1 µg/ml not significantly different to the contraction in untreated control gels (49.4 ± 2.8%; Figure 1). Only a concentration of 10 µg/ml anti-CD49b antibody reduced contraction significantly (to 11.2 ± 1.6%).
The effect of blocking integrin α subunits on contraction of cellular collagen gels. Free-floating gels containing C6 cells at a density of 4 × 106 cells/ml were incubated in the presence or absence of different concentrations of anti-CD49a antibody or anti-CD49b antibody for 24 h then % contraction was assessed. A dose–response curve was calculated for anti CD49a using nonlinear regression (GraphPad Prism software), IC50 approximately 0.3 µg/ml. Data are means ± SEM, n = 10 gels per condition. *P < 0.05, ***P < 0.001 for anti-CD49a, ###P < 0.001 for anti-CD49b; One-way ANOVA with Dunnett’s post test to compare each concentration to the 0 µg/ml control which was 51.9 ± 5.9% and 49.4 ± 2.7% for the anti-CD49a and anti-CD49b experiments, respectively. Control treatment with 1 µg/ml rabbit IgG mean ± SEM; 50.2 ± 3.4%.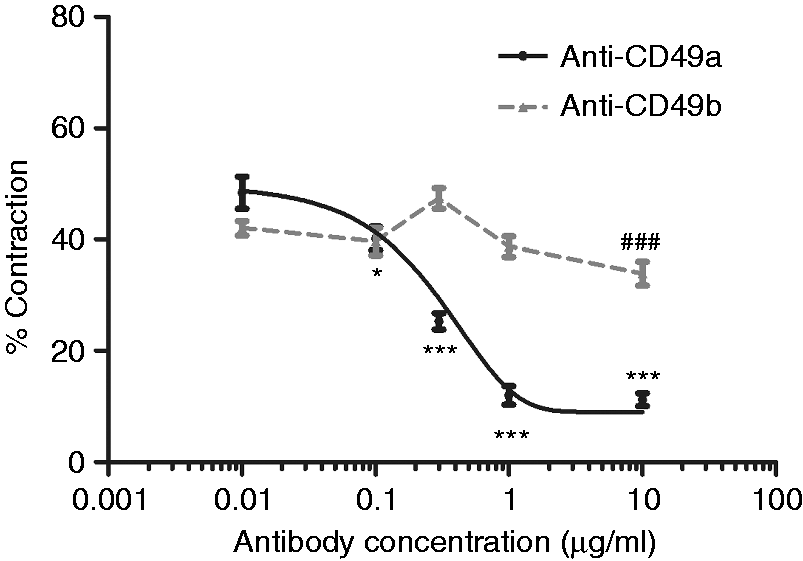
Blocking integrin α1 subunit inhibits cellular alignment within tethered collagen gels
Following the assessment of the effect of anti-CD49a and anti-CD49b on contraction of free-floating gels in the 96-well plate assay, the effect on cellular alignment of 1 µg/ml of anti-CD49a antibody was tested. After 24 h, the control gel with no anti-CD49a adopted the characteristic shape associated with the generation of tension, narrowing by 38% in the centre (Figure 2(a)). In contrast, an equivalent gel incubated with anti-CD49a showed little change in shape (∼15% narrowing; Figure 2(b)).
The effect of anti-CD49a antibody on cellular self-alignment. Tethered collagen gels containing 4 × 106 C6 cells/ml were incubated for 24 h in the absence (a) or presence (b) of 1 µg/ml anti-CD94a antibody. The width of the gel was measured to obtain % change in width in each case. Gels were fixed and cells stained with H & E to enable the angle of deviation of cellular processes relative to the long axis of the construct to be determined in the middle and side regions (c). The drawing illustrates the position of the regions that were sampled. Boxes show interquartile range and median values, whiskers indicate maximum and minimum angles. A Mann-Whitney test was used to compare equivalent regions in the presence or absence of anti-CD49a antibody (***P < 0.001, n = approx 72 cell processes from three gels).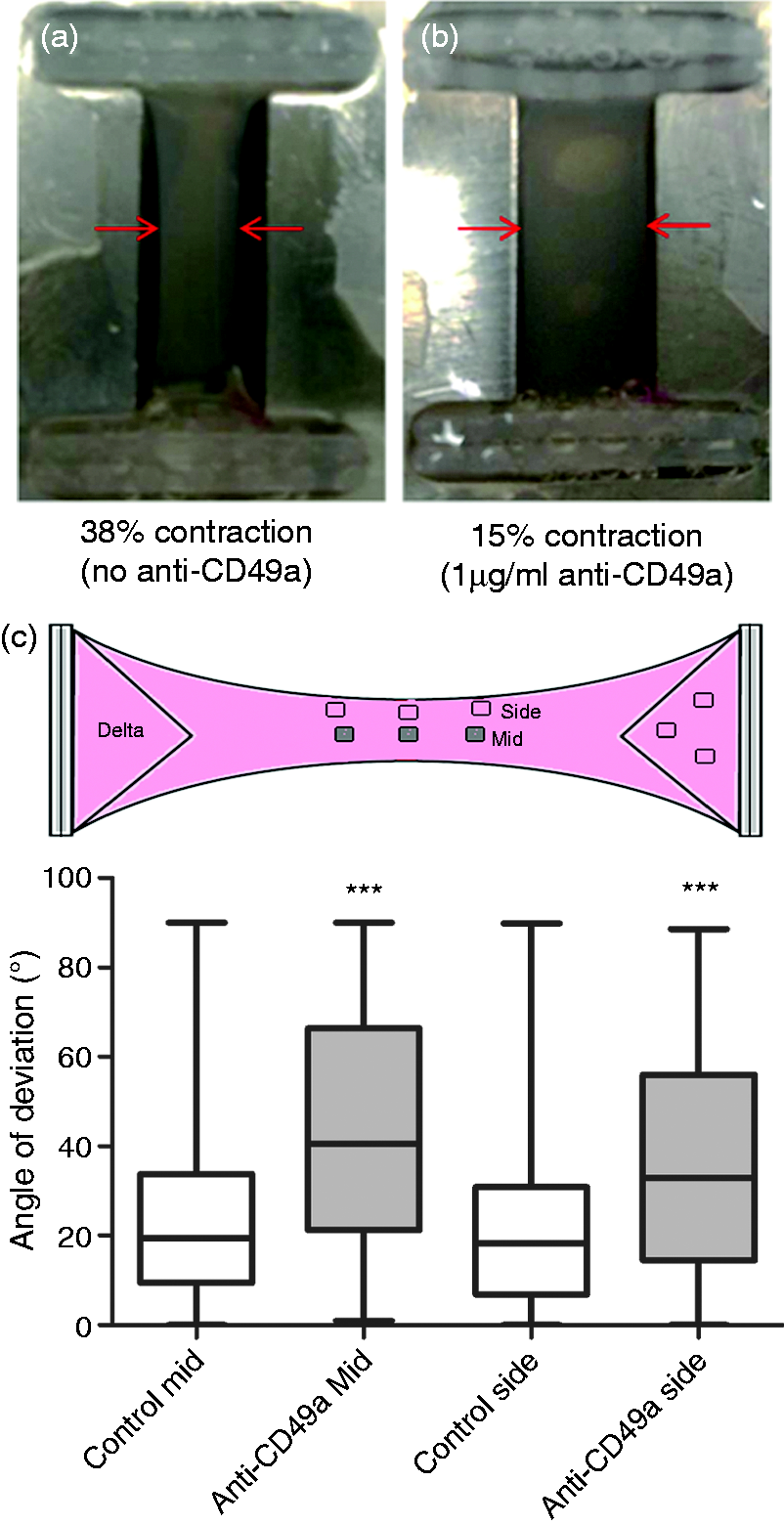
The alignment of C6 cells within the tethered gels was assessed using image analysis to determine the angle by which the principal axis of each cell process deviated from the longitudinal axis of the gel. Specific fields within the gels were assessed based on previous studies that showed how both the ‘middle’ and ‘side’ regions of tethered gels contain aligned cells after 24 h. 4 Anti-CD49a (1 µg/ml) reduced the alignment of cells in both the middle and side regions of the tethered gel (Figure 2(c)).
Using contraction assays to optimise parameters for cellular self-alignment
When a previously untested cell type is used in a tethered collagen gel system it can be time-consuming and costly to determine the optimal cell seeding density required to generate sufficient tension for self-alignment. Using C6 cells as an exemplar, a simple multi-well plate assay was therefore used to quantify contraction over a range of cell densities, in order to relate this to the extent of cellular alignment generated in tethered aligned gels of equivalent cell density. Figure 3(a) shows an example of free-floating gels with different cell seeding densities in a 96-well plate after 24 h contraction. Contraction increased with increasing cell density up to 3 × 106 cells/ml before reaching a plateau at ∼70% (Figure 3(b)).
Contraction assay for varying seeding densities of C6 cells. Typical photograph showing free-floating gels containing C6 cells at various seeding densities after 24 h in culture (a). Images were processed to calculate % contraction, enabling a contraction profile to be constructed (b). Data are mean ± SEM, n = 10 gels per condition from a total of three experimental gel batches.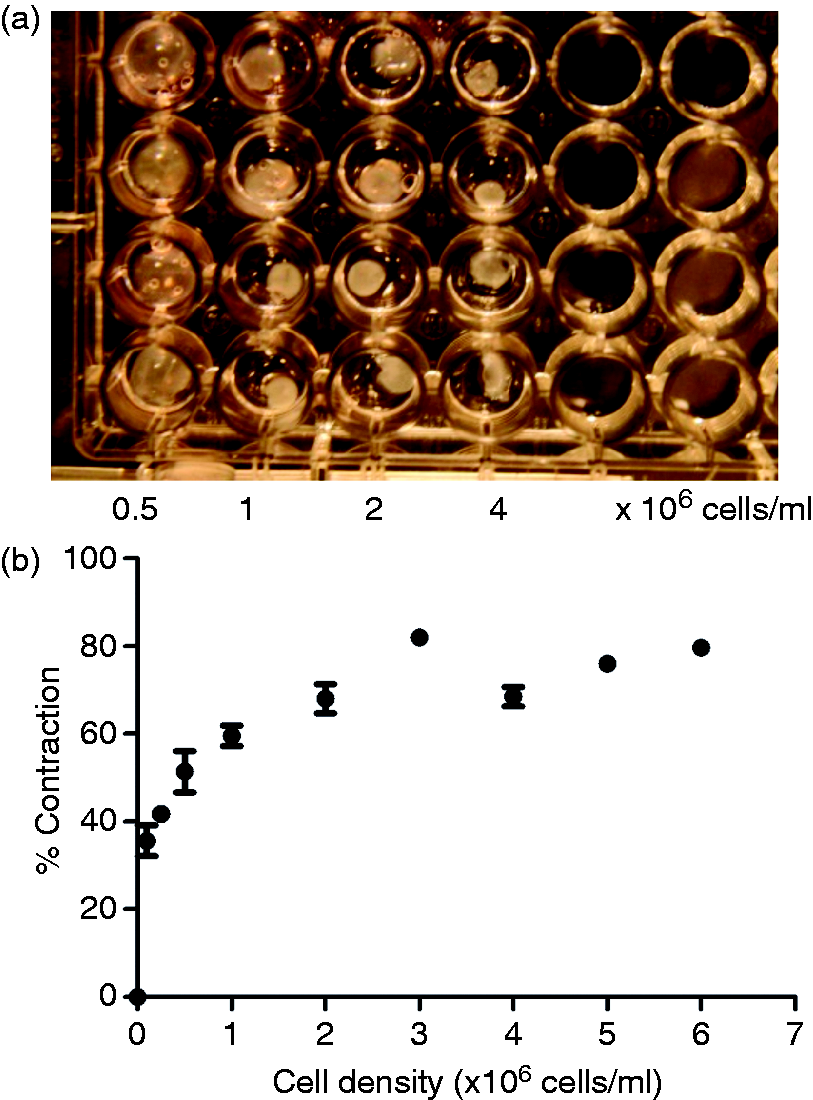
The relationship between % contraction of free-floating gels in the multi-well plate assay and the extent of cellular self-alignment achieved using the equivalent seeding density in tethered gels was investigated. Images of specific regions from each gel were used to determine the pattern of cellular alignment in each case (Figure 4). The three gel regions analysed (Figure 2(c)) were selected based on previous work showing that distinct alignment can be detected in each, with cells in the side regions more highly aligned than the middle and those in the delta zones randomly orientated.
4
The gels with lower cell densities (0.5 and 1 × 106 cells/ml) only showed a significant difference in alignment between the side regions and delta zones, whereas gels with higher cell densities (2 × 106 to 4 × 106 cells/ml) showed a significant difference in both middle and side regions compared to delta zones, indicating cellular alignment throughout the central part of the gel. Table 1 shows the mean angle of deviation from the longitudinal axis of gel for the cell processes in each region. Within the side regions and the middle regions from the higher cell density gels, a smaller mean angle of deviation can be noted in comparison to the delta zones and the middle regions from the lower cell density gels. The C6 cell seeding density that resulted in significant cellular self-alignment throughout the middle and side regions corresponded to the seeding densities that produced >60% contraction in the multi-well plate assay.
Assessment of cellular self-alignment in tethered gels with varying seeding densities of C6 cells. Micrographs showing typical H & E staining of gels containing 4 × 106 C6 glioma cells/ml in the three different areas of the tethered aligned system after 24 h (a). Tethered gels containing C6 cells at 0.5 × 106 to 4 × 106 cells/ml were maintained for 24 h then fixed and stained to enable cellular alignment to be assessed. The angle by which the orientation of cell processes deviated from the longitudinal axis of the gel was measured for approx 30 cells in 3 fields within delta, middle and side regions of gels (b). Boxes show interquartile range and median values, whiskers indicate maximum and minimum angles. Middle and side populations were compared to delta zones using a Kruskal-Wallis test with Dunn’s post-test, ***P < 0.001, n = 3 gels. Summary of cellular self-alignment in tethered gels with varying seeding densities of C6 cells. Note: Mean angle data ± SEM were calculated from the cell populations shown in Figure 4. Middle and side populations were compared to delta zones using a Kruskal-Wallis test with Dunn’s post-test. ns: not significant. P < 0.001, n = 3 gels. The contraction profile result for each equivalent cell density (Figure 3(b)) is also shown.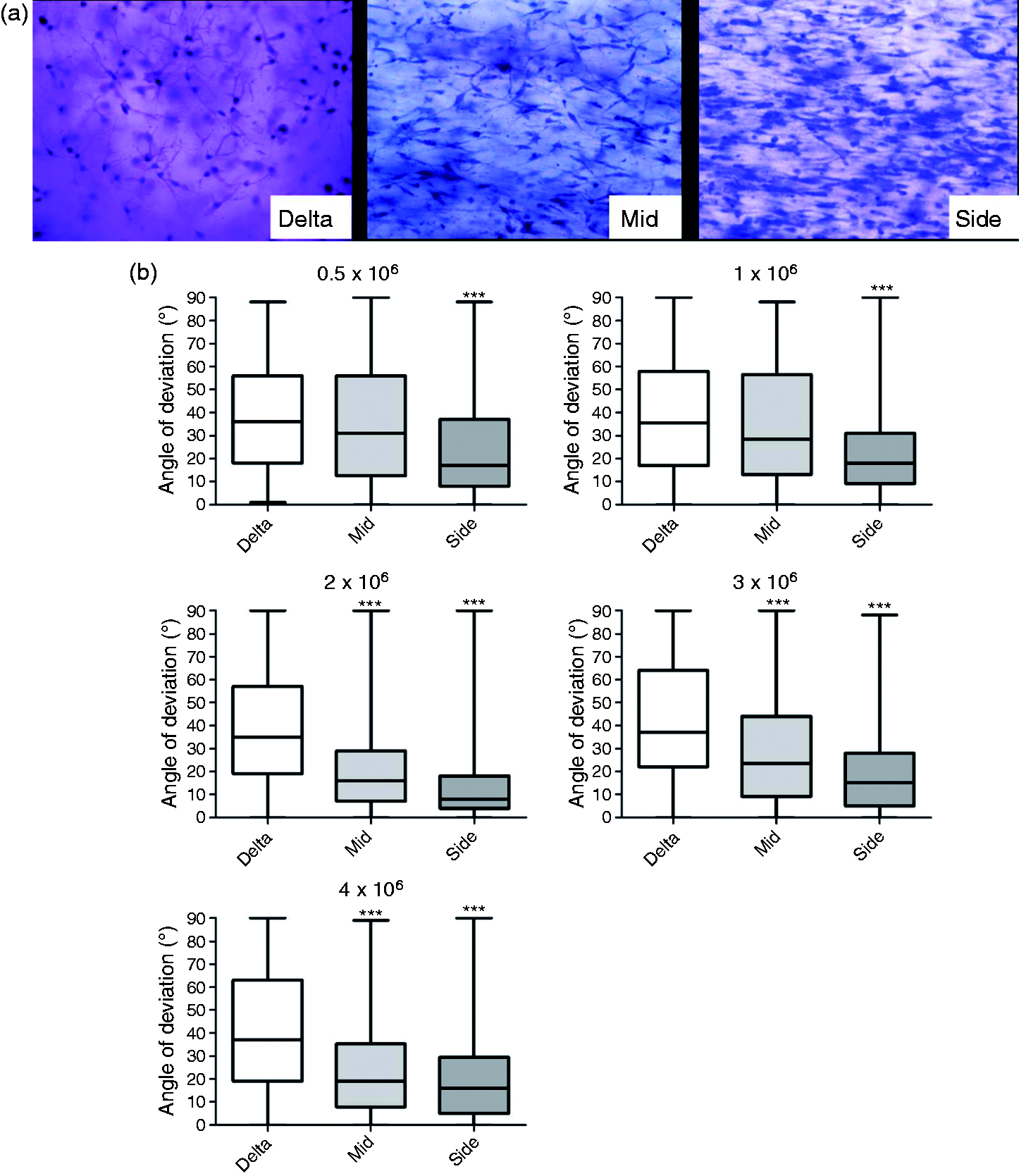
Validation of contraction assay as a way to select seeding density for cellular self-alignment
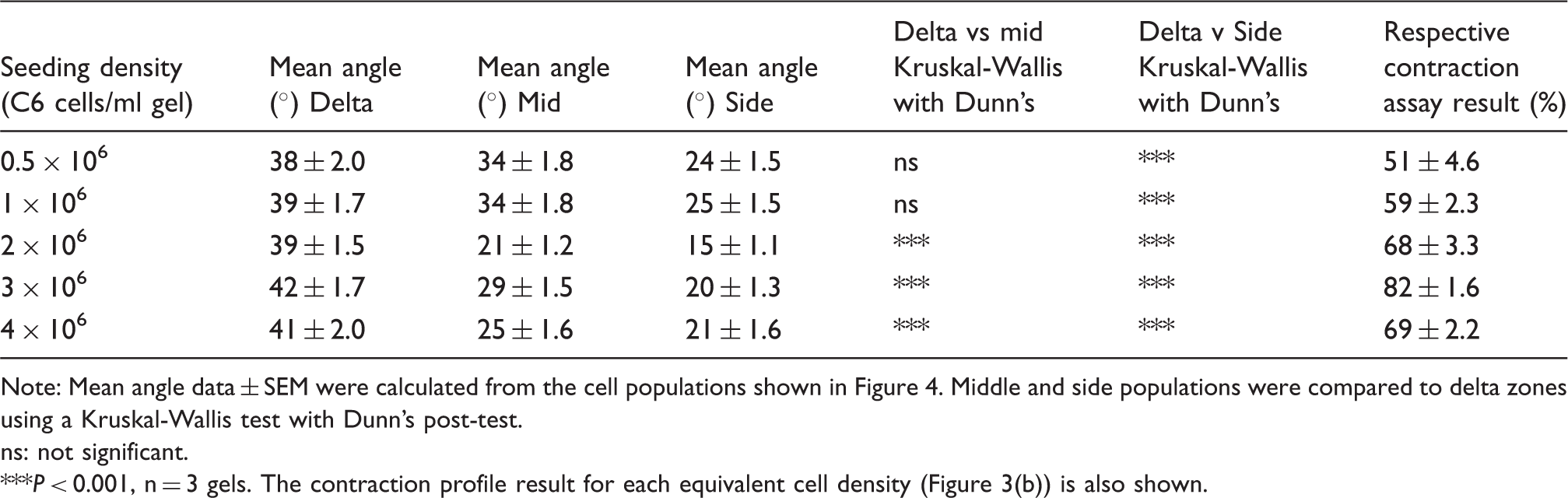
Having established the link between % contraction in the 96-well plate assay and subsequent cellular self-alignment using C6 cells, a validation experiment was conducted using primary rat astrocytes. Previous work showed that stimulation of primary rat astrocytes in collagen gels using TGFβ1 caused them to adopt a more reactive phenotype with an accompanying increase in gel contraction.4,16 Contraction assays were conducted using varying cell densities in the presence or absence of TGFβ1 (Figure 5(a)). Based on this, a cell seeding density of 4 × 106 cells/ml in the presence of TGFβ1 was selected as providing between 60 and 80% contraction and subsequently tested for cellular self-alignment within the tethered aligned system. Figure 5(b) shows that the primary astrocytes became highly aligned in both the middle and side regions.
Using the contraction assay to predict the seeding density for successful alignment of primary rat astrocytes. Twenty-four hour contraction assay (collagen gels in 96-well plates) for primary rat astrocytes in the presence or absence of TGFβ1 (a). Data are mean ± SEM, n = 5 gels per condition from a total of two experimental gel batches. On the basis of the contraction assay, a cell density of 4 × 106 primary rat astrocytes/ml was chosen to assess cellular self-alignment within tethered gels following 24 h in culture (b). Middle and side populations were compared to delta zones using a Kruskal-Wallis test with Dunn’s post-test, n = 2 gels.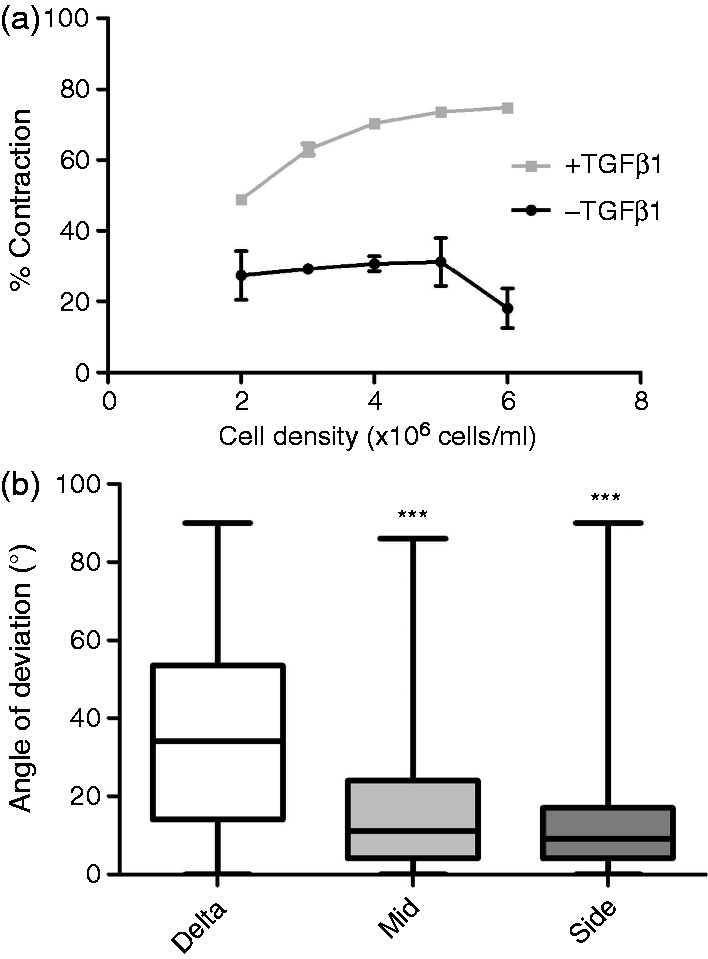
The effect of collagen concentration on contraction of cellular gels
To test the hypothesis that the concentration of collagen present will affect the extent of cell-mediated hydrogel contraction, gels were made using a range of collagen concentrations (1–5 mg/ml). Contraction profiles, using C6 cells revealed that in all cases contraction increased with cell density, and the gels with lower collagen concentration contracted more than those with higher collagen concentrations (Figure 6).
The effect of collagen concentration on contraction. Free-floating gels were made in 96-well plates using different collagen concentrations and seeded with a range of C6 cell densities. Their % contraction was assessed after 24 h. Data are means ± SEM, n = 6 gels per seeding density from at least four cultures.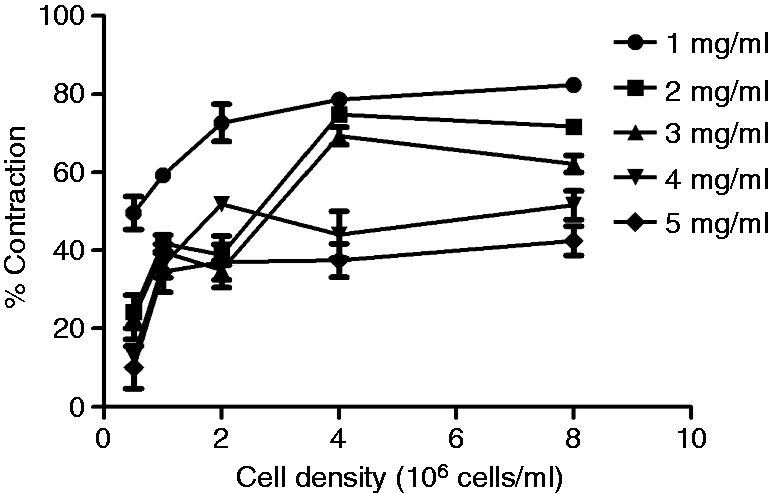
Time-course of collagen gel contraction over 24 h
To complete the investigation of the factors affecting contraction of collagen gels by C6 cells, the time-course was examined, using two different seeding densities in collagen gels.
Figure 7 shows that most of the contraction took place in the initial 4 h for cells seeded at 2 × 106 cells/ml and the first 6 h for cells seeded at 4 × 106 cells/ml, then the rate of contraction was slower for the remaining time. By 24 h, similar contraction was achieved at both cell densities.
Time-course for collagen gel contraction. Free-floating gels were made in 96-well plates using 2 or 4 × 106 C6 cells/ml. Cultures were photographed at various times throughout the 24 h incubation period and the % contraction determined at each stage. Data are means ± SEM, n = 5 gels at each density.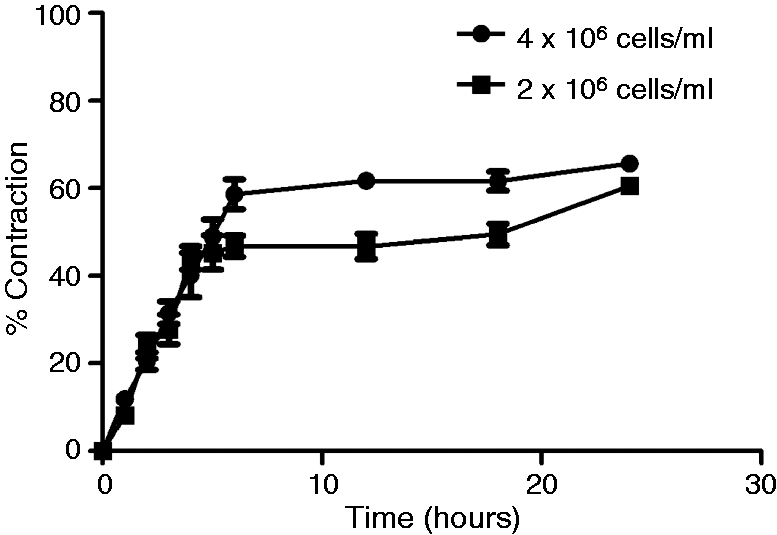
Discussion
Characterisation of the cell-mediated contraction of collagen gels is important for the development of engineered tissues that exploit the cellular self-alignment that accompanies generation of tension in tethered gels. Here we have shown how simple 96-well plate assays can be used to investigate cell–ECM interactions and optimise parameters such as cell seeding density, collagen concentration and incubation time to facilitate subsequent generation of aligned neural cell constructs. The desired end result of this investigation is to achieve consistent, predictable self-alignment of neural cells within hydrogels to mimic the aligned tracts in the CNS and peripheral nerves for regenerative medicine11 and to aid analysis and quantification.9,18 Self-alignment to achieve cell and matrix anisotropy within a 3D collagen environment is dependent on sufficient tension being developed through cell–ECM interaction and cytoskeletal activity. Parameters such as seeding density, collagen concentration and incubation time therefore need to be optimised for each new cell type. Since neural cells are often valuable and in short supply, and the measurement of alignment in tethered gels is time-consuming, the development of the simple 96-well plate assay described here is a useful new tool to speed up and simplify the construction of aligned neural tissues. In addition to being useful for characterising the ability of new cell types to self-align, this assay can potentially be used in testing of batches of cells for engineered neural tissue (EngNT) construction, release criteria, or product assessment.
Functional attachment of cells to collagen is an essential step in the self-alignment process and the ability of neural cells to align in this system will therefore depend on their expression of relevant integrins. It is accepted that α1β1, α2β1, α10β1 and α11β1 are the integrins responsible for direct attachment of cells to collagen fibrils.19–21 Here we used the 96-well assay to demonstrate that blocking the α1 integrin subunit of C6 cells inhibited contraction of gels in a dose-dependent manner, indicating the critical role played by α1 integrin in C6 cell-collagen interaction. Blocking the α2 integrin subunit however, was less effective, with only a high concentration of antibody reducing contraction. Inhibition of contraction achieved with anti-CD49a was not total, and anti-CD49b had minimal effect, therefore indicating the presence of other binding mechanisms, for example directly via α10β1 and α11β1 integrins or indirectly via other ECM molecules present in the cultures (e.g. fibronectin from the FCS which can bind to both cells and collagen). Integrin expression in neural cells varies according to the specific cell type and phenotype22,23; for example, Schwann cells which have been used previously in the tethered aligned collagen gels express α1β1 to a greater extent than α2β1, and the pattern of integrin expression is altered by interactions with axons. 24 The C6 cells used here and in many other in vitro studies are glioma-derived and their β1 integrin expression is associated with invasion of brain tissue, 25 which may improve their ability to contract and align in tethered gels compared for example to primary astrocytes. 4 The diversity of integrin expression among neurons and glia adds to the unpredictability associated with generation of self-alignment of untested neural cells in collagen gels and reinforces the importance of developing systems for optimisation.
The effects of time and cell density upon collagen contraction have been studied with regard to the behaviour of fibroblasts.14,26 Here we used C6 cells to determine the relationship between % contraction of free-floating gels 14 and self-alignment of cells in equivalent tethered gels. In this case, the free-floating gels were prepared in standard 96-well plates, using small volumes of gel (75 µl), making the assay suitable for widespread adoption and minimising the materials used in the optimisation of conditions for new cell types. This assay format could also be automated. The results showed that the free-floating gel contraction which produced reliable alignment throughout the middle and side zones of the equivalent tethered gel was above 60%. Contraction of less than 60% in the free-floating gels was associated with less alignment; specifically, alignment of the middle zone of the equivalent tethered gel did not differ significantly from that in the unaligned delta zones. The value of the contraction profile as a means of determining optimal cell density for self-alignment of other cell types was tested using primary astrocytes. The primary rat astrocyte contraction profile indicated that between 4 and 6 × 106 cells/ml in the presence of TGFβ1 would achieve sufficient contraction for self-alignment in tethered gels. Using the lowest seeding density in the predicted range in the tethered aligned system, highly aligned middle and side regions were observed, validating the use of the contraction assay as a tool for the optimisation of seeding density to generate aligned engineered constructs. Furthermore, the relationship between C6 cell and primary astrocyte seeding densities and collagen gel contraction was similar to that seen previously with other cell types, where contraction increased with increasing cell density until a plateau was reached between 75 and 80%.27,28
In addition to cell seeding density, another factor that affects the cellular contraction of collagen gels is the initial collagen concentration.14,26,29–31 To explore this in detail, 5 mg/ml collagen was diluted to give concentrations of 1, 2, 3, and 4 mg/ml, and gels of each concentration assessed in terms of contraction after 24 h with a range of C6 cell densities. The gels prepared with lower concentrations of collagen contracted more than those prepared with higher concentrations at each cell seeding density, which is similar to previous studies using fibroblast-populated gels. 32 Similarly, the time course of C6 cell-mediated collagen gel contraction leading up to the standard 24 h end point was investigated using two seeding densities. Interestingly, with both cell densities there was a rapid initial contraction phase in the first 6 h, followed by a much slower increase in contraction. This suggests that the 96-well contraction assay might be further refined by assessing gels at an earlier time point than 24 h in order to determine the likely ability of the cell condition under test to result in successful self-alignment in a tethered construct. Such a modification would potentially be beneficial in terms of improving the efficiency of the assay.
In summary, we have demonstrated how a simple accessible assay system using minimal quantities of reagents and cells can be used to establish optimal conditions for cellular self-alignment in tethered collagen gels. This will be particularly useful in the area of neural tissue engineering, where aligned cultures containing neurons and/or glia are being increasingly used for the engineering of implantable repair constructs or in vitro tissue models. Minimising the number of cells used to achieve successful self-alignment is of critical economic importance in the scale-up of engineered tissues, so an assay to help optimise this without the need to quantify alignment in numerous different tethered gels is a beneficial development. This study also demonstrated for the first time the involvement of α1 integrin in the self-alignment within collagen gels of C6 cells, a cell line that is widely used in neuroscience research. Furthermore, a detailed investigation of the relationship between cell density, collagen initial concentration and % contraction was conducted, providing a useful reference framework for future development of self-aligned neural tissues.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a PhD studentship funded by TAP Biosystems and the Open University.