
Abstract
Sepsis is a life-threatening problem by organ dysfunction influenced by negative inflammatory responses and stimulated oxidative stress, which most of sepsis patients about 40–60% are accompanied with myocardial injury. Recently, stem cells derived exosomes could effectively apply in the numerous diseases by combined with natural therapeutic agents. In the present investigation, Sweroside functionalized with exosomes to control inflammatory responses by sepsis and significantly proved the function of depreciated myocardial injury-induced by LPS. The sweroside could have effectively delivered to cardiomyocytes cells via exosome carriers. The induced-SMI rats exhibited severe myocardial injury and apoptosis by in vivo experiments and treatment of sweroside-functionalized exosomes (SWO/EX) reassured the phenotypes. Importantly, SWO/EX significantly downregulated the ROS generation in the SMI rat models. The SOD and GSH activity were also suppressed in SMI rat models, and treated models with SWO/EXO could have effective liberating activity in the Rats. Meanwhile, SWO/EXO treated LPS-induced cardiomyocytes displayed that significant reduction of pro-inflammatory cytokines (IL-1β, IL-6 and TNF-α) levels and also increasing cell survival and prevented apoptosis. Thus, we demonstrate that MS-cells derived exosome with sweroside could have effectively impede sepsis-induced myocardial injury. SWO/EX formulations might be applied as a potent therapeutic agent for SMI therapy.
Highlights
1. Stem cells derived exosomes could effectively apply in sepsis-induced SMI. 2. Sweroside functionalized with exosomes to control inflammatory responses. 3. Sweroside vesicle significantly downregulated the ROS generation. 4. SWO/EXO provide a significant reduction of pro-inflammatory cytokines. 5. SWO/EXO formulations might be applied as a potent therapeutic agent for SMI therapy.
Introduction
Sepsis is characterized by elevated secretion of inflammatory cytokines, oxidative stress, and dysfunction of multiple human organs. It is a systemic inflammatory reaction by upregulation of inflammatory cytokines. 1 Sepsis frequently leads to myocardial injury (SMI) and dysfunction as a consequence. Sepsis-related systemic abnormalities, including cardiac damage and injury, have been linked to the uncontrolled release of pro-inflammatory cytokines by activated inflammatory cells.2,3 It has been proven that myocardial injury happens up to 60% of the time in the first 3 days of being admitted for septic shock, which lead to high fatality risk for infected patients. 4 According to previous reports, excessive inflammation, mitochondrial dysfunction, an imbalance of oxidants and antioxidants, increased apoptosis, and problems with the autonomic nervous system all contributed to sepsis-induced heart damage. 5 The myocardial injury and dysfunction after sepsis have been significantly lowered left ventricular ejection fraction, impaired myocardial contractility, and transient biventricular dilation in patients. Therefore, it is essential to find a specific biochemical process and mechanisms of SMI in order to create new therapeutic approaches. 6
Exosomes are significant paracrine mediators, and more recent biomedical researches has shown that the treatment impact of MSC-based therapies is mostly attributed to the paracrine production of bioactive substances.7,8 Exosomes are lipid bilayer vesicles that range in size from 30 to 200 nm and can be generated by different types of cells. These exosome nanoformulations have wide range of bioactive compounds including proteins, lipids, miRNAs and mRNAs, which are acquired from the parental cells.9,10 The exosome’s lipid bilayer membrane can preserve the active ingredients including structural stability, coherence, and biological efficacy. Exosome vesicles are produced when multivesicular endosomes branching inward combine with the plasma membrane before being discharged into the extracellular microenvironment to affect the physiology and function of recipient cells.11,12 Exosomes derived from MSCs have been demonstrated to have substantial therapeutic potential without any adverse effects in a variety of illness models, including cardiovascular diseases, neurological diseases, and acute inflammatory injury. These exosomes are essential players in cell-cell communication in many diseases diagnosis therapies.13,14 Recent research indicates that exosome-derived from MSCs may contribute as an alternative to stem cell transplantation therapy in myocardial infarction by controlling the biological process and molecular pathways of MSCs. The treatment of MSCs-derived exosomes may enhance myocardial healing in myocardial injury by transforming M1 pro-inflammatory macrophages to M2 anti-inflammatory macrophages. Regulation of a healthy connection between M1 and M2 macrophages may be a potential beneficial therapeutic strategy in cardiac repair since macrophage polarization plays a significant role after myocardial injury.15,16 Additionally, when treated with low concentrations of pro-inflammatory agents like lipopolysaccharide (LPS), MSCs-derived exosomes retain information from toll-like receptors and have improved therapeutic efficacy in reducing inflammation. This is significant because immune-inflammatory cells communicate some aspects with MSCs-derived exosomes. 5
Natural sweroside, a Chinese traditional medicinal compound extracted from Swertia pseudochinensis Hara’s plant, which said to have potent antioxidant, anti-inflammatory, anti-bacterial, anti-fungal, anti-diabetic, and hepatoprotective properties.17,18 According to previous research, pre-treatment with sweroside prevented cardiomyocyte oxidative stress and intercellular ROS generation caused by aconitine. Sweroside has also been discovered to be capable of reducing inflammatory responses brought on by LPS or IL-1β by inhibiting the NF-κB signalling pathway. 19 Noticeably, a recent study revealed that treatment of sweroside could prevent the activation of the NLRP3 inflammasome.20,21 The present investigation demonstrated that increased the cardiomyocytes cell viability in LPS-induced or E. coli-induced sepsis model, inhibited cell apoptosis, simultaneously decreased IL-1 and TNF levels, and upregulated the expression of superoxide dismutase (SOD) and glutathione peroxidase (GSH-PX) in the plasma and culture medium. Further research revealed that SWO/EX reduced LPS-induced SMI by activating the Nrf2/HO-1 pathway and suppressing the TRAF6/NF-B signalling pathway. Overall, the current researchers indicated the therapeutic potential of SWO/EX in sepsis-induced MI, providing guidelines on how to treat SMI.
Experimental section
In vitro cell culture method
Bone marrow-derived mesenchymal stem cells (BM-MSCs) and H9C2 cardiomyoblasts were obtained from Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The cell proliferation medium for both cells was supplemented with Gibco Dulbecco’s modified Eagle’s medium, supplemented with nutrient mixture F-12 (DMEM/F-12), fetal bovine serum (10% (v/v); FBS; Gibco), with streptomycin (100 mg/mL) and addition of penicillin G (100 U/mL). The conditions for all co-culture incubation were 37°C, 5% CO2, and 100% humidity.
Extraction of MSCs-derived exosomes
Exosomes from BM-MSCs were extracted as stated previously with some minor modifications.16,22 When the BM-MSCs were 85–95% present (%) cell confluency, the α-MEM supplemented FBS (10%) was switched out for an exosome-free FBS (10%) osteogenic medium. FBS media was ultracentrifuged at 100,000g for 12 h to get Exosome-free FBS medium. The BM-MSCs medium was recovered after 48 h, and cells and cellular debris were removed by centrifuging it at 400g for 15 min, 2000g for 15 min, and 10,000g for 40 min. A sterile filter (0.22 µm) was then used to filter the medium. To get the exosomes, the supernatant was then ultracentrifuged at 100,000g for 2 hours. Hitachi transmission electron microscopy (TEM; H-7650, Japan) was used to determine the morphology of BM-MSCs-derived exosomes. Malvern NanoSight machine (LM10, UK) were used to assess the particle size distribution.
Exosome-uptake assay
BM-MSCs-derived Exosomes were stained with red Dil dye (Beyotime, China) in accordance with the package recommendations. The human fibroblast (L929) cells were then treated with the Dil-labeled exosomes for 4 and 24 h. The L929 cells were then labelled with a DAPI after being fixed. The results were visualized using confocal laser scanning microscopy.
Preparation of sweroside -loaded exosomes
The procedure for encapsulating sweroside in BM-MSCs derived exosomes was as follows: The obtained sweroside (1 mg) compound was weighed and dispersed in DMSO (200 µL) solution containing Tween-80 (2%), along with extracted exosomes (1.5 mg). This mixture was then constantly blended at 4°C for an entire night before being incubated continuously with ultrasound in an ice-water bath. To remove the unloaded sweroside, the resulting SWO-EXO was washed twice. High performance liquid chromatography (HPLC) was performed to measure unbound sweroside in the supernatant after reaction.
Analysis of ROS content
The intracellular amount of ROS was examined using a ROS test kit from the Beyotime Institute of Biotechnology, China. In brief, H9C2 cardiomyoblasts were plated onto a 12-well culture plate, treated for 6, 12, and 24 h with prepared samples and treated with or without LPS (10 mg/mL; dissolved in ddH2O). After that, cells were introduced and thoroughly covered with 10 M DCFH-DA for 20 min. We examined the cells using an OLYMPUS fluorescent microscope (Japan) after three rounds of washing in serum-free growth media.
Analysis on in vitro anti-oxidant enzyme assays
RIPA lysis buffer (Beyotime, China) was used to lyse the heart tissue section or/and H9c2 cells on ice while also adding protease inhibitor pellets. By centrifuging lysates at 13,000g for 20 min at 4°C, the supernatants were collected, and the concentrations were determined using a Thermo Fisher BCA-kit (Waltham, MA). The supernatant was then collected on ice and kept in a freezer at 80°C. The anti-oxidant enzyme kits (Beyotime Institute of Biotechnology, China) were used to measure the activities of glutathione peroxidase (GSH-Px), malondialdehyde (MDA), and superoxide dismutase (SOD). The OD value of each well assay was measured that used a microplate reader in the prescribed order, and the corresponding value was measured.
In vitro analysis of mitochondrial function
The amount of MMP on LPS-induced cells and treatment of prepared samples were examined using the mitochondrial membrane potential (MMP) detection kit JC-1 (Beyotime Institute of Bio-technology, China). H9C2 cardiomyoblasts were co-reacted with JC-1 (2 g/ml) on six-well culture plates for 5 minutes in a 37°C incubator. The H9C2 cardiomyoblast cells were examined using an OLYMPUS IX53 fluorescent microscope (Tokyo, Japan) after being washed twice with PBS. MMP was expressed by the ratio of red to green fluorescence. The ATP assay kit was used to determine the relative ATP contents in accordance with the manufacturer’s instructions.
In vivo animal models and experimental grouping
The Shanghai Slac Laboratory Animal Co., Ltd (Shanghai, China) supplied male C57/B6J mice (six-week-old), which were housed in the animal facility of the Dongying People’s Hospital, Dongying, China. The ideal housing circumstances included standard cleanliness, a temperature control of 25°C, a light/dark cycle (12 h/12 h), and free access to standard mouse feed and water. The Animal Ethics Committee of Dongying People’s Hospital granted its consent for the experiments to be carried out in accordance with the National Institutes of Health’s Guidelines for the Use of Laboratory Animals. The care and management of the animals were conducted in accordance with Dongying People’s Hospital stated policies. In this study, 72 mice were used, and they were randomly assigned to five different groups: (i) control group (n = 9); (ii) sham group (n = 9, intraperitoneal injection of menstruum); (iii) LPS group (n = 18, injection of a single dose LPS at an intensity of 8 mg/kg); (iv) LPS + EXO and (v) LPS + SWO-EXO treatment. Following the administration of LPS, treatment samples were injected immediately. Three mice from groups (i) and (ii) and six mice from each of groups (iii), (iv), and (v) were sacrificed, and their heart tissue sections were removed for further study at 6, 12, and 24 h post-LPS/menstruum injection.
Histopathological observation of H&E staining
After a 24-h period with or without menstruation or LPS treatment, myocardial tissue sections from three mice in each group were fixed in 4% formaldehyde, dried using graded ethanol solution, cleaned in xylene solution repeatedly, simultaneously dipped in wax, and then paraffin embedded with a thickness of 4 m. Moreover, the tissue section underwent stretching, roasting, dewaxing, hematoxylin impregnation, rinsing, returning to blue with ammonia solution (0.5%), dyeing with eosin, rinsing, dewaxing, sealing, and drying in proper order process. Under a microscope (Olympus, Tokyo, Japan), the pathological section state was viewed from various angles, and photographs were taken in various multiples. The pathological outcomes were subsequently examined.
Apoptosis observation in myocardial cells by TUNEL assay
The TUNEL assay kit (TUNEL assay kit-FITC, Abcam, Shanghai, China) instructions were followed in order to detect apoptosis in cardiac tissue in situ. The treatment paraffin-embedded tissue sections were then processed with PBS (pH 7.4–8.0) diluted Proteinase K (2 mg/ml), and the sample was further incubated for 30 min at atmospheric temperature. The buffer solution was thawed and combined with the FITC-12-dUTP labelling mix. After removing the parafilm, the tissue section was immersed in the PBS solution and allowed to incubated for 10 min. The excess liquid was poured off, and the tissue sample was then incubated for a further 10 min at room temperature with PBS solution. The glass slide was placed inside the staining container with the DAPI solution and left there for 20 min. After being cleaned, the sample was placed on a glass slide, which was then let to immersed in deionized water for 5 min at room temperature. This process was carried out twice. An examination of the samples was done using OLYMPUS IX53 fluorescent microscope (Tokyo, Japan). 23
Expression analysis by real-time PCR analysis
We purified total RNA from tissues or cells using the TRIzol reagent in accordance with the manufacturer’s (Invitrogen) guidelines. The Transcriptor First Strand cDNA Synthesis Kit and an oligo dT primer (Promega Biotech Co., China) conjunction significantly worked to reverse transcription of total RNA (2 µg) into cDNA. On a 7500 cycler (Applied-Biosytems), amplification and quantitative RT-PCR analyses were carried out using Promega SYBR Green. Target genes' relative mRNA expression levels were standardized to GAPDH. The expressions level of mRNA was evaluated by computing the values of the Δcycle threshold (ΔCt), optimizing the average Ct value to that of the endogenous control (GAPDH or -action), and then calculating 2−ΔΔCt values.
Western blotting analysis
SDS-PAGE gels (Beyotime Biotechnology, Jiangsu, China) were used to separate the samples before they were electroblotted onto PVDF membranes. Tris-buffered saline Tween 20 (TBST) was used to block the membranes with 5% non-fat milk for 1 h. Overnight, primary antibodies such as phospho-p38MAPK (1:1000; Abcam), phospho-JNK ½ (1:1000; Abcam), phospho-ERK (1:1000; Abcam), p62 (1:1000; Abcam), NF-κB p65 (1:1000; Abcam) and GAPDH (1:1000; Abcam) were treated with the membranes. Secondary antibodies then reacted with the fasteners at room temperature for 2 h before being detected using a chemiluminescence ECL kit (Beyotime Biotechnology, Jiangsu, China).
Statistical analysis
For statistical analysis, GraphPad Prism 8.0 software was used. All Data are stated as the mean ± SEM. One-way analysis of variance and the Student-Newman-Keuls test were used to evaluate grouped data. A p value <.05 was considered as statistically significant.
Results and discussion
The MSCs extracted EXO had a spherical-shaped morphology with a mean diameter of 80–120 nm, according to TEM examination (Figure 1(a)). The diameter of EXO ranged from 50 to 150 nm, according to the results of the particle size distribution analysis (Figure 1(b)). The EXO expressed the exosomal markers CD9, CD63, and CD81, as determined by the Western blotting observations (Figure 1(c)). These data all indicated to the EXO having successfully isolated from MSCs. EXO are considered as essential and can be utilized for tissue regeneration applications among the many paracrine factors that MSCs secrete. The MSCs-extracted EXO can influence the biological processes of target cells by changing the gene expression and protein translation of receptor cells, according to previous studies.
8
MScs-derived EXO can also transmit proteins, miRNAs, and mRNA between the cells. Figure 1(d) exhibits fluorescence microscopic visualization of cells ingesting MSCs-derived EXO formulations. The findings demonstrated that the H9C2 cardiomyoblast cells absorbed the EXO labelled with red-fluorescent Dil throughout the course of 4 to 24 h. Even more exosomes EXO were absorbed and dispersed throughout the nucleus after the EXO were incubated with the cells for 24 h. Characterization of exosomes-derived from MSCs. (a) TEM observations of exosomes morphology. (b) Particulate size of the exosomes by DLS analysis and (c) Dil-labelled exosomes ingested with L9C2 cardiomyoblast cells in different incubation time of 6 h and 26 h (scale bar = 25 µm).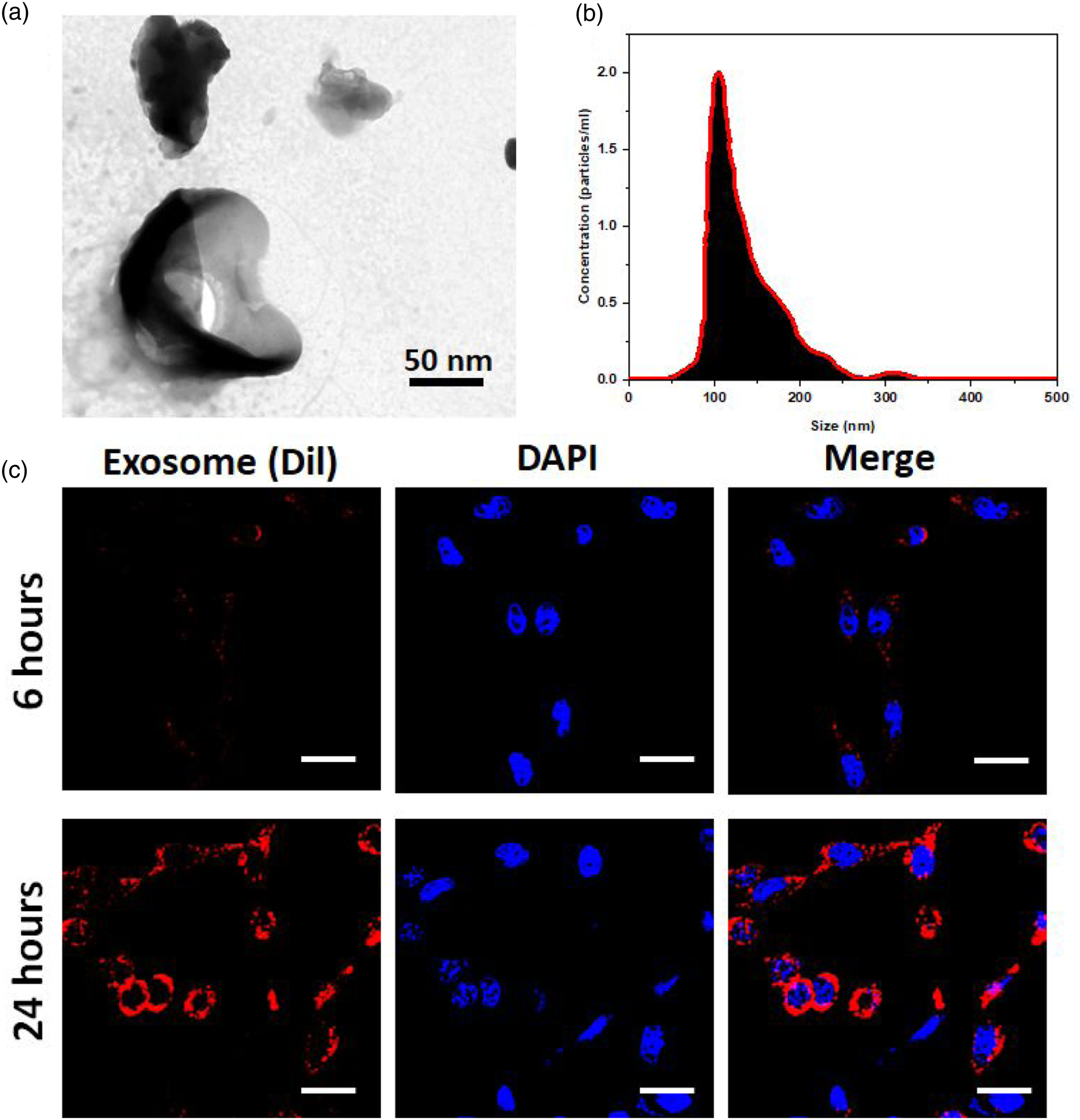
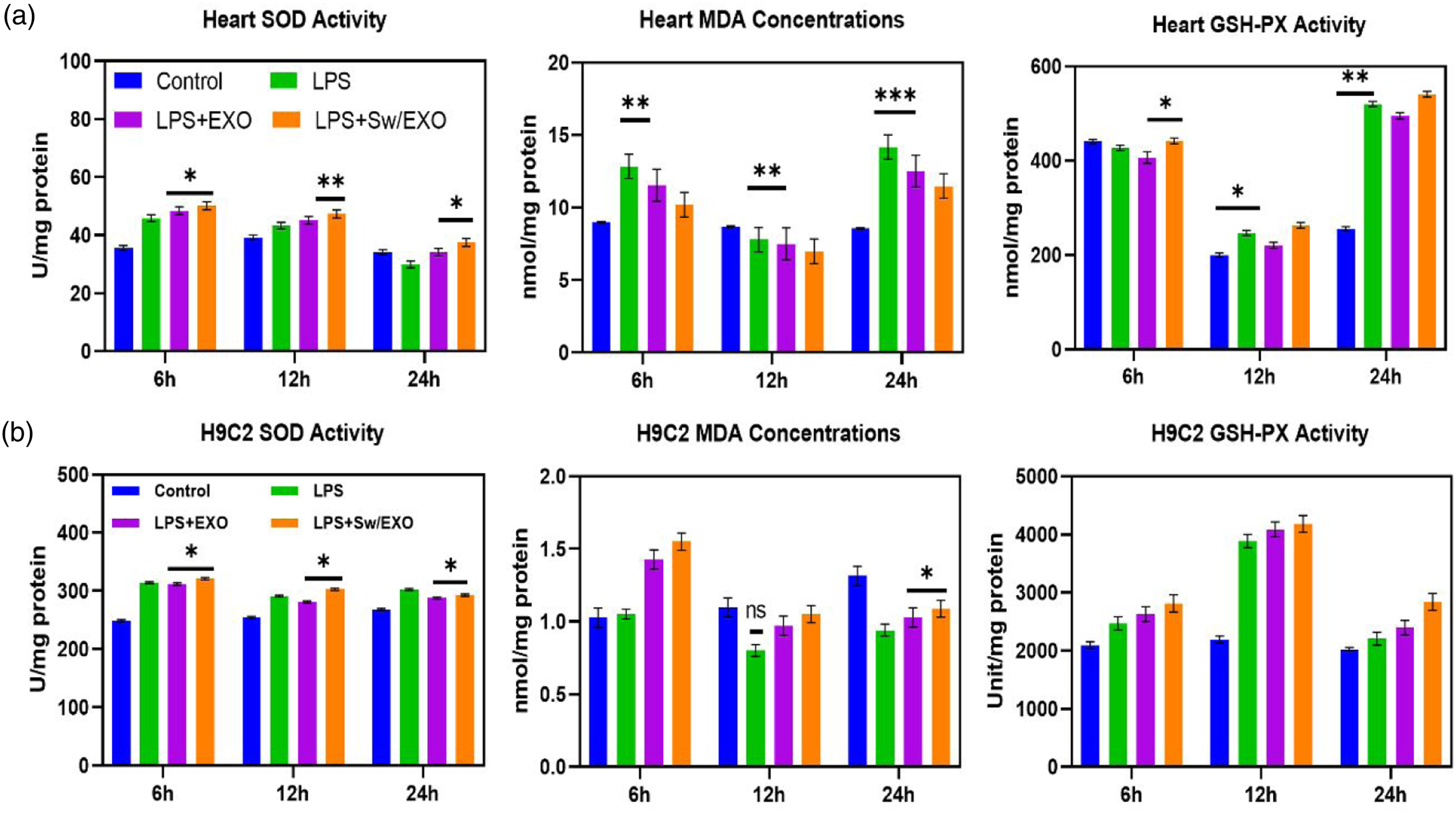
In the study of intracellular ROS generation using H9c2 cells, DCFH-DA staining demonstrated that the LPS treated groups have greater ROS production when compared to the control group. SWO/EXO pre-incubation significantly decreased ROS overproduction after LPS treatment, and there was no noticeable difference between the control, EXO, and SWO/EXO groups. Figure 2(a) illustrates that ROS concentration in the LPS group considerably enhanced when compared to the control group after 12 h. Despite the fact that LPS administration led to an overproduction of ROS, SWO/EXO treatment considerably reduced ROS production to the LPS group as exhibited in Figure 2(a) and (b) by Olympus fluorescent microscope. The expression parameters of ROS, MDA, SOD, and GSH-Px can all be used to determine cellular oxidative stress treatment of developed compounds by in vitro (Figure 3(a)) and in vivo analyses (Figure 3(b)). The addition of natural component of SWO effectively increased the functions of SOD and GSH-Px that were suppressed by LPS-induction while greatly reducing the levels of ROS and MDA content that were triggered by LPS. The observed levels of SWO/EXO treated cells can demonstrated the status of oxidative stress, which indicated that SWO and EXO are important antioxidant molecules. It was revealed that the addition of SWO caused the increased expression of EXO to nearly twofold. These findings indicate that SWO embedded EXO molecules may be able to reduce oxidative damage brought on by LPS. Systemic observations of in vitro and in vivo oxidative stress treatment with SWO/EXO vesicles: (a) fluorescence microscopic observation of H9C2 cells-treated with SWO/EXO vesicle after LPS-induction for 6 h, 12 h and 24 h and fluorescence ratio measurements and measured fluorescence intensity (b). Data are expressed as mean ± SE. *p < .05, **p < .01, ns: no significant difference. The relative expression levels of oxidative stress factors (SOD, MDA and GSH-Px) for in vitro (a) and in vivo (b) experiments in each treatment groups (control, LPS, LPS + EXO and LPS + SWO/EXO). All graph exhibits mean ± SE, *p < .05 versus LPS-induced group (scale bar = 50 µm). Data are expressed as mean ± SE. *p < .05, **p < .01, ns: no significant difference.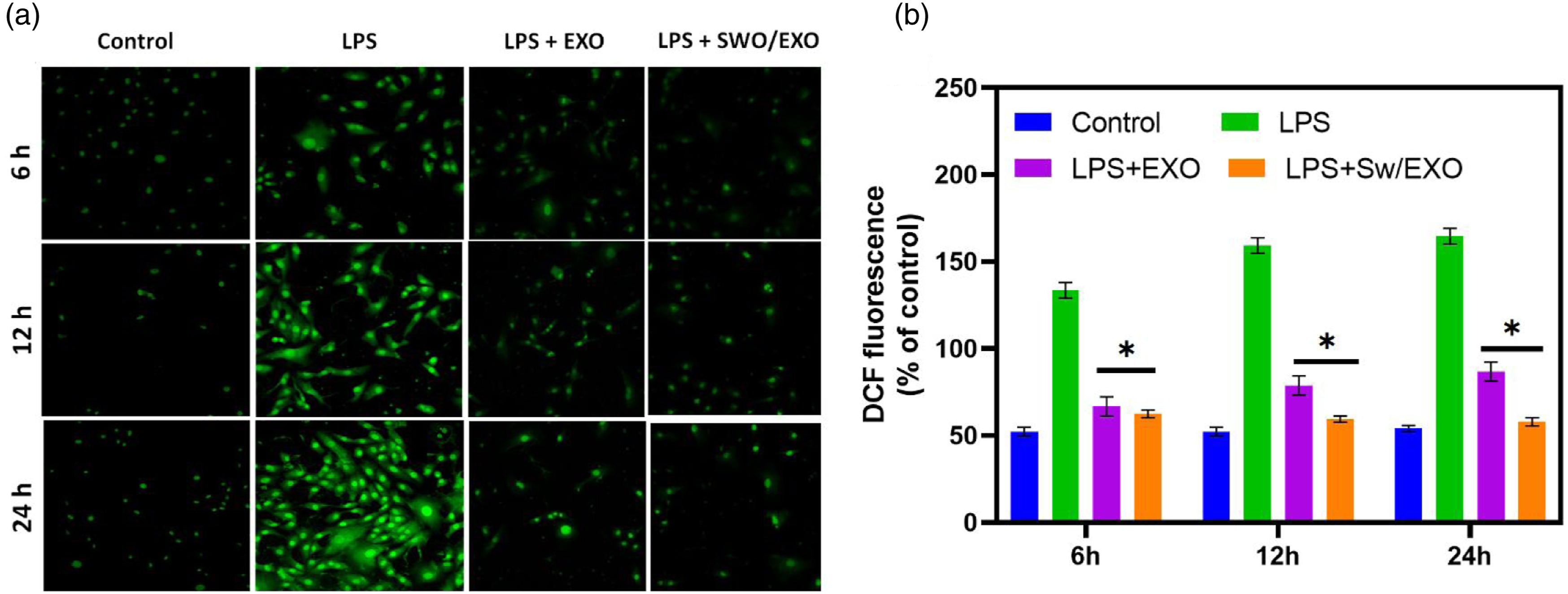
As exhibited in Figure 4(a), exposure to LPS caused depolarization of mitochondrial membrane potential (MMP), which resulted in mitochondrial damage and cell apoptosis of L9C2 cells. The mitochondrial membrane potential (MMP) reduced following LPS treatment, according to the investigation of the mitochondrial activity. EXO and SWO/EXO pre-treatment reversed this negative effect. In contrast, neither the EXO nor the SWO/EXO groups significantly differed in MMP when compared to the control group. Throughout this time observation, the ATP level was noticeably elevated by the administration of the EXO and SWO/EXO groups after 6 h of LPS exposure (Figure 4(b)). Although, there was no apparent time-dependent effect when treatment with SWO/EXO group. The structural damages of myocardial mitochondrial and reduction of mitochondrial density have been greatly associated to SMI. However little previously recognized about preventing cardiac mitochondrial damage to enhance sepsis outcomes. The major observations of our study demonstrated that delivery of SWO-loaded EXO vesicles effectively decreased inflammation and apoptosis and protected MMP, to diminish mitochondrial damage, and subsequently enhanced cardiac functions in LPS-induced rat models. Furthermore, SWO/EXO treatment significantly reduced the LPS-activated NF-κB signal pathway’s ability to further downregulated excessive inflammatory response. These observations suggest a potential role for the mitochondrial-specific targeted SWO/EXO in preventing LPS-induced inflammatory processes, mitochondrial functional deficits, and redox balance degradation in cardiomyocytes. Study on mitochondrial activity of L9C2 cells after LPS-induction and treatment with EXO and SWO/EXO vesicles. (a) Fluorescence microscopic images of mitochondrial membrane potential of LPS-induced L9C2 cells treated with samples. (b and c) In vivo and in vitro cardiac metabolism were evaluated by ATP content of heart and H9C2 cells. Data are expressed as the mean ± SE, *p < .05 versus LPS-induced group (scale bar = 50 µm).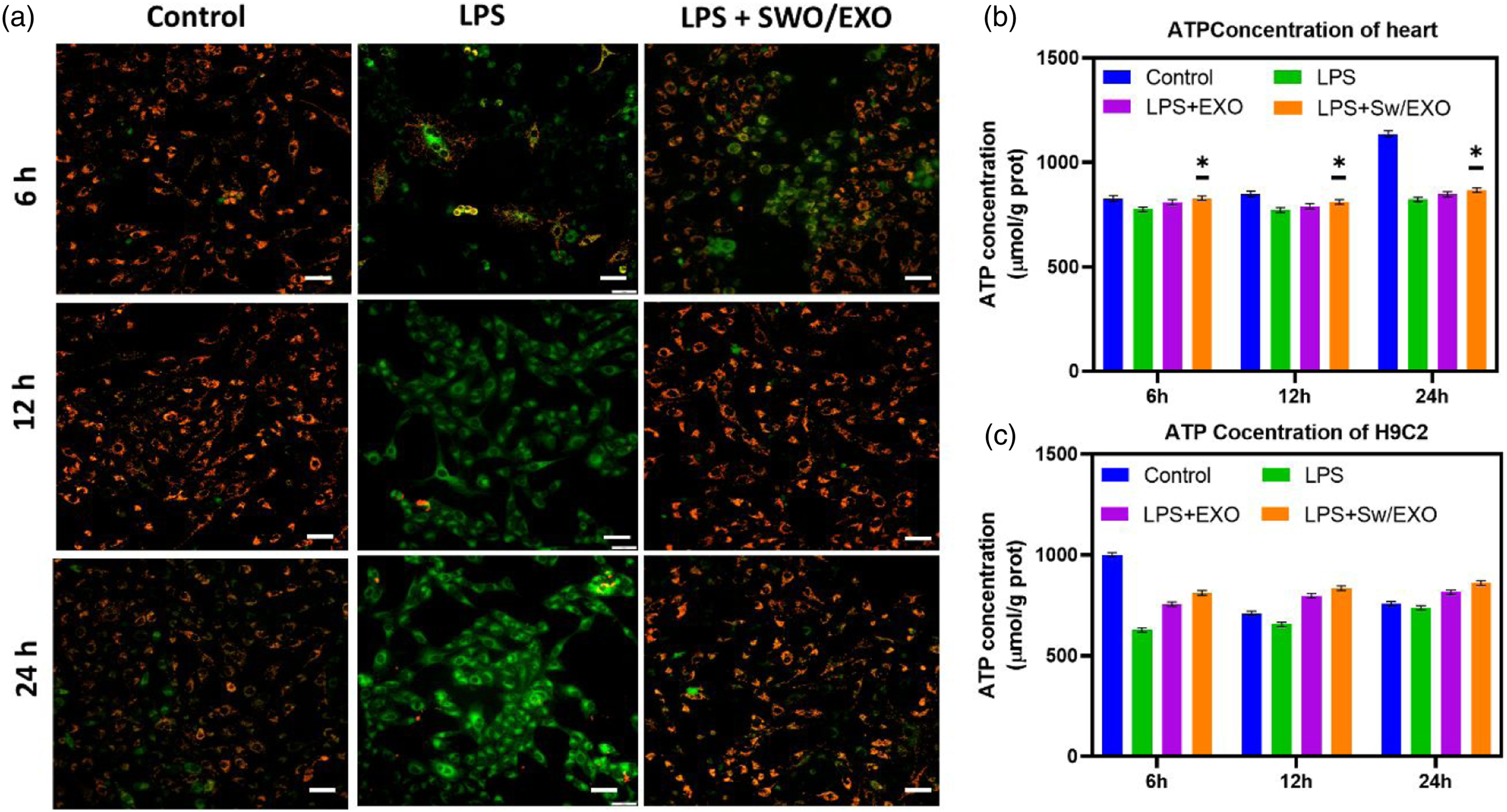
LPS-induction of intraperitoneal administration causes cardiomyocyte injury in vivo rat model. According to previous studies, after being exposed to LPS for 12 h, cardiomyocytes in the LPS group showed signs of cardiac injury as shown by the degree of apoptosis and inflammatory cell infiltration (Figure 5).24,25 Histopathological observation (H & E staining) revealed no apparent changes between the SWO/EXO group and the sham group. However, cardiomyocytes in the LPS group showed signs of myocardial injury after being exposed to LPS for 12 h as shown by the degree of necrosis and inflammatory cell infiltration as well as less obvious myocardial cross-striations. Figure 5 illustrates how the myocardial morphological improvements following after treatment of SWO/EXO delivery were greatly superior than those in the LPS group. The myocardial morphological changes of treated rat heart sections were observed by histopathological study: Rat groups were administered by LPS and intraperitoneally injection of prepared EXO and SWO/EXO vesicles. Heart tissue section were harvested and then stained with H&E staining. The microscopic observations were revealed that inflammatory infiltration and cardiomyocyte disruption (scale bar = 200 µm).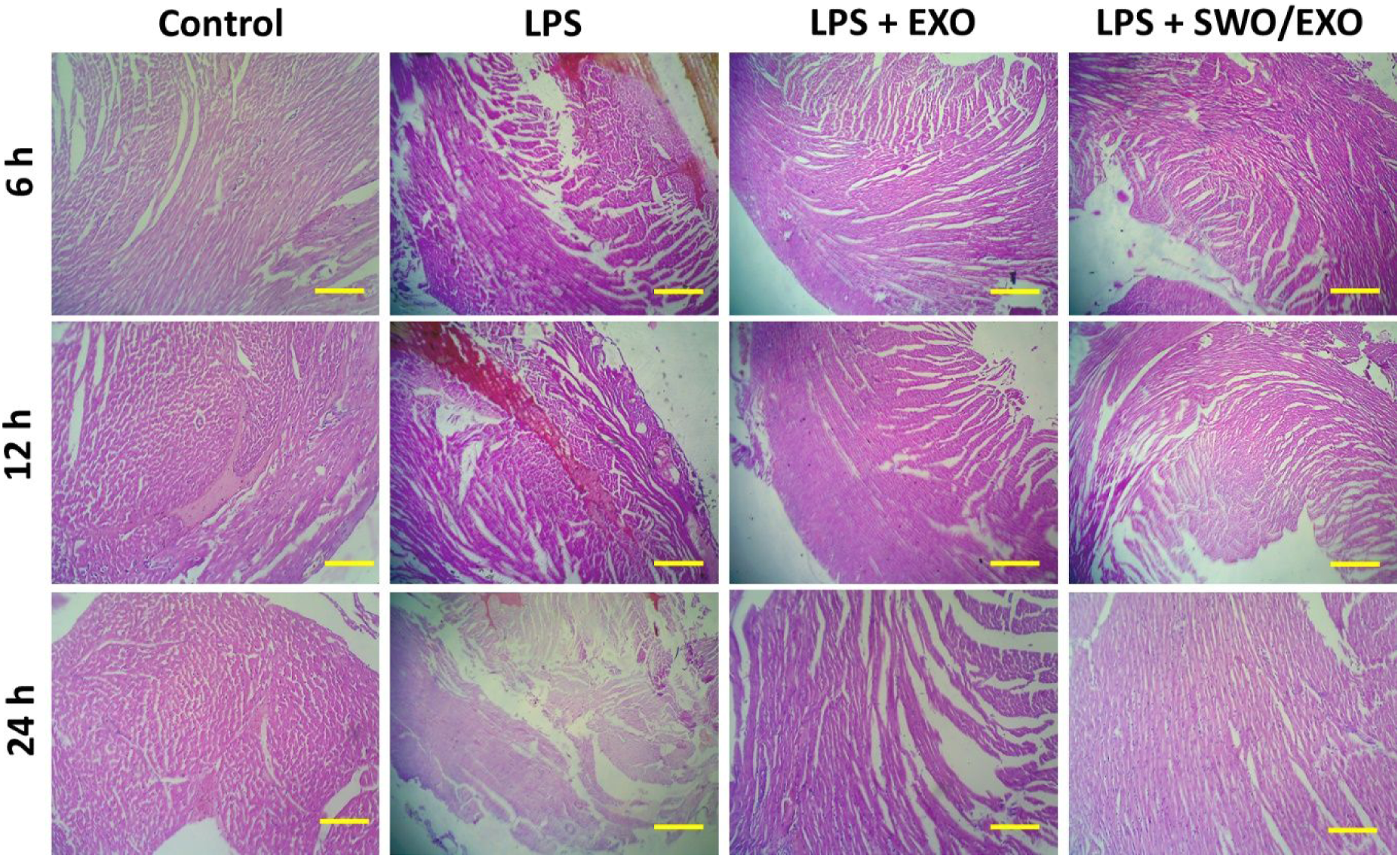
The control group’s nuclei were exhibited pale blue in colour with clear and intact morphological observation using TUNEL staining, whereas green florescent stain displayed apoptotic nuclei. The results demonstrated that LPS-induced treatment significantly augmented the number of TUNEL positive cells in comparison to the sham group after 24 h treatment. While SWO/EXO vesicle administration significantly reduced LPS-induced augmentation of the number of apoptotic cells, meanwhile EXO vesicle administration has little effect influence to prevent the elevation in apoptotic cells (Fig SI 1). These results indicate that the addition of SWO into the EXO vesicle considerably increases the anti-apoptotic action of SWO/EXO vesicle. Then, we examined into whether introducing SWO/EXO vesicles could impact the systemic inflammatory response to sepsis. The induction of LPS significantly enhanced the mRNA levels of inflammatory cytokines including TNF-α IL-1β and IL-6, in both experimental models of in vivo and in vitro as shown in Figure 6. Importantly, SWO/EXO prevented the LPS-induced elevation of inflammatory cytokines in H9C2 cells (Figure 6(b)). These observations indicated that, the treatment of SWO/EXO reduced the expression of pro-inflammatory cytokines when compared to the LPS-induced group (Figure 6(a)). During sepsis, it appeared that SWO/EXO had no noticeable time-dependent effect. Prepared SWO/EXO reduced cardiac inflammatory responses of LPS-induced heart sections and H9C2 cells. The mRNA expressions of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) was measured by RT-PCR at different incubation hours (6 h, 12 and 24 h). (a, b & c) Rat models were injected with LPS, LPS + EXO and LPS + SWO/EXO vesicle and (c) L9C2 cells were influenced with LPS, LPS + EXO and LPS + SWO/EXO vesicle. Data are expressed as mean ± SE. Data are expressed as mean ± SE. *p < .05, **p < .01, ns: no significant difference.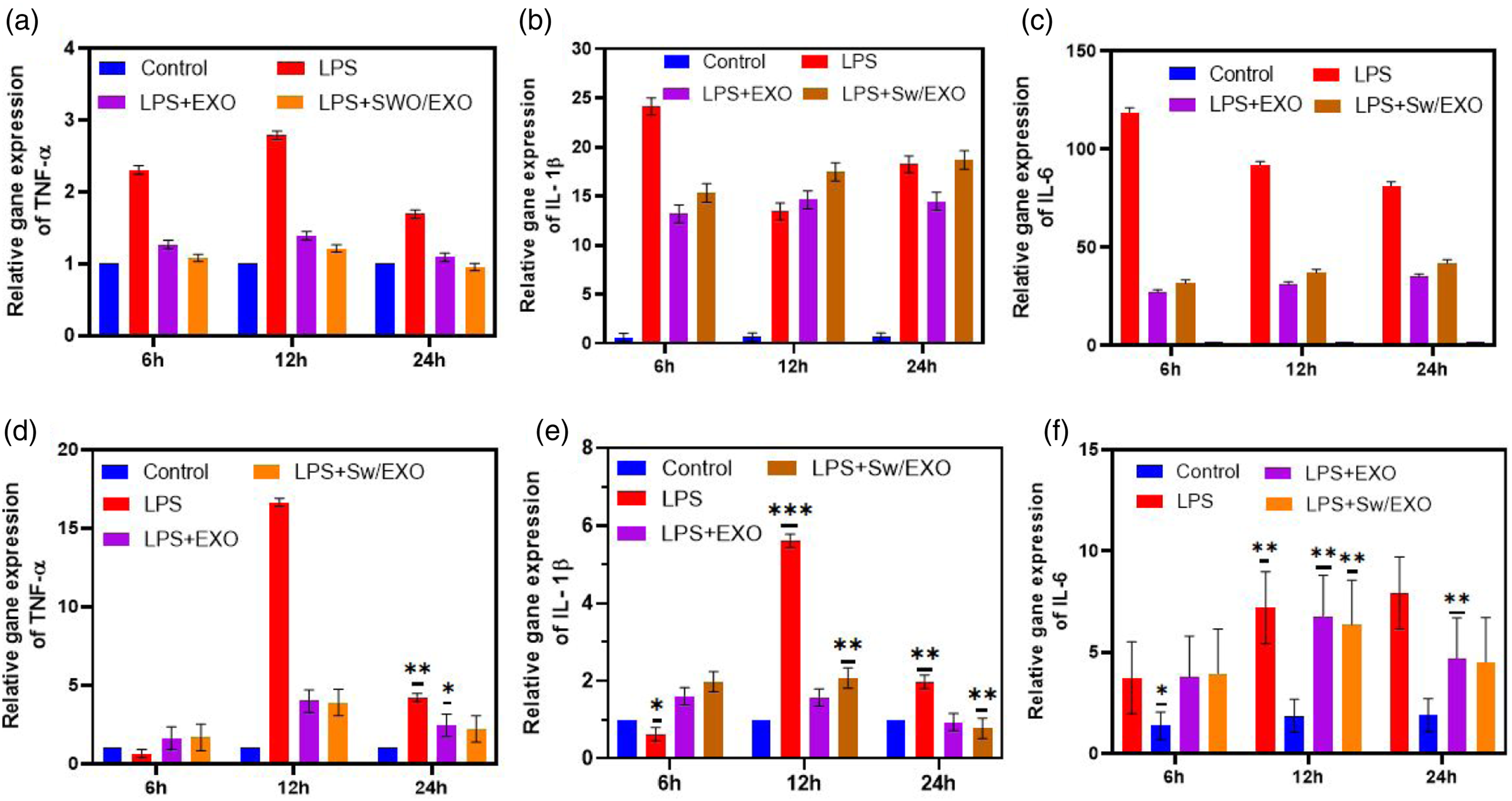
Diverse signalling pathways, including NF-κB and MAPK, were known to play a role in the progression of sepsis. p-P38MAPK, p-JNK1/2, p-ERK, p-62, and endonuclear p65 levels have been determined by western blotting methodology. Figure 6 shows that LPS treatment induced p65 phosphorylation and nuclear translocation, which would be inhibited by SWO/EXO administration as exhibited in Figure 7(a)–(e). However, p62, JNK1/2, and p-p38MAPK levels were shown to be significantly different between the LPS and LPS + EXO groups. In both sepsis models of animals and humans, myocardio-depressant factor suppresses mitochondrial function, which plays a crucial role in the development of sepsis.25,26 Studies indicate that reducing oxidative stress and inflammation reduces mitochondrial dysfunction, and that myocardial depressive factors like IL-1β, TNF-α, and IL-6 are committed to increasing mitochondrial damage. In addition to damaging mitochondria, high levels of inflammatory mediators promote the formation of peroxides that damage neighbouring tissues, impair heart function, trigger multi-organ failure, and ultimately led to mortality.1,10 Anti-infection therapy seems not to significantly increase the survival rate of sepsis patients. In sepsis, it has been shown that inhibiting IL-1β specifically had no effect on mortality or morbidity. Furthermore, anti-TNF-α therapy was successful in extending the lifespans in infected animals but failed to decrease deaths in sepsis patients. Previous reports have also emphasised that despite the critical importance of anti-inflammatory medicines, their administration in clinical trials failed to reduce the overall mortality of patients with sepsis.8,27 We found that levels of IL-1β, TNF-α, and IL-6 were all significantly reduced after treatment with SWO. A number of cardiac inflammatory responses during sepsis have been linked to the NF-κB signalling pathway, one of many kinases involved in this process. In fact, we found that SWO had some suppressive effect on NF-κB. The RNA expression of pro-inflammatory factors induced by LPS may be suppressed by SWO-loaded Exo vesicles through regulation of the NF-κB signalling pathway. Effects of developed SWO/EXO on signaling pathway. The quantification value of proteins (p-P38MAPK, JNK, p-ERK, p-62 and NF-κB p65) by fold change from collected heart sections for the detection of signaling pathways after LPS induction for different incubation hours 6 h, 12 h and 24 h. All the proteins were normalized to GAPDH before quantification. Data are expressed as mean ± SEM. *p < .05 versus the control group.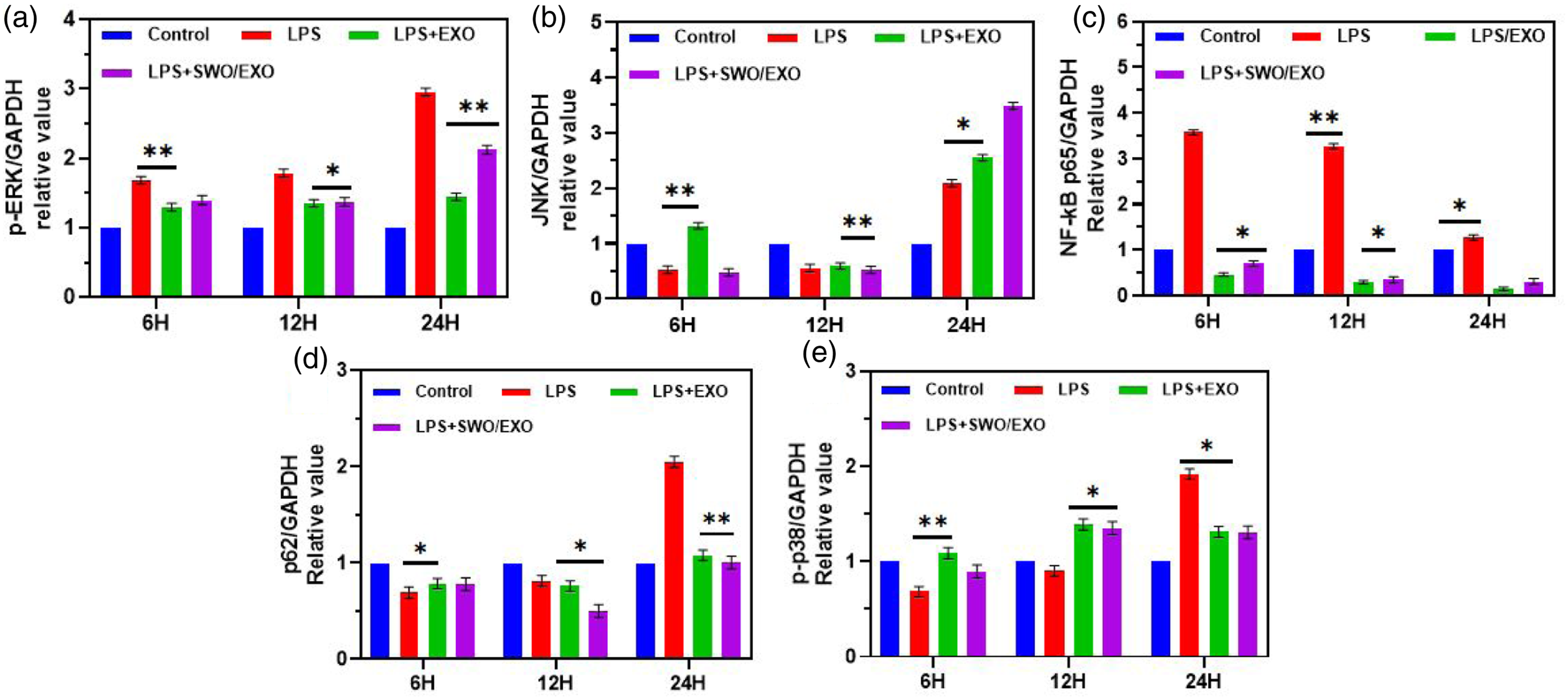
The LPS-induced group’s 72-h mortality was 70%, but the LPS + EXO and LPS + SWO/EXO groups’ 72-h mortality rates were 30% and 20%, respectively, confirming extended survival of rat models (Figure 8(a)). Echocardiography was used to examine heart function measures in order to look at how SWO loaded EXO vesicles influenced the rat model of sepsis caused by LPS. According to the echocardiography observations, LPS-induced mice exhibited considerably deteriorated cardiac function as shown by noticeably decreased HR (Figure 8(b)), FS% (Figure 8(c)), EF% (Figure 8(d)), and LVPWs (Figure 8(e)), as well as noticeably increased LVIDs (Figure 8(f)). Treatment with SWO/EXO significantly reversed these adverse effects. In terms of LVIDd and LVPWd, there were no noticeable differences between the groups. These results demonstrate that SWO/EXO treatment enhanced survival rates and conserved cardiac function in animal models of LPS-induced myocardial damage. Prepared SWO/EXO vesicle protected cardiac function and improved the survival rate in LPS-induced rat models. (a) SWO/EXO vesicle was administered intraperitoneally injected after LPS-induction for 2 h and then for administered for three consecutive days. The mortality rate of rat was observed for 72 h (n = 8 rats), (b) heart rate, (c) fractional shortening, (d) ejection fraction, (e) LVIDs, (f) LVIDd, (g) LVPWs and (h) LVPWd. Data are expressed as mean ± SE. *p < .05, **p < .01, ns: no significant difference.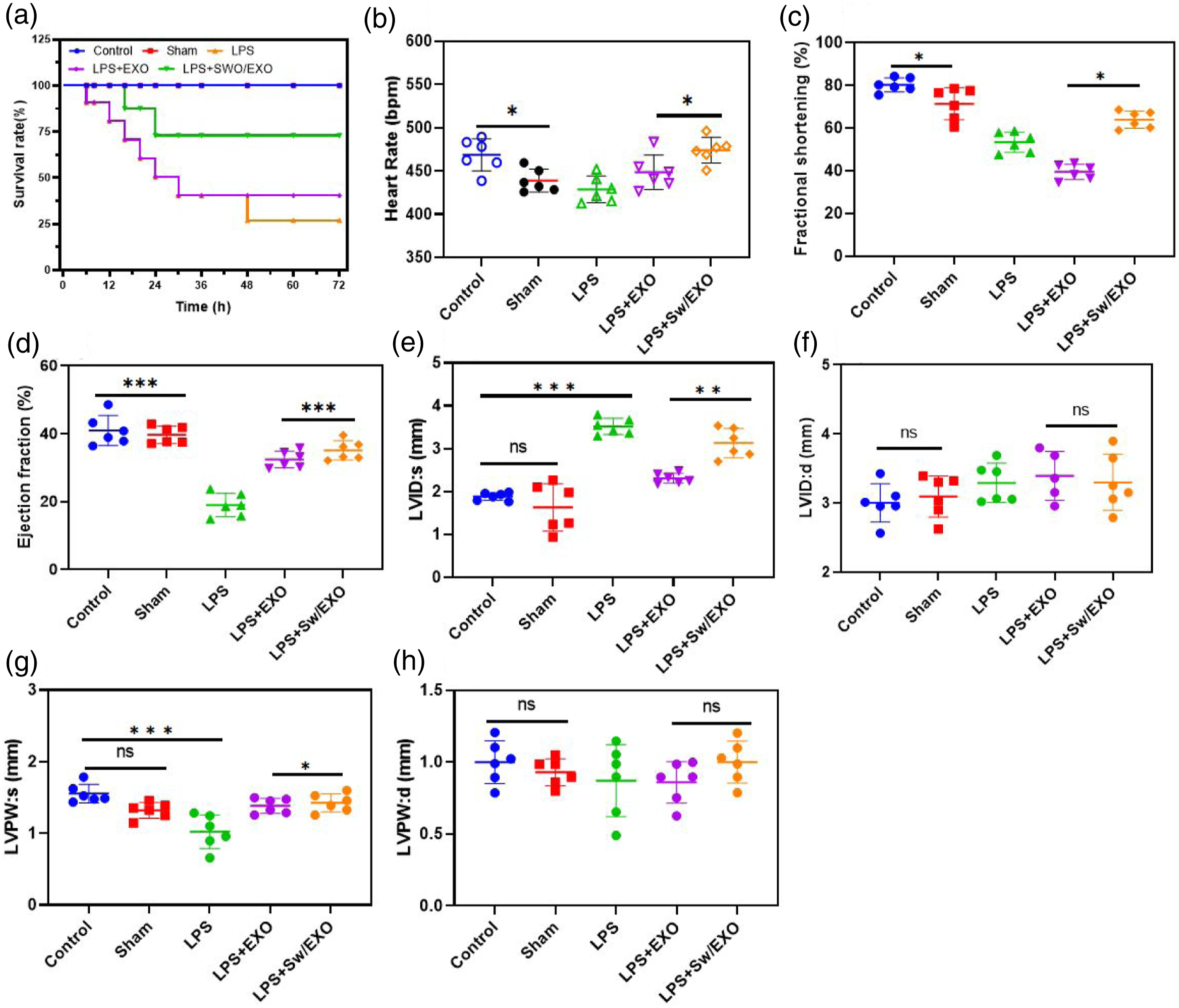
In an LPS-induced SMI model, the current work investigated the therapeutic benefits of SWO loaded EXO vesicles. According to the previous findings, SWO/EXO vesicles reduced the systemic oxidative stress and inflammation induced by sepsis and lead to diminish myocardial damage. 21 Severe infections from a variety of pathogenic causes might lead to sepsis, a disease defined by a systemic inflammatory response. Myocardial damage and heart failure are among the most frequent sequelae of sepsis and one of the main risk factors influencing the outcome of severe sepsis, respectively.28,29 Multi-organ injury and dysfunction are prominent complication of sepsis. Recently, biomedical researches focus on reducing myocardial damage in severe sepsis and preserving heart function.30,31 Several medications have been shown to be beneficial against myocardial injury by reducing systemic oxidative stress and inflammation as a consequence of increased research on the condition. Salidroside, for instance, can protect against LPS-induced myocardial damage by preventing the ROS-mediated PI3K/AKT/mTOR signalling from being activated. 32 Natural oxymatrine also reduces the expression of pro-inflammatory protein expressions of IL-1 and TNF, inhibiting the Janus kinase 2/STAT3 signalling pathway and reducing myocardial injury brought on by septic shock. 3 Additionally, it has been revealed that several traditional Chinese medications include anti-inflammatory and antioxidant properties.33,34 Therefore, it is crucial to research how natural components from traditional Chinese medicine extracts can prevent sepsis-related cardiac damage.
Conclusion
In vivo and in vitro protection against LPS-induced myocardial injury is demonstrated in this study by SWO-loaded EXO vesicles. SWO/EXO mixture greatly reduced LPS-induced myocardial injury by inhibiting cell apoptosis, oxidative damage and mitochondrial dysfunction instigated by sepsis. Our research has provided preliminary evidence that SWO-encapsulated EXO might protect mitochondria from sepsis-induced myocardial injury. Therefore, a promising treatment approach for avoiding sepsis-related myocardiopathy involves combining native SWO component with EXO vesicles. Future clinical application of the conventional pharmaceutical SWO material in sepsis will require a great deal more research.
Supplemental Material
Supplemental Material - Sweroside functionalized with MS cells derived exosomes attenuates sepsis-induced myocardial injury by modulating oxidative stress and apoptosis in rats
Supplemental Material for Sweroside functionalized with MS cells derived exosomes attenuates sepsis-induced myocardial injury by modulating oxidative stress and apoptosis in rats by Jianghai Wang, Xiaochen Ma, Xuepeng Si and Wang Han in Journal of Biomaterials Applications.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.