
Abstract
Exploring biological “orthogonality” between plant and animal kingdoms provides a unique opportunity to mimic biological features of both kingdoms. This novel approach can provide solutions to problems that animal kingdom faces. In this study, we aimed to develop and characterize the Neolamarckia cadamba leaf-derived cellulose scaffolds (NCCS). Fresh Neolamarckia cadamba leaves were decutinized using an n-hexanes-diethyl ether and decellularized using 5% sodium dodecyl sulfate (SDS) for 120h and 2% sodium deoxycholate (SDC) solution for 48h. The leaves incubated in deionized water were acted as controls. Finally, the leaves were bleached in 4% NaOCl solution for 12 h. Histology, DAPI staining, and SEM study of scaffold showed complete decellularization with significantly reduced DNA quantity. Water vapor transmission rate and swelling percentage increased significantly and the scaffolds were hemocompatible. Contact angle revealed a significant increase in hydrophilicity and at maximum load of 2.83 N, percent elongation at break was 0.094 mm/mm. The FT-IR bands were not disturbed, and average roughness (Ra) and root mean square roughness (RMS) (Rq) of NCCS was 193.27 nm and 153.48 nm, respectively. Specific surface area, pore volume, and pore radius of NCCS were increased. The Madin-Darby canine kidney (MDCK) cells were adhered on NCCS and cell viability on decellularized leaf was 88.18% ± 16.12%. Subcutaneously implanted NCCS revealed infiltrated host cells. These characteristics establish NCCS as a sustainable, ethical and cost-effective substitute to mammalian tissue-derived scaffolds for biomedical applications.
Keywords
Introduction
The application of decellularized plant tissue presents opportunities for the creation of scaffolds for tissue engineering. In recent years, collagen scaffolds derived from mammalian tissues have been tested in rabbits and clinically implanted in veterinary patients,1–4 but use of mammalian tissue leads to ethical concerns and social prohibition. Autologous, artificial and animal grafts currently used for tissue replacement have limitations due to their decreased biocompatibility, limited availability and high cost and ethical issues. 5 There are also worries about the possible spread of disease to host after transplantation of decellularized mammalian tissues. 6 Because of its availability, affordability, biocompatibility, quick ability to degrade and configurable biomechanical qualities, studies have now focused on creating cellulose scaffolds made from plant materials. 7 Surprisingly, even though plants and animals circulate fluids and biomolecules via essentially distinct methods, their vascular systems have significant analogies. The circulatory systems of plants and humans have comparable characteristics. 8 Lignin, hemicellulose, and cellulose, the predominant biological polymer, make up most of the plant cell wall. 9 A stable type of the extracellular matrix (ECM), which makes up much of the cell wall, is cellulose. The structural protein makeup of plant cell walls and animal extracellular matrix is the main distinction between the two. Extracellular matrix in animal cells as well as plant cell walls similarly controls the cell division and expansion. 10 Cellulose present in the decellularized plant material, is compatible with host tissue and easily biodegradable, 11 but studies are going on to find out its in vivo response. The enzymes needed to break down cellulose molecules is lacking in mammalian tissue. However, cellulose could potentially be able to remain at the implantation for long duration due to its resistance to the enzymatic destruction by mammalian cells.12,13
Neolamarckia cadamba leaves have a well-developed vascular network and are mechanically stronger and thicker than the thin and delicate spinach leaves. The improved structural integrity of N. cadamba may allow the leaves to better withstand decellularization processes. Moreover, it has more thickness of leaves, more density of vein and good cellulose-rich matrix providing better mechanical stability, porosity and surface properties which may support cell attachment and soft tissue regeneration.
Decellularization entails treatment of tissues with biological detergents to remove cellular components including proteins and nuclear materials while maintaining the structural strength of the remaining components of the tissue. To generate a scaffold, a range of decellularization approaches can be used, including physical, chemical, and biological ones. 9 Numerous tissues and organs from animals have been successfully decellularized using ionic detergents such as sodium deoxycholate 14 and sodium dodecyl sulphate, 3 as well as non-ionic detergents such Triton X-1001. Plant tissues can be readily and swiftly decellularized to remove the cellular material, leaving behind an extracellular matrix (ECM) structure. Despite the decellularization procedure, cellulose along with other vital cell wall constituents including hemicellulose, pectin, and certain glycoproteins continue to be essential for maintaining structural integrity and tensile strength. The decellularization procedure is confirmed by histological analysis, fluorescent immunostaining, DNA estimation, and scanning electron microscopic examination of samples. The scaffold surface’s physico-chemical characteristics are crucial for cell adhesion and migration. The AFM approach is rapidly emerging as the gold standard for acquiring more precise details on surface and interface features as compared to other imaging methodologies. 15 Contact angle (CA) assessment provides essential details on the wettability of the scaffold surface, which directly affects cell adhesion, and expansion. This modality proves crucial for the construction of biological scaffolds in tissue engineering research, where scaffolds are usually designed with surfaces that are both porous and hydrophilic. The ECM’s ideal pore size is essential for promoting cell penetration in the scaffold. Specific surface area and pore distributions can be determined using Brunauer-Emmett-Teller (BET) surface area analysers. For the cytocompatibility analysis, Madin-Darby canine kidney (MDCK) epithelial cells were chosen since MDCK epithelial cells are a good model for epithelial cells and closely mimic the adhesion and barrier properties that are critical for tissue engineering matrices. 16
Therefore, in this study, N. cadamba leaves were selected for their distinctive morphological and mechanical characteristics and decellularized using sodium dodecyl sulphate (SDS) and sodium deoxycholate to prepare leaf-based cellulose scaffold for soft tissue engineering and regenerative medicine.
Materials and methods
All the reagents used in this study were supplied by Sisco Research Laboratories (SRL Pvt. Ltd, Mumbai, India) unless otherwise mentioned.
Collection, decutinization and decellularization of N. cadamba leaves
Native Neolamarckia cadamba leaves were incubated in a solution of n-hexanes and diethyl ether (9:1) for 10 min to eliminate the cuticle. After rinsing with sterile deionized water, the leaves were decellularized using 100 ml of 5% sodium dodecyl sulfate (SDS) under constant agitation at 180 rpm for 120 h at room temperature. Subsequently, the samples were incubated in 2% sodium deoxycholate (SDC) solution for 48 h. At the same time, dewaxed native leaves (control) were also soaked in sterile deionized water for 168 h. Dewaxed leaves were used as the control throughout the study. The decellularized leaves were then carefully cleaned 6 times with deionized water for 20 min each under continuous agitation on orbital shaker (150 rpm) at room temperature to eliminate residual decellularizing agents and cellular debris. Both dewaxed native and decellularized leaves were eventually bleached in a 4% sodium hypochlorite (NaOCl) solution for 12 h at room temperature.
Characterization of the decellularized scaffolds
Gross observations
The coloration, texture, puffiness, and elasticity of control and decellularized N. cadamba leaves were investigated grossly.
Microscopic observations
Hematoxylin and eosin (H&E) staining was used to observe the microstructures of both native (control) and decellularized N. cadamba leaf samples. The samples were fixed in 10% formalin for 24 h, paraffin blocks were prepared, 6 μm sections were cut and fixed on a glass slide. The sections were cleared in xylene, rehydrated by gradually lowering the ethanol concentration from 99.99% to 30%, followed by a wash with distilled water and immersion in hematoxylin for 7-8 min. After washing, the slides underwent counterstaining with eosin dye for 2 min, were dehydrated by increasing the ethanol concentration, cleared in xylene, and subsequently mounted for microscopic observations.
Scanning electron microscopy
Scanning electron microscopy (SEM) was performed to examine the surface topography of both the native and decellularized N. cadamba leaf scaffolds using a CaRl-Zeiss model EVO-18. The samples were fixed in 2.5% glutaraldehyde, subsequently processed through a series of graded ethanol, and then fully dehydrated by immersing them in hexamethyl disilazane (HMDS). After dehydration, the specimens were coated with a thin layer of gold-palladium at 180°C (15 mA for 5 min). The ultramicrographs were captured from various positions at 10 kV. Additionally, the dimensions of the stomatal pores, including length (nm), width (nm), and area (nm 2 ), were measured from the SEM images to assess the stomatal pore size.
DAPI (4,6-diamidino-2-phenylindole 2HCl) fluorescent immunostaining
The native (control) and decellularized sections of N. cadamba leaves were first rehydrated and then rinsed thrice with 0.2% Tris buffered saline Tween-20 (TBST). The tissue sections were stained with DAPI (Elabscience® Buffer saline solution, India), incubated in the dark at 37°C for 15 min and examined for cold blue fluorescence under an inverted fluorescent microscope (AE31 Elite model, Motic, China).
Measurement of leaf thickness
The thickness of native (control) and decellularized N. cadamba leaves (n = 6) were measured using digital vernier calipers (Themisto Vernier Calliper, TH-M61).
DNA quantification
DNA extraction of both the native (control) and decellularized N. cadamba leaf (n = 3; 50 mg each) was carried out BioLit plant DNA extraction mini kit (SRL Pvt. Ltd). The dsDNA extracted from both the tissues was subsequently quantified using spectrophotometry (NanoDrop ND1000 spectrophotometer (Thermo Scientific, USA).
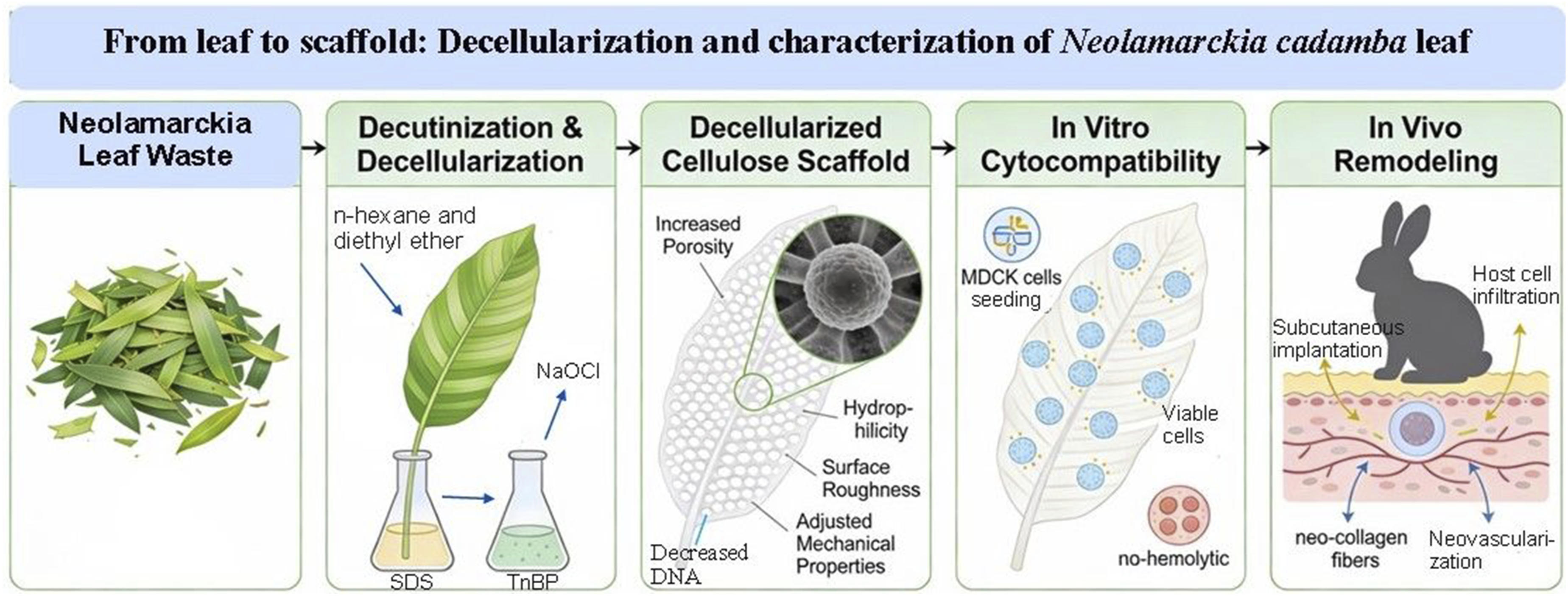
Water vapor transmission rate (WVTR)
Water vapor transmission rate of dewaxed native (control) and decellularized Neolamarckia cadamba leaf tissues (n = 6) was performed.
17
Each house received a specific volume of deionized water, and the samples of size 20 × 20 mm2 were incubated at 37°C and relative humidity of 40 %. The initial weight of the leaf’s samples and weight after 24h were measured. WVTR was computed from the following formula.
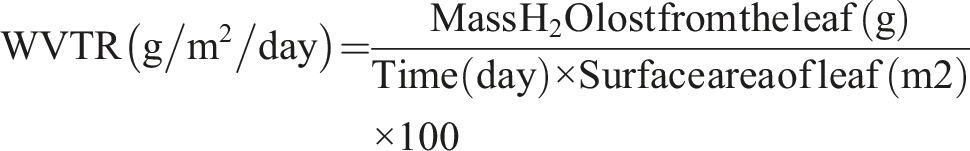
In vitro hemocompatibility
In vitro hemocompatibility of the dewaxed native and decellularized Neolamarckia cadamba leaf tissue (n = 4) was carried out as per the protocol developed previously.
4
Briefly, the leaf scaffolds of size 1 cm2 were exposed to anticoagulated RBCs dissolved in normal saline (NS) in siliconized test tubes. Negative control tubes contain 10 mL NS only and distilled water acts as positive control to induce maximal lysis of erythrocytes. All test tubes were incubated in the water bath for 1 h. Thereafter, the test tubes were centrifuged at 100 × g for 5 min and the O.D. of the supernatant was noted at a wavelength of 540 nm using UV-VIS single beam spectrophotometer (Model LT-291, Labtronics, Haryana, India). The percentage of hemolysis was calculated using the following formula:
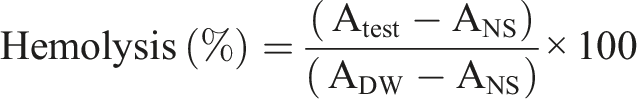
Swelling property
The native (control) and decellularized Neolamarckia cadamba leaves (n = 4) were assessed for water absorption.
18
First, the weight of the samples was measured and then the samples were placed in a tube containing 5 ml PBS at a constant condition (37°C, humidity 35%). The samples were immersed for 1, 3, 24, 72, and 240 h and then weighed. The percentage of swelling ratio was computed using the following formula:
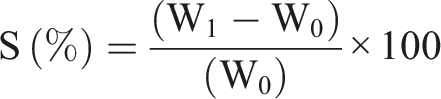
‘S’ represents the percentage of swelling, W0 weight of the dry sample and W1 is the weight of the sample after swelling.
Contact angle analysis
The hydrophilicity of both the native and decellularized N. cadamba leaf samples (n = 3) was measured using the sessile water drop contact angle method with KRUSS Advance, Drop Shape Analyzer (model DSA25). Freeze-dried samples of sizes 10 × 10 mm2 were placed and secured to the transparent glass slides using transparent two-sided tape to ensure stability. The angle formed between a droplet of deionized water, suspended from the tip of a microliter syringe, and the surface of the scaffold was digitally captured using Image-J software. 19
Mechanical test
The native and decellularized N. cadamba leaves tissues (n = 3) of size 30 × 20 mm2 were taken for measurement of tensile strength using a tabletop universal testing machine (Model-EZ50, LLOYD, AMETEK Inc.) equipped with a 1 kN load cell and a crosshead speed of 5 mm/min. The leaf sample was clamped, and uniaxial tensile force was employed till rupture of leaf. The resulting load (N) versus displacement (mm) value was captured for leaf samples. At the rupture points, values for maximum tensile stress (MPa) and strain were documented. Load and displacement were normalized against the cross-sectional area and initial gauge length, respectively, to create stress-strain curves. Young’s modulus was determined from the linear sections of these curves.
20
Based on the load-displacement relation that was monitored during the test, the stress (σ) and strain (ε) were calculated by using the following formulas
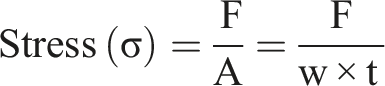
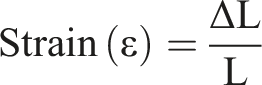
Fourier transform infrared (FTIR) spectroscopic analysis
Fourier transform-infrared spectroscopic analysis of the control and decellularized N. cadamba leaves was carried out using FT-IR spectrophotometer (PerkinElmer, USA) and the software PerkinElmer (Spectrum IR version 10.7.2 model). Every spectrum was captured between the 4000–450 cm−1 range at a resolution of 4 cm−1.
Atomic force microscopy
The surface morphology of the freeze dried native and decellularized N. cadamba leaf sample (10 × 10 mm2) was also characterized by atomic force microscope (Oxford Instrument, Asylum research AFM, ARC2 Controller, Tapping mode, MFP3D). Cantiliver/probe model-PPP-NCLAu-10 with resonance frequency 146-236 KHz was used. The leaf tissue was mounted on transparent glass slides and surface roughness of the leaf tissues was studied as a root mean square (RMS) and arithmetic mean (Ra) value from the AFM images with an area of 5 × 5 μm2.
Brunauer-Emmett-Teller (BET) analysis
Surface area (m2/g), ratio of pore volume to weight (cc/g) i. e pore volume and pore radius (Å) was measured using Brunauer–Emmett–Teller (BET) instrument (Quantachrome® AsiQwin™© 1994-2012, Quantachrome Instruments v3.01). Origin-Pro software was used to assess the collected data. Brunauer–Emmett–Teller (BET) analysis was performed by applying nitrogen adsorption/desorption isotherms at bath temperature −196°C. First, scaffolds were placed in a degassing machine (Autosorb iQ Station 1) at a temperature of 120°C for 3.1h. Nitrogen as adsorbate and Neolamarckia cadamba leaf tissue as adsorbent was taken. Nitrogen molecular cross-sectional area was assumed to be 16.20 Å and liquid density was 0.808 g/cc21. The leaf sample weight was taken 0.08 g and pore volume was determined from the high P/Po area of the isotherm where volume of nitrogen adsorbed is constant. Barrett-Joyner-Halenda (BJH) and Dollimore Heal (DH) method of cumulative adsorption–desorption isotherm curve was utilized to evaluate the cumulative specific surface area (m2/g), ratio of pore volume to weight (cc/g) and pore radius (Å). Dubinn-Radushkevic (DR) method was utilized to determine the micropore area (m2/g), micropore volume (cc/g), micropore half pore width (Å). Density Functional Theory (DFT) method was employed to assay the cumulative surface area, cumulative pore volume and pore radius (Mode). Horvath-Kawazoe (HK) and Saito-Foley (SF) methods were used to estimate the micropore volume (cc/g) and pore radius (Mode) (Å). Dubinin-Astakhov (DA) method was used to calculate only pore radius (Mode) (Å) of sample. 22
Seeding of cells on leaf scaffold
The hydrated native and decellularized leaf were cut into discs (n = 3) with a radius of 3 mm and treated with 70% ethanol for 5 min. The discs were inserted into the wells of an untreated 96-well tissue culture plate and exposed to UV rays overnight. The Madin-Darby canine kidney (MDCK) cells were diluted in low glucose Dulbecco’s Modified Eagle’s Medium (DMEM; HiMedia) supplemented with 10% FBS and 1% penicillin-streptomycin and a total of 15 × 103 cells were seeded onto the surface of the leaf scaffolds, and incubated at 37°C in 5% CO2 for 72 h. Thereafter, the scaffolds were observed under epifluorescence microscope and then scaffolds were fixed with 2.5% glutaraldehyde and 10 % formal saline for SEM and histopathological study, respectively.
MTT cell viability assay
To exclude any cytotoxic effect that may result from leachable chemical residues or DNA leftover, hydrated native and decellularized leaf scaffolds (n = 3) were cut into small round shape pieces to fit into the wells of a 96 well plate and exposed to UV rays overnight. The Madin-Darby canine kidney (MDCK) cells were diluted in low glucose Dulbecco’s Modified Eagle’s Medium (DMEM; HiMedia) supplemented with 10% FBS and 1% penicillin-streptomycin and seeded onto the surface of the leaf scaffolds at a concentration of 15 × 103 cells/well, and incubated at 37°C in 5% CO2 for 48 h. In parallel, the MDCK cells cultured in standard DMEM were used as a control. Thereafter, MTT solution (100 µl) was added into each well and incubated at 37°C in 5% CO2 for 3 h. Now DMSO was added to each well to dissolve the formazan crystals and gently mixed on orbital shaker and again incubated at 37°C in 5% CO2 for 30 min. Absorbance of formazan product was measured at 570 nm using Multiskan SkyHigh Microplate Spectrophotometer (Thermo scientific, USA).
In vivo biocompatibility evaluation
The Institutional Animal Ethics Committee approved the proposed experiment (reference no. IAEC/CVSc-ANDUAT/2025/1/22). A total of 04 animals were included in this study. Following aseptic preparation, both native and decellularized leaf scaffolds (4 samples each) were implanted subcutaneously on either side of the dorsum of New Zealand white rabbits under anesthesia induced by xylazine hydrochloride (6 mg/kg I/M) and ketamine hydrochloride (60 mg/kg I/M). 23 The leaf tissue was secured in position by applying an interrupted suture using sterile non-absorbable monofilament nylon 0-1. Tissue samples were collected on days 20 for histopathological analysis using H&E, Masson’s trichrome, and DAPI stains. Additionally, the ultrastructure of the biopsy tissue was observed by evaluating SEM images using Image-J software.
Statistical analysis
The data analysis was accomplished by SPSS software version 18.0 (SPSS, Inc., Chicago, IL). A one-way ANOVA (analysis of variance) was used to compare the mean values of control and the treatment group.
Results
Gross observations
The mixed solution of normal hexane and ethoxy ethane (9:1) entirely decutinized the leaves of N. cadamba within 10 min. After decutinization, the leaves transformed into a bright green hue. Following treatment with SDS (5%) for 120 h, SDC (2%) for 48 h, the color of the leaves gradually changed to brown as chlorophyll and intracellular contents were disrupted and removed by the detergent treatment. However, the native leaves (control) turned into a greenish-brown color due to natural content of chlorophyll pigments along with phenolic and other cellular components which increased with age. After bleaching with 4% NaOCl solution for 12 h, chlorophyll was entirely removed from the decellularized leaves and the leaf was transparent, indicating substantial elimination of cellular materials and pigments while preserving the underlying cellulose-rich vascular architecture. However, decolorization in the native (control) leaves was incomplete and leaves were milky white in appearance. The petiole was brown, but the color of the central leaf vein faded gradually towards the tip of the leaf (Figure 1). Gross observation of native (a,b) and decellularized (c,d) Neolamarckia cadamba leaves scaffold prepared using 5% SDS (120h), 2% SDC (48h) and depigmented by 4% sodium hypochlorite (12h).
Histological observations
The micrographs of decellularized Neolamarckia cadamba leaves stained with H&E showed entire removal of cuticle from both the adaxial and abaxial surfaces of the epidermis. The leaves underwent thorough decellularization with minimal cellular damage, resulting in a well-structured, nucleus-free palisade tissue. The architecture of leaf tissue was well preserved post-treatment. The absence of purple-blue dots and chlorophyll in the stroma of the treated leaves indicates complete decellularization (Figures 2(a) and 2(b)). Micrograph of native (a) and decellularized (b) Neolamarckia cadamba leaves scaffold (H & E; 20X; Scale bar: 200 µm). Cu– cuticle, ade– Adaxial epidermis, abe-Abaxial epidermis, pp-palisade parenchyma, sp-spongy parenchyma, st-stomatal pore, vb-vascular bundle, mp-mesophyll; Ultrastructure of native (c) and decellularized (d) leaves. Scale bars: 10 µm. DAPI fluorescent stained micrographs of native (e) and decellularized (f) leaves. Native leaves showed cold blue fluorescence indicative of intact nuclei.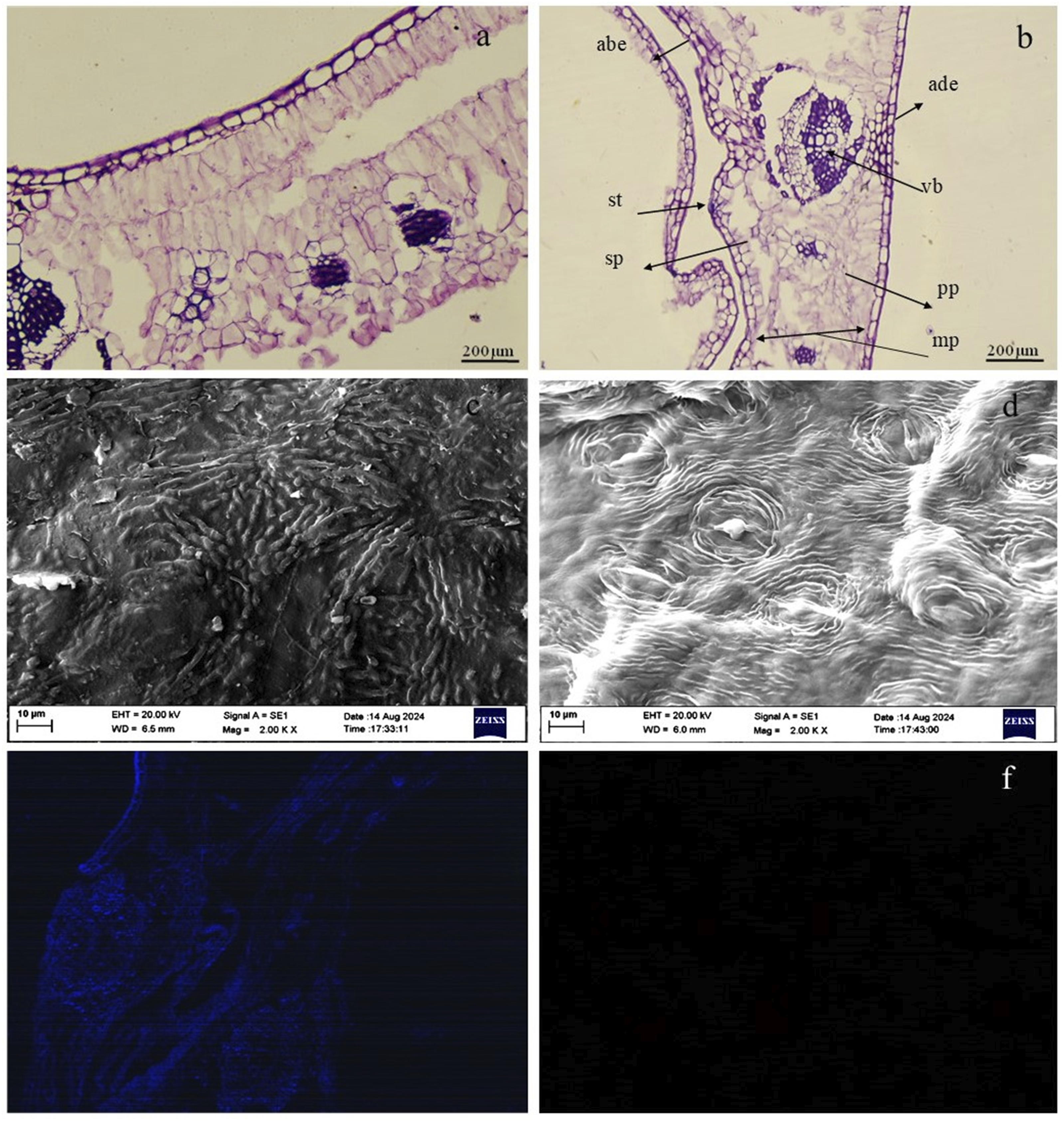
Scanning electron microscopy
The ultrastructure of the native leaf (control) showed the epidermal cells with the majority of the stomatal pores covered by cuticular wax (Figure 2(c)). In contrast, following the decellularization process, the cuticular wax was entirely removed, while other morphological structures remained intact with clearly visible guard cells and stomatal pores, although nuclear content was absent (Figure 2(d)). The topographical structure of the N. cadamba leaf tissue showed non-significant alterations after decellularization. The decellularized leaf scaffolds displayed a well-organized vascular system with increased porosity compared to the native leaf (control) tissue. Mean ± SE of stomatal pore length (nm) and stomatal pore width (nm) of decellularized leaf tissues samples (n = 5) was 12.54 ± 0.59 nm and 3.73 ± 0.27 nm, respectively
DAPI staining
The histological outcomes were further confirmed by DAPI fluorescent immunostaining. The decellularized leaf tissues obtained at 120 h interval demonstrated a nearly complete elimination of nuclear components from the native leaf tissues (Figures 2(e) and 2(f)).
Leaf thickness
The leaf thickness (mm) of the decellularized leaf scaffold was significantly (P < 0.05) decreased as compared to native (C) leaf. The thickness of native leaf and decellularized leaf scaffold was 126.70 ± 6.67 µm and 65.00 ± 5.63 µm, respectively (Figure 3(a)). (a) Leaf thickness of native and decellularized Neolamarckia cadamba leaves scaffold (n = 6) was measured using digital vernier callipers. (b) DNA quantity was determined using a BioLit Plant DNA extraction mini kit (p = 0.000; n = 3). (c) WVTR of native and decellularized leaves (p = 0.000; n = 6). (d) Percent hemolysis caused by native and decellularized leaves (p = 0.000; n = 4). (e) Swelling percentage of native and decellularized leaves at different time intervals (p = 0.001; n = 4). (f) Contact angle of native and decellularized leaves (p = 0.000; n = 3). Representative images of contact angle of native (g) and decellularized leaf (h).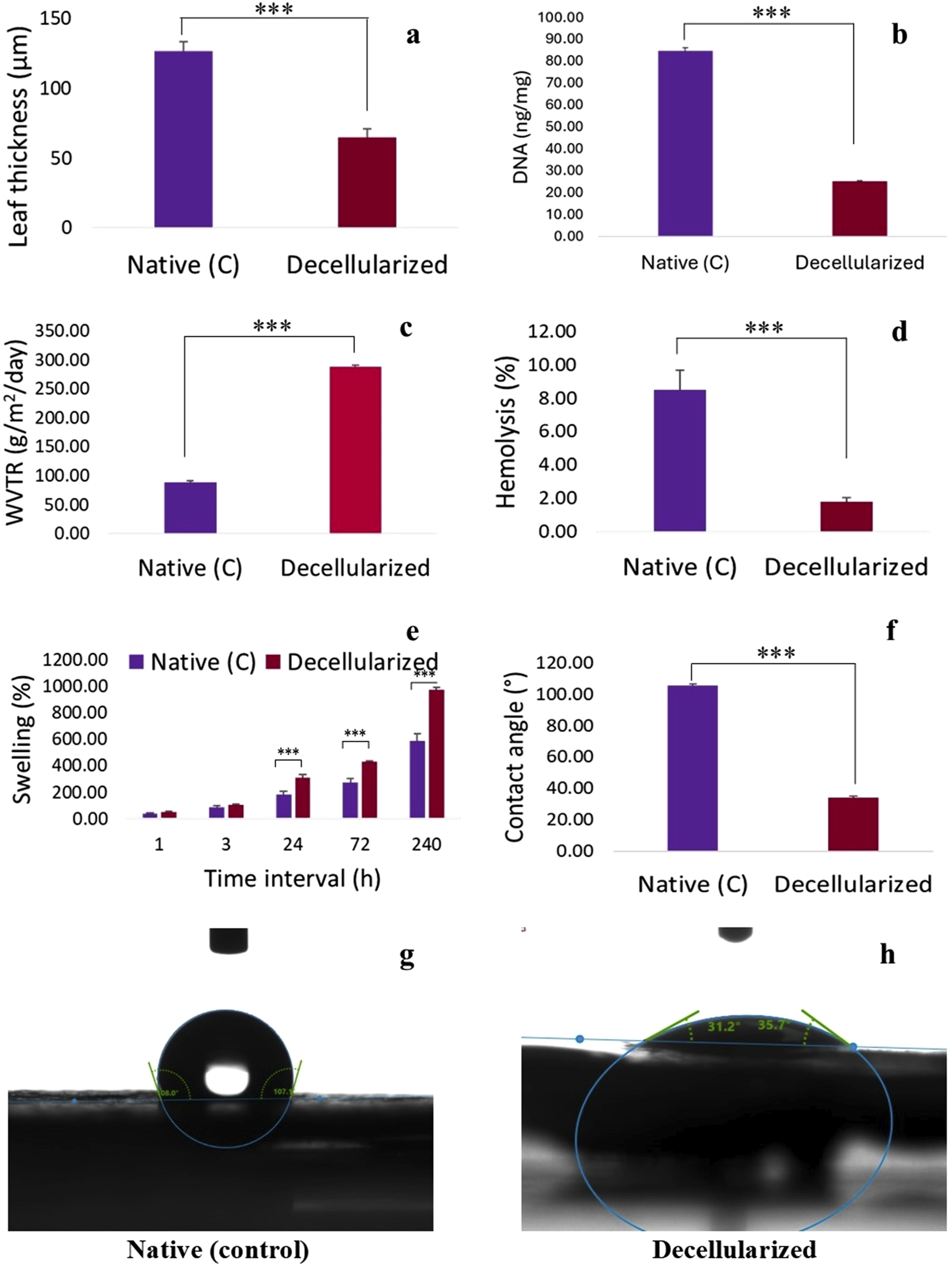
DNA quantification
The quantitative assessment of residual DNA in Neolamarckia cadamba leaf tissues that underwent decellularization indicated a significant decrease in DNA levels when compared to the native leaf tissues. The DNA content of native leaf and decellularized leaf scaffold was 84.93 ± 1.33 ng/mg and 25.31 ± 0.43 ng/mg of tissue, respectively (Figure 3(b)).
Water vapor transmission rate (WVTR)
The observations revealed a significant (P < 0.05) increase in WVTR of the decellularized Neolamarckia cadamba leaves tissue as compared to control group leaves. The WVTR (g/day/m2) of native leaf and decellularized leaf scaffold was 88.44 ± 2.43 g/day/m2 and 287.51 ± 2.66 g/day/m2, respectively (Figure 3(c)).
Hemocompatibility
The hemolysis (%) caused by decellularized Neolamarckia cadamba leaves tissue (1.78 ± 0.27 %) was significantly (P < 0.05) lower as compared to control group leaves (8.50 ± 1.19 %) (Figure 3(d)).
Swelling percent
The swelling percent of decellularized Neolamarckia cadamba leaf tissue was significantly increased (P < 0.05) with increase in time intervals i.e., 1h, 3h, 24h, 72h and 240h when compared with control leaf tissues collected at same time intervals (Figure 3(e)).
Contact angle
The contact angle of decellularized Neolamarckia cadamba leaves tissue was decreased significantly (P < 0.05) as compared to control group leaves. The contact angle of native leaf and decellularized leaf scaffold was 105.65 ± 0.71° and 34.13 ± 0.81°, respectively (Figure 3f–(h)).
Mechanical test
Stress-strain curve of the native and decellularized Neolamarckia cadamba leaves scaffold is illustrated in Figure 4(a) and 4(b). The maximum tensile stress of the native leaves was approximately 0.85 ± 0.00 MPa. In contrast, the maximum tensile stresses for the decellularized Neolamarckia cadamba leaves was 0.59 ± 0.00 MPa which was significantly less when compared to the native leaves (Figure 4(c)). The decellularized Neolamarckia cadamba leaf showed the young’s modulus value of 8.3296 ± 0.001 MPa. However, it was significantly lesser than that of the native (control) leaf samples, which measured 24.49 ± 0.007 MPa (Figure 4(d)). Percent elongation at break for native and decellularized leaf was 0.048 mm/mm and 0.094 mm/mm, respectively (Figure 4(g)). Stress-strain curve of native (a) and decellularized (b) Neolamarckia cadamba leaves scaffold. Tensile stress (c) Young’s modulus (d) and Percent elongation at break (e) of native and decellularized leaves (p = 0.05; n = 3).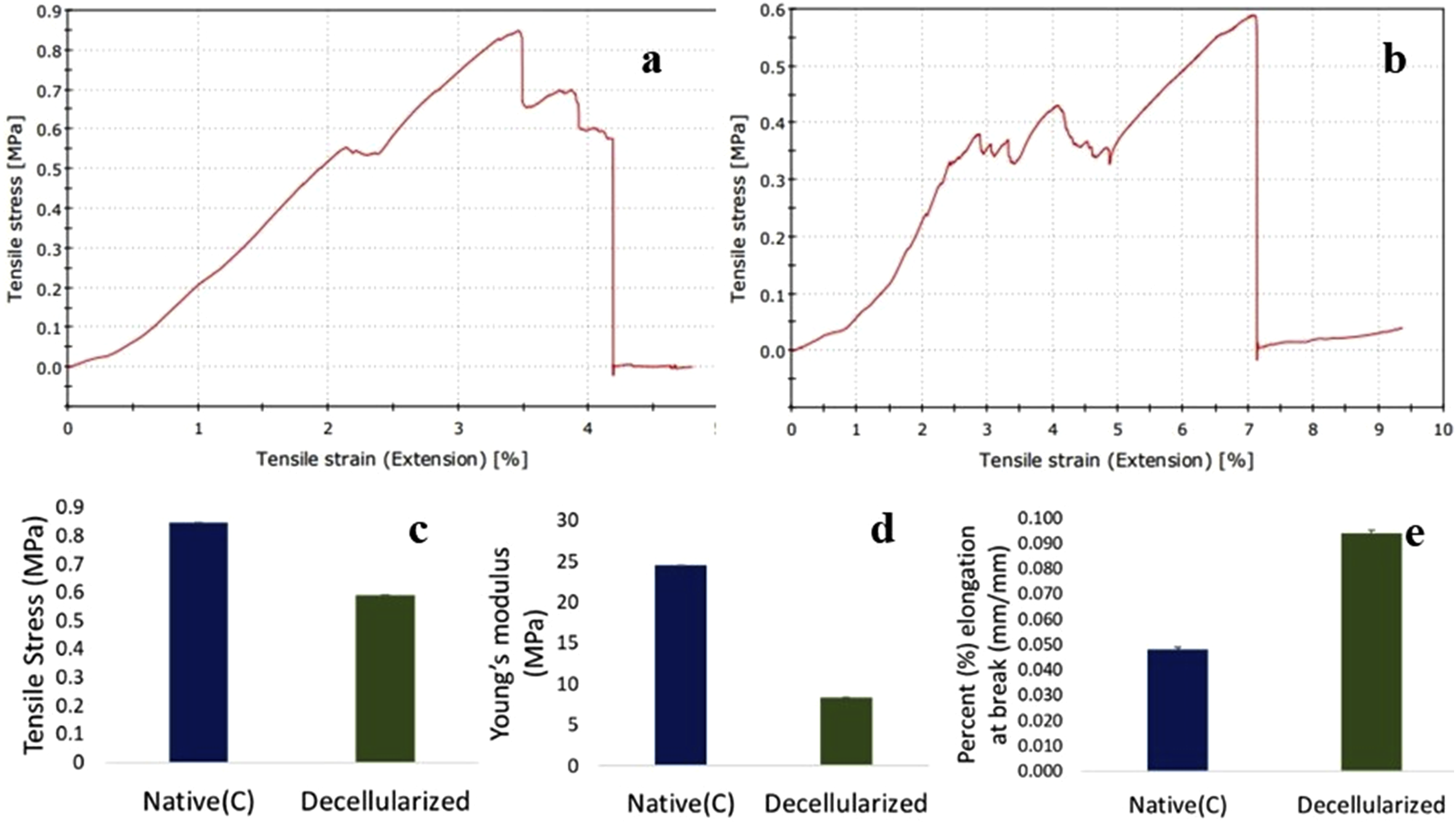
Fourier transform infrared (FTIR) spectroscopy
Fourier transform infrared spectroscopic (FTIR) analysis of control and decellularized Neolamarckia cadamba leaf samples was carried out to compare the alterations in the chemical moiety after treatment with chemicals and detergents. Both control and decellularized leaf samples showed well defined bands between 3384 and 3425 cm
−1
; 2920 and 2957 cm
−1
; 1618 and 1638 cm
−1
; 1385 and 1399 cm
−1
and between 735 and 771 cm
−1
in the spectra (Figure 5a–(b)). The band at 3335 cm
−1
indicated the stretching of H-bonded hydroxy groups intra- and inter molecular hydrogen bonding of cellulose. The absorption band at 2921 cm
−1
is assigned to asymmetric stretching vibrations of the aliphatic C–H bonds (CH2 groups) that exist in the backbone of the cellulose. The fact that this peak persisted after decellularization demonstrates that the plant scaffold has retained its basic polysaccharide structure. The bands observed between 1618 and 1638 cm
−1
in both control and decellularized leaves which indicate aromatic C = C stretch and asymmetric carbonyl (C = O) stretch in carboxyl group of lignin or asymmetric carboxyl group of pectin. The band observed between 1385 cm
−1
and 1399 cm
−1
indicates C–H distortion in cellulose, N = O stretch, CO-H bend (aldehydes), O-H bend (alcohols). The band at 1324.84 cm
-1
was present only in the FTIR spectra of decellularized scaffolds. The absorption band detected at 1324 cm
-1
was assigned to C-O stretching of syringyl (S) units in the lignin structure. Presence of the bands between 1065 and 1081 cm
−1
in both groups corresponds to anti-symmetric stretching of C–C. Presence of the bands between 1027 and 1033 cm
−1
in decellularized leaves corresponds to carbonyl stretching vibrations in polysaccharides and alcohols found in cellulose. The band at 894 cm-1 demonstrate the C-H stretch bond in cellulose and hemicellulose. The bands between 735 and 769 cm
−1
were noted in both the groups which indicated the probable functional groups like C-N stretch (Amines), C-H bend (hydrocarbons) and C-C stretch (Chlorides present in sodium hypochlorite). FTIR spectrum of native (a) and decellularized (b) Neolamarckia cadamba leaves scaffold recorded in the wavenumber range of 4000-500 cm-1.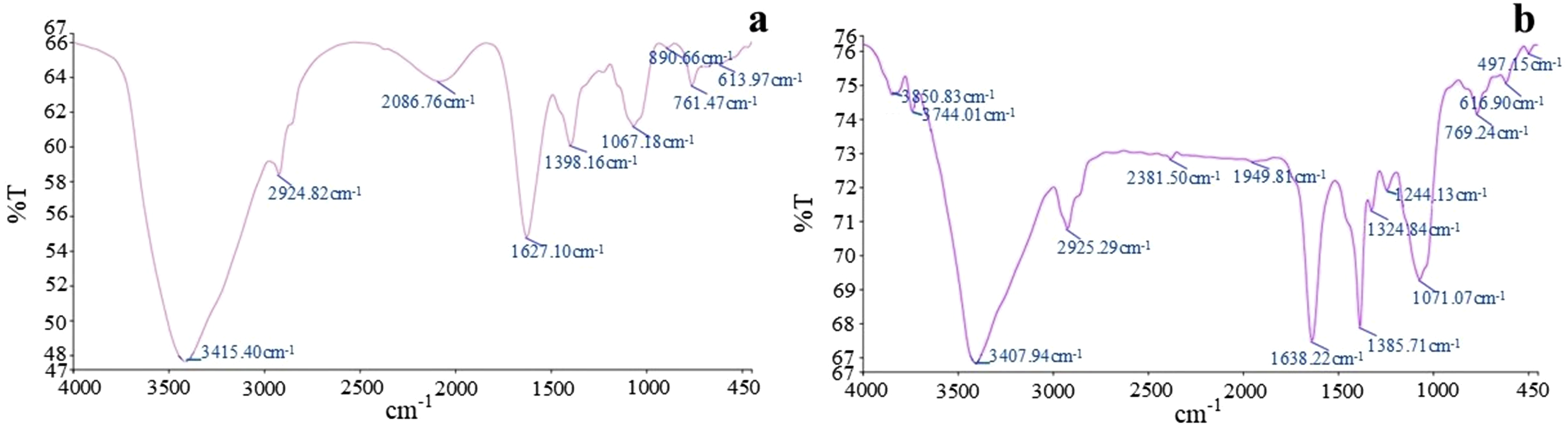
Atomic force microscopy
Atomic-force microscopy (AFM) was done to evaluate the surface roughness characteristics of the native (control) and decellularized Neolamarckia cadamba leaf. The findings indicated that the native leaf sample had fewer, more spaced-out peaks, whereas the decellularized leaf scaffolds exhibited a greater number of peaks that were very close to each other. The surface roughness for both native and decellularized leaf scaffolds was depicted through the bright and dark regions on the images (Figure 6a–(b)). The AFM-derived 3D images clearly illustrate the surface morphology and topography of both native and decellularized leaf samples. The average roughness Ra and root mean square roughness (RMS) roughness (Rq) of the native (control) leaf tissue were 89.28 nm and 113.69 nm, respectively. However, the average roughness Ra and root mean square roughness (RMS) roughness (Rq) of the decellularized leaf scaffold was 191.40 nm and 238.10 nm, respectively. AFM morphological images of native (a) and decellularized (b) Neolamarckia cadamba leaves scaffold. (Scan area 25.5 µm2 and 34.8 µm2, respectively) acquired in tapping mode. The surface shows ridges (bright regions) and valleys (darker regions) which were more pronounced in decellularized scaffolds indicating preserved cellulose microarchitecture after decellularization. The RMS value of native and decellularized leaf was 59.01 nm and 193.26 nm, respectively.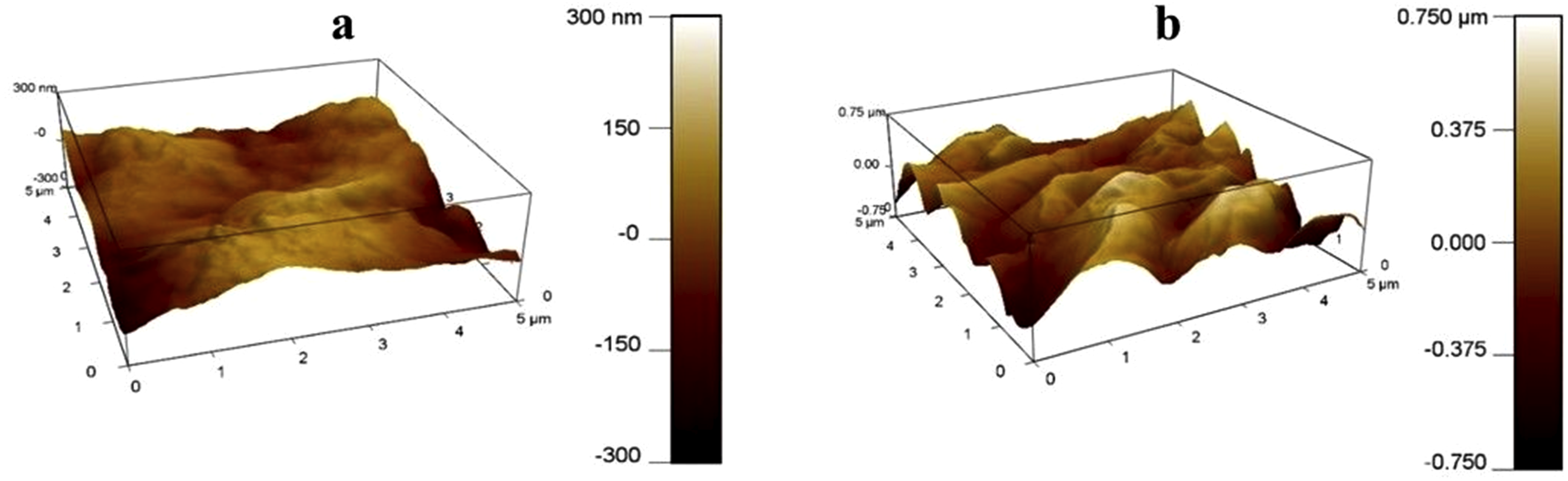
Brunauer-Emmett-Teller (BET) test
Result of the native (control) and decellularized Neolamarckia cadamba leaves are illustrated in Figure 7. A typical adsorption–desorption curve for the control and decellularized The MultiPoint BET test revealed that the specific surface area for control and decellularized Neolamarckia cadamba leaves scaffold was 10.251 m2/g and 19.493 m2/g, respectively. The ratio of total pore volume to the weight of control sample was 0.025 cm3/g for pores having a radius <1857.34 Å at relative pressure (P/Po) value equal to 0.994. However, the ratio of the whole pore volume to the weight of decellularized sample was 0.029 cm3/g for pores radius smaller than 2312.36 Å at relative pressure (P/Po) value equal to 0.996. The average pore radius of control and decellularized leaves matrices was 29.59 Å (2.959 nm) and 49.05 Å (4.905 nm), respectively. The pore radius of control and decellularized Neolamarckia cadamba leaves scaffolds calculated using the Dubinn-Radushkevic (DR) protocol were 43.34 Å and 45.53 Å, respectively. (a-b). Nitrogen adsorption-desorption type IV isotherm and corresponding analysis of Brunauer-Emmett-Teller (BET) specific surface area of control (a) and decellularized (b) Neolamarckia cadamba leaves tissue at multipoint. Type-IV isotherm of control (c) and decellularized (d) leaves tissue. Barrett-Joyner-Halenda (BJH) analysis showed pore volume of control (e) and decellularized (f) leaves tissue at multipoint. BET analysis of pore radius (Å) of control (g) and decellularized (h) leaves tissue at multipoint.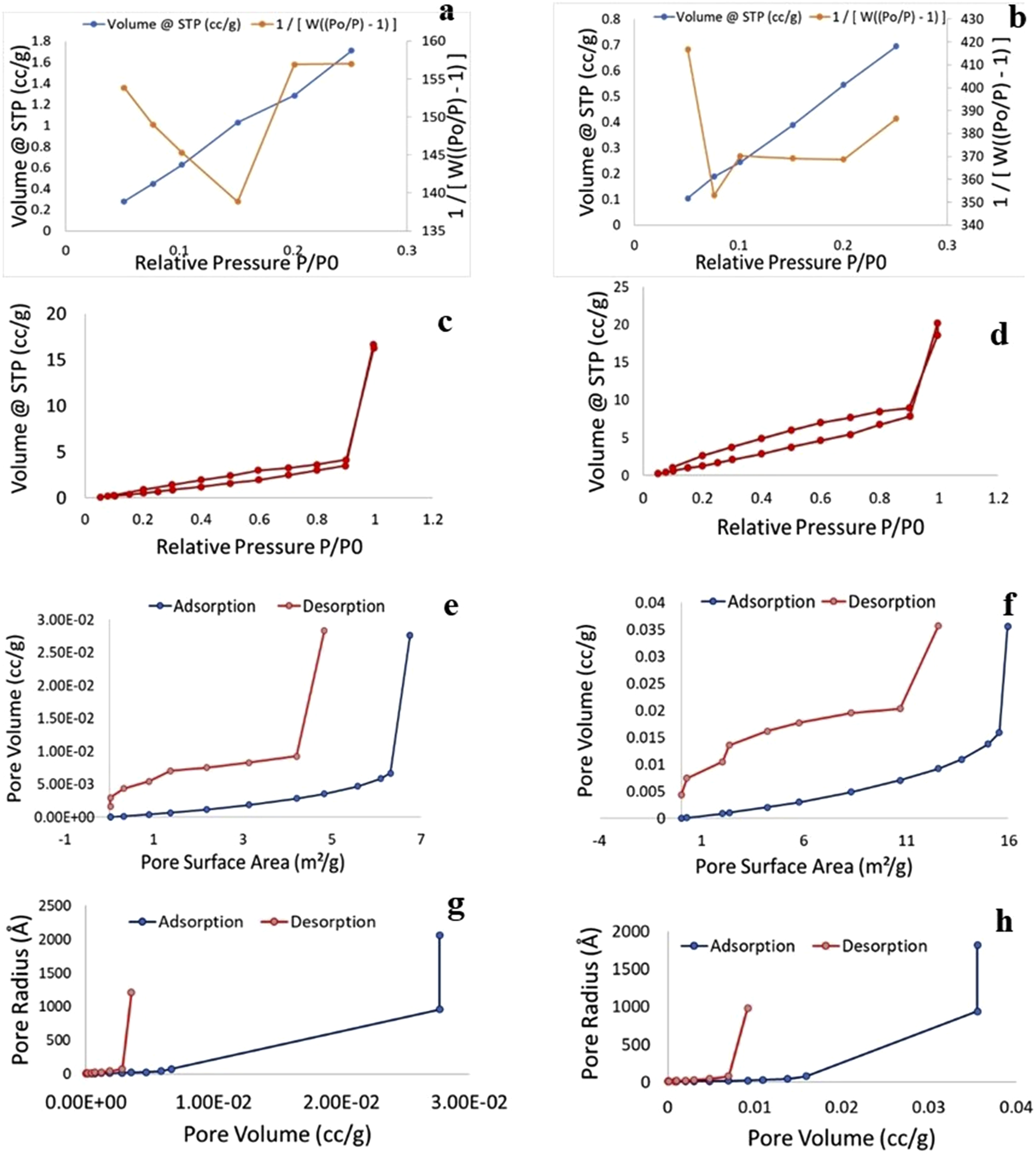
The observations of specific surface area (m2/g) of the control and decellularized Neolamarckia cadamba leaves evaluated by Barrett-Joyner-Halenda (BJH) and Dollimore Heal (DH) method showed a specific type IV adsorption–desorption isotherm graph showing the existence of distinct mesopores. For control leaf sample the cumulative adsorption and desorption values of specific surface area obtained by BJH procedure were 6.75 (m2/g) and 11.61 (m2/g), respectively. Moreover, in decellularized leaves sample the cumulative adsorption and desorption values were 15.95 (m2/g) and 27.05 (m2/g), respectively. For control leaf tissue the values of cumulative adsorption and desorption evaluated by DH method were 6.93 (m2/g) and 12.40 (m2/g), respectively. However, in decellularized tissue the quantity was 6.40 (m2/g) and 29.04 (m2/g), respectively. In control leaves, the external surface area (m2/g) of the pore obtained by the t-method was 10.25 m2/g. However, in decellularized leaves sample its quantity was 16.43 m2/g. The mean value of micropore area of control and leaves sample received using the Dubinn-Radushkevic (DR) method was 14.30 m2/g and 30.75 m2/g, respectively. The cumulative surface area determined using the Density Functional Theory (DFT) approach was 2.78 m2/g and 6.68 m2/g, for control and decellularized leaves, respectively.
Observations of pore volume to weight (cc/g) ratio of the control and decellularized Neolamarckia cadamba leaves received by both Barrett-Joyner-Halenda (BJH) and Dollimore Heal (DH) method showed a typical type IV cumulative adsorption–desorption pore volume isotherm with a specific hysteresis loop. Initial loop depicts single to many layers adsorption and another loop depict desorption. Relative pressure (P/Po) uptake had a substantial limit. Employing the BJH and DH methods, the control sample’s total adsorption and desorption pore volumes were 0.0277 and 0.0284cc/g, respectively. In contrast, the total adsorption and desorption pore volumes for the specimen of decellularized leaves were 0.0356 and 0.0357cc/g, respectively. The Dubinn-Radushkevic (DR) approach yielded 0.00,508 and 0.01,087cc/g for control and decellularized Neolamarckia cadamba leaves, respectively; the Horvath-Kawazoe (HK) method yielded 0.000,604 and 0.001,587cc/g, respectively; and the Saito-Foley (SF) method yielded 0.0000158 and 0.0000142 cc/g, respectively. Using the Density Functional Theory (DFT) approach, the cumulative pore volume of control and decellularized leaves was found to be 0.005,970 and 0.01,297 cc/g, respectively.
The control Neolamarckia cadamba leaves’ cumulative pore radius (Å) for adsorption and desorption, as determined by the Barrett-Joyner-Halenda (BJH) and Dollimore Heal (DH) techniques, was 10.96 Å and 9.04 Å, respectively. In contrast, the total adsorption and desorption pore radius for the specimen of decellularized leaves were 12.29 Å and 9.05 Å, respectively. The Barrett-Joyner-Halenda (BJH) procedure’s pore radius (Å) results for both native (control) and decellularized Neolamarckia cadamba leaves revealed a typical type-IV adsorption–desorption pore radius isotherm characterised by the prominent existence of a hysteresis loop. When the condensation of capillary takes place in a mesopore, a hysteresis loop is observed. Mesopores are defined as having pore sizes between 2 and 50 nm. Single to multilayer adsorption is represented by the first loop, while desorption is represented by the subsequent one. Using the Dubinn-Radushkevic (DR) approach, the micropore half-pore width of control and decellularized leaves was measured; the results were 43.34 and 45.53(Å), respectively. Several techniques were used to assess the pore radius (mode) of control and decellularized leaves. The Dubinin-Astakhov (DA) approach yielded pore radius (mode) values of 16.40 and 17.10 (Å) for control and decellularized leaves, respectively; the Horvath-Kawazoe (HK) approach yielded measurements of 9.24 and 9.69 (Å), respectively; the Saito-Foley (SF) approach yielded measurements of 16.66 and 18.87 (Å); and the DFT approach yielded measurements of 13.85 and 14.48 (Å) respectively. The MultiPoint BET of decellularized leaf revealed increased specific surface area, total pore volume and average pore radius as compared to native leaves scaffolds. It may be attributed to removal of cells and debris from the native leaves without affecting the architecture of leaf during the process of decellularization to develop decellularized leaf cellulose scaffolds. The BET analysis of the decellularized spinach leaf scaffold, that exhibits a high pore volume to scaffold weight ratio (0.06,255 cm/g) and a significant specific surface area (28.540 m/g). The mean pore diameter, total pore volume and specific surface area of the decellularized onion scaffold was 5.8 nm, 0.00,972 Cm/g and 6.636 m/g, respectively and the architectural study revealed that the scaffold provided an even surface and a good specific surface area.
Cells seeded on leaf scaffold
A total of 15 × 103 MDCK cells were seeded onto the surface of leaf scaffolds to investigate whether cells could adhere to and maintain functionality. Decellularized leaf scaffolds provided structural support for the seeded cells to adhere, multiply and differentiate (Figure 8(a)). The ultrastructure of the seeded scaffold showed extensive attachment and proliferation of the cells onto the surface after 48 h of seeding. Most of the scaffold surface was covered with interconnected cells and extracellular matrix (Figure 8(b)). These attributes demonstrated the excellent cytocompatibility of the N. cadamba leaf-based decellularized scaffolds. Representative images (n = 3) of Madin-Darby canine kidney (MDCK) cells after 48h of culture on decellularized grafted plant scaffolds. Cells remained viable and adhered to the scaffold surfaces as demonstrated by the round shaped structures (a); SEM ultrastructure of seeded leaf scaffold showed adhesion and proliferation of cells (b); Cellular viability (%) at the scaffold was measured using an MTT assay. There was a significant (P < 0.05) difference in cell viability on the native leaf and the decellularized leaf scaffold (c).
MTT cell viability assay
The cell viability on the decellularized N. cadamba leaves samples was significantly higher compared to native leaf samples. MTT assay revealed that the native leaf had an adverse effect on MDCK cell viability compared to control culture medium. The viability of cells grown in the native and decellularized leaf seeded media was 74.1% ± 3.88% and 116.2% ± 21.25%, respectively after 4 h. (Figure 8(c)).
In vivo biocompatibility
Grossly, the biopsy sample of native N. cadamba leaves collected from the implanted site on day 20 was surrounded by fibrous tissue. Microscopically, the tissue showed severe inflammatory reaction within the native leaf tissue with dense infiltration of the polymorphonuclear cell like granulocytes (Figure 9(a)). Both axial and abaxial epidermis, palisade parenchyma, spongy parenchyma, chlorophyll, and mesophyll of the leaf were clearly visible. Necrotic areas with cellular debris were identified within the stroma. These findings clearly showed an acute foreign body reaction against the native leaf tissue (C). The decellularized leaves scaffold implanted tissue was infiltrated with host tissue with accumulation of well-organized collagen fibers and neovascularization (Figure 9(b)) which indicates active tissue remodeling. Masson’s Trichrome staining revealed distinction in collagen formation and deposition between groups. The native scaffold displayed irregular and sparse collagen organization (Figure 9(b)), while the decellularized scaffold presented noticeable, blue-stained collagen fibers with improved distribution and maturation suggesting host-derived connective tissue deposition, an important sign of constructive remodeling and integration of the scaffold with surrounding tissue (Figure 9(f)). In the control biopsy, DAPI signal also showed sparse nuclei and leaf debris within the native leaf matrix because it is less conducive to infiltration (Figure 9(c)). In the decellularized leaf implanted biopsy tissue, evidence of spilled DAPI-positive nuclei within the scaffold demonstrates deep host cell infiltration (Figure 9(g)). SEM typically reveals a more compact, less remodeled surface with fewer signs of cellular occupancy in the native tissue (Figure 9(d)). Decellularized biopsy was rougher, with exposed fibrous architecture and possible surface colonization by host cells or extracellular deposits, in line with a scaffold capable of supporting attachment, penetration and remodeling (Figure 9(h)). Micrograph of biopsy samples of native (a) and decellularized (e) Neolamarckia cadamba leaves scaffold transplanted tissue (H & E; 20X; Scale bar: 200 µm). fb– fibroblasts, ep– epidermis, nc-neocollagen, ls-remnants of leaf scaffolds with infiltration of host cells, pp-parenchyma, n-necrosed tissue, pmn-polymorphonuclear cells. Masson’s Trichrome stained native tissue displayed irregular and sparse collagen organization (b), while the decellularized scaffold presented blue-stained collagen fibres deposition and remodeling, and integration of the scaffold with surrounding tissue (f). DAPI signal showed sparse nuclei and leaf debris within the native leaf matrix (c) and decellularized leaf implanted biopsy tissue demonstrated spilled DAPI-positive nuclei within the scaffold indicating host cell infiltration (g). Ultrastructure reveals a less remodeled surface with fewer signs of cellular occupancy in the native tissue (d) and decellularized biopsy tissue showed fibrous architecture and possible surface colonization by host cells and extracellular deposits (h).
Discussion
Plant-based tissue engineering has the potential to contribute significantly to the production of cellulose scaffolds7
The decellularization protocol included 4% sodium hypochlorite, 5% SDS and 2% sodium deoxycholate to ensure elimination of cellular components while maintaining the structural integrity of the plant-derived scaffold. Sodium hypochlorite is a powerful oxidant that removes chlorophyll, nucleic acids and cellular debris, improving decellularization efficiency. 24 SDS is an anionic detergent that is frequently used for tissue decellularization as it is very effective at disrupting the cell membrane and solubilizing intracellular proteins, resulting in extensive cellular removal. 9 However, prolonged treatment with SDS may negatively affect the matrix structure. Thus, sodium deoxycholate was used as a milder ionic detergent to eliminate leftover cellular elements with minimal harm to the cellulose architecture. 27 To the best of our knowledge, this is the first report describing the application of SDC in leaf decellularization. We hypothesized that the milder amphipathic nature of SDC would facilitate elimination of residual cellular debris while minimizing damage to the cellulose structure and vascular network. The effective preservation of scaffold morphology, vascular architecture and physicochemical properties reported in the present study, support this hypothesis and emphasize the potential of SDC as an alternative or adjunct detergent in plant-derived scaffold preparation. The concentrations used (4% sodium hypochlorite, 5% SDS and 2% sodium deoxycholate) were selected based on earlier published methods for plant decellularization and optimized to maintain the vascular structure and physicochemical properties of the Neolamarckia cadamba leaf scaffold.24,28 Simple immersion technique effectively decellularizes the leaves, because of the stomata, which allows for rapid exchange of detergents without requiring a perfusion system. 29 Moreover, processing multiple leaves simultaneously in one container should be avoided, as it may lead to inconsistent decellularization.
The time it takes for a leaf to become transparent can vary from 24 h to several weeks, influenced by factors such as its size, thickness, lobing, and chemical makeup. 24 Apart from this, leaf maturation stage may alter the duration of complete decellularization. The complete absence of nuclei and chlorophyll in the leaf stroma indicates thorough decellularization. 30 The ultrastructure of decellularized N. cadamba leaf tissue exhibited a significant increase in stomatal pore length, width, and area. This enhancement may be the outcome of the removal of the cuticle, cellular components, and barriers, including protective cells around the stomata, during the decellularization process. Following decellularization, the scaffold became more porous. 31 Furthermore, the overall topographical architecture of the specimens remained largely unchanged after applying the decellularization technique. 20
A key aspect of this decellularization process is the removal of nearly all cellular DNA. 32 A significant decrease in the plant DNA content (less than 50 ng double-stranded DNA/mg tissue) and absence of cold blue fluorescence in the DAPI stained tissue sections of the leaf scaffold confirmed a successful decellularization. WVTR is a crucial metric to consider assessing the body fluids lost by evaporation. The elimination of several regulatory barriers, including stomatal protective cells (guard cells) while decellularization and the development of pores after the decellularization procedure, may be the cause of the elevated WVTR. Porosity raises the vapour permeability of scaffolds, which in turn raises the rate of water vapour transmission. While a very low WVTR may result in the buildup of wound exudate, an exceptionally high WVTR may promote dehydration of wound. 33 Under static settings, the dressings’ WVTR should be ranged from 90 ± 10 to 2892 ± 337 g/m2/day. 34
Since hemocompatibility is necessary for any biological tissue to be implanted in the body, the haemolysis investigation was utilised in this work to rule out the immunological response brought on by the existence of any nuclear or cellular material inside the decellularized leaf scaffold. Materials are categorized as haemolytic if they cause more than 5% haemolysis, somewhat haemolytic if they cause less than 5% haemolysis, and nonhemolytic if they cause less than 2% haemolysis. 35 If the biomaterial causes a higher degree of RBC lysis, which could lead to graft rejection and such scaffolds are not suitable for in vivo transplantation. Both synthetic and biological, prostheses shouldn’t cause more than 2% haemolysis. 36 The decellularized N. cadamba leaf scaffolds showed a hemolysis percentage of 1.78 ± 0.27%, which is below the permissible limit, indicating good hemocompatibility. The decellularized samples’ swelling ratio increased because of the increased liquid adsorption rates brought about by the elimination of cell material caused by the decellularization procedure. 37
The decellularized samples’ swelling ratio increased because of the increased liquid adsorption rates brought about by the elimination of cell material caused by the decellularization procedure. Decellularized leek samples swelled over 2.5 times more in PBS and 1.5 times more in DMEM than untreated tissues. 20 Dehydrated decellularized baby spinach leaves could achieve 1696 ± 223% water absorption in 120 min. 37 By virtue of roughness, amorphous frameworks are more permeable to fluids than highly organized crystalline sections. 38
Contact angle is one of the indicators of cell adherence and biocompatibility. The wetting ability of the exterior of the material significantly affects the proteins’ adhesion, which is evidently an intermediate step of cell binding. 39 The contact angle of the decellularized N. cadamba leaf sample was significantly reduced, indicating an enhancement in hydrophilicity. This outcome is consistent with previous research on plant-derived scaffolds, including bamboo stems 40 and cabbage matrices. 22 The resulting hydrophilic environment is critical for tissue engineering, as it actively promotes strong cell adhesion, facilitates the efficient uptake of nutrients and growth factors, and supports optimal cell growth. 41 The expansion of the fibroblasts was considerably suppressed on the “exceptionally” hydrophobic and hydrophilic surfaces, most probably because of the absence of adhesion proteins. Conversely, the fibroblasts that adhered to the modestly wettable interfaces showed linear growth of cells. 42
The decellularization of the leaves using SDS and SDC solution followed by incubation in a 4% sodium hypochlorite solution considerably changes the mechanical characteristics in comparison to native leaf. The decellularization process led to a significant reduction in mechanical strength of the leaf scaffold which might be due to the removal of cellular content during decellularization and subsequent development of void spaces in the scaffolds, which ultimately weakens their mechanical strength. 43 Tensile strength and elastic modulus may be decreased by prolonged contact with harsh detergents, which additionally degrade the ECM architecture and deteriorate its mechanical qualities. The oxidative qualities of bleach, which can further harm cellulose and cause fibre thinning, could be the cause of this effect. 44 The decellularization leads to damage of certain venations and most cellular and nuclear materials, significantly diminishing the mechanical properties of the plant-based scaffolds in contrast to non-decellularized tissues.20,24 For certain plant species, it may reach a comparable range as the majority of human tissues, including soft tissues (1–20 kPa), cortical bone (20 GPa) and muscles (10 kPa).40,44
Lignin, cellulose, and hemicellulose make up most plant cell walls. The bands in the region of protein are related to the different plant proteins that remained in the scaffold and were not removed during the decellularization procedure. 45 The band at 3334 cm-1 is correlated with the stretching of H-bonded hydroxy groups. 46 The alteration of the band at 2900 cm-1, that represents the alkane (C-H) stretching and the decrease in its intensity, lends additional credence to this. 47 The C = O stretching of xylan observed in hemicelluloses or functional groups in the structural makeup of lignin may be represented by the band between 1700 and 1740 cm-1 wavelength. 48 In cellulose and hemicellulose, the band at 1420 cm-1 denoted alkanes (C-H) and hydroxy (O-H) group wag. 20 The band at 894 cm-1 demonstrates the C-H bond stretch in cellulose and hemicellulose. Decellularized Neolamarckia cadamba leaf tissues lack this peak indicating rupture of glycosidic bonds of cellulose and ultimate decline in strength of the scaffold. At the same time presence of “S unit” of lignin, a building block of lignin polymer, in the decellularized scaffolds only may provide the structural support to the decellularized leaf and ultimately increase in tensile strength. 49
For cell expansion, migration, differentiation, and protein absorption, the scaffolds must have a sufficiently rough topography. 45 Scaffolds with roughened surfaces can enhance cell adhesion and interactions. 27 Topographical imaging of scaffold surfaces can be improved by a method that essentially involves eliminating the low-frequency surface undulations. 50 The decellularized leaf scaffolds exhibited a greater number of closely spaced peaks, likely because of the increased surface roughness. The loss of water-soluble pectin during the decellularization process may also disrupt the cell wall structure, contributing to this roughness. The increased value of RMS, arithmetic mean (Ra), and irregular surface geometry of the decellularized leaf tissue may be due to the removal of cuticular wax as well as nuclear and cytoplasmic materials during decellularization. The surface organization of this scaffold precisely corresponds to SEM images and contact angle values. Decellularization of green spinach and walnut leaves had no statistically significant impact on the surface characteristics.22,51 However, in present experiment the roughness of the decellularized N. cadamba leaf scaffolds was increased. This increased surface roughness in the decellularized leaf scaffold may facilitate greater cell attachment, growth, proliferation, and differentiation, thereby promoting the regeneration of the target tissue. 22 The decellularization method may reduce the surface roughness of the decellularized grass matrix. 52
The MultiPoint BET analysis of decellularized N. cadamba leaf demonstrated an increase in specific surface area as compared to native leaf scaffolds which is important for their functionality in bioengineering. The increase in surface area in decellularized tissues is largely because of the removal of cellular material and the preservation of the extracellular matrix’s native architecture. This reflects increased pore volume, porosity and average pore radius which are generally beneficial for host tissue infiltration, cell migration, neovascularization and nutrient and waste flow in scaffold. 53 This increase may be attributed to variations in morphology and anatomical structure between the native and decellularized leaf scaffolds. 45
Although some research indicates that the residual plant proteins like lectins may trigger an early inflammatory response by activating immune pathways. 11 The Madin-Darby canine kidney (MDCK) cells were adhered to the N. cadamba leaf scaffolds on day 3 post cell seeding. The cell viability on decellularized leaf was significantly higher. Cellulose derived from decellularized apple hypanthium can promote the proliferation of mammalian cells. 54 Human umbilical vein endothelial cells (HUVEC) and human mesenchymal stem cells (hMSC) were found to be compatible in decellularized plant scaffold. 24 Remaining vegetal DNA may be the cause of the noticeably decreased viability percentage of cells on the native leaf tissue when compared to the decellularized leaf scaffold. Preliminary studies showed that cells seeded on soft plant scaffolds function significantly differently from cells seeded on conventional, hard tissue culture plastic flasks.20,26 Cancer cells cultured on decellularized spinach leaf matrices showed fewer faster rates of growth and more rounded cell morphologies than those cultured on conventional tissue culture flasks. 44 Further research has shown conflicting findings that did not suggest a difference in cell configuration or proliferation over soft plant and stiff synthetic substrates, despite evidence of changed cell shapes and multiplication rates on decellularized vegetal scaffolds being confirmed.20,26,45 Madin–Darby Canine Kidney cells showed strong adhesion with the cellulose scaffold which included flattened cell bodies and partial cobblestone-like architecture in specific areas. The cells covered the whole scaffold and there was no indication of cellular rounding, detachment, or cytotoxic alterations. Decellularized plants scaffolds have intriguing characteristics such as minimal cytotoxicity, 55 biocompatible, 11 affordable, and a natural architecture that promotes attachment of cells, growth, and differentiation.22,56,57 The plant material interfaces encouraged cell proliferation without exhibiting any cytotoxic effects.26,45
Despite plant-based cellulose scaffolds having several benefits, their restricted degradation may be an obstacle to their use in vivo. The cellulose is not easily broken down in the mammalian body due to the lack of cellulolytic enzymes. 58 Host cells readily populated the NCCS, featuring the buildup of collagen fibers and the development of new blood vessels within the void spaces of scaffolds. The original leaf material gradually absorbed and replaced by host tissue, highlighting the superior biodegradability and biocompatibility of plant cellulose in vivo.11,54,59 Furthermore, mammalian cells not only adhered and survived but also proliferated on these grafted leaf scaffolds without triggering a negative immune response.24,60 Cell growth was primarily observed on the upper surfaces of the scaffolds, with the highest density aligned along the collagen fibers, potentially resulting in uneven outer surfaces of the scaffold. 61 Like other natural and synthetic 3D scaffolds, plant-based cellulose scaffolds demonstrate a capacity to adhere to host cells. 62
Conclusions
Cellulose-based scaffolds derived from leaves of Neolamarckia cadamba (NCCS) were successfully fabricated by way of decellularization with sodium dodecyl sulfate (SDS) and sodium deoxycholate (SDC). The developed scaffolds showed good physicochemical and biological properties. These features underline the potential of NCCS as a low-cost, ethically sourced and sustainable alternative to traditional mammalian tissue-derived scaffolds. NCCS is highly suitable for soft tissue engineering applications, especially as a wound dressing matrix and scaffold for skin tissue regeneration. A limitation of the present study is the absence of complementary cell proliferation analyses. Thus, more investigations with other proliferation markers are necessary to improve the understanding of scaffold-mediated regeneration and long-term host tissue integration.
Footnotes
Acknowledgement
We acknowledge Shraddha Singh, Junior Technical Superintendent BSBE Department; Deepali Ubale, Senior Project Associate, Nanoscience and Nanotechnology in Nanofabrication and Mr S K Agnihotri, Senior Technical Superintendent, Indian Institute of Technology, Kanpur for providing the facilities of SEM, AFM, BET and contact angle.
Ethical considerations
All animal housing and experiments were conducted in strict accordance with the institutional Guidelines for Care and Use of Laboratory Animals at College of Veterinary Science and Animal Husbandry, Acharya Narendra Deva University of Agriculture and Technology, Ayodhya 224,229, Uttar Pradesh, India (reference no. IAEC/CVSc-ANDUAT/2025/1/22).
Author contributions
Sangeeta Devi Khangembam performed experiments; Anil Kumar Gangwar, Sangeeta Devi Khangembam and Vipin Kumar Yadav conceived and designed the study; Sangeeta Devi Khangembam and Anil Kumar Gangwar wrote the manuscript; Sangeeta Devi Khangembam and Anil Kumar Gangwar collected and analysed the data; Sangeeta Devi Khangembam, Sonal Srivastava, Surendra Pratap Chakraverty Sameer Srivastava performed the Cell seeding and MTT assay. Yogendra Singh, Surendra Pratap Chakraverty, Anil Singh and Rajesh Kumar Verma and Prafull Kumar Singh performed the histological study and interpreted the FTIR spectroscopic data; All authors read and approved the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by hon’ble Vice Chancellor of the university.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article [and its supplementary information files].