
Abstract
Three dopamine genes (DAT1, DRD2, and DRD4) have been associated with interpersonal delinquency, aggression, and violence when individuals experience adverse environmental exposures. Guided by the catalyst model of aggression, risk alleles identified in previous studies were hypothesized to be associated with intimate partner violence (IPV) perpetration in the presence of financial stressors, a possible environmental trigger. This hypothesis was tested using weighted, clustered logistic regression with data from the National Longitudinal Study of Adolescent to Adult Health. The direct effects DAT1, DRD2, and DRD4 on IPV perpetration, and the interaction of DAT1, DRD2, and DRD4 and financial stressors on IPV perpetration were assessed. Due to cell size, only White men and women were included in this analysis. Increasing number of financial stressors was associated with increased odds of IPV perpetration, regardless of DAT1, DRD2, and DRD4 alleles. As predicted, increasing number of financial stressors was more strongly associated with IPV perpetration among individuals with high-risk DAT1 alleles, than individuals with low-risk alleles. However, this relationship was inverted for DRD2. Although there was still a significant interaction between DRD2 and financial stressors, individuals with low-risk alleles had higher odds of IPV perpetration in the presence of financial stressors. A similar, nonsignificant relationship was found for DRD4. These findings indicate that these genes may interact differently with environmental exposures and types of violent behavior. In addition, the findings may, if replicated, suggest dopamine plays a different role in IPV perpetration compared with other forms of aggression and violence.
Intimate partner violence (IPV) has significant implications for the mental, physical, and economic well-being of men and women. Approximately, 5% of men and women in the United States experience physical IPV victimization each year (Black et al., 2011). Beyond the trauma and injury associated with physical violence victimization (Wu, Huff, & Bhandari, 2010), many men and women experience lasting physical and mental health issues (Afifi, Henriksen, Asmundson, & Sareen, 2012; Beydoun, Beydoun, Kaufman, Lo, & Zonderman, 2012; Devries et al., 2014; Okuda et al., 2011; Trevillion, Oram, Feder, & Howard, 2012; Vives-Cases, Ruiz-Cantero, Escribà-Agüir, & Miralles, 2011; Weiss, Duke, & Sullivan, 2014). Despite the high prevalence of IPV and the significant health consequences, a small proportion of the IPV literature examines risk and protective factors or prevention strategies for perpetrators. The limited understanding of factors contributing to IPV perpetration limits the development of effective intervention and prevention activities for perpetrators (Bell & Naugle, 2008; Eckhardt, Murphy, Black, & Suhr, 2006).
From the existing literature, it is clear the etiology of IPV perpetration is multifactorial and complex (Bell & Naugle, 2008). Individual personal characteristics may interact with physical and social experiences throughout life to shape the propensity to perpetrate IPV (Bell & Naugle, 2008). Although there are several etiological frameworks for IPV, existing frameworks often oversimplify the antecedents to IPV, and fail to account for recent findings in the field of violence research (Bell & Naugle, 2008), including potential genetic contributions. The potential genetic contribution to IPV perpetration, especially in light of growing evidence on the role of genetic factors in the perpetration of other forms of aggression (Barnes & Jacobs, 2013; Beaver, Gibson, DeLisi, Vaughn, & Wright, 2011; T. J. Chen et al., 2005; Ferguson, 2010; Ferguson & Beaver, 2009; Ferguson, Ivory, & Beaver, 2013; Guo, Roettger, & Shih, 2007; Vaske, Boisvert, & Wright, 2012), is of growing interest to the field of violence research and has the potential to contribute to the understanding of the etiology of IPV perpetration.
Based on a comprehensive literature review, four studies of genetic factors for IPV perpetration were identified (Stuart et al., 2014). Two of the four studies used a twin design, which compares the similarities and differences between pairs of monozygotic and dizygotic twins to determine the relative contribution of genetic and environmental factors (Meaney, 2010). These studies were not able to identify specific genes or the interaction between those genes and the environment to predict violence (Meaney, 2010). However, the studies found genetic factors explained up to 54% of the variance in IPV perpetration, which supported the hypothesis that genetic factors contribute to IPV perpetration (Barnes, TenEyck, Boutwell, & Beaver, 2013; Hines & Saudino, 2004). There have also been two studies of the contribution of specific genes to IPV perpetration (Stuart et al., 2014). In a small study of men with hazardous alcohol behaviors in batterer intervention programs, the Monoamine Oxidase A (MAOA) gene and the serotonin transporter gene (SLC6A4) were significantly associated with more frequent IPV perpetration (Stuart et al., 2014). However, this study did not include any nonperpetrating control participants, so the generalizability of this study to the population was unclear. The second study examined the interaction between alcohol use and a cumulative genetic risk scale created by summing the number of individual alleles associated with higher risk of impulsivity as a result of dopamine processing (Foshee et al., 2015). This study found evidence that the association between the genetic risk scale and adolescent dating violence was significantly different depending participants’ alcohol use (Foshee et al., 2015). Together, these findings provided further support for the genetic contributions to IPV perpetration, at least in the presence of environment risk factors.
Genetic Polymorphisms
Although many genetic polymorphisms have been examined as potential contributors to violence perpetration, three dopaminergic genes (DAT1, DRD2, and DRD4) emerged as particularly promising. Research has found that these genes play a role in the processing of dopamine (Anselme, 2013), a neurotransmitter with significant influence on motivation and learning (Anselme, 2013; Wassum, Ostlund, Balleine, & Maidment, 2011). In addition, variations in dopamine processing have been found to modulate the relationship between exposure to stressors and stress responses (Valenti, Gill, & Grace, 2012). In studies of the functional implications of these genes, DAT1 influenced the dopamine reuptake process in the presynaptic neuron (Fuke et al., 2001), and the 10-repeat allele was associated with changes to the rate of dopamine reuptake in the synapse and increased violence perpetration in young men (T. J. Chen et al., 2007; Guo et al., 2007). Carrying a 10R/10R genotype (two 10-repeat DAT1 alleles) was associated with lower levels of self-control (Li & Lee, 2012; Wright, Schnupp, Beaver, Delisi, & Vaughn, 2012) and deficits associated with impulse control, including attention, cognitive flexibility, and planning (Cornish et al., 2005).
Two dopamine receptor genes (DRD2, DRD4) have also been shown to be associated with violence perpetration (Beaver et al., 2007). As a result of fewer D2 dopamine receptors (Pohjalainen et al., 1998), the DRD2 A1 allele was associated with increased susceptibility to environmental triggers (Bakermans-Kranenburg & van Ijzendoorn, 2011), elevated response to stressful situations (Berman & Noble, 1997), and an increased propensity for violence perpetration (Beaver, Gibson, Jennings, & Ward, 2009; Beaver et al., 2007; Guo et al., 2007). Similar to DRD2, DRD4 was associated with the availability of D4 dopamine receptors (Beaver et al., 2007). In interactions with the environment, the presence of a single 7-repeat allele was associated with reduced response to dopamine (Asghari et al., 1995), poor impulse control (Bakermans-Kranenburg & van Ijzendoorn, 2011), and increased odds of violence perpetration (Beaver et al., 2007; Dmitrieva, Chen, Greenberger, Ogunseitan, & Ding, 2011).
Environmental Exposures
Stress, defined as an interaction between individuals and their environment that results in the perception that the situation exceeds resources and compromises well-being (Lazarus & Folkman, 1984), has been associated with IPV perpetration across the life span (Cano & Vivian, 2001; Capaldi, Knoble, Shortt, & Kim, 2012; Mason & Smithey, 2012; Roberts, McLaughlin, Conron, & Koenen, 2011). Stressors, defined as the specific environmental trigger of the stress response (Selye, 1950), have received less attention in the literature as a potential antecedent to IPV perpetration, although specific forms of stressors, such as relationship conflict, have been consistently shown to be an antecedent to IPV perpetration (Byun, 2012; Elkins, Moore, McNulty, Kivisto, & Handsel, 2013; Langhinrichsen-Rohling, McCullars, & Misra, 2012; Shortt, Capaldi, Kim, & Tiberio, 2013; Whitaker, 2013).
Financial stressors, one potential environmental trigger of IPV (Byun, 2012; Neff, Holamon, & Schluter, 1995; Slep, Foran, Heyman, & Snarr, 2010; Schwab-Reese, Peek-Asa, & Parker, 2016), have been reported as stressful to both individual partners and the couple as a dyad (Mason & Smithey, 2012). Several studies of females have found an association between financial stress and IPV victimization (Golden, Perreira, & Durrance, 2013; Khalifeh, Hargreaves, Howard, & Birdthistle, 2013; Matjasko, Niolon, & Valle, 2013). However, the associations of financial stressors and IPV were less consistent. In one study, increasing perceptions of financial stress were significantly associated with IPV victimization, but potential financial stressors, such as income to needs ratio, were not associated with IPV victimization (Benson, Fox, DeMaris, & Van Wyk, 2003). Another recent study found financial stressors, such as nonpayment of housing or food insecurity, were significantly associated with increased odds of IPV perpetration among both men and women (Schwab-Reese et al., 2016), which may provide preliminary support for the association between financial stressors and IPV perpetration.
Understanding the multifactorial etiology of IPV perpetration is an important foundation for the development and implementation of effective IPV prevention and intervention programs (Bell & Naugle, 2008). Given the dearth of literature on the relationships of specific polymorphisms to IPV perpetration, the purpose of this analysis was to contribute to the understanding of the etiology of IPV perpetration through increased understanding of the interaction between three dopaminergic genes and adulthood financial stressors.
Method
Study Population
Data from the National Longitudinal Study of Adolescent to Adult Health (Add Health) were used for this analysis. Between 1994 and 1995, more than 90,000 students completed an in-school questionnaire. From the in-school sample, a subset of 20,745 adolescents participated in a prospective cohort study from 1995 to 2008 that included four waves of data collection (Harris et al., 2009). Of the original 20,745 participants, 15,701 participants completed data collection during Wave IV, which occurred when participants were 24 to 32 years old (Harris et al., 2009). Further information on the Add Health Study is available elsewhere (Harris et al., 2009).
Measures
Outcome
Participants reported IPV perpetration during Wave IV (2008-2009; Harris et al., 2009). Participants reported, “How often (have/did) you (slapped/slap), hit, or (kicked/kick) [partner’s initials]?” (Harris et al., 2009). Response options included, “Never,” “This has not happened in past year, but happened before then,” and specific numbers of times it occurred in the past year (Harris et al., 2009). Participants were coded as perpetrators if they reported slapping, hitting, or kicking their partner at least once in the prior year, and as nonperpetrators if they did not report these actions in the prior year.
Exposure
The research team collected genetic data for all participants who consented. Further detailed information on the protocol, equipment, genotyping, and data cleaning for each gene is available elsewhere (Harris, Halpern, Smolen, & Haberstick, 2006; Smolen et al., 2013). For this analysis, each allele of DAT1, DRD2, and DRD4 was categorized as empirically associated with violence perpetration (“risk gene”) or not (“nonrisk gene”) based on prior literature on the functional differences and associations with violence perpetration. For DAT1, the 10-repeat allele was coded as high risk and the 9-repeat allele as low risk (T. J. Chen et al., 2007; Guo et al., 2007). Individuals with the 11-repeat (n = 4), 3-repeat (n = 39), 7-repeat (n = 53), and 8-repeat (n = 181) were excluded from the DAT1 analysis because the functional implications of these alleles were unclear (Barnes & Jacobs, 2013). For DRD2, the A1 allele was coded as high risk and the A2 allele as low risk (Beaver et al., 2009; Beaver et al., 2007; Guo et al., 2007; Pohjalainen et al., 1998). For DRD4, the 7-repeat allele and 8-repeat allele were coded as high risk and the 2-repeat, 3.39-repeat, 3-repeat, 4-repeat, 5-repeat, and 6-repeat alleles as low risk (Beaver et al., 2007; Dmitrieva et al., 2011). Based on the literature, each pairs of alleles was categorized as high risk if there was at least one high-risk DRD2 or DRD4 allele and two high-risk DAT1 alleles (Thibodeau, Cicchetti, & Rogosch, 2015).
Interacting variable
During Wave IV, participants reported whether they had experienced each of the following six financial stressors during the prior 12 months: without phone service due to financial reasons; did not pay the full amount of the rent or mortgage due to financial reasons; evicted from housing for nonpayment; did not pay full amount of a gas, electricity, or oil bill due to financial reasons; gas or electric service turned off or oil company would not deliver because of nonpayment; worried that food would run out before there was money to buy more (Harris et al., 2009). As was consistent with prior literature (Halliday Hardie & Lucas, 2010), the total number of stressors reported by each participant was summed with scores ranging from zero to six
Statistical Analysis
In accordance with guidelines provided by the Add Health study research team, all analyses were weighted and clustered to account for the complex survey design. Distributions of polymorphisms differed by race, so all analyses were stratified by race. Given the relative rarity of combinations of the risk alleles, financial stressors, and IPV perpetration, the analysis was limited to White individuals due to insufficient sample size among the remaining racial/ethnic groups.
Univariate analysis was used to estimate the mean number of financial stressors, and the number of high-risk genes. Weighted, clustered simple logistic regression models were used to examine relationship between financial stressors, high-risk genes, and IPV perpetration and weighted, clustered logistic regression models including main effects of each of the dopaminergic genes and financial stressors, and interaction effect of the two variables to estimate the odds of perpetrating physical IPV for each combination of exposure compared with having neither exposure and the odds of perpetrating IPV associated with financial stressors comparing participants in the same dopaminergic risk category. Finally, the predicted probability of perpetrating IPV by each level of financial stressor and risk allele was calculated.
To be eligible for this analysis, participants must have been in a romantic relationship during the past 12 months, answered questions on the exposure, interaction, and outcome variables, and completed genotyping. Participants who were not in a relationship during the past 12 months were excluded because their reports of IPV perpetration occurred more than 12 months prior to the interview. Because financial stressors were measured during the prior year, it would have been temporally ambiguous to include participants who reported relationship experiences that occurred prior to the time frame for financial stressors.
Of the nearly 16,000 participants who completed Wave IV data collection, approximately 11,500 were included in this analysis. Most of the excluded participants (~3,320) did not have a romantic partner in the prior 12 months so could not report experiences with IPV. The remaining participants (~900) were excluded due to missing data for IPV perpetration, financial stressors, or other covariates. However, participants were not removed from the data set. As directed by the Add Health research team, the weights for these participants were set to 0.000001, which included the participants in the weighting process but did not include them in the estimates produced by the model (P. Chen & Chantala, 2014).
Results
Financials Stressors, Polymorphisms, and Perpetration of IPV
The mean number of financial stressors was 0.5 (95% CI = [0.4, 0.5]; Table 1). Approximately three quarters (77.5%, 95% CI = [75.8, 79.2]) reported no financial stressors in the past year. Approximately, 16.0% (95% CI = [14.3, 17.8]) reported one financial stressor and 6.5% (95% CI = [5.0, 7.9]) reported more than one financial stressor. As the number of financial stressors increased, the odds of perpetrating IPV increased 1.34 times (95% CI = [1.26, 1.44]). More than half had two DAT1 risk alleles (55.7%, 95% CI = [53.9, 57.58]) and approximately one third had one DRD2 (38.1%, 95% CI = [36.7, 39.5]) or DRD4 (37.0%, 95% CI = [35.8, 38.3]) risk allele. None of the risk alleles was directly associated with IPV perpetration.
Estimated Percent and 95% Confidence Intervals Financial Stressors and Dopamine Genes, and Their Associations With IPV Perpetration Among White Men and Women.
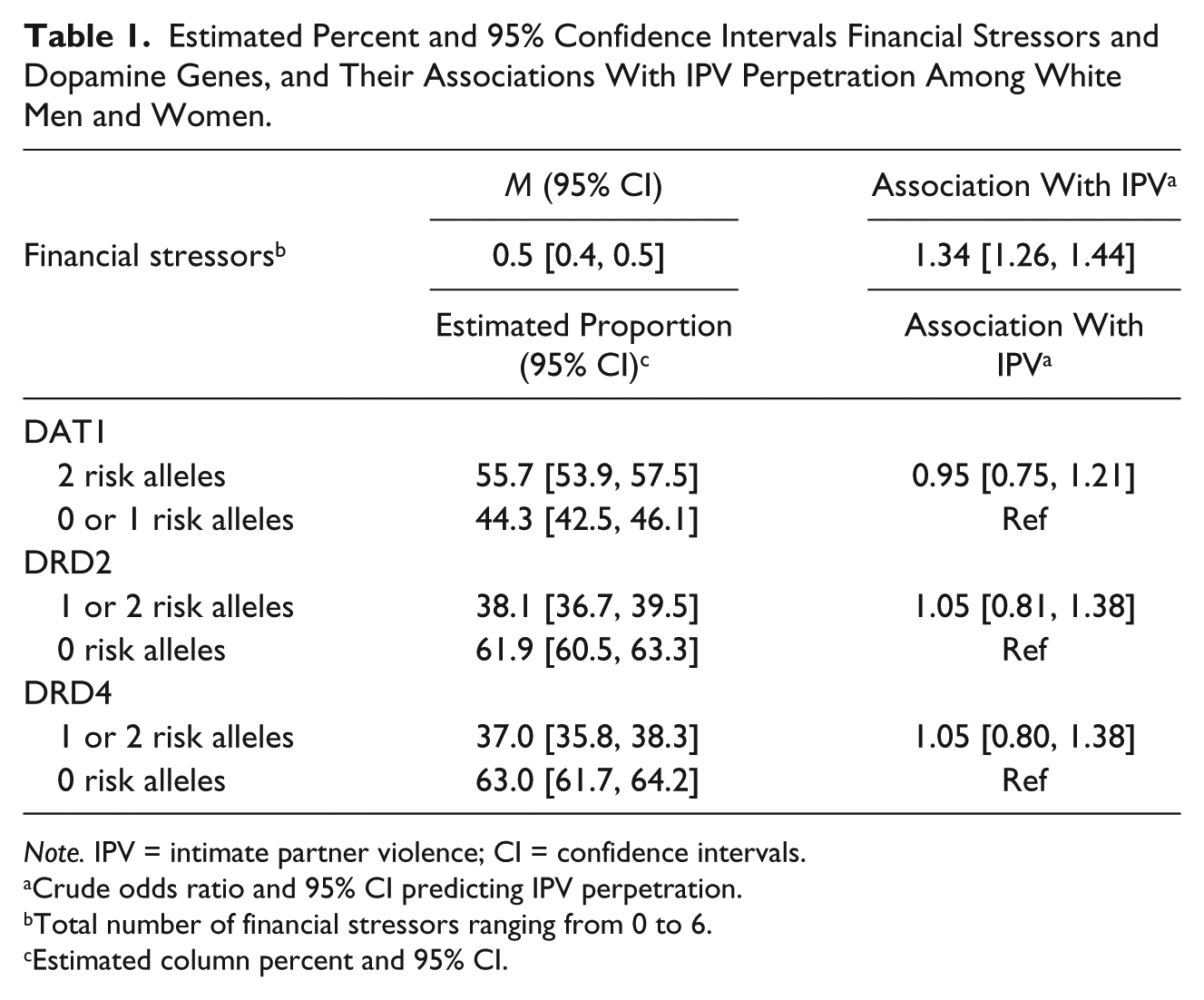
Note. IPV = intimate partner violence; CI = confidence intervals.
Crude odds ratio and 95% CI predicting IPV perpetration.
Total number of financial stressors ranging from 0 to 6.
Estimated column percent and 95% CI.
Interaction of Polymorphisms and Financial Stressors
Increased financial stressors significantly increased the odds of perpetrating IPV across all genes and allele categorizations, when compared with individuals with no financial stressors and no risk alleles (Table 2). However, the magnitude of the increase varied across allele categorizations. Individuals with two DAT1 risk alleles and six financial stressors had 10.07 times (95% CI = [5.07, 20.03]) higher odds of perpetration compared with individuals with no risk alleles and no financial stressors. Among individuals with zero or one DAT1 risk allele, the increase in the odds associated with a greater number of financial stressors was smaller (odds ratio [OR] = 5.11; 95% CI = [2.68, 9.74]). As the number of financial stressors increased, the probability of perpetration (Figure 1) was significantly higher among individuals with two DAT1 risk alleles, compared with individuals with no or one DAT1 risk allele (interaction p = .05). This significant interaction indicated that the presence of financial stressors significantly moderated the relationship between DAT1 alleles and IPV perpetration.
Associations Between Perpetrating Physical Intimate Partner Violence and the Interaction of Dopaminergic Polymorphism and Financial Stressors Among White Men and Women.
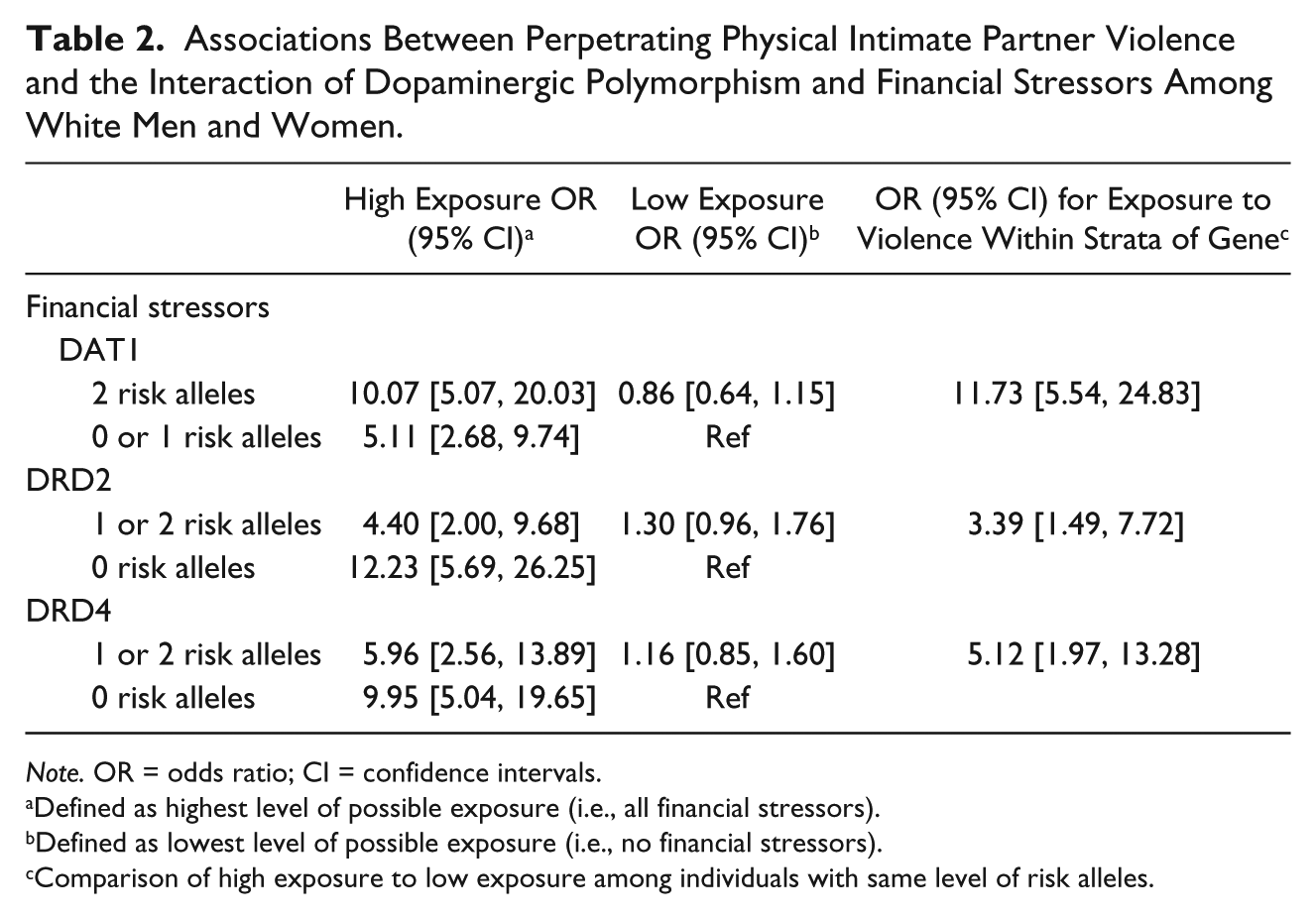
Note. OR = odds ratio; CI = confidence intervals.
Defined as highest level of possible exposure (i.e., all financial stressors).
Defined as lowest level of possible exposure (i.e., no financial stressors).
Comparison of high exposure to low exposure among individuals with same level of risk alleles.
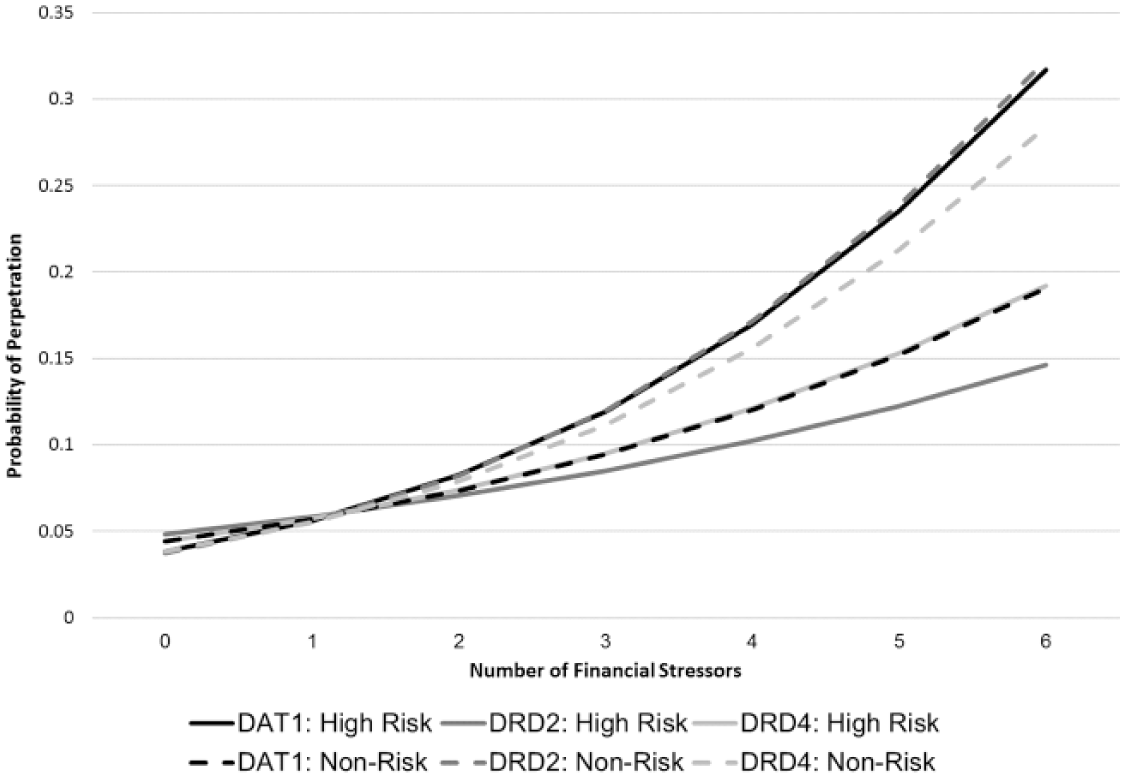
Predicted probability of perpetrating physical IPV, by polymorphism and financial stressor.
In contrast, individuals with one or two DRD2 risk alleles and six financial stressors had 4.40 (95% CI = [2.00, 9.68]) times higher odds of perpetrating IPV compared with individuals with no DRD2 risk alleles and no financial stressors. This increase in the odds of perpetration associated with financial stressors was significantly less (interaction p = .02) than among individuals with no risk alleles (OR = 12.23; 95% CI = [5.69, 26.25]). Although both groups experienced significantly increased probability of perpetration as number of financial stressors increased, the probability of perpetration was significantly higher among individuals with no DRD2 risk alleles compared with individuals with one or two DRD2 risk alleles (Figure 1). Thus, the presence of financial stressors significantly moderated the relationship between DRD2 risk alleles and financial stressors, although in the opposite direction as the DAT1 moderation. The results of the models of DRD4 were similar, however, there was not a significant interaction between financial stressors and DRD4.
Discussion
These findings contributed to the small body of literature on genetic contributions to IPV perpetration (Stuart et al., 2014) and suggested that the etiology of IPV perpetration is complex and multifactorial. Although having DAT1 risk alleles increased the probability of perpetrating IPV as number of financial stressors increased, the associations between DRD2 and DRD4 with financial stressors did not support the initial hypothesis. In fact, having DRD2 risk alleles was associated with significantly smaller increases in the probability of IPV perpetration, compared with peers without DRD2 risk alleles. Although financial stressors significantly increased the probability of perpetrating IPV, the presence of DRD4 risk alleles did not significantly influence that relationship. The results did not indicate that DAT1, DRD2, and DRD4 polymorphisms were significantly associated with IPV perpetration, at least in the absence of environmental experiences. Any contribution of dopaminergic polymorphisms was likely the result of their interaction with the environment and their influence on other factors (e.g., differential response to stress; Stuart et al., 2014). At a minimum, the findings suggested the interactions between dopamine genes and exposure to stressors were better predictors of IPV perpetration compared with dopamine genes alone.
These analyses contributed to the understanding of the specific genetic polymorphisms to IPV perpetration, which, to our knowledge, had only been examined by two previous studies (Stuart et al., 2014). In the initial study of specific genes, the MAOA gene and the SLC6A4 were associated with more frequent IPV perpetration (Stuart et al., 2014). In contrast, the results of this analysis suggested that the dopaminergic genes under consideration do not directly contribute to IPV perpetration. The second study examined the interaction between alcohol use and dopamine genes (Foshee et al., 2015). Similar to this study, this analysis suggested the interaction between genes and environmental experiences, such as financial stressors, was a better predictor of IPV perpetration than genes alone. This gene and environment interaction was consistent with prior findings on contributions of genes to other forms of aggression (Barnes & Jacobs, 2013). In prior research, the association between violence perpetration and a higher number of risk alleles was dependent upon harmful environmental experiences, such as neighborhood disadvantage (Barnes & Jacobs, 2013). However, this relationship was found only for men and not for women (Barnes & Jacobs, 2013) and this study found no significant differences by gender. These results may have implications for the development of IPV prevention and intervention activities, but additional research will be necessary to clarify the role of genetic and environmental interactions. However, financial stressors significantly increased the odds of IPV perpetration, regardless of polymorphism, so interventions to reduce financial stressors may benefit all individuals, regardless of genetic factors.
It is crucial to carefully interpret the results of these analyses, particularly in light of the limitations. There were two limitations associated with gene by environment analysis of complex behavioral outcomes. First, the gene by environment analysis of complex behaviors was based on probabilistic methods and was not able to identify specific functional implications associated with the polymorphisms (Meaney, 2010). As a result, this type of study was unable to determine a causal relationship between dopaminergic polymorphisms and IPV perpetration (Meaney, 2010). Second, this analysis was not able to account for epigenetic differences, which may mitigate the effects of specific polymorphisms by altering transcription (Essex et al., 2013).
There were also some limitations associated with the study design. As a secondary data analysis, the possible variables were limited to those collected by the study investigators. Several potentially important variables, including child maltreatment and exposure to parental IPV, were not available in the data. In addition, these results depended upon perpetrator self-report, which may differ from victim report (Renner, Schwab-Reese, Peek-Asa, & Ramirez, in press). In addition, these analyses were at the level of the individual, rather than at the event level. Exposure to financial stressors may change over time so an analysis at the event level would have been able to determine the specific levels of exposure at the time of perpetration. For example, a recent study asked participants to keep daily diaries of their affect and their experiences with their partners (Elkins et al., 2013). By tracking affect every day, these researchers were better able to determine the temporal association between affect and IPV. However, this type of data was expensive to collect, time-consuming for both participants and researchers, and subject to high costs. Finally, this analysis was not able to account for whether IPV perpetration occurred during the relationship or after a relationship ended. In addition, individuals who were not in a partnership during the past 12 months may have continued to perpetrate against former partners, but that perpetration was not captured in the current data structure. The termination of relationships has been identified as a high-risk period for IPV perpetration (Saftlas et al., 2010), so it may be important to understand how the relationships of stressors and genes influence IPV among former partners.
Despite these limitations, these findings may have important implications for future IPV research and practice. A better understanding of the interactions between genes and the environment may result in more effective intervention and prevention activities that can be tailored to the individual and those at highest risk for perpetration. Ultimately, pharmacological or behavioral intervention may be tailored to individuals based on their composition of genetic and environmental risk factors.
Footnotes
Acknowledgements
Special thanks to Marizen Ramirez, Sato Ashida, and Carri Casteel for their feedback on earlier drafts of the article.
Authors’ Note
Laura M. Schwab-Reese is now at Kempe Center for the Prevention and Treatment of Child Abuse and Neglect, Department of Pediatrics, School of Medicine, University of Colorado Anschutz Medical Campus, Aurora, Colorado, USA. Information on how to obtain the Add Health data files is available on the Add Health website (![]() ).
).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by a pilot project through the University of Iowa Injury Prevention Research Center. This research uses data from Add Health, a program project directed by Kathleen Mullan Harris and designed by J. Richard Udry, Peter S. Bearman, and Kathleen Mullan Harris at the University of North Carolina at Chapel Hill, and funded by Grant P01-HD31921 from the Eunice Kennedy Shriver National Institute of Child Health and Human Development, with cooperative funding from 23 other federal agencies and foundations. No direct support was received from Grant P01-HD31921 for this analysis. Special acknowledgment is due to Ronald R. Rindfuss and Barbara Entwisle for assistance in the original design by the Add Health Team.