
Abstract

Keywords
The breast is a unique organ in that it fully matures during pregnancy and lactation, when it undergoes complete remodeling of its epithelium and stroma to support the synthesis and secretion of milk and its delivery to the infant. 1 Mammary stem cells are responsible for the changes that occur in the breast during this period, which can be repeated many times during the life of a female. What is less well known is that these stem cells, together with other cell types from the breast, filter down into breast milk and are ingested by the breastfeeding child. Interestingly, stem cells in human breast milk have been recently shown to have pluripotent features. This means that they are able to differentiate into not only breast-specific cells but also many other cell types, including neural-like cells. 2 This provides new grounds to support the argument that non-nutritional breastfeeding benefits to infants may extend well beyond immunological protection, 3 with the discovery of breast milk stem cells having spurred scientists to investigate their potential functions in the development of the infant’s brain and other organs. Furthermore, the non-invasive source and easily accessible nature of these cells emphasize their potential to be used in cell replacement therapies (CRTs) in patients with damage in the central nervous system (CNS). 1 Faced now with this new discovery, the dynamic nature of breast milk stem cells may hold great promise for both the lactation field and regenerative medicine.
From the Breast to the Milk
Behind the changes in the female breast during pregnancy and lactation are mammary stem cells (MaSCs). MaSCs are capable of producing both cell types of the mammary epithelium, the myoepithelial cells and the luminal cells, which include the lactocytes. 4 In 1998, it was shown that epithelial clearing from mouse mammary fat pads and transplantation of mammary epithelial cells from another mouse resulted in reconstitution of fully formed mammary glands. 5 This suggested that only a small number of cells, which were thought to be bipotent stem cells, can create a full gland. Since then, this has been repeated with transplantation of single MaSCs, which were able to reconstitute a full functional mammary gland in mice. 6 In vitro spheroid assays confirmed the presence and assisted characterization of bipotent MaSCs. 7 Most studies, however, have been conducted in the mouse mammary gland, which is now known to contain MaSCs with the signature of CD49fhigh/CD29+/CD24low/Sca-1+.4,8 Human MaSCs have been less characterized, particularly in the fully mature organ, due to the scarcity of human lactating breast tissue.1,9 It has been proposed that MaSCs in the human resting (in non-pregnant, non-lactating females) breast have the phenotype CD49fhigh/CD29+/CD24low/EPCAMlow/- and also express cytokeratin 5.1,10 Marker profiles for more differentiated progenitors (EPCAM+/CD49f+) and myoepithelial (CK14+) and luminal mammary cells (CK18+) are also known, but the mammary cellular hierarchy and the properties of each developmental stage still remain elusive.1,10 MaSCs are scarce in the resting breast but become activated during pregnancy and lactation, fueling the remodeling of the breast during this period. Interestingly, MaSCs of the lactating breast are found in breast milk,1,2,11,12 and the term human breast milk stem cells (hBSCs) has been coined by Hassiotou et al 2 to describe these cells.
Breast Milk Stem Cells and Their Promise
Cregan et al 11 first identified cells with mammary stem/ progenitor properties in human breast milk-derived cultures by staining for MaSC markers. A subsequent study by Thomas et al 12 reported expression of the epithelial progenitor marker p63 by a subpopulation of cells in breast milk. The presence of MaSCs in breast milk was confirmed by another research group, 13 and investigations commenced to further characterize the properties and differentiation potential of hBSCs. 2
hBSCs display the 2 hallmarks of “stemness,” that is, self-renewal and the potential to differentiate into many cell types.2,14 Recently, we reported that hBSC populations express pluripotency stem cell markers (OCT4, SOX2, NANOG, SSEA4, and TRA-1-60/81). 2 Expression of these markers was found to be minimal in the resting breast epithelium but upregulated in the lactating epithelium, pinpointing to the origin of these cells and the marked changes the breast undergoes during pregnancy and lactation.2,14 When compared with human embryonic stem cells (hESCs), hBSCs displayed remarkable similarities in morphology, phenotype (gene expression) (Figure 1A), and functionality (differentiation capabilities). 2 They were also shown to be capable of differentiating both spontaneously and in a directed way into cells from all 3 germinal layers in vitro, including mammary cells producing milk proteins, osteoblast-like cells, chondrocyte-like cells, adipocyte-like cells, hepatocyte-like cells producing albumin, pancreatic beta-like cells producing insulin, cardiomyocyte-like cells, and neuron-like cells1,2 (Figure 1B). Neuron-like cells were obtained both from spontaneous differentiation of hBSCs and from their directed differentiation in specialized media, 2 commonly used for the culture of neural cells and/or the differentiation of hESCs and neural stem cells towards neural cells. The resulting hBSC-derived cells had neuronal morphology and expressed the markers β-III tubulin and nestin, which are expressed in neurons. 2 In addition, more recent research is showing that neural-like cells deriving from hBSCs also express the neuron marker MAP2 and are not restricted to neurons only but also extend to glia-like cells expressing the glial marker s100b. 15
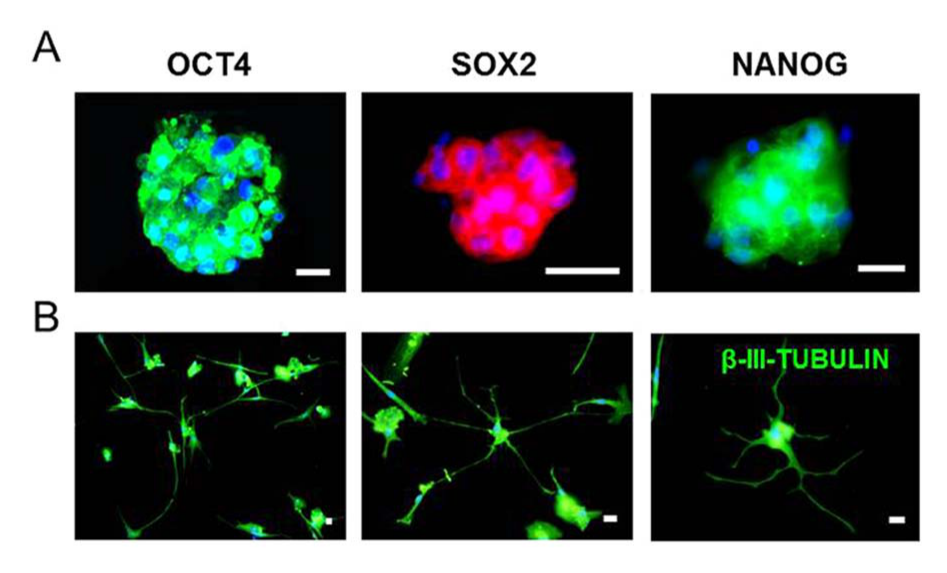
A. Human Breast Milk Stem Cells Cultured as Spheroids. B. Neuron-like Cells Differentiated from Human Breast Milk Stem Cells.
These findings have generated numerous implications, including the physiological role of hBSCs for the breastfed infant. Animal studies, including primate models, have previously demonstrated that live milk-ingested leukocytes can pass through the intestinal layer of the neonate into the systemic circulation and from there be transported and integrate into different organs.16-18 Is it possible that stem cells from human breast milk do the same in human babies? Live breast milk stem cells may integrate and differentiate into neural cells as well as other cell types in the infant’s body, potentially secreting neurotrophic factors and contributing to tissue homeostasis, development, and/or regeneration in the infant.1,19 Attempts are currently being made to further investigate the differentiation potential of hBSCs towards the neural and other lineages both in vitro and in vivo, in an effort to answer some of these questions and with the hope that they may be used in regenerative medicine.
Of great interest, hBSCs may be of use in CNS-related CRTs. Damage to the CNS, involving the brain and spinal cord, either through injury or disease, often leads to devastating and irreversible disabilities due to loss of functional neural cells and their slow regeneration.20,21 Therapeutic interventions for these conditions include CRTs, which involve injecting either undifferentiated stem cells or partially differentiated progenitor cells into the areas of damage along the CNS. 22 A number of stem cell candidates have been investigated for use as CRTs, including human embryonic stem cells (hESCs), induced pluripotent stem cells (iPSCs), and human mesenchymal stem/progenitor cells (hMSCs). However, all these stem cell types are associated with a number of transplantation issues, including formation of multilineage tumors 23 and risk of disease transmission to the donor, 24 which is why they have not yet been implemented into routine therapies. Accessing hESCs raises serious ethical issues because of their derivation from early embryos. 25 Thus, the limitations associated with their use in CRTs call for the need to explore better cell candidates for in vivo human therapies.24,26,27
Breast milk has been recently identified as a novel source of stem cells for CRTs, with potential application in the CNS due to their neural differentiation capabilities. 2 Perhaps the common origin of the CNS and the mammary gland from the ectodermal germinal layer is associated with the potential of hBSCs to differentiate towards the neural lineage. Importantly, similar to the pluripotent very small embryonic-like (VSEL) cells and the multilineage-differentiating stress-enduring (MUSE) cells from the bone marrow,28,29 tumors are not formed by hBSCs in the teratoma assay, 2 suggesting their safety for CRTs. Further tests are needed to demonstrate integration of hBSCs in vivo in adult organs and contribution to tissue repair and regeneration, similar to what has been shown for VSEL and MUSE cells.30,31
Future Prospects
From the 2007 discovery of stem-like cells in breast milk, we now know that hBSCs exist and their properties extend far beyond what was originally thought. Importantly, hBSCs are not associated with the issues prohibiting routine use of other stem cell candidates in CRTs. Instead, hBSCs offer many attractive features: they are readily available, can be accessed non-invasively, and are found in all lactating women. Research thus far suggests that hBSCs can differentiate into neural-like cells, including both neuron-like and glia-like cells, which is the first step for discerning whether these stem cells are suitable for CRTs in CNS-damaged patients and have a function in infant brain development. hBSCs may survive the neonate gut and be transported via the systemic circulation to distant sites. It is exciting to speculate that there they integrate into the infant’s tissues, including the brain, suggesting a role for hBSCs in supporting tissue homeostasis, regeneration, and development early in life. These hypotheses certainly merit further investigation, especially because they may provide a mechanistic basis for the faster brain development and higher IQ of breastfed infants.
Footnotes
Acknowledgements
Many thanks are extended to Dr Anne Merewood and Dr Donna Geddes for editorial comments on an earlier version of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Foteini Hassiotou and Peter E. Hartmann were supported by an unrestricted research grant from Medela AG (Switzerland) and a Women and Infants Research Foundation grant to Foteini Hassiotou. Neither of these organizations had any input on the article.