
Abstract
Background:
Human milk contains messenger ribonucleic acid (mRNA), a key player in protein production and a source of gene expression information for understanding lactation physiology. The mRNA is renowned for its fragility and exists in several milk fractions, creating practical challenges for mRNA isolation and analysis. While research teams have developed protocols for the use of human milk samples in transcriptomic applications, it is unclear whether there are best practices in sample collection, storage, and processing procedures for the collection of the highest quality mRNA.
Research Aim:
We aim to review current practices in the collection, storage, and processing of human milk relevant for its use in transcriptomic applications (using mRNA) in the last 10 years.
Method:
The PubMed database was systematically searched for publications addressing methodological considerations or the use of mRNA derived from human milk samples.
Results:
Most sources described the collection of human milk from small cohorts of term mothers in a mature lactational stage. While fresh collection of human milk was common, some was frozen or preserved. Several centrifugation protocols exist, and considerations for collection, storage, and processing may vary by transcriptomic application. There is inconsistency in the description and reporting of mRNA quality used in analyses that obfuscates the determination of best practices for best mRNA quality.
Conclusions:
Research into the effects of human milk collection, processing, and storage on mRNA quality metrics is warranted.
Keywords
Background
Human milk feeding is recommended by the American Academy of Pediatrics and the World Health Organization as the ideal primary source of nutrition for infants exclusively until at least 6 months of age (Meek & Noble, 2022). Continued human milk feeding is recommended for 2 years or beyond and is strongly associated with numerous beneficial maternal and infant health outcomes (Meek & Noble, 2022). Human milk is well recognized as a complex, nutritive substance that contains a variety of components including carbohydrates, protein, fat, vitamins, minerals, enzymes, and hormones. Additionally, it may be considered a living tissue that is rich in mammary and immune cells and contains a multitude of bioactive molecules critical to developmental programming (Hassiotou, Geddes, et al., 2013; Tingö et al., 2021). Decades of intensive study have revealed that human milk contains biological markers that provide information about the health of mothers and infants, as well as keys to understanding processes by which human milk feeding is able to support health (Dastmalchi et al., 2022; Hassiotou, Geddes, et al., 2013). The transcriptome, or entire collection of gene reads present in a cell or source, has been recognized as a valuable resource in the field of lactation physiology (The National Institutes of Health, 2020). During the last 2 decades, study and sequencing of expressed transcripts within human milk, human milk transcriptome sequencing, has grown in popularity for its ability to provide insights into gene expression and inferred function (Lemay, Ballard, et al., 2013; Maningat et al., 2009; Twigger et al., 2015).
Research seeking to understand milk cellular function may use a methodology designed to sequence ribonucleic acid (RNA), specifically messenger RNA (mRNA), that act as a blueprint for cell protein production (Lemay, Ballard, et al., 2013; Schroeder et al., 2006). These mRNA are often referred to as coding RNAs as many mRNA code for proteins. Other types of non-coding RNA exist in milk that act as cellular regulators, like microRNAs (miRNA), long non-coding RNAs (lncRNA), and circular RNAs (circRNA; Tingö et al., 2021). Human milk may contain RNA in several places, including the cellular components of the milk, skim (or whey), lipid (or fat/milk fat globules) and extracellular vesicle components (including exosomal), and therefore research employing human milk transcriptome sequencing methods may vary (Alsaweed et al., 2015; Maningat et al., 2009; Raymond et al., 2022).
Practical considerations may limit the feasibility of human milk transcriptomic research in population-based cohorts. Unlike deoxyribonucleic acid (DNA), RNA is renowned as fragile, and its components are susceptible to degrading enzymes, like RNases (Schroeder et al., 2006). The gold standard of RNA quality assessment is calculation of the RNA integrity number (RIN), which is performed using an electrophoresis tool like the Agilent Bioanalyzer and results in a numerical value from 1 (degraded) to 10 (intact; Schroeder et al., 2006). Previous research suggests that RNA in fresh (unfrozen) human milk degrades markedly as the distance from the collection site to the laboratory increases, with sites up to 25 miles away often providing RINs considered unsuitable for whole transcriptome sequencing (Lemay, Hovey, et al., 2013). This suggests that research teams conducting studies in animals or in laboratory-adjacent settings may be able to process milk samples individually or quickly, while collection from early postpartum (outpatient, potentially location-bound) participants could present challenges in sample batching and transport. For many tissue types, snap-freezing to -80 °C may be employed to protect against cellular degradation during transport to the laboratory or long-term storage. While the snap-freezing of human milk at the time of collection has been performed in previously conducted studies using human milk transcriptome sequencing, it is currently unclear whether snap-freezing effectively stabilizes the sample or compromises its integrity (Barboza et al., 2012).
Heterogeneity in approaches to human milk transcriptome sequencing pose significant challenges to the field of lactation science. The presence and use of a multitude of methodologies related to the collection, storage, and processing of samples may obfuscate the appraisal and consumption of published research. Furthermore, it presents a barrier to scientific teams attempting to plan and reliably execute their own human milk studies. Additional challenges likely include variable description or disclosure of study protocols and procedures. Here, we sought to review current practices in the collection, storage, and processing of human milk samples relevant to transcriptome sequencing of coding RNA (mRNA) in the last 10 years.
Methods
Design
This research is a state of the science review with a focus on procedures used in the collection, storage, and processing of human milk samples for the study of gene expression or transcriptomic applications. We present a synthesis of the most recent evidence guiding current procedures. A formal review protocol is available in the online Supplementary Materials (S1). No IRB review was necessary for this review of the literature.
Sample: Defining the Articles Reviewed
Inclusion criteria specified that sources must either evaluate sample collection, storage, or processing methodology relevant to (mRNA) human milk transcriptome sequencing or use the methodology in pursuit of a primary study aim. Due to an overwhelming surplus of sources to describe human milk RNA sequencing (including rapidly growing interest in miRNA), this review focuses specifically on the sequencing of coding RNA (mRNA). For non-coding RNA (miRNA, lncRNA, etc.), a high-quality systematic review exists to meet this need (Tingö et al., 2021). As an additional inclusionary note, although the primary focus of this review was transcriptomic (global gene expression), we included sources using RT-qPCR techniques (targeted expression) for their potential to reveal critical insights into how collection, processing, or storage of samples may affect mRNA quantity or quality in human milk. Non-primary sources (i.e., reviews), sources without peer-review, those examining primarily viral RNA (non-host) or non-coding RNA, those examining RNA from exosomes (often serving as packages for miRNA), and animal studies were excluded from this synthesis. Sources that included both animal and human data were included if all other inclusion criteria were met. Lastly, those published in a language other than English were not reviewed by our English-speaking study team. Sources were first screened against inclusion and exclusion criteria independently by two reviewers. The final sample size for this state of the science review includes 24 sources (Figure 1; Page et al., 2021).
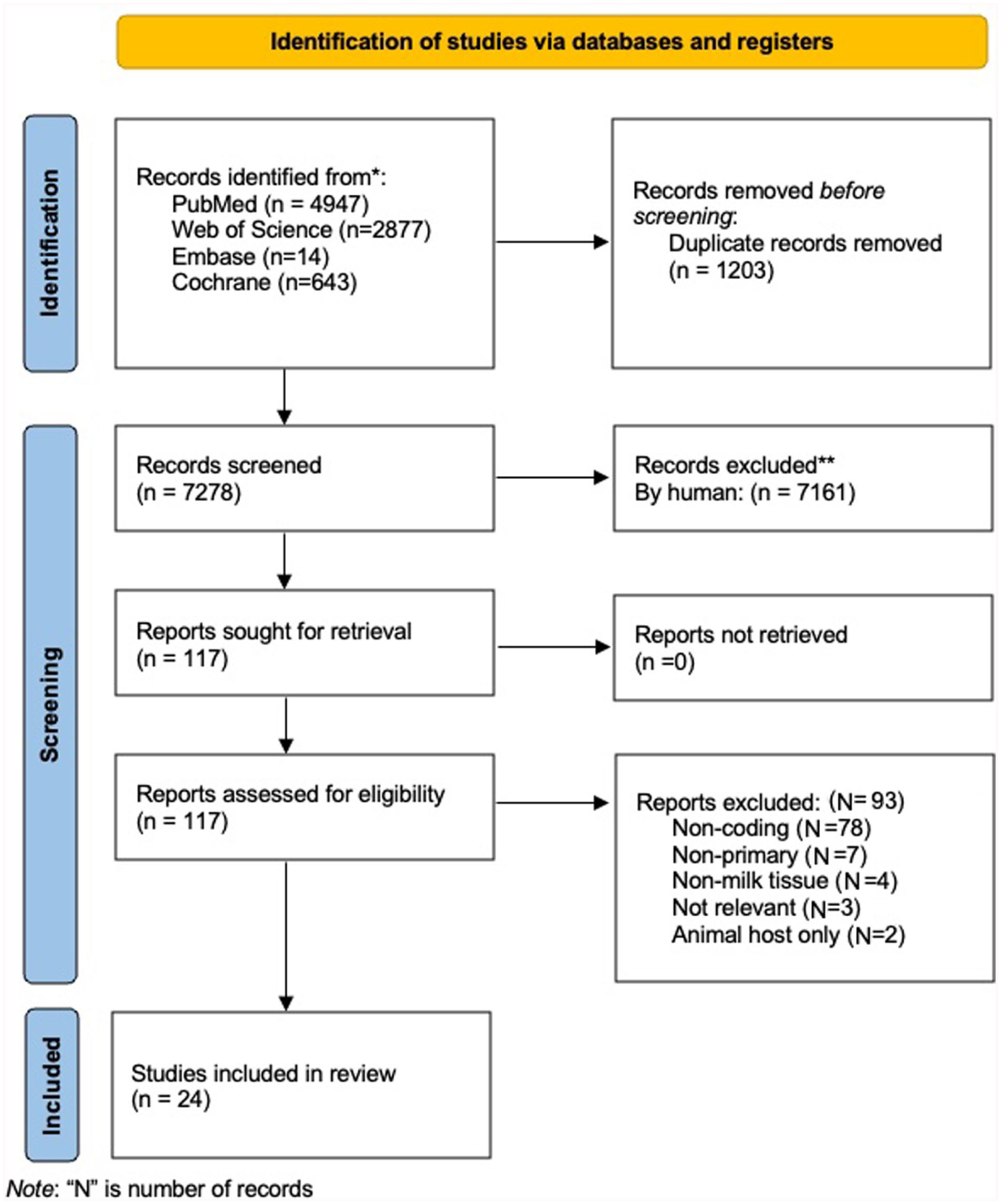
Search and Screen for Included Sources.
Data Collection: The Search Strategy and Process
The authors initially met with a senior academic librarian at the University of Wisconsin – Madison to collaboratively develop a rigorous search strategy. Articles published through September 12, 2024 were considered for review, with date and English filters applied to return sources published during the last 10 years (2013–2023, with extension to 2024). Primary sources either addressing sample collection, processing, or storage for (mRNA) human milk transcriptome sequencing, or those using the methodology in pursuit of a primary research aim, were identified in the PubMed, Web of Science, Embase, and Cochrane databases, and considered potentially relevant. The full search strategies are available in the online Supplementary Materials (S2). Following the search, all results were uploaded to the Covidence platform for two-author review (Veritas Health Innovation, n.d.).
Measurement
We identified and collected relevant variables from the sources that include (1) source demographic information, (2) first author and year of publication, (3) the number of included participants and whether they were term or preterm populations, (4) the date of antenatal or postpartum sample collection, (5) the experimental method (i.e., qPCR, RNA-seq), (6) sample storage conditions provided, (7) which milk fraction was used (i.e. cells, milk fat globule), (8) time of day the sample was collected, (9) the time, temperature and speed (x g or rpm) of initial centrifugation, (10) the quantity of human milk collected, (11) which isolation kit (if any) was used, and (12) descriptions of RNA concentration, purity, or integrity assessment. Two authors participated in data extraction from sources that met full-text inclusion criteria using a piloted form and table of variables.
Data Analysis
Data was synthesized quantitatively through categorization and comparison for a total of 24 sources (N = 24) to provide a summary and analysis of source demographics and variables. Summaries of extracted data are available, with demographics listed in Table 1 and measured variables in Table 2. Further synthesis was conducted within sub-groups according to experimental method (i.e., qPCR, RNA-seq, scRNA-seq) when relevant.
Demographic Information for Included Sources.
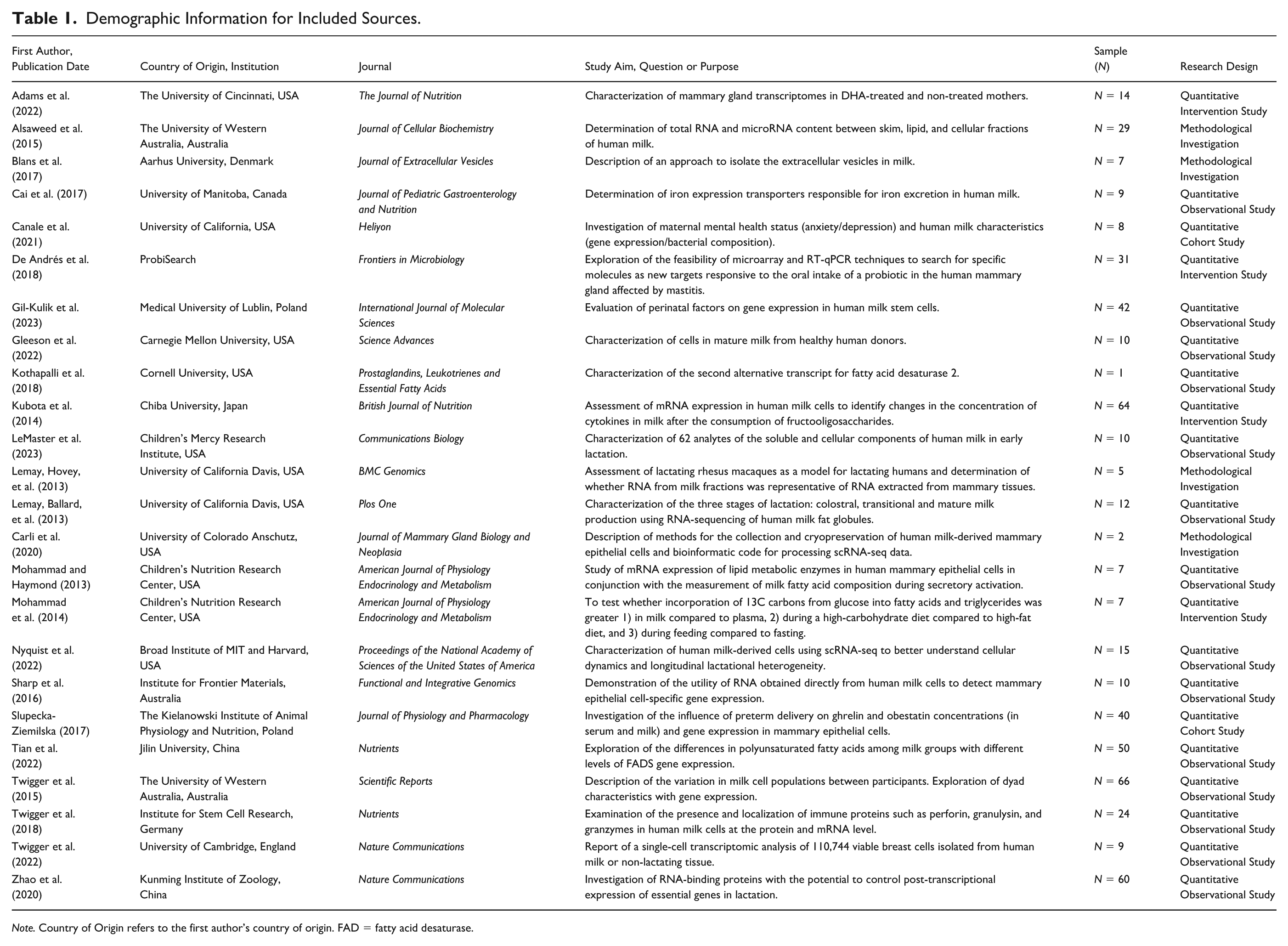
Note. Country of Origin refers to the first author’s country of origin. FAD = fatty acid desaturase.
Description of Included Sources.
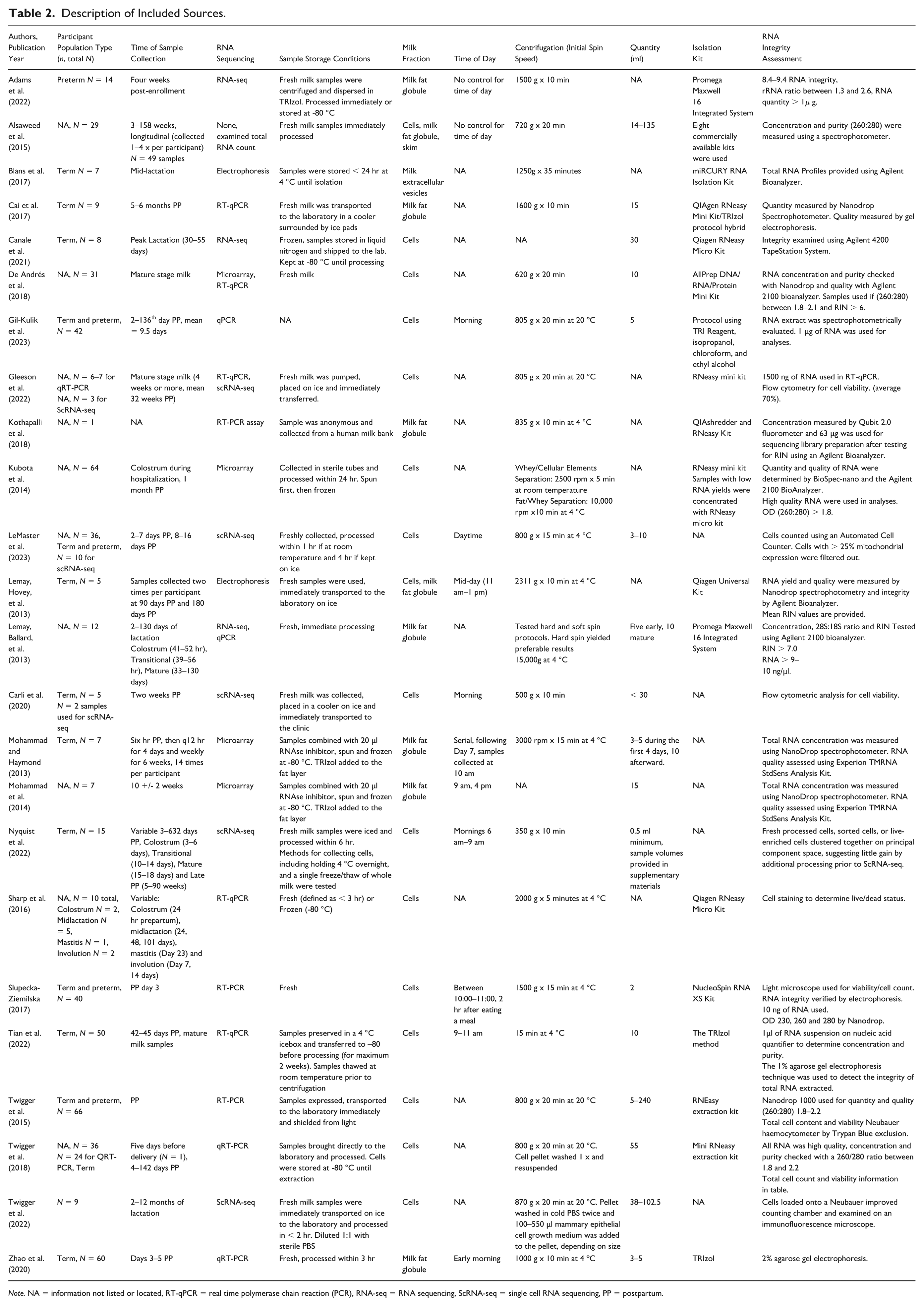
Note. NA = information not listed or located, RT-qPCR = real time polymerase chain reaction (PCR), RNA-seq = RNA sequencing, ScRNA-seq = single cell RNA sequencing, PP = postpartum.
Results
Study Characteristics
Through our literature search, we identified a total of 24 journal articles meeting our inclusion criteria. Publications examining human milk mRNA or pursuing transcriptome sequencing remained relatively consistent throughout this period, with 11 studies published before 2018 and 13 published in 2018 or later (see Table 2). We found that the earliest published studies (prior to 2018) used techniques including basic RNA quantification (N = 1; Alsaweed et al., 2015) and electrophoresis (N = 2; Blans et al., 2017; Lemay, Hovey, et al., 2013), microarrays (N = 3; Kubota et al., 2014; Mohammad & Haymond, 2013; Mohammad et al., 2014), qPCR (N = 5; Cai et al., 2017; Lemay, Ballard, et al., 2013; Sharp et al., 2016; Slupecka-Ziemilska, 2017; Twigger et al., 2015), and RNA-seq (N = 1; Lemay, Ballard, et al., 2013). While qPCR (N = 7; De Andrés et al., 2018; Gil-Kulik et al., 2023; Gleeson et al., 2022; Kothapalli et al., 2018; Tian et al., 2022; Twigger et al., 2018; Zhao et al., 2020) and RNA-seq (N = 2; Adams et al., 2022; Canale et al., 2021) remain commonly used in later published work (2018 and later), recent years have brought a rise to the use of single-cell RNA sequencing technologies (N = 4; Carli et al., 2020; Gleeson et al., 2022; LeMaster et al., 2023; Nyquist et al., 2022; Twigger et al., 2022).
Human milk samples in the included studies appeared to be mainly sourced from small groups of postpartum mothers delivering term infants. Most studies (N = 10; Blans et al., 2017; Cai et al., 2017; Canale et al., 2021; Carli et al., 2020; Lemay, Hovey, et al., 2013; Mohammad & Haymond, 2013; Nyquist et al., 2022; Tian et al., 2022; Twigger et al., 2018; Zhao et al., 2020) sampled only mothers of term infants, although some included preterm infants (N = 1; Adams et al., 2022), combined populations of term/preterm (N = 4; Gil-Kulik et al., 2023; LeMaster et al., 2023; Slupecka-Ziemilska, 2017; Twigger et al., 2015), or did not directly specify. Most milk samples were provided by participants from the mature lactational stage (
Sample Properties
We found that during the last 10 years, investigators pursued research in the field of human milk mRNA transcriptomics using cells (N = 16; Alsaweed et al., 2015; Canale et al., 2021; Carli et al., 2020; De Andrés et al., 2018; Gil-Kulik et al., 2023; Gleeson et al., 2022; Kubota et al., 2014; LeMaster et al., 2023; Lemay, Hovey, et al., 2013; Nyquist et al., 2022; Sharp et al., 2016; Slupecka-Ziemilska, 2017; Tian et al., 2022; Twigger et al., 2015, 2018, 2022), the milk fat globule (N = 9) (Adams et al., 2022; Alsaweed et al., 2015; Cai et al., 2017; Kothapalli et al., 2018; Lemay, Ballard, et al., 2013; Lemay, Hovey, et al., 2013; Mohammad et al., 2014; Mohammad & Haymond, 2013; Zhao et al., 2020), skim (N = 1; Alsaweed et al., 2015) and milk extracellular vesicles (MEV; N = 1) (Blans et al., 2017). Research teams used a variety of fresh (N = 16; Adams et al., 2022; Alsaweed et al., 2015; Cai et al., 2017; Carli et al., 2020; De Andrés et al., 2018; Gleeson et al., 2022; Kubota et al., 2014; LeMaster et al., 2023; Lemay, Ballard, et al., 2013; Lemay, Hovey, et al., 2013; Sharp et al., 2016; Slupecka-Ziemilska, 2017; Twigger et al., 2015, 2018, 2022; Zhao et al., 2020), preserved (N = 5; Blans et al., 2017; Canale et al., 2021; Mohammad & Haymond, 2013; Mohammad et al., 2014; Nyquist et al., 2022; Tian et al., 2022), or combination (N = 1; Nyquist et al., 2022) sample preparations. Notably, one study sourced human milk from a human milk bank (N = 1; Kothapalli et al., 2018). Many did not specify expression method or the time of day that the sample was collected, but in those that did, morning collection was common (morning N = 5; Carli et al., 2020; Gil-Kulik et al., 2023; Nyquist et al., 2022; Slupecka-Ziemilska, 2017; Tian et al., 2022). Others practiced either mid-day (N = 1; Lemay, Hovey, et al., 2013) or serial collections (N = 2; Mohammad & Haymond, 2013; Mohammad et al., 2014). The sample volume collected ranged drastically from 0.5 ml to 240 ml (Nyquist et al., 2022; Twigger et al., 2015). While many studies collected sample volumes of 1 fluid ounce (30 ml) or less (N = 13; Cai et al., 2017; Canale et al., 2021; Carli et al., 2020; De Andrés et al., 2018; Gil-Kulik et al., 2023; LeMaster et al., 2023; Lemay, Ballard, et al., 2013; Mohammad & Haymond, 2013; Mohammad et al., 2014; Nyquist et al., 2022; Slupecka-Ziemilska, 2017; Tian et al., 2022; Zhao et al., 2020), several studies did not disclose the sample volumes of human milk used in analyses or sampled across a considerable range (e.g., 6–240 ml). Notably, 10–15ml of mature milk was common among those studies that isolated milk fat globules (Cai et al., 2017; Lemay, Ballard, et al., 2013; Mohammad & Haymond, 2013; Mohammad et al., 2014) while volumes for cellular isolation varied dramatically (0.5–240 ml). In those collecting colostrum, small volumes were ubiquitous (0.5 ml–5 ml; Gil-Kulik et al., 2023; Nyquist et al., 2022; Slupecka-Ziemilska, 2017).
Sample Processing and Storage
Human milk samples were routinely centrifuged for separation of milk components (cells, skim, fat) prior to RNA isolation and sequencing. A variety of sample centrifugation protocols were used, and the results suggest that the milk component targeted for isolation (i.e., cells, milk fat globule, extracellular vesicle) and planned experimental method (i.e., scRNA-seq) were important considerations in protocol selection. For cellular components, 620 g–15,000 g for 5–20 min (Alsaweed et al., 2015; De Andrés et al., 2018; Gil-Kulik et al., 2023; Gleeson et al., 2022; Lemay, Hovey, et al., 2013; Sharp et al., 2016; Slupecka-Ziemilska, 2017; Tian et al., 2022; Twigger et al., 2015, 2018) or 2500 rpm for 5-min (Kubota et al., 2014) protocols were used. Temperature settings varied (20 °C N = 3, 4 °C N = 4). Notably, those pursuing single-cell RNA sequencing used centrifugation protocols with lower force, 350–870 g for 10–20 min at 4 °C or 20 °C (Carli et al., 2020; Gleeson et al., 2022; LeMaster et al., 2023; Nyquist et al., 2022; Twigger et al., 2022). In studies isolating the milk fat globule, centrifugation speeds ranged from 720–2311 g for 10–20 min (Adams et al., 2022; Alsaweed et al., 2015; Cai et al., 2017; Kothapalli et al., 2018; Lemay, Ballard, et al., 2013; Lemay, Hovey, et al., 2013; Zhao et al., 2020) or 3,000 rpm for 10 min (Mohammad & Haymond, 2013; Mohammad et al., 2014) at 4 °C. The singular study examining the milk extracellular vesicles listed a centrifugation protocol of 1250 g x 35 min (Blans et al., 2017).
Later processing steps included RNA isolation, RNA quantification, and integrity testing. Results indicated that a variety of commercially available RNA isolation kits were used, but that Qiagen kits were most common (N = 9; Alsaweed et al., 2015; Cai et al., 2017; Canale et al., 2021; De Andrés et al., 2018; Kubota et al., 2014; Lemay, Hovey, et al., 2013; Sharp et al., 2016; Twigger et al., 2015, 2018). Of those examining the milk fat globule, approximately half (N = 3) opted for an extraction protocol including TRIzol or a similar reagent (Cai et al., 2017; De Andrés et al., 2018; Zhao et al., 2020). Few studies explicitly reported RNA integrity numbers (N = 4; Adams et al., 2022; De Andrés et al., 2018; Lemay, Ballard, et al., 2013; Lemay, Hovey, et al., 2013), quantities (N = 7; Adams et al., 2022; Gil-Kulik et al., 2023; Gleeson et al., 2022; Kothapalli et al., 2018; Lemay, Ballard, et al., 2013; Slupecka-Ziemilska, 2017; Tian et al., 2022), or purity (N = 5; Alsaweed et al., 2015; De Andrés et al., 2018; Kubota et al., 2014; Twigger et al., 2015, 2018). Ultimately, the total number of studies that did not provide descriptive or quantitative measures of RNA quantity or integrity beyond mention of method (Nanodrop, Agilent Bioanalyzer) used was high. For sources using scRNA-seq, several provided quality control metrics to describe cellular count and viability (Carli et al., 2020; Gleeson et al., 2022; LeMaster et al., 2023; Nyquist et al., 2022; Twigger et al., 2022).
Discussion
Following review and synthesis of 24 published sources describing recent methodological inquiries and applications of human milk mRNA-based experimental technique, we report significant heterogeneity in sample collection, processing, storage, and reporting. It is unclear whether the application of diverse study protocols reliably produces human milk mRNA suitable for sequencing. Based on our synthesis, we will discuss relevant considerations and present several opportunities for further development of rigorous methods in human milk mRNA handling, processing, and storage.
Sample Collection
Human milk is well known to be a dynamic tissue, and therefore samples collected from small cohorts, across lactational stages, and by differing expression methods, may result in heterogeneity. Collection of mature milk samples from small cohorts were the most common among the studies included in this review. While the high cost of RNA-sequencing services has depreciated considerably in recent years, (Hu et al., 2021) it is possible that project budget or feasibility constraints limited cohort sizes.
We can also speculate that parent availability and mobility, expressed volume, and RNA quality may favor inclusion of mature lactating populations of term-born infants in studies to reduce the challenges inherent in the collection of samples in populations producing colostrum and transitional milk. As with RNA from other tissue types, RNA in human milk samples decreases over time (Lemay, Hovey, et al., 2013). Colostrum collected antenatally and immediately postpartum may be feasible if lactating populations are inpatient or accessible in ways that facilitate transport of samples to a laboratory for processing. Notably, in healthcare systems designed to discharge maternal populations shortly after birth, samples collected after 2–4 days of life may be difficult to acquire as maternal populations move into residences that may not facilitate sample collection and transportation (i.e., longer transport times) necessary to preserve human milk RNA. Notably, the lactational stage itself may affect the quality of sample RNA. Data on RNA integrity suggest that RNA quality increases with progressive lactational stage, with 27% of colostrum samples, 60% of transitional samples and 78% of mature milk samples providing RNA suitable for RNA-seq (RIN
Whether the milk expression technique affects the integrity of human milk samples for transcriptome sequencing remains unclear. We found that descriptions of collection methods (i.e., hand expression, hand/mechanical breast pump), interval since last expression, and time of sample collection were often vaguely described or omitted. If research teams seek to isolate RNA from cellular or milk fat globule sources, time of day and expression practice may be relevant. Human milk cellular content has been observed to fluctuate with circadian rhythm and may be higher in hindmilk than foremilk (Alhussien & Dang, 2018; Hassiotou, Hepworth, Williams, et al., 2013). Nursing frequency may also affect a cellular sample with higher numbers of apoptotic cells potentially accumulating with longer between-expression intervals or individuals with higher breast capacities (Martin Carli et al., 2021). Because mRNA is quickly degraded, apoptotic cells may lack high-quality mRNA. Similarly, it is reasonable to speculate that RNA in the milk fat globule may be most accessible in milk with higher fat fractions, such as hindmilk (Hassiotou, Hepworth, Williams, et al., 2013). Whether inter-breast variation is a factor is also unclear, and therefore research teams may consider adherence to best practice protocols for general human milk collection whenever possible (McGuire et al., 2021).
Sample Quantity
Sample quantity is a critical consideration in the planning of human milk gene expression or transcriptomic studies. Research teams must collect the smallest amount of human milk necessary to ensure minimal burden to research participants while still collecting a sample that will reliably yield RNA appropriate for analyses. Although many studies did not describe the quantity of human milk collected, we found that those studying colostrum collected smaller amounts (maximum 5ml) than those working with transitional or mature milk (commonly
Fresh Collection or Preservation
The results of our synthesis suggest that freshly collected or frozen human milk may be appropriately used if correctly handled for specific experimental applications. Stabilization techniques may offer a lucrative advantage, as they may preserve or prevent RNA from RNase degradation and broach significant feasibility concerns in the sampling of population-based or disease phenotype cohorts of childbearing people. Although most studies used fresh human milk samples (Adams et al., 2022; Alsaweed et al., 2015; Cai et al., 2017; Carli et al., 2020; De Andrés et al., 2018; Gleeson et al., 2022; Kubota et al., 2014; LeMaster et al., 2023; Lemay, Ballard, et al., 2013; Lemay, Hovey, et al., 2013; Sharp et al., 2016; Slupecka-Ziemilska, 2017; Twigger et al., 2015, 2018, 2022; Zhao et al., 2020), several that were subjected to preservation methods such as freezing (Blans et al., 2017; Canale et al., 2021; Nyquist et al., 2022; Tian et al., 2022), or the addition of a stabilizing additive were used in analyses. Our ability to determine the effect of these preservation methods on RNA quantity or quality is severely limited in cases where RIN or other quality assurance metrics were not reported. Notably, the appropriateness of sample preservation and processing technique may vary based on experimental application and whether viability or lysis is a particular concern (as with ScRNA-seq). While freshly collected samples for ScRNA-seq have been shown to yield reliable high-quality data, holding samples at 4 °C overnight or freezing following cell isolation using a cell cryopreservation protocol have demonstrated success (Carli et al., 2020; Nyquist et al., 2022; Twigger et al., 2022). Samples frozen prior to cell isolation may result in poorer quality cells (Nyquist et al., 2022). Similarly, concern surrounding RNA integrity may vary based on whether a qPCR or RNA-seq application is pursued. While RIN values > 7 are strongly preferred for RNA-seq (Puchta et al., 2020), samples with quality scores > 5 may be acceptable for qPCR (Fleige & Pfaffl, 2006). These less stringent integrity requirements may explain the infrequent reporting of RNA integrity in studies using qPCR. Further research is needed to understand the effects of preservation techniques on human milk RNA of the various milk fractions (cellular, skim, lipid) and whether cellular lysis and loss of mRNA occur. It is possible that rapid freeze protocols or use of a stabilizing agent is more feasible when RNA integrity is not a paramount concern, or for use in select experimental applications. Scientists within the human milk and/or transcriptomics field may consider development and validation of remote collection procedures to support inclusion of difficult-to-access (e.g., rural, low-incidence/special) populations.
A variety of extraction and centrifugation protocols exist to guide the fractionation of human milk samples prior to RNA extraction. In general, the softest spins of 350–870 g were used for ScRNA-seq applications, likely to protect the viability and prevent lysis of cells. In the isolation of milk fat globules, research teams used slightly higher spins of 720–2311 g in contrast to the harder spins of cellular isolation at 620–15,000 g. Notably, one team found that a harder spin protocol (15,000 g) produced the highest quality cellular RNA (Lemay, Ballard, et al., 2013). Temperature settings during centrifugation varied between room temperature (20 °C) and refrigerator temperature (4 °C). Considering that many fresh samples were stored on ice for transport to the laboratory, understanding the effects of centrifuge temperature on RNA quality may be warranted, particularly for protocols involving longer spin cycles (20 min). Investigators may consider approaching RNA extraction using a column-based phenol-free method, as it demonstrates high performance for all three fractions of human milk (Alsaweed et al., 2015).
Limitations
We respectfully acknowledge that this synthesis of recent work has limitations. Results suggest that reporting of RIN, RNA quantity, and purity is not a standard practice for all experimental applications. Without this data, our ability to provide insight into the success of human milk sample collection, storage, and processing procedures for gene expression quantification or transcriptomic applications was limited. Furthermore, despite our best efforts to extract this information, it is possible that RNA quality metrics were reported elsewhere, in previous publications or locations that evaded our extraction procedures. Additional considerations outside the scope of this review, including downstream bioinformatic analysis, should be synthesized and critically evaluated in consideration of the fact that data cleaning and handling are widely recognized for the capacity to affect results. Finally, it is possible that applicable publications were published after our search or not retrieved by our search procedures.
Conclusions
The field of human milk science is in critical need of rigorously conducted studies that clearly describe the effects of sample collection, processing, and storage on human milk mRNA isolated from the cellular, skim, and milk fat globule fractions. Without this knowledge, it is unclear whether prospective studies designed to examine mammary gene expression can expect RNA with quality suitable to support various transcriptomic applications. Future studies may seek to investigate novel methods—including remote human milk collection, freezing, and transport of the sample—that facilitate large, multi-center trials. Research teams previously engaged in human milk transcriptomic work may also consider publishing study protocols with inclusion of data that describes RNA sample quantity, quality, and purity. Considering the heterogeneity in reporting practices, we also suggest that a human milk reporting checklist may be beneficial to ensure that future studies can provide the most complete and accurate descriptions of their work.
Supplemental Material
sj-docx-1-jhl-10.1177_08903344251387654 – Supplemental material for The Collection, Storage, and Processing of Human Milk Samples as a Source of mRNA: A State of the Science Review
Supplemental material, sj-docx-1-jhl-10.1177_08903344251387654 for The Collection, Storage, and Processing of Human Milk Samples as a Source of mRNA: A State of the Science Review by Katelyn Desorcy-Scherer and Kerry McNamara in Journal of Human Lactation
Footnotes
Acknowledgements
The authors would like to acknowledge the expertise and guidance of Mary Hitchcock, Senior Academic Librarian at the University of Wisconsin – Madison.
Author Contributions
Katelyn Desorcy-Scherer: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing - original draft, Writing - review & editing
Kerry McNamara: Conceptualization, Formal analysis, Investigation, Methodology, Project administration, Writing - review & editing
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Disclosures and Conflicts of Interest
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The authors have no conflicts of interest to disclose. The authors worked collaboratively with K.M.D.S., Assistant Professor, as an academic mentor to K.M., DNP student
Supplemental Material
Supplementary Material may be found in the "Supplemental material" tab in the online version of this article.