
Abstract
Background
Peritoneal membrane research has provided important insights into the physiology and pathophysiology of this tissue that is of vital importance for peritoneal dialysis patients. Among the various tools and methodologies used to study the peritoneum, we have extensively used adenovirus-mediated gene transfer.
Methods
A literature review was carried out. Information from reviewed papers was combined with the authors’ experience and results.
Results
We have used first-generation adenoviruses that are simple to construct and can infect a wide range of dividing and nondividing cell types. These vectors are restricted, however, in that they provide only a short duration of transgene expression and may elicit an inflammatory response. Modifications to this technology with helper-dependent adenovirus may circumvent these problems but with increased complexity of construction. Adenovirus-mediated gene transfer has been used to evaluate the effect of several cytokines and growth factors on peritoneal membrane physiology. We have used intraperitoneal delivery of transforming growth factor-β to generate an experimental model system of resolving peritoneal fibrosis and epithelial mesenchymal transdifferentiation. We have studied the effects of the inflammatory cytokines interleukin-1β and tumor necrosis factor alpha on the peritoneum, and have shown that antiangiogenic factors such as sFLT-1 and angiostatin can reduce the damaging effects of exposure to peritoneal dialysis solutions in an animal model.
Conclusions
The use of recombinant adenoviruses to genetically modify cells and tissues is now a common laboratory research tool. This technique has provided important advances in our understanding of the peritoneal membrane.
Keywords
Adenoviral Vector Technology
The adenoviruses have been extensively studied (3-5) and the reader is encouraged to take advantage of these more detailed papers to supplement the brief description provided in this review. Adenoviruses comprise a large group of non-enveloped icosahedral DNA viruses, characterized by an outer protein shell surrounding an inner nucleoprotein core (Figure 1). The linear double-stranded DNA genome of 36 kilobases (kb) is divided into five transcription units that contain genes for the viral proteins expressed during the early and late phases of the viral life cycle (produced before and after DNA replication respectively) (Figure 1). Adenoviruses have been isolated from dogs, mice, pigs, primates, and humans, with many of the human serotypes causing respiratory, ocular, and gastrointestinal disease. There are 51 distinct serotypes divided into 6 subgroups (A – F) based on their hemagglutinating properties and DNA sequence homology.
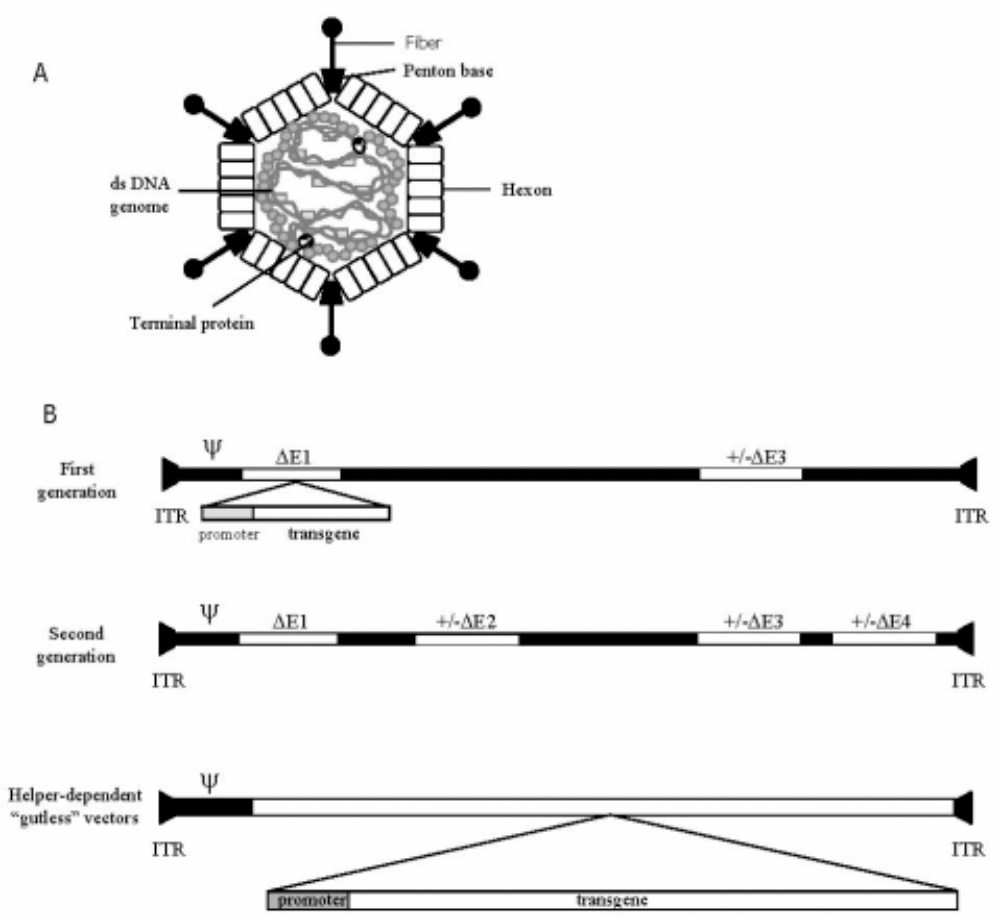
Adenovirus structure and vector constructs. Schematic diagram of an adenovirus particle showing the outer capsid proteins and the nucleoprotein core (A). The genomes of first- and second-generation vectors, and helper-dependent, or “gutless,” vectors show the viral DNA sequences (solid line), the inverted terminal repeats (ITRs), and the packaging sequence (ψ) (B). The first-generation vector shows E1 and E3 deletions (open boxes), with a transgene expression cassette inserted in E1. A second-generation E2/E4 deleted vector still retains a backbone of viral sequences. In the helper-dependent “gutless” vectors, all viral material except the ITRs and ψ is deleted, allowing for insertions of up to 38 kb of exogenous DNA containing regulatory and promoter regions and transgene coding sequences.
Adenoviruses have been used extensively in research and therapeutic applications, from investigation of general genetic organization and cellular processes of differential splicing and mRNA processing, to vaccine development and delivery. The human serotypes 2 and 5 (Ad2 and Ad5) are most commonly used for research and medical applications. Together with retrovirus and adeno-associated virus (AAV) vectors, they are the most commonly used viral vectors in gene therapy applications.
Adenoviruses exhibit a number of properties that make them attractive as gene transfer agents. Wild-type adenoviruses can be rendered replication deficient through deletion of the genetic regions responsible for virus replication. The removal of viral genes allows for the insertion of exogenous genetic material containing the coding regions and regulatory sequences of the desired protein (i.e., the transgene or gene of interest) and prevents unwanted viral replication and release of infectious viral particles. Adenoviruses can infect a number of cell types and readily infect both dividing and quiescent cells. Viral infection occurs through initial binding of the virus fiber to the Coxsackie virus B and adenovirus receptor (CAR), a cellular transmembrane receptor. Internalization of the virus particle then proceeds through the a v cell surface integrins. Upon infection, the viral genetic material does not integrate into the host genome but remains epichromosomal, decreasing the potential for insertional mutagenesis. The duration of transgene expression can be affected by the epichromosomal nature of the viral genome: as the viral genome will be diluted out and eventually lost through cell turnover, the duration of expression will be dictated in part by the rate of cell turnover. Important for research purposes, cell lines have been developed that provide the deleted viral proteins in trans, allowing for growth of recombinant replication-deficient viruses to high titers in cell culture systems. Finally, adenoviral vectors are also easy to manufacture in high concentration and purity, making them good candidates for commercial production and clinical use.
Adenoviruses exhibit a number of characteristics that make them preferable to other gene transfer agents. Adenoviral vectors have large packaging capacity compared with AAV (38 kb vs 4.7 kb). They do not integrate into the host genome, as do retroviral and AAV vectors: retroviruses insert randomly into the host genome, risking insertional mutagenesis, while AAV integrates at a specific location in chromosome 19. Adenoviral vectors have a high expression and wide tropism for resting and dividing cells compared with retroviruses, which require replicating cells for gene transfer and expression. On the other hand, adenoviral vectors — especially first- and second-generation constructs — produce only a transient gene expression in immunocompetent recipients, while retrovirus-mediated transgene expression is longer lived. Adenoviruses are highly immunogenic and in high dose can induce a fatal inflammatory response; retro-viruses and AAV are significantly less immunogenic. Non-viral transfection reagents, such as Lipofectamine (Invitrogen, Carlsbad, California, USA), have also been tested for gene transfer to the peritoneal mesothelium, but in our hands were significantly less efficient in both in vitro and in vivo gene transfer.
Adenoviral Vector Construction
Adenoviral vector technology has been continuously challenged to improve along several parameters, including vector construction and production, transgene capacity, antigenicity, and repeat delivery. The first-generation adenoviral vectors were derived by deletion of the early E1 region, which severely compromised the replicative ability of the virus [Figure 1(b)]. E1-encoded proteins are responsible for activating transcription of other adenoviral genes and also induce cell cycling through interaction with retinoblastoma protein (3). E1-deleted viruses are cultured on 293 cells, a human embryonic kidney cell line that has been transformed with the deleted E1 sequences and now provides their functions in trans. First-generation vectors may also have a deletion of the early E3 region. Proteins encoded by the E3 region subvert the host immune response through inhibition of major histocompatibility complex class I peptide loading (3). The success of these early vectors as gene transfer agents was compromised by the stimulation of a cellular immune response directed against infected cells, and a low level of replication competent viruses that arose in virus preparations due to recombination with the viral sequences in the 293 cells.
Second-generation viruses have been engineered with deletions in the early E2 and E4 regions in addition to E1 (3). These deletions effectively eliminate production of proteins essential for viral DNA replication (E2) and cell cycle control and transformation (E4), yielding viruses that are virtually incapable of DNA replication and have a larger transgene capacity. While these viruses are significantly less immunogenic, the innate immune response to the viral capsid proteins will be intact. Construction of these vectors is slightly more complicated as the packaging cell line has to be engineered to supply all of the deleted functions in trans.
Helper-dependent, or gutless vectors [see Figure 1(b)], may hold the most promise for minimizing host response and prolonging transgene expression (6). These vectors have had all the viral sequences deleted, with only the two inverted terminal repeats and the packaging signals (ψ) remaining of the wild-type genome (7). These viruses require a helper virus for replication and assembly, but purification of the helper-dependent vectors away from the helper viruses has thus far been problematic.
The short duration of transgene expression is a limiting feature of adenoviral vectors. First-generation adenoviruses tend to express for 4 – 10 days in the peritoneum (8), with transgene expression limited by the epichromosomal nature of the viral vector, the rate of mesothelial cell turnover, and the host immunogenic response directed against infected cells. Second-generation adenoviral vectors have been demonstrated to have prolonged expression, up to 70 days in one study (9), and transgene expression from helper-dependent vectors is substantially increased, lasting out to 80 days (7).
Deletion of viral sequences allows for the insertion of an expression cassette for the gene of interest. The size of the expression cassette usually corresponds to that of the deleted sequences, as the viral particle has a limited packaging capacity of up to 38 kb. The E1 deletion allows for an insertion of 5.1 kb; an E1/E3 deletion frees up approximately 8.2 kb of genetic material, while the gutless or helper-dependent vectors can accommodate an insertion of up to 38 kb, a significant increase over the first- and second-generation vectors. The expression cassette contains, at minimum, the cDNA for the gene of interest, promoter sequences, and other regulatory elements. Commonly used promoter sequences include the cytomegalovirus (CMV) promoter and promoter of the major late adenovirus transcription unit, both of which allow for constitutive expression of the transgene. Tissue-specific promoters, inducible promoters (e.g., responsive to tetracycline and rapamycin), and promoters responsive to physiological stimuli (e.g., hypoxia) can also be used for regulated transgene expression.
Cell Culture and Virus Purification
Several techniques can be employed for adenoviral vector construction and purification. We have employed a co-transfection technique exclusively (10). Using this strategy to create a first-generation adenoviral vector, a recombinant plasmid engineered with the adenovirus packaging signal, promoter, and gene of interest is cotransfected into 293 cells with a second plasmid containing the remaining adenoviral genome less the E1 and E3 regions. Homologous or site-specific (i.e., Cre-lox) recombination will lead to a viable, non-replicative adenovirus genome. The inclusion of the packaging sequence with the gene of interest increases the likelihood that the rescued adenovirus will contain the transgene.
The recombinant adenovirus is then amplified on 293 cells and purified by cesium chloride gradient centrifugation. Transgene protein expression can be assessed by Western blot analysis or ELISA of infected cell supernatant and lysate and, if possible, through a functional protein assay. The concentration of functional adenovirus in the preparation is quantified by plaque titer on 293 cells (11) and expressed as plaque-forming units (pfu), and the concentration of total adenovirus particles is assessed by optical density. While the number of infectious particles (pfu) is always less than the total number of virus particles, we usually achieve an adenovirus concentration of 0.5 – 2 × 1011 pfu/mL, with a total adenovirus particle count that is less than 50 times the active adenovirus concentration. It is advantageous to maintain this ratio between total and infectious particles, as higher total adenoviral particle counts lead to increase in the innate immune response to the adenovirus that may distort the transgene response, especially at early time points in experimental animal systems.
Experimental Parameters of Intraperitoneal Delivery
Experimental protocols for intraperitoneal gene transfer should optimize transgene expression while minimizing any side effects associated with the virus itself. Several parameters can affect the amount, distribution, and duration of transgene expression. The dose of adenovirus (e.g., the number of pfu or viral particles delivered) has a direct effect on the number of resident peritoneal cells infected and the total amount of transgene product produced (12). The volume in which the adenoviral vector is delivered dictates the distribution of transgene expression, both within the confines of the peritoneal cavity and systemically. As infection of the cells requires direct contact between the virus and the cell membrane, smaller volumes maximize uptake but minimize the total serosal surface area that will be exposed and infected. As the infused volume increases, the likelihood of lymphatic uptake and systemic expression increases. We have found that, in general, infused volumes of 100 μL or less result in a more patchy distribution of gene product, especially on the small intestine and mesentery, and on a smaller area of the ventral parietal peritoneum. In our hands, volumes in excess of 250 μL led to transgene expression in the parenchyma of both liver and spleen due to lymphatic uptake of the virus suspension. We routinely use volumes of 100 – 150 μL in our studies; however, as virus preparations can vary from batch to batch and between laboratories, we highly recommend that pilot studies to optimize viral load and dose be performed with any viral vector preparation before embarking on larger studies.
There is likely a modest dose effect on duration of transgene expression; however, this has not been carefully documented. The duration of expression is likely more a function of the immune response mounted toward the adenoviral vector, and this is a function of the number of delivered viral particles, production of viral proteins, the immunogenicity of the transgene, and the immune status of the host. It is also dependent on the nature of the transgene itself, as overexpression of proinflammatory cytokines and profibrotic and angiogenic factors can have a significant impact on peritoneal physiology, which in turn can affect viability of the transgene-expressing cells. In addition, expression of proteins from a species other than the host may elicit an immune response against the protein and the transduced cells.
Peritoneal Transgene Expression
We have carried out studies in a rat model to define delivery parameters using AdlacZ, a first-generation adenoviral vector that produces the bacterial protein β-galactosidase (β-gal). This protein remains intracellular and the enzymatic activity can be identified upon incubation with the chromogenic substance 5-bromo-4-chloro-3-indolyl β-D-galactosidase (x-gal) and appropriate stain. The staining procedure can be performed on both tissue sections and whole tissue, providing a quick and easy means of visualizing transgene expression.
In preparation for injection of the viral vector, we generally anesthetize the animals, carefully prepare and clean the anterior abdominal wall, and make an injection in the right lower quadrant using a 23-gauge needle. In a pilot study, increasing viral doses of 5 × 107 to 5 × 109 pfu AdlacZ were delivered intraperitoneally (IP) to 175 – 275 g rats in volumes of 50 – 500 μL (13). Cellular uptake of the virus and subsequent transgene expression were rapid, and all of these conditions resulted in β-gal expression on parietal and visceral surfaces within 24 hours, with coverage ranging from 40% to 90% of parietal peritoneum, spleen, liver, diaphragm, and small intestine and mesentery surface areas (Figure 2).
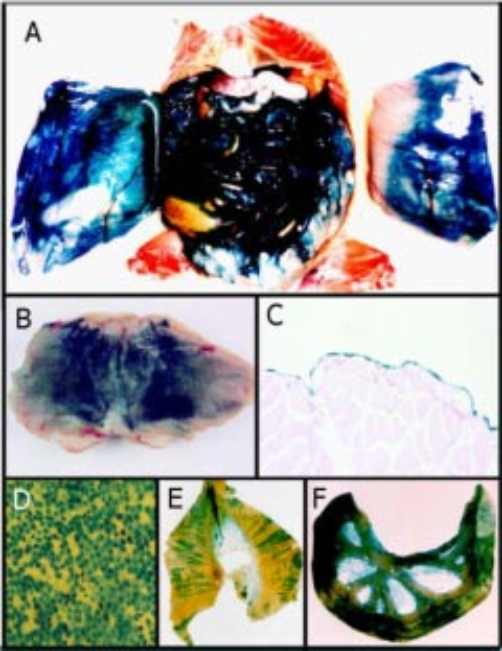
Adenovirus-mediated gene transfer to the peritoneal mesothelium. Twenty-four hours after intraperitoneal delivery of 2 × 109 plaque forming units of AdLacZ in 100 μL phosphate-buffered saline in a rat model, the peritoneal tissues were fixed and stained for β-galactosidase activity in situ. There is widespread staining of all peritoneal surfaces (A), with uniform staining on the parietal peritoneal wall (blue staining) (B). Section of the parietal wall shows that transgene expression is limited to the mesothelial monolayer. This section was counterstained with nuclear fast red. Original magnification x100 (C). β-galactosidase staining of the anterior parietal peritoneal wall (original magnification x200) (D), diaphragm (E), and small intestine and mesentery (F). Panels B and C from: Margetts PJ, Kolb M, Galt T, Hoff CM, Shockley TR, Gauldie J. Gene transfer of transforming growth factor-β1 to the rat peritoneum: effects on membrane function. J Am Soc Nephrol 2001; 12(10):2029–39. Reproduced by permission of Lippincott, Williams & Wilkins.
The primary target in the peritoneal cavity for adenovirus-mediated gene transfer is the mesothelium, the serosal lining of all parietal and visceral serosal surfaces. Mesothelial cells are relatively long-lived, with turnover rates estimated at 0.1% – 3% per day (15). Viral transduction (with AdlacZ) is limited to the mesothelial cell monolayer, with no evidence of transgene expression in the submesothelial interstitium or skeletal muscle, even in extreme cases when the overlying mesothelium has been experimentally denuded prior to virus delivery (Hoff CM, unpublished observations). In our hands, delivered volumes of up to 250 μL restricted transgene expression to peritoneal surfaces, while volumes in excess of 250 μL led to β-gal expression in the parenchyma of both liver and spleen, as well as the diaphragmatic lymphatics due to lymphatic uptake of the virus suspension. However, Setoguchi et al. reported no detectable transgene RNA in any tissues other than parietal and visceral peritoneum after delivery of 109 pfu of the adenoviral vector for α-1-antitrypsin (Ada 1AT) in 500 μL in 140 – 160 g rats (16).
As mentioned above, the volume of infusate is a determining factor in the distribution of expression. Martin–Martinez and colleagues have reported that delivery of the adenoviral vector in larger volumes [1 × 109 and 3 × 109 pfu AdβGal in 15 mL phosphate-buffered saline (PBS)] resulted in significant β-gal staining in the lymph nodes, lungs, liver, heart, and kidneys (12). This systemic expression did not appear to induce any physiological changes, as there were no associated changes in liver enzymes or renal function. Beta-galactosidase expression was also noted in the endothelial cells of the capillaries and venules, adipocytes, and mesothelial cells of the omentum, and in the mesenteric region of the visceral peritoneum 24 hours postdelivery (Figure 3). We suspect this pattern of transgene expression may be directly related to the higher volume of fluid (15 mL) used to deliver the adenovirus. This observation increases the flexibility of adenovirus as a tool to study peritoneal function. At smaller volumes, targeted tissues are limited to mesothelium; at higher volumes, targeted tissues also include important submesothelial structures.
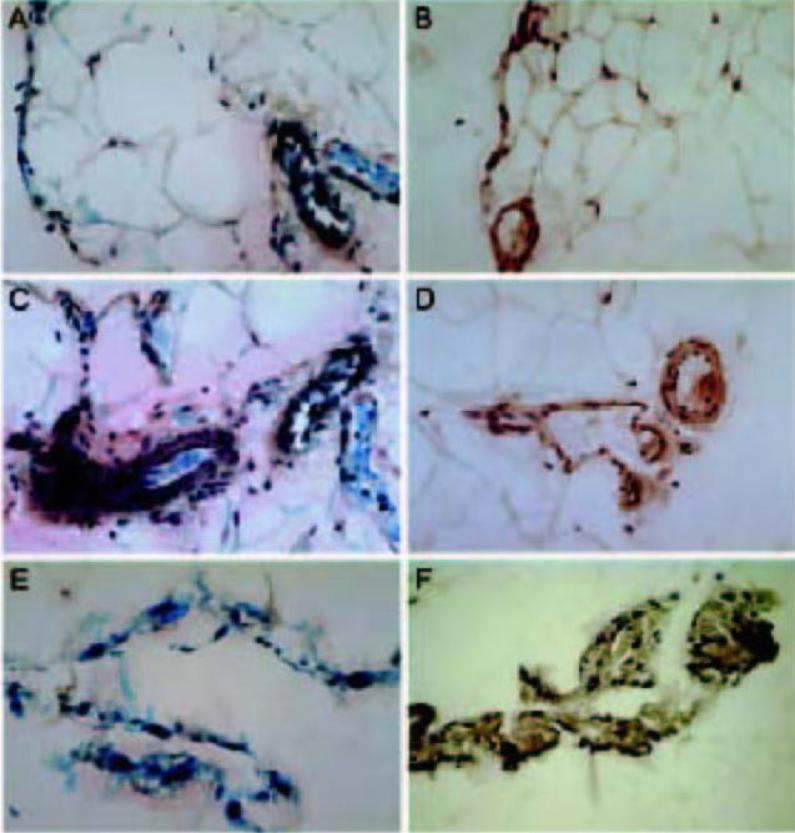
β-galactosidase (β-gal) staining in the visceral peritoneum of control and adenovirus-treated rats. Representative sections of the omentum (A–D) and visceral peritoneum (E, F) 24 hours after delivery of 3 × 109 plaque forming units of AdβGal in 15 mL buffer (A, C, E) or buffer alone (B, D, F). Specific β-gal staining (indicated by blue nuclei) is observed in the mesothelial cells (A, E), endothelial cells (A, C), and adipocytes from adenovirus-treated rats. The lack of staining indicates the lack of endogenous β-gal activity in tissues from control rats. β-gal staining in all panels. Original magnification: (A–F) x300. [From Martin-Martinez DM, Stoenoiu M, Verkaeren C, Devuyst O, Delporte C. Recombinant adenovirus administration in rat peritoneum: endothelial expression and safety concerns. Nephrol Dial Transplant 2004; 19(5):1293–7, by permission of the European Dialysis and Transplant Association – European Renal Association.]
The virus is usually administered in PBS; however, delivery in icodextrin (15% w/v, pH 7.4) has been reported to enhance gene expression in a rabbit and a mouse model due to prolonged retention and distribution of the virus solution in the peritoneal cavity (17). Based on β-gal activity, the authors demonstrated that animals treated with adenovirus in icodextrin all had >50% expression in the serosal tissue, whereas those treated with a similar adenovirus dose and volume of PBS mostly had <20% expression in the serosal tissues.
Peritoneal Response to the Virus
The adenoviral vector itself has a specific impact on peritoneal physiology, affecting the recipient animals’ well-being, peritoneal inflammation, cellular and humoral immunity, and peritoneal structure and function. The inclusion of a control, or null, virus that does not express a transgene allows differentiation between virus-specific effects and those due to specific expression of the transgene: we routinely use AdDL70, a deleted virus that contains no exogenous sequences or expression cassette (18).
We and other investigators have noticed that recipient animals commonly experience weight loss in the first few days post infection (19). In our studies, we have not seen animal mortality associated with IP administration of null adenovirus alone, and this was confirmed in a safety study using very high viral doses in rodents (20). We have observed mortality in about 15% of rats treated with an adenoviral vector expressing active transforming growth factor beta1 (TGF-β1) treated at a dose of 2 × 109 pfu in 100 μL PBS. These animals succumbed to massive fibroproliferation and bowel obstruction, not direct effects of the adenovirus per se, but most likely due to the expression of TGF-β1.
The recipient's immune response to the virus is one of the key factors in limiting the duration of transgene expression. As outlined below, the immune response is directed against the viral capsid proteins, translated viral proteins, and, in some cases, possibly the transgene itself. In immunocompetent animals, transgene expression is transient, with peak expression around 3 – 4 days. Expression then declines rapidly and is undetectable by 10 – 14 days post infection. This temporal expression has been noted by investigators using adenoviral vectors containing different transgenes and is due in large part to host response to the virus.
The acute innate immune response is likely related to the number of viral particles. It should be noted that the number of viral particles is usually 50 times the number of viable plaque-forming viruses. The ratio of particles to pfu can be variable, so for applications where a number of pfu are administered, the number of viral particles, and therefore the inflammatory response, may be different. The null adenoviral vector may also evoke an acute inflammatory response, as demonstrated by increased peritoneal cell counts (largely neutrophils) and an increase in effluent tumor necrosis factor alpha (TNF-α) that has been seen after administration of both a null virus, AdDL70, and transgene-specific vectors. The inflammatory response is accompanied by a change in mesothelial cell morphology, with the cells assuming a more rounded shape that is most prominent 2 days after infection. The inflammatory response peaks by day 3 and resolves completely by day 4. Martin–Martinez et al. also noted a focal mononuclear infiltration in the submesothelial area of the visceral peritoneum at day 5 after delivery of AdlacZ in 15 mL. The inflammatory response may vary between species of animal and the vector de- livered: Osada et al. did not see an inflammatory response in mice after delivery of a null vector or a vector containing the gene for erythropoietin (21).
A humoral immune response to the adenoviral vector itself results in the appearance of neutralizing antibodies against the viral capsid proteins within 7 days. The expressed protein may also elicit a humoral response, depending on its immunogenicity; for example, if a human protein is expressed in rodents. Setoguchi et al. found that anti-transgene antibodies raised against the initial administration of Ada 1AT prevented transgene expression from a second injection of this vector 1 week later, and that the antibody titer against a 1AT correlated with the virus load (16). When the interval between injections was lengthened to 3 months, a repeat injection was effective in producing measurable amounts of the transgene. Morrissey and colleagues have found that the humoral response was related to the dose of adenoviral vector used (20). We have found that a second IP injection 9 days after the first was possible but less effective, and produced a mild inflammatory infiltrate in the submesothelial tissue (8). In the lung, we have found that repeated applications of adenoviral vectors are possible, but with greatly reduced efficacy. The efficacy of repeated adenoviral administration was improved with concomitant use of a steroid for immunosuppression (22). The use of different viral serotypes for initial and subsequent administration may moderate host response.
The rapid loss of transgene expression noted by investigators is most likely due to a T-cell mediated response directed against transduced cells. We noted that the disappearance of β-gal-producing cells from peritoneal surfaces 7 days after delivery of AdlacZ to immunocompetent rats coincided with an increase in peritoneal lymphocytes, which were significantly above background levels by 4 days and remained elevated out to at least 14 days. Cyclosporine A treatment beginning 1 day prior to virus administration prevented an elevation in peritoneal lymphocytes and led to the maintenance of β-galexpressing cells on peritoneal surfaces out to at least 28 days (23). Steroids have also been used to reduce the anti-adenoviral immune response and prolong transgene expression (12). Immunomodulated animals provide another avenue for research where the immune response to the adenovirus can be limited and transgene expression prolonged. The nude rat, an athymic, T-cell deficient strain, has been used and would be a suitable model for PD research (24).
Functional Gene Transfer to the Peritoneal Cavity
We and other investigators have used adenovirus-mediated gene transfer of functional genes to the peritoneal cavity to investigate the role of a variety of genes in pathophysiological pathways. These have included proinflammatory cytokines, pro- and anti-fibrogenic factors, and angiogenic and antiangiogenic factors.
AdTGF-β: An Experimental Model of Peritoneal Fibrosis
Adenovirus-mediated gene transfer of TGF-β1 can provide an experimental model of peritoneal fibrosis, a condition commonly found in peritoneal biopsies from PD patients and thought to be a factor in ultrafiltration failure. TGF-β has been implicated in several studies as being involved in the progressive changes found in the peritoneum of PD patients (25).
A first-generation adenoviral vector containing the cDNA for the active form of TGF-β1 (AdTGF-β1) was delivered by IP injection in a rat model (14). This viral construct contains the human TGF-β1 gene that is mutated at residues 223 and 225; the resulting protein does not bind to its latency-associated protein, allowing production of active TGF-β1 (26). We have found that administration of AdTGF-β1 at doses of 0.75 – 2.0 × 109 pfu/ 150 μL in a 250 – 300 g rat, and 5 × 108 pfu/100 μL in a 140 – 160 g rat, gives good expression with minimal adenovirus-related toxicity. In preliminary studies using this same vector in mice, we have found that 1.5 × 108 pfu in 100 μL provides a histopathological effect similar to that observed in rats (19).
In our rat model, peritoneal effluent expression of TGF-β1 peaked 4 days after delivery and resulted in elevation of factors downstream of TGF-β, such as vascular endothelial growth factor (VEGF), plasminogen activator inhibitor-1, tissue inhibitor of metalloproteinase-1, and connective tissue growth factor, in mesothelial cells and peritoneal effluent, as well as an autocrine increase in rat latent TGF-β1 [Ref. (14) and Hoff C. M., unpublished results]. This change in cytokine and growth factor profile resulted in altered structural and functional membrane parameters, with increased submesothelial thickness and hypercellularity, increased collagen deposition, and angiogenesis (Figure 4). In addition, there were alterations in peritoneal transport with decreased net ultrafiltration and an increase in small solute transport that did not return to normal until days 21 – 28, coincident with a decline in peritoneal vascularization. These experiments, and those detailed below, were all controlled using a similar dose of the null adenovirus AdDL70. Adenovirus infection by itself did not appear to significantly impact peritoneal membrane function. AdDL70 caused a minimal reduction in net ultrafiltration at day 4, but values were indistinguishable from control by day 7 (14). There was no significant effect on parameters of small solute transport and the null virus appeared to have no lasting effect on peritoneal histology. The early inflammatory response in null adenovirus-treated animals was not associated with the induction of fibrogenic genes (27).
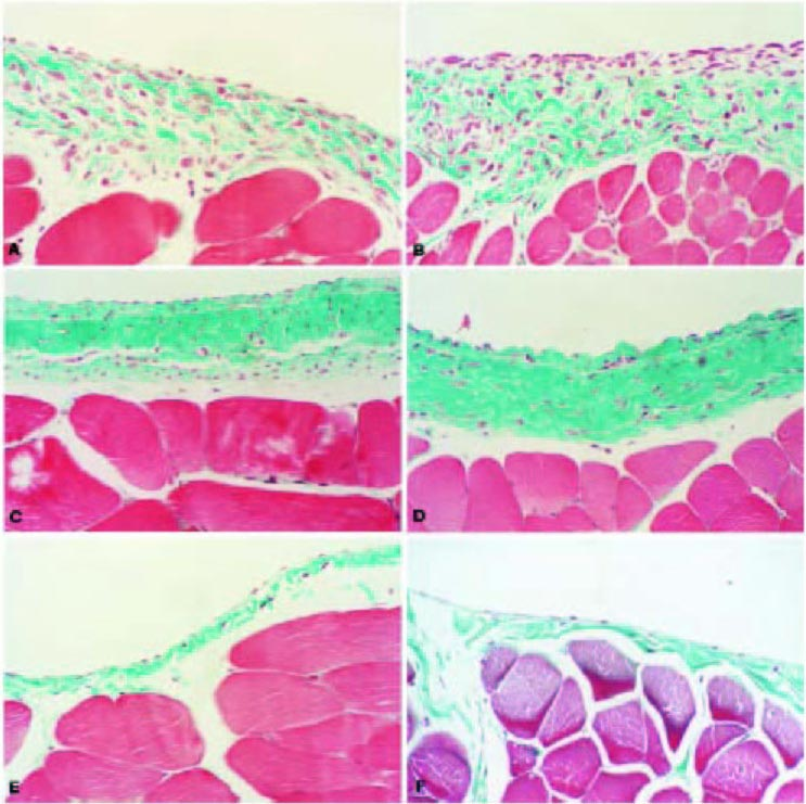
Histological features of the anterior abdominal wall after exposure to AdTGF-β1. On day 4 after infection, there is a fibroproliferative response, with increased thickness, cellularity, and vascularization (A). On day 7, these changes are exaggerated, with some increase in collagen deposition (B). By day 21, the cellularity of the peritoneal membrane has decreased but there is ongoing vascularization and extensive collagen deposition (C). The collagen deposition remains 28 days after infection but vascularity is decreased (D). Four days after control virus (AdDL70) infection, the peritoneum exhibits no acute virus-related changes (E). Normal rat peritoneum on day 21 after phosphate-buffered saline treatment is also shown (F). All sections Masson's trichrome stain. Magnification x200. From: Margetts PJ, Kolb M, Galt T, Hoff CM, Shockley TR, Gauldie J. Gene transfer of transforming growth factor-β1 to the rat peritoneum: effects on membrane function. J Am Soc Nephrol 2001; 12(10):2029–39. Reproduced by permission of Lippincott, Williams & Wilkins.
This model was also used to further elucidate the role of TGF-β1 in epithelial mesenchymal transition (EMT), a normal process involved in tissue healing and repair. Epithelial mesenchymal transition has also been identified in peritoneal tissues of PD patients and may be a contributing factor in the development of peritoneal fibrosis. Intraperitoneal delivery of AdTGF-β1 led to an early increase in genes associated with EMT and fibrosis, such as the zinc finger regulatory protein Snail, as well as genes for type 1 collagen A2 and alpha smooth muscle actin (27). By day 7 there was evidence of a transition to a myofibroblast phenotype and reorganization of the membrane, with cells exhibiting both an epithelial and a myofibroblast phenotype (Figure 3). These changes were resolving by day 14, with myofibroblasts less pronounced and restoration of the normal submesothelial architecture. These studies support the central role of TGF-β1 in peritoneal fibrosis and provide an experimental model system in which to study the sequence of events involved in TGF-β1-induced EMT.
The TGF-β -mediated model of peritoneal fibrosis was used to examine the therapeutic potential of FLT-1, a soluble VEGF type 1 receptor, and the soluble receptor for TGF-β, sTGFβRII, in blocking inflammatory angiogenic and profibrotic pathways respectively. Intraperitoneal delivery of AdTGF-β1 in a mouse model resulted in increased serum levels of active TGF-β1 and marked thickening of the parietal peritoneum with increased collagen deposition (19). Although we have less experience with this mouse model of peritoneal fibrosis, preliminary data suggest that the effects of AdTGF-β1 on peritoneal pathophysiology are similar in rat and mouse. An increased inflammatory response to the virus was noted with a transient weight loss, elevated levels of serum amyloid-P and interleukin (IL)-12, and increased monocyte chemoattractant protein (MCP)-1 gene expression. Pretreatment of recipient animals by intramuscular gene transfer of 100 μg of FLT-1 cDNA 4 days prior to induction of peritoneal fibrosis significantly attenuated the development of inflammation and fibrosis, as evidenced by an 81% decline in collagen deposition and decreased levels of intercellular adhesion molecule (ICAM)-1 and MCP-1 mRNA levels. On the other hand, sTGFβRII gene transfer, while reducing the levels of active TGF-β1, had no effect on TGF-β1-mediated collagen deposition and actually enhanced the inflammatory response, as seen by increased levels of MCP-1 and ICAM-1 mRNAs and circulating levels of TNF-α and IL-12. These findings suggest that, in this murine model of inflammation and fibrosis, intervening in peritoneal fibrosis by blocking VEGF signaling may be of therapeutic value. It also confirms both the pro- and the anti-inflammatory actions of TGF-β1.
Proinflammatory Cytokines IL-1β and TNF-α: Contributions to Peritoneal Inflammation and Fibrosis
Peritonitis is a common complication of PD, producing an acute inflammation that can have not only an immediate impact on membrane function, but long lasting effects on the membrane through the expression of pro-inflammatory cytokines and their downstream effector molecules (2,28,29). Intraperitoneal delivery of the adenoviral vectors for IL-1β (AdIL-1β) (30) and TNF-α (AdTNF) (31) was shown to induce changes in the peritoneal membrane of rats similar to those alterations seen in patient episodes of peritonitis (32). This strategy provides an experimental system in which to examine the individual roles of proinflammatory cytokines and their relevance to peritoneal pathophysiology.
Delivery of 1.5 – 2.0 × 109 pfu/mL of either IL-1β or TNF-α to the rat peritoneum led to an acute inflammatory response, an upregulation of VEGF, enhanced angiogenesis, and changes in solute permeability and ultrafiltration (32). This was accompanied by the transition to a profibrotic state, with increased expression of TGF-β, fibronectin, and collagen deposition. The magnitude of these changes and their resolution differed, however. TNF-α -induced ultrafiltration dysfunction normalized by 7 days and histological changes returned to baseline parameters by 21 days after infection, while increased vasculature, submesothelial thickening, and other indicators of fibrosis persisted in AdIL-1-treated animals. These differences were observed despite a similar dose of adenovirus being used, and a similar initial inflammatory and angiogenic response being elicited. This study helped to further delineate the effects of IL-1β and TNF-α on membrane structure and function, and may have relevance to the changes in patients experiencing an episode of peritonitis.
Adenovirus-Mediated Gene Transfer for Therapeutic Intervention: Angiostatin and Decorin
Adenovirus-mediated gene transfer can be used not only to establish a condition in the peritoneal environment, but also to intervene in a pathogenic process. Adenoviral vectors expressing decorin and angiostatin were delivered to the rat peritoneum to investigate the potential of antifibrotic and antiangiogenic intervention in an acute fibroproliferative model of PD. Decorin is a proteoglycan that can bind and inactivate TGF-β, and angiostatin is a recognized inhibitor of angiogenesis. In this study, animals received an IP injection of either the null virus AdDL70, AdDecorin, or AdAngiostatin 2 days prior to, and 7 days after, initiation of daily infusion with a high glucose dialysate (8). Four weeks of infusion [coupled with administration of lipopolysaccharides (LPS)] resulted in a thickening of the submesothelial layer, increased collagen deposition with angiogenesis, and evidence of membrane dysfunction with a decrease in net ultrafiltration and an increase in membrane permeability to small solutes (33). In rats that received AdDecorin, there was a reduction in hydroxyproline content of the membrane but no effect on angiogenesis and little impact on membrane transport parameters. Intraperitoneal delivery of angiostatin led to a reduction in number and density of peritoneal vessels, an improvement in net ultrafiltration and small solute transport, and a slight reduction in submesothelial thickness. This study, while preliminary, helps to define the individual contributions of peritoneal fibrosis and angiogenesis to peritoneal transport dysfunction, and demonstrates the value of the adenovirus-mediated gene transfer in not only evaluating the therapeutic potential of anti-fibrogenic and antiangiogenic molecules, but elucidating the role of these processes in membrane dysfunction.
Adenovirus-Mediated Gene Transfer for Systemic Delivery
Adenovirus-mediated gene transfer to the peritoneum has also been used for systemic delivery of an expressed protein to affect physiologic activity at a distant site. Intraperitoneal delivery of the gene for erythropoietin (AdEPO) was evaluated in cotton rats (34) and in a mouse model of polycystic kidney disease (35). Expression in rats led to a marked increase in the proportion of erythroid precursors throughout the 4 – 24 day observation period, with a significant increase in hematocrit that peaked at 14 days postdelivery. Intraperitoneal administration of AdCMV-EPO in DBA/2FG-pcy mouse, a model of polycystic kidney disease, improved the state of anemia. Serum EPO levels peaked at day 10, accompanied by an increase in circulating reticulocytes and an elevated hematocrit. In these models, it is likely that the transgene is expressed locally by the peritoneal mesothelium, but that the expressed protein is taken up and delivered to the systemic circulation via the lymphatics.
Summary
Adenovirus-mediated in vivo gene transfer provides an efficient means to study peritoneal membrane pathophysiology and potential therapeutic interventions. The experimental parameters for vector preparation and delivery have been well documented, as has the virus-specific response to the vector. Investigators have the opportunity to create an unlimited number of vectors for their research, thanks to a variety of do-it-yourself kits and companies devoted to constructing adenoviral vectors. The positive aspects of the adenovirus system must be balanced by its side effects, including acute inflammation and an immune response, and the transient nature of transgene expression, which makes the study of chronic expression of the transgene difficult. In this review, we have generally discussed the application of first-generation adenoviral vectors in PD research. These limitations of these vectors are outlined and should be addressed by the next generation of gutless vectors. Future studies in applying gene transfer to study the peritoneal membrane may also include the combination of gene delivery and chronic exposure to PD solutions, or gene transfer in uremic animals to more fully mimic the peritoneal membrane pathophysiology of PD patients.