
Abstract
Background
Peritoneal fibrosis (PF) limits the long-term use of peritoneal dialysis (PD), with effective therapies lacking. Ferroptosis, an iron-dependent cell death process, has been implicated in organ fibrosis, but its role in PD-related PF remains unexplored. Quercetin, a natural flavonoid, possesses potential anti-fibrotic and anti-ferroptotic properties.
Methods
PD effluent cells from patients with different dialysis durations were analyzed for the expression of fibrosis markers (α-smooth muscle actin and collagen I) and ferroptosis-related markers (glutathione peroxidase 4 (GPX4) and solute carrier family 7 member 11 (SLC7A11)). In vitro, human peritoneal mesothelial cells (MeT-5A) exposed to high glucose were treated with quercetin to examine its effects on mitochondrial ultrastructure and marker expression. A rat model of PF was established through daily intraperitoneal injection of high-glucose dialysate, with or without quercetin administration, to evaluate histological and molecular changes in the parietal peritoneum.
Results
Prolonged dialysis duration was associated with upregulated fibrotic markers and downregulated ferroptosis-related genes in patient samples. In vitro, high glucose induced mitochondrial damage and a profibrotic phenotype in MeT-5A cells, which were significantly attenuated by quercetin. Quercetin restored the expression of GPX4 and SLC7A11, comparable to the effects of the ferroptosis inhibitor ferrostatin-1. In vivo, quercetin treatment markedly alleviated high-glucose-induced peritoneal thickening and fibrosis while enhancing the expression of ferroptosis suppressors.
Conclusion
Our findings demonstrate that ferroptosis contributes to the pathogenesis of PD-associated PF. Quercetin mitigates fibrotic progression by modulating ferroptosis, highlighting its promise as a novel therapeutic agent for preventing or treating this complication.
Keywords
Introduction
Chronic kidney disease (CKD) is increasingly prevalent, and patients with end-stage kidney disease require kidney replacement therapy to maintain internal fluid–electrolyte homeostasis. 1 Peritoneal dialysis (PD) is favored for its ease of use and low cardiovascular impact, but long-term use can lead to peritoneal fibrosis (PF), a serious complication that affects treatment efficacy. 2 Approximately 3% of PD patients lose ultrafiltration capacity in the first year, increasing to over 30% by six years. 3 Factors, such as hypertonic glucose dialysate and acidic pH, damage mesothelial integrity, causing fibrosis and vascular issues. This process involves downregulation of E-cadherin (E-cad) and upregulation of α-smooth muscle actin (α-SMA), collagen I (COL-1), and fibronectin (FN), indicating mesothelial-to-mesenchymal transition (MMT) and contributing to fibrosis.4,5 Despite the clinical importance of PF, its initiating cellular processes and regulatory mechanisms remain poorly understood, and effective therapeutic strategies are lacking.
Ferroptosis is a form of regulated cell death characterized by iron-dependent lipid peroxidation and redox imbalance, primarily resulting from reduced expression of glutathione peroxidase 4 (GPX4). 6 GPX4 and solute carrier family 7 member 11 (SLC7A11) are key regulators of this process. SLC7A11, a component of the cystine/glutamate antiporter system Xc−, facilitates glutathione (GSH) synthesis and maintains GPX4 activity, with the latter essential for scavenging lipid peroxides. SLC7A11 deficiency drives GSH depletion, reduces GPX4 activity, promotes lipid peroxide accumulation, and triggers ferroptotic cell death. 7 Ferroptosis has been implicated in multiple pathological conditions, including cancer, inflammation, ischemia-reperfusion injury, neurodegeneration, and fibrosis in the heart, liver, lungs, and kidneys.8–10 Our preliminary findings have demonstrated downregulation of SLC7A11 and GPX4 in the peritoneal tissue of rats exposed to high-glucose dialysate, suggesting that ferroptosis may be a contributing mechanism in PD-associated PF. However, further evidence is needed to clarify its role, and effective ferroptosis-targeted interventions remain lacking.
Quercetin is a polyphenolic flavonoid widely found in fruits, vegetables, tea, and red wine. It exerts diverse biological activities, including antioxidant, anti-inflammatory, anti-tumor, and anti-fibrotic effects, largely through its capacity to reduce oxidative stress.11,12 Quercetin has been shown to regulate the activity of enzymes involved in GSH metabolism and exhibits significant anti-fibrotic activity in preclinical models of liver and kidney fibrosis.13,14 Moreover, it confers protection in murine models of diseases such as acute kidney injury, type 2 diabetes, and alcoholic fatty liver, in part through modulation of ferroptotic pathways.15–17 Therefore, it is of interest to investigate the inhibitory effect of quercetin on ferroptosis in PF, which has not yet been reported.
In this study, we aimed to further examine the role of ferroptosis in PF development and to evaluate the therapeutic potential of quercetin. We first analyzed PD effluent cells from patients with different dialysis durations and found altered expression of fibrosis- and ferroptosis-related markers. Consistent patterns were observed in high-glucose-treated MeT-5A mesothelial cells and in a high-glucose-induced rat model, both showing increased fibrosis and reduced GPX4 and SLC7A11 expression. These changes were significantly alleviated by quercetin treatment. Our findings suggest that ferroptosis contributes to PD-associated fibrosis and that quercetin may attenuate this process by modulating a ferroptosis-related mechanism, offering a potential therapeutic strategy.
Materials and methods
Chemicals
Quercetin (purity >98%) and Ferrostatin-1 (Fer-1) were purchased from Sigma and APExBio, respectively. For in vitro studies, stock solutions were prepared in dimethyl sulfoxide (DMSO) (quercetin: 100 mM; Fer-1: 10 mM) and stored at −20°C. For in vivo administration, Fer-1 was diluted in saline containing 0.5% DMSO to 2.5 mM, and quercetin was freshly prepared in Tween 80/saline at 10 mg/mL.
Patients’ recruitment and sample collection
Exfoliated mesothelial cells were isolated from the PD effluent of patients undergoing regular PD at the Department of Nephrology, Hunan Provincial People's Hospital, between August and December 2024. A total of 30 patients were enrolled and stratified into three groups based on treatment duration: new-onset (≤1 month), short-term (>1 to ≤12 months), and long-term (>12 months) (n = 10 per group). All patients were on continuous ambulatory PD and routinely used 1.5% or 2.5% glucose dialysate. Key exclusion criteria included active malignancy, peritonitis within the previous 3 months, current immunosuppressive therapy, or a history of major abdominal surgery. Baseline clinical characteristics of the patients are summarized in Online Supplemental Table S1.
Fresh PD effluent was collected after an 8-h dwell time. Primary human peritoneal mesothelial cells were isolated and purified using the standard adherence-based method. Briefly, exfoliated cells were harvested by centrifugation and resuspended in complete culture medium. The cells were then plated and allowed to adhere. Non-adherent cells were removed by washing with phosphate-buffered saline during medium changes within the first 48–72 h. The adherent cell population, which exhibits the characteristic cobblestone morphology of mesothelial cells, was expanded in culture for one week. Based on morphological uniformity and the established efficiency of the adherence-based purification protocol, we estimate the purity of the cultured peritoneal mesothelial cells to be >90%. This adherent, cultured cell population was subsequently used for total RNA extraction and gene expression analysis.
This study was conducted in accordance with the Declaration of Helsinki and was approved by the Institutional Review Board of Hunan Provincial People's Hospital (approval number: 2025-266). Written informed consent was obtained from all participants.
Immunofluorescence staining
Exfoliated mesothelial cells were isolated from 8-h PD effluent by centrifugation. After fixation in 4% paraformaldehyde and paraffin embedding, 3 μm sections were blocked with 10% goat serum (20 min) and incubated overnight at 4°C with an anti-vimentin polyclonal antibody (1:200; ThermoFisher). After secondary antibody incubation and 4′,6-diamidino-2-phenylindole (DAPI) counterstaining, images were acquired using a fluorescence microscope.
Cell culture and treatment
MeT-5A cell line (CRL-9444) was cultured at 37°C in Medium 199 containing 10% fetal bovine serum, 1 × insulin-transferrin-selenium supplement, 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 1.5 g/L sodium bicarbonate, 400 nM hydrocortisone, 3.3 nM epidermal growth factor, and 1 × penicillin/streptomycin. Upon reaching 60% to 80% confluency, cells were treated with the indicated concentrations of glucose solution (GS) ± Fer-1 (1 μM) or quercetin (20 μM) for 24 h. Cells were subsequently harvested for morphological examination, mitochondrial analysis, or molecular assays.
Cell viability
MeT-5A cells were seeded in transparent 96-well plates and allowed to attach overnight. The medium was refreshed, and cells were treated with or without 2.5% GS ± 5–40 μM quercetin for 24 h. Cell viability was assessed using the cell counting kit-8 (CCK-8) assay, and absorbance was measured at 450 nm using a microplate reader.
Transmission electron microscopy (TEM)
MeT-5A cells were fixed in 2.5% glutaraldehyde for 5 min, centrifuged, and further fixed in osmium tetroxide for 30 min. After washing, samples were dehydrated through graded ethanol or acetone. Epoxy resin infiltration was performed, followed by embedding and curing at 60°C to 70°C. Ultrathin sections (50–90 nm) were cut and mounted on copper or nickel grids. Sections were stained with uranyl and lead salts to enhance contrast, then rinsed and dried. Finally, the samples were imaged by TEM (ThermoFisher) and quantified using ImageJ software.
Animals
Specific-pathogen-free-grade male Sprague–Dawley rats (4 weeks old, 180–220 g) were obtained from Hunan Slake Jingda Experimental Animal Co., Ltd (license no. SYXK [Xiang] 2020-0017). Throughout the experiment, all animals were housed under standardized conditions of temperature, humidity, and light cycle, and maintained on an identical diet and water. Animal care and experimental manipulations were approved by the Institutional Animal Care Committee of Hunan Provincial People's Hospital under protocol no. 2021-138.
Quercetin administration
After acclimation, rats were randomly assigned to five groups (n = 6 per group): blank, model, solvent, Fer-1, and quercetin. Except for the blank group, all animals received daily intraperitoneal injections of 4.25% glucose PD solution (100 mL/kg) for 6 weeks. In addition, the solvent group received 1 mL of 0.5% DMSO daily; the Fer-1 group received Fer-1 (2.5 µmol/kg/day) in 0.5% DMSO; and the quercetin group received quercetin (50 mg/kg/day) via oral gavage. Solvent, Fer-1, or quercetin was administered at least 1 h prior to the glucose PD solution. All intraperitoneal injections were performed on the same side of the abdomen.
After 6 weeks, rats were euthanized under anesthesia, and peritoneal tissues were collected from the contralateral side (not used for injection). Tissues were either processed for histology or frozen at −80°C for molecular analyses.
Histological analysis
Parietal peritoneal tissues were fixed in 4% paraformaldehyde, dehydrated, embedded in paraffin, and sectioned at 5 μm. Sections were stained with hematoxylin and eosin (H&E) or Masson's trichrome according to standard protocols.
Peritoneal thickness was measured in Masson trichrome-stained sections under a light microscope. Eight high-power fields were randomly selected per specimen, and four measurements were taken per field. The average peritoneal thickness was calculated.
Quantitative real-time polymerase chain reaction (RT-PCR)
The TRIzol reagent (Aikerui, Changsha, China) was utilized to extract total RNA, with precisely 1 µg being subjected to reverse transcription using a reverse transcription-PCR kit (Aikerui). Additionally, the RT-PCR was conducted using an ABI 7300 instrument from Foster City, CA, USA, alongside the SYBR Green Pro Taq HS Mix (Aikerui) for the RT-PCR analysis. The conditions for PCR amplification included an initial denaturation at 95°C, followed by 40 cycles of denaturation at 95°C for 10 s and annealing/extension at 60°C for 30 s. The relative gene expression levels were quantified using the 2−ΔΔCt method. The primers used are listed in Online Supplemental Table S2.
Western blot
MeT-5A cells were washed with ice-cold phosphate-buffered saline and lysed in radioimmunoprecipitation assay (RIPA) buffer with protease inhibitors. Rat parietal peritoneal tissues were homogenized in RIPA buffer. Lysates were centrifuged, and supernatants were denatured with loading buffer. Proteins (20 μg) were separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels (6%, 10%, or 12%) and transferred to polyvinylidene fluoride membranes. After blocking with 5% bovine serum albumin, membranes were incubated with primary antibodies overnight, followed by a horseradish peroxidase-conjugated secondary antibody. Primary antibodies included anti-α-SMA, COL-I, SLC7A11, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Proteintech), GPX4, FN (Abcam), E-cad (BD Biosciences), and β-actin (Proteintech). Signals were developed with an enhanced chemiluminescence substrate and captured using a ChemiDoc system (Bio-Rad). Band intensities were quantified with ImageJ software.
Statistical analysis
Quantitative data are presented as mean ± standard error of the mean (SEM). Group differences were assessed using the t-test for two groups and one-way analysis of variance for multiple groups. Categorical variables were analyzed using the χ2 test. A P-value <0.05 was considered statistically significant. Statistical analyses were performed using GraphPad Prism version 10.1 or SPSS 25.0.
Results
Duration-dependent increase in fibrotic markers and decrease in ferroptosis regulators in PD patients
Analysis of exfoliated cells from PD patients revealed a strong correlation between dialysis duration and molecular markers of fibrosis and ferroptosis. Exfoliated cells were positive for the mesothelial marker vimentin (Figure 1(a)) and, after 1-week culture, exhibited adherent growth with a spindle-like or rounded morphology (Figure 1(b)), confirming their mesothelial origin. Notably, expression of the fibrosis-associated markers ɑ-SMA and COL-I was significantly elevated in the short-term group compared to the new-onset group, with a further increase observed in the long-term group. In contrast, E-cad expression showed an opposite trend (Figure 1(c)). Conversely, the expression of the key ferroptosis regulators GPX4 and SLC7A11 was significantly suppressed in the short-term group and declined even further in the long-term group (Figure 1(d)). This inverse relationship at the mRNA level suggests that ferroptosis is implicated in the pathogenesis of PF as early as the initial months of dialysis and may drive its progression during long-term PD.
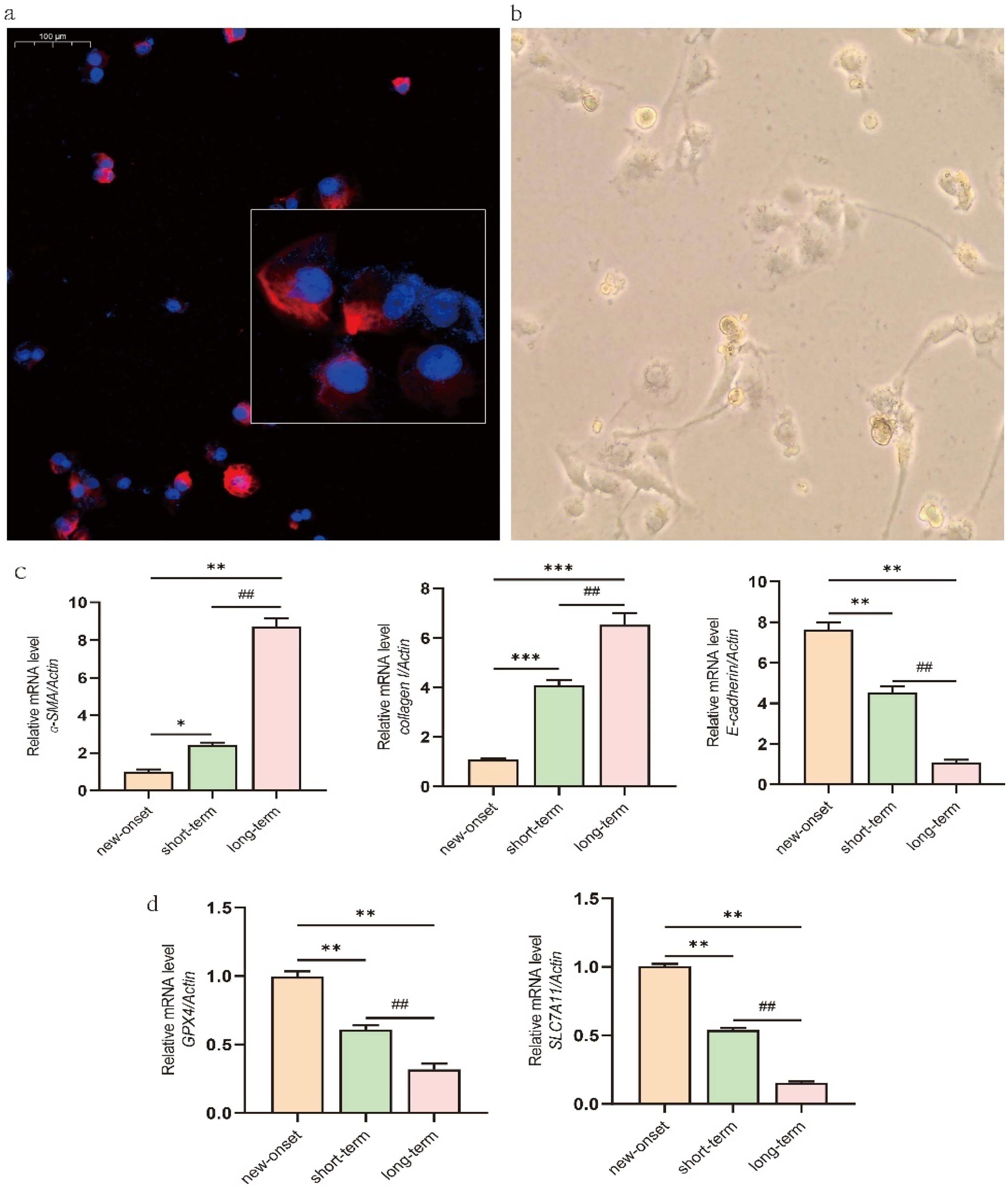
Identification of exfoliated mesothelial cells and mRNA expression of fibrosis- and ferroptosis-related markers. (a) Exfoliated cells collected from 8-h peritoneal dialysate from patients undergoing PD were subjected to immunofluorescence staining for vimentin (red) in the cytoplasm, with nuclei counterstained using DAPI (blue). 400× magnification. (b) After one week of culture, the primary exfoliated cells became adherent and exhibited a spindle-shaped or oval morphology (×100 magnification). (c) The mRNA expression of fibrosis-associated markers ɑ-SMA, E-cad, and collagen I was determined by qRT-PCR. (d) The mRNA expression of ferroptosis-related markers GPX4 and SLC7A11. Data are expressed as mean ± SEM (n = 10), or representative images are shown. *P < 0.05 and **P < 0.01 versus the new-onset group; #P < 0.05 and ##P < 0.01 versus the short-term group.
Quercetin attenuates high glucose-induced cytotoxicity, morphological alterations, and mitochondrial ultrastructural changes in MeT-5A cells
To establish an in vitro model of glucose-induced peritoneal injury, MeT-5A cells were exposed to different concentrations of high glucose (1.5% to 4.25%) for 24 h. High glucose triggered pronounced morphological alterations, including cell elongation and pseudopodia formation, indicative of a shift toward mesenchymal-like phenotypes (Online Supplemental Figure S1). These changes were accompanied by significant upregulation of the fibrotic markers α-SMA and COL-I at the mRNA level (Online Supplemental Figure S2). Since 2.5% glucose elicited a more moderate effect on cell viability while still inducing clear fibrotic responses (Online Supplemental Figure S1), this concentration was selected for subsequent experiments.
We then evaluated the protective effects of quercetin. After determining that quercetin at 5 to 20 μM did not significantly affect basal cell proliferation (Figure 2(a)), we found that pretreatment with these concentrations significantly attenuated the 2.5% glucose-induced decrease in cell viability, with 20 μM showing the strongest protective effect (Figure 2(b)). This concentration was therefore chosen for further mechanistic studies.
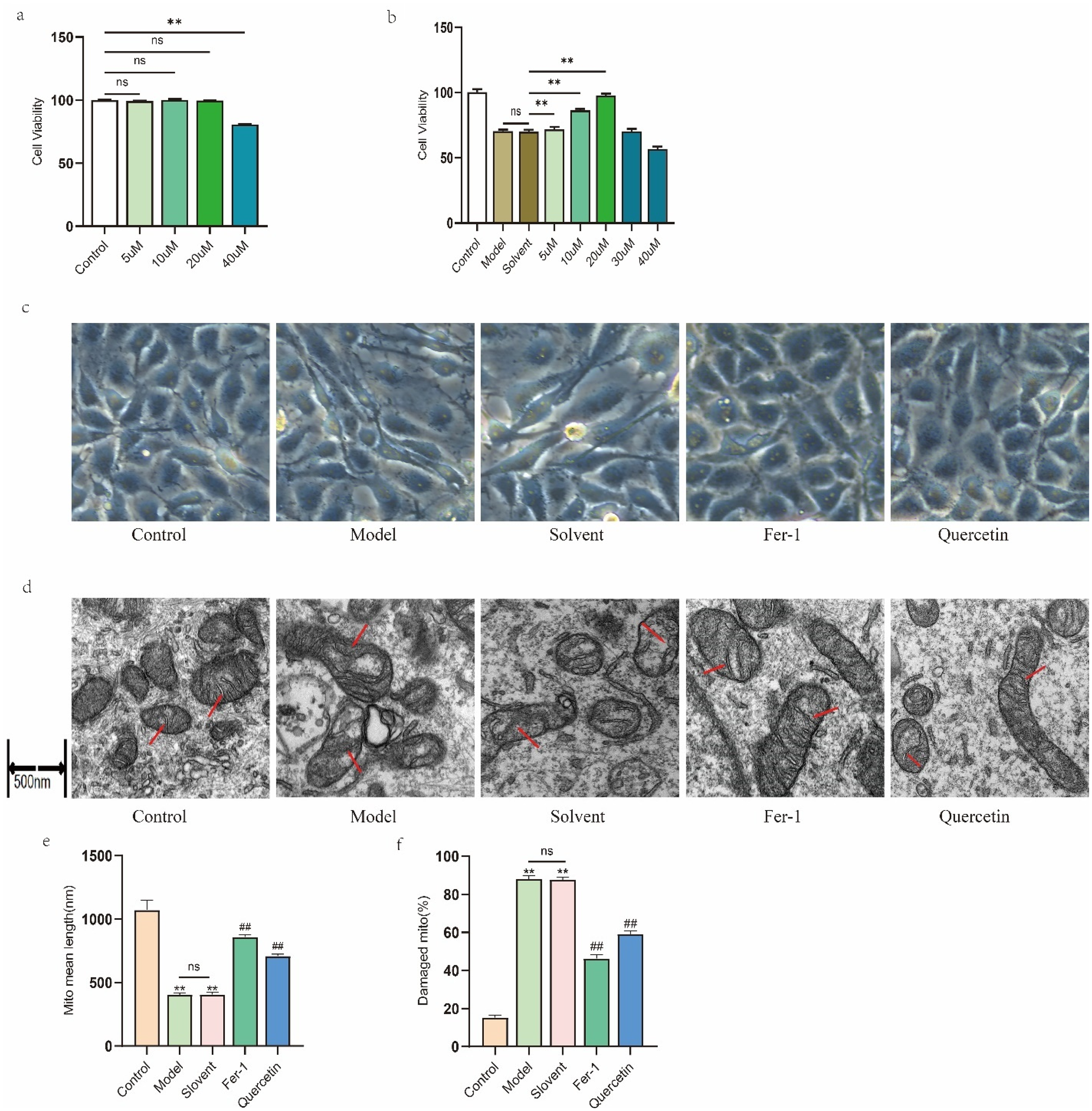
Effects of quercetin on high glucose-induced cytotoxicity, morphological alterations, and mitochondrial ultrastructural changes in MeT-5A cells. (a) Cell viability assessed by CCK-8 assay after 24 h treatment with 5–40 μM quercetin. (b) Cell viability after 24 h exposure to normal medium (Control), 2.5% glucose (Model), or 2.5% glucose co-treated with 0–40 μM quercetin. Solvent: 2.5% glucose plus DMSO for 24 h. (c) Representative light microscopy images (×200) of cells treated for 24 h with 2.5% glucose, in the presence or absence of 20 μM quercetin. Fer-1 (1 μM) served as a positive control, and 0.1% DMSO was used as the solvent control. (d) As in (c), but mitochondrial morphology was examined by TEM. Red arrows indicate characteristic changes (swelling, outer membrane disruption, and/or cristae loss). Scale bar = 500 nm. (e) and (f) Quantitative analysis of (c), where (e) represents the length of the mitochondrial long axis and (f) indicates the proportion of damaged mitochondria. Data are expressed as mean ± SEM (n = 3), or representative images are shown. **P < 0.01.
Light microscopy revealed that untreated cells exhibited a typical cobblestone-like morphology, whereas exposure to 2.5% glucose, with or without the DMSO vehicle, induced a spindle-shaped, mesenchymal-like appearance. Remarkably, co-treatment with either the ferroptosis inhibitor Fer-1 (1 μM) or quercetin (20 μM) largely preserved the normal epithelial morphology (Figure 2(c)).
TEM observation showed that glucose-exposed cells exhibited severe mitochondrial damage, including cristae loss, outer membrane disruption, and swelling. Further quantitative analysis confirmed that mitochondrial length was significantly reduced, while the proportion of damaged mitochondria was significantly increased. These ultrastructural abnormalities were markedly attenuated in cells co-treated with Fer-1 or quercetin (Figure 2(d) to (f)). These results suggest that quercetin preserves cellular and mitochondrial morphology under high-glucose conditions.
Quercetin antagonizes high glucose-induced fibrotic and ferroptotic responses in MeT-5A cells
To determine whether quercetin protects peritoneal mesothelial cells from glucose-induced cellular injury by modulating fibrosis and ferroptosis, we assessed the expression of related markers in MeT-5A cells. In the presence or absence of quercetin (20 μM) or Fer-1 (1 μM), the model group was treated with 2.5% glucose for 24 h, while the solvent group was treated with 2.5% glucose plus DMSO for 24 h. High glucose exposure significantly upregulated the fibrosis-associated markers α-SMA and COL-I at both mRNA (Figure 3(a)) and protein (Figure 3(b)) levels, while significantly downregulated the ferroptosis-associated markers GPX4 and SLC7A11. Meanwhile, compared with the model group, the solvent group showed no significant differences (Figure 3(c) and (d)). Quercetin treatment markedly reversed these glucose-induced alterations, showing efficacy comparable to that of the ferroptosis inhibitor Fer-1. These results suggest that quercetin attenuates glucose-induced fibrotic responses in MeT-5A cells, potentially through inhibition of ferroptosis.
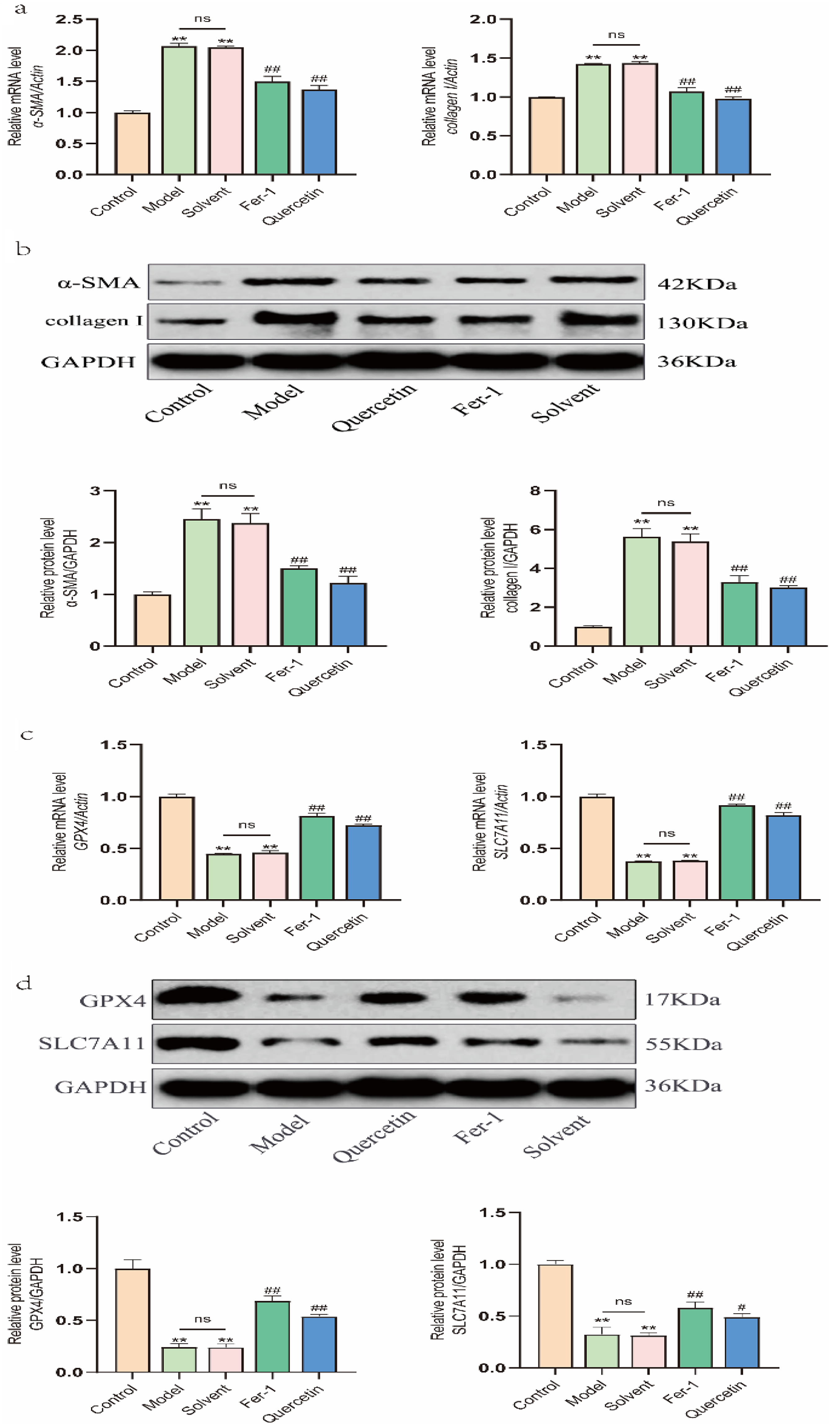
Effects of quercetin on high glucose-induced alterations in the expression of fibrosis- and ferroptosis-related markers in MeT-5A cells. Cells were treated with 2.5% glucose in the presence or absence of quercetin (20 μM) or Fer-1 (1 μM) for 24 h. (a) α-SMA and collagen I mRNA expression. (b) Protein expression of α-SMA and collagen I was assayed by Western blot, and the band densities were quantified and normalized to those of GAPDH. (c) GPX4 and SLC7A11 mRNA expression. (d) GPX4 and SLC7A11 protein expression. Data are expressed as mean ± SEM (n = 3) or representative blots are shown. *P < 0.05 and **P < 0.01 versus the control group; #P < 0.05 and ##P < 0.01 versus the solvent group or model group.
Quercetin ameliorates the histopathological progression of PF in rats
To evaluate the in vivo relevance of our findings, we employed a rat model of PF as described in the Methods section. H&E staining revealed that, compared to the blank control group, the parietal peritoneum in both the model and solvent groups was markedly thickened, with a disorganized and loose submesothelial matrix, noticeable inflammatory cell infiltration, collagen fiber deposition, and new capillary formation. In contrast, the Fer-1 and quercetin treatment groups exhibited reduced parietal peritoneal thickness, improved submesothelial matrix organization, and notably decreased collagen deposition and neovascularization compared to the solvent group (Figure 4(a)).
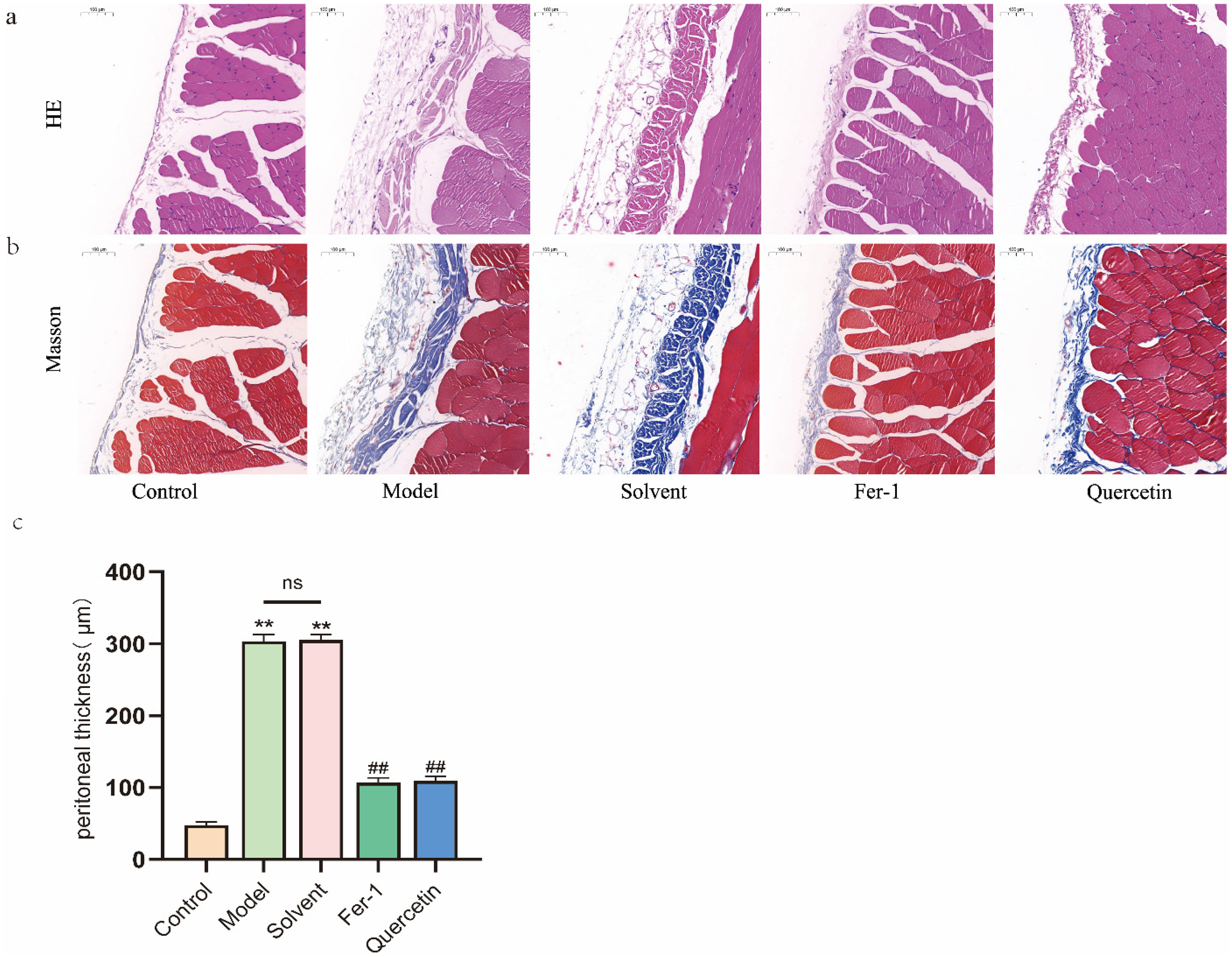
Effects of quercetin on peritoneal fibrosis induced by high-glucose dialysate in rats. Except for the normal control group, rats received daily intraperitoneal injections of 4.25% glucose peritoneal dialysis solution (100 mL/kg) for 6 weeks, concurrently administered with 1 mL of 0.5% DMSO (solvent), quercetin (50 mg/kg), or Fer-1 (2.5 µmol/kg) daily. (a, b) H&E staining and Masson trichrome staining of parietal peritoneal tissue. Scale bar = 100 μm. (c) Peritoneal thickness was measured in Masson-stained sections under a light microscope by averaging measurements from eight randomly selected high-power fields per specimen, with four measurements taken in each field. Data are expressed as mean ± SEM (n = 6), or representative images are shown. *P < 0.05 and **P < 0.01 versus the control group; #P < 0.05 and ##P < 0.01 versus the solvent group or model group.
Masson trichrome staining showed increased submesothelial collagen fiber deposition and new capillary formation in the model and solvent groups relative to the blank control group. These pathological changes were significantly ameliorated by both Fer-1 and quercetin treatment (Figure 4(b)).
Measurement of parietal peritoneal thickness revealed no significant difference between the model and solvent groups; however, both exhibited a significant increase compared to the blank control group. In contrast, treatment with Fer-1 or quercetin significantly reduced peritoneal thickening relative to the solvent group (Figure 4(c)). These results demonstrate that quercetin protects against PF induced by high-glucose dialysate in vivo.
Quercetin modulates expression of fibrotic and ferroptotic markers in rats with PF
To explore the mechanism underlying quercetin's protection against PF, we analyzed the expression of key molecular markers in the parietal peritoneum of rats. At the mRNA level, the model group exhibited significantly increased expression of FN and COL-I, alongside decreased E-cad, compared to the blank control group, changes that were not observed in the solvent group. Both Fer-1 and quercetin treatment significantly reversed these alterations, reducing FN and COL-I and restoring E-cad expression relative to the solvent group (Figure 5(a)). Consistent with these findings, protein analysis confirmed elevated FN and COL-I and reduced E-cad in model and solvent groups, effects that were markedly attenuated by Fer-1 or quercetin co-treatment (Figure 5(b)).
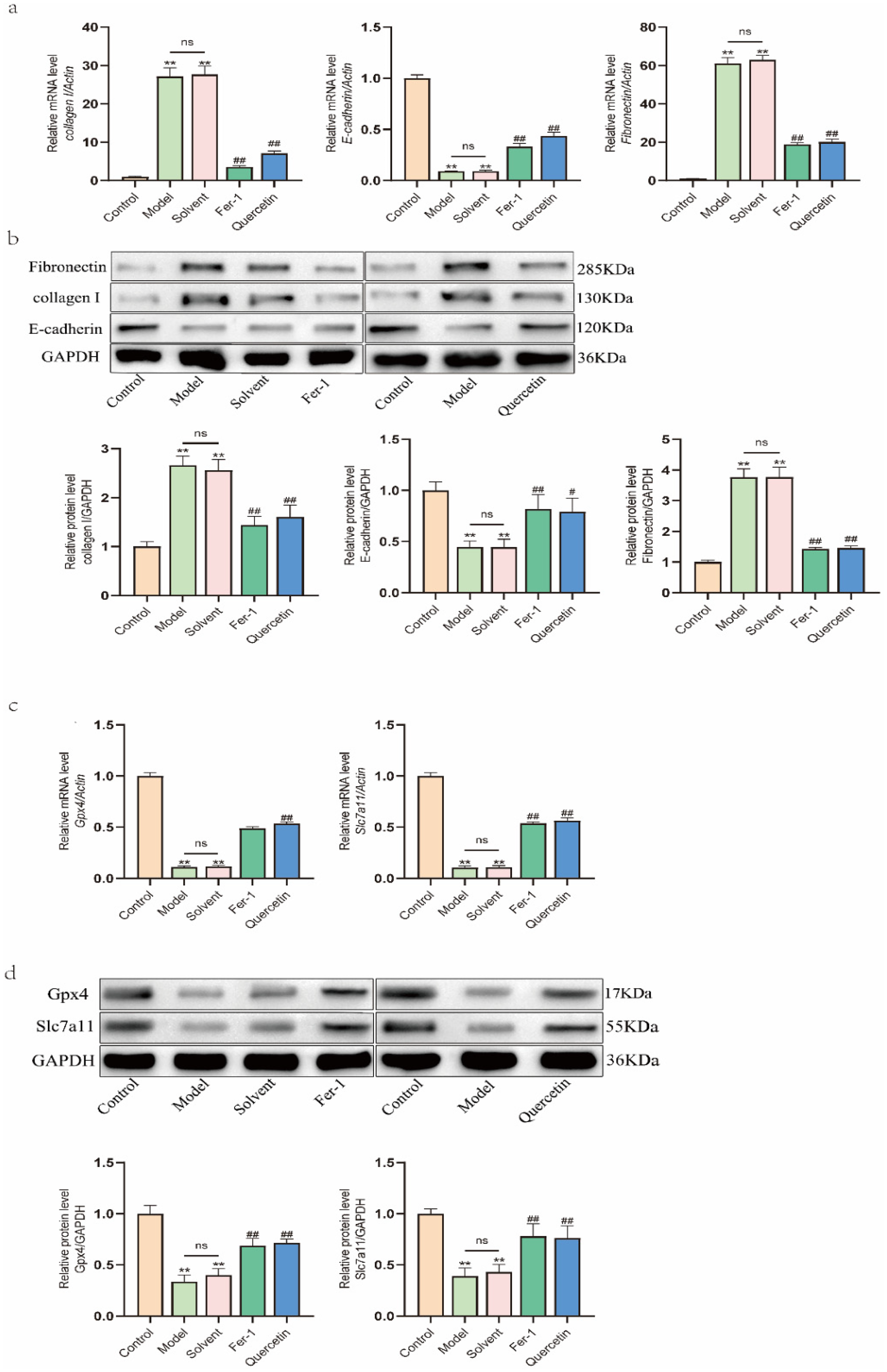
Effects of quercetin on the expression of fibrosis- and ferroptosis-related markers in rats treated with high-glucose dialysate. (a) The mRNA expression of fibrosis markers collagen I, E-cad, and FN. (b) The protein expression of collagen I, E-cad, and FN, with band densities quantified and normalized. (c) The mRNA expression of ferroptosis-related markers GPX4 and SLC7A11. (d) The protein expression of GPX4 and SLC7A11. Data are expressed as mean ± SEM (n = 6) or representative blots are shown. *P < 0.05 and **P < 0.01 versus the control group; #P < 0.05 and ##P < 0.01 versus the solvent group or model group.
We next evaluated markers associated with ferroptosis. mRNA and protein expression of Gpx4 and Slc7a11 were significantly downregulated in both model and solvent groups compared to blank controls. Importantly, Fer-1 and quercetin treatment robustly restored their expression (Figure 5(c) and (d)). These results indicate that quercetin counteracts the dysregulation of both fibrotic and ferroptotic markers in vivo, supporting the notion that its anti-fibrotic effects are mediated, at least in part, through inhibition of ferroptosis.
Discussion
PF remains a pivotal barrier to long-term PD due to an incompletely understood pathogenesis and a lack of effective therapies. Our study reveals ferroptosis as a novel contributor to PD-associated PF and identifies the natural flavonoid quercetin as a potent therapeutic agent that alleviates this condition by targeting ferroptotic dysregulation.
The development of PF centers on mesothelial injury under chronic high-glucose exposure. This condition promotes a glycolytic metabolic shift, activating hypoxia-inducible factor 1-alpha and upregulating profibrotic mediators.18–20 Concurrently, high glucose disrupts mitochondrial integrity and fosters lipid accumulation, exacerbating oxidative stress.4,21 However, we acknowledge that the cellular composition of PD effluent evolves with treatment duration, including potential variations in the relative abundance of mesothelial cells, fibroblasts, and immune cells. 22 In interpreting our patient-derived data, it is crucial to distinguish between changes driven by shifting cell populations and those indicative of active MMT. Our interpretation of MMT is supported by the coordinated expression pattern of key phenotypic markers, specifically the concurrent downregulation of the epithelial marker E-cad and upregulation of mesenchymal markers (e.g. α-SMA), within the same adherent cell populations from long-term PD patients. This dual pattern is an established hallmark of MMT and cannot be explained solely by an increase in the proportion of preexisting myofibroblasts.22,23 Within this pathogenic milieu, ferroptosis, a regulated cell death pathway executed through iron-dependent lipid peroxidation, emerges as a critical mechanism. 24 Our analysis of patient-derived cells directly links longer dialysis duration with a molecular signature of elevated fibrosis markers and diminished ferroptosis defenses (GPX4, SLC7A11), providing translational evidence for its role in human PF progression.
Ferroptosis has been increasingly implicated in fibrotic processes across various tissues. In doxorubicin-induced cardiomyopathy models, inhibition of ferroptosis via activation of the SIRT1/nuclear factor erythroid 2-related factor 2 (Nrf2) pathway alleviated cardiac dysfunction and reduced myocardial fibrosis. 25 In pulmonary fibrosis, PM2.5-induced ferroptosis accelerates fibrotic progression through heme oxygenase-1 (HO-1)-mediated mitochondrial dysfunction and NCOA4-dependent ferritinophagy. 26 Similarly, in a 5/6 nephrectomy-induced CKD model, ferroptosis modulators altered renal fibrosis independently of apoptosis. 27 Building on these established connections and our clinical findings, we utilized high glucose-treated MeT-5A cells to simulate the PD milieu. Beyond confirming the upregulation of fibrotic markers and suppression of SLC7A11 and GPX4, we observed hallmark ultrastructural features of ferroptosis, including mitochondrial shrinkage and outer membrane rupture, indicating its activation during mesothelial cell injury. Critically, these morphological and molecular changes were reversed by the ferroptosis inhibitor Fer-1, which preserved cellular architecture. These results substantiate the functional role of ferroptosis in propagating PF and underscore the therapeutic promise of its inhibition.
Fer-1, a well-characterized ferroptosis inhibitor, confers protection by integrating into cell membranes and directly neutralizing lipid peroxyl radicals, thus halting the propagation of lipid peroxidation. 28 Despite its utility in preclinical research, its translational applicability remains limited. In contrast, natural compounds derived from traditional herbal medicine, with their multi-target activities, favorable pharmacokinetic profiles, and established safety records, present promising candidates for ferroptosis-targeted therapies. Quercetin, a bioactive flavonoid abundant in herbs such as honeysuckle, forsythia, and formulations including Huangkui capsules and Xuefu Zhuyu capsules, has recently garnered attention for its anti-fibrotic properties.29–32 Growing evidence positions quercetin as a regulator of ferroptosis across disease contexts. For example, quercetin alleviated postmenopausal atherosclerosis in ovariectomized mice by mitigating lipid peroxidation, reducing iron deposition, and suppressing endothelial ferroptosis. 33 In cerebral ischemia models, it counteracted ferroptosis by decreasing levels of malondialdehyde, reactive oxygen species, and Fe2+, while upregulating GPX4 and ferritin heavy chain 1. 34 Furthermore, in diabetic nephropathy, quercetin inhibited ferroptosis by modulating iron homeostasis, enhancing GPX4 and SLC7A11 expression, and activating the Nrf2/HO-1 axis. 35 However, its role in regulating ferroptosis within the context of PF had not been investigated prior to this study.
Therefore, we evaluated the ability of quercetin to modulate ferroptosis in our in vitro PF model. In MeT-5A cells, quercetin significantly attenuated glucose-induced cytotoxicity, morphological alterations, and mitochondrial ultrastructural damage. Similar to Fer-1, quercetin reversed the upregulation of α-SMA and COL-I and restored the expression of GPX4 and SLC7A11, suggesting anti-ferroptotic activity. This effect is likely attributable to its antioxidant nature. Quercetin enhances the activity of antioxidant enzymes, including superoxide dismutase, catalase, and GSH peroxidase, while reducing lipid peroxidation products such as malondialdehyde. 15 Through activation of the Nuclear factor erythroid 2-related factor 2-antioxidant response element pathway, quercetin upregulates antioxidant genes (e.g. HO-1 and SOD1) and suppresses pro-oxidative signaling pathways (e.g. NLRP3 inflammasome, nuclear factor-κB, and mitogen-activated protein kinase), thereby contributing to redox homeostasis.36,37 Its phenolic structure also facilitates direct scavenging of free radicals, mitigating oxidative stress under both environmental (e.g. PM2.5 and ultraviolet B) and pathological (e.g. diabetes and ischemia-reperfusion) conditions.38,39 Additionally, quercetin-metal ion complexes may help regulate intracellular iron homeostasis, potentially influencing ferroptosis-related proteins such as ACSL4 and LOXs.13,40
In parallel, in vivo studies using a rat model of high-glucose dialysate-induced PF revealed that quercetin administration significantly alleviated peritoneal thickening and structural disruption. We speculate that quercetin exerted the anti-PF role by inhibiting the mesenchymal transition of peritoneal mesothelial cells, at least partially through modulation of the ferroptosis process. Molecular analyses revealed reduced expression of FN and COL-I, increased E-cad expression, and restored GPX4 and SLC7A11 levels in the parietal peritoneum following quercetin co-administration, with effects comparable to those of Fer-1. These findings reinforce the mechanistic link between ferroptosis and PF and demonstrate the in vivo efficacy of quercetin in modulating this pathway.
Several limitations of this study should be considered. First, a key consideration regarding our clinical findings is that the mesothelial cells analyzed were derived from PD effluent (i.e. detached cells), which may exhibit a phenotype distinct from that of the intact, adherent mesothelial layer. Effluent cells may be in a state of stress or injury, and their profile may not fully represent the functional peritoneal barrier. However, analyzing these cells remains clinically relevant, as their detachment and altered phenotype are direct consequences of bioincompatible dialysate and are related to peritoneal membrane dysfunction. 41 The consistent ferroptosis-related changes seen in these patient-derived cells, alongside the protective effects of quercetin observed in both adherent cells in vitro and intact peritoneum in vivo, support the pathophysiological relevance of our findings. Future studies using paired peritoneal biopsies would help delineate the contributions of different mesothelial cell states. Currently, no standardized protocols exist for establishing a rat model of PF. In this study, rats were subjected to continuous intraperitoneal injections of 4.25% glucose dialysate for 6 weeks. 42 Histopathological evaluation and fibrosis marker analysis confirmed successful induction of dialysis-associated PF. Although the selected quercetin dose was pharmacologically active and relatively safe, it remains unclear whether its anti-fibrotic effects are dose-dependent. Other limitations include the relatively small patient cohort size and grouping based solely on dialysis duration without adjusting for potential confounders such as underlying diseases. In both in vitro and in vivo models, the precise molecular targets of quercetin in regulating ferroptosis require further investigation.
Taken together, our clinical, cellular, and animal data consistently demonstrate that ferroptosis contributes to PF during PD and that quercetin exerts anti-fibrotic effects by preserving ferroptosis regulatory balance. These findings highlight quercetin's promising therapeutic potential to prevent or attenuate PF by modulating ferroptosis-related mechanisms, offering a viable strategy for clinical application.
Supplemental Material
sj-docx-1-ptd-10.1177_08968608261463605 - Supplemental material for Quercetin attenuates peritoneal fibrosis by upregulating ferroptosis-related glutathione peroxidase 4 (GPX4) and solute carrier family 7 member 11 (SLC7A11) in MeT-5A and rat models: Supported by clinical mRNA expression data
Supplemental material, sj-docx-1-ptd-10.1177_08968608261463605 for Quercetin attenuates peritoneal fibrosis by upregulating ferroptosis-related glutathione peroxidase 4 (GPX4) and solute carrier family 7 member 11 (SLC7A11) in MeT-5A and rat models: Supported by clinical mRNA expression data by Shaxi Ouyang, Yu Chen, Siyuan Yan, Xiangyi Li, Yunling Liu, Yuling Xiao, Guoli Li, Zangyue Sun and Xiaofang Hu in Peritoneal Dialysis International
Footnotes
Acknowledgements
We thank the Department of Laboratory Animals at Hunan Provincial People's Hospital for their assistance and support during this study.
Ethical approval
The research protocols for both animal experiments and patient-related studies were approved by the Ethics Committee of the First Affiliated Hospital of Hunan Normal University (Hunan Provincial People's Hospital), with ethics approval numbers 2021-138 and 2025-266, respectively.
Author contributions
SO, YC, and SY: Validation, conceptualization, and writing—original draft. XL and YL: Formal analysis and investigation. YX and ZS: Formal analysis and data curation. GL: Methodology and conceptualization. SO and XH: Resources, funding acquisition, conceptualization, and writing—review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Natural Science Foundation of Hunan Province, National Natural Science Foundation of China, and Hunan province college students research learning and innovative experiment project (grant numbers 2024JJ9280, 82500884, and S202510542396).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.