
Abstract
Spinal cord injuries (SCI) are extremely difficult to treat due to the limited capacity for neural regeneration across the injury site. However, V2a interneurons have been a point of interest in SCI research over the last decade, as they have been shown to contribute to the promotion of neuroplasticity after injury. These excitatory interneurons contain either long or short projections that are effective at driving rhythmic motor firing. By possessing ipsilateral, contralateral, or propriospinal projections, subtypes of V2a interneurons expressing the visual system homeobox-2 (Vsx-2) gene have been shown to extend their projections past the site of injury and restore injured spinal circuits that contribute to the respiration and right-left coordination. Moreover, Vsx-2/Zfhx3-expressing V2a interneurons in the midthoracic region of the spinal cord are a point of interest due to their unique ability to extend long projections caudally past the injury site and into the lumbar region, which resulted in substantial improvement in hind limb function after SCI in mice. Here, we collectively summarize the origin, subtypes, and the role Vsx-2 V2a interneurons play after SCI. We further describe the various techniques utilized to promote the accumulation and growth of these interneurons across or around the site of injury, effectively rewiring motor networks to contribute to functional recovery.
Introduction
Spinal cord injury (SCI) remains a significant medical challenge due to the limited capacity for neural regeneration in the adult central nervous system (CNS). The complexity of the injury environment, characterized by inhibitory molecules, the formation of an astrocyte border, and a lack of intrinsic regenerative capacity, necessitates innovative approaches to promote functional recovery. 1 Among the various cell types involved in neural regeneration, the Visual System Homeobox-2 (Vsx-2) V2a interneurons have emerged as critical players in promoting axonal regrowth and functional recovery after SCI.2,3
Vsx-2, as its name implies, is a homeobox gene known for its role in the development of the visual system. However, recent studies have highlighted its potential role in neural repair mechanisms outside of the visual system.2,3 Vsx-2 expressing V2a interneurons have the capacity to enhance neuroplasticity after injury or disease onset.4–7 Research has demonstrated that interneurons are capable of undergoing changes following SCI, such as modifications in their connectivity. These alterations can result in detrimental outcomes such as neuropathic pain, spasticity, and autonomic dysreflexia. However, interneurons are also a promising target for therapeutic intervention to support recovery. By activating dormant relay circuits and facilitating the establishment of new connections, interneurons play a crucial role in enhancing both motor and sensory functions. 8 Despite these promising findings, the mechanisms by which Vsx-2-expressing V2a interneurons enhance recovery after SCI remain to be fully elucidated.2,7 This review aims to provide a comprehensive overview of the current understanding of Vsx-2-expressing interneurons in the context of mammalian SCI, highlighting key studies, potential therapeutic applications, and future directions for research.
The VSX-2 Gene: Structure and Function
Vsx-1 and Vsx-2 are homeobox genes that are members of the PRX-superclass gene family.9,10 PRX genes can be categorized into paired-class (Prd-C) or paired-like (Prd-L) based on the presence of a paired-box domain, a paired homeodomain, or a combination of both. Prd-C genes feature either a solitary paired-box or both a paired-box and paired homeodomain, whereas Prd-L genes exclusively possess a paired homeodomain. 11 Prd-C genes encompass all Pax genes and various Drosophila genes like paired pox neuro and pox meso.12–14 Due to the presence of only a paired homeodomain, Vsx-1 and Vsx-2 are classified as Prd-L genes. Vsx-2, as well as Vsx-1, also contains a region known as the CVC domain, which is comprised of an additional 54 amino acids that are conserved and located next to the C-terminus of the homeodomain. This further categorizes Vsx-1 and Vsx-2 into a subclass of Prd-L genes known as paired-like: CVC (Prd-L:CVC) genes.9,10 These genes share the common characteristic of being highly expressed in both adult and embryonic retinas as well as sensory interneurons, highlighting their role within the neural network of the sensory system.15,16 In addition, Vsx-1 and Vsx-2 are expressed in interneuron populations in vertebrates’ spinal cord, hindbrain, and retina. 17
Vsx-2, also known as Chx10, HOX10, RET1, and MCOP2, is a 23 kb homeobox gene encoding for the 361 amino acid Vsx-2 protein.18–20 This protein functions as a transcriptional regulator by binding to DNA at the consensus sequence 5′-[TC]TAATT[AG][AG]-3′ located upstream of gene promoters. 21 The Vsx-2 protein is primarily involved in the development of the eye and plays a critical role in the proliferation and differentiation of sensory retinal progenitor cells.18–20 Vsx-2 also mediates the specification of V2a neurons in the spinal cord during development. 19
The Vsx-2 (or HOX10) gene is part of a larger family known as HOX genes in the spinal cord. Through interactions with morphogens, such as fibroblast growth factor (FGF) and retinoic acid, HOX genes play a critical role in regulating the positional orientation of neurogenic precursor cells in specific regions of the spinal cord.22,23 By interacting with gradients of accessory factors like FoxP1, HOX proteins play a significant role in determining the subtype diversification of motor neurons. 24 Table 1 summarizes the genes that directly interact with Vsx-2, based on Gene and BioGRID databases.
VSX-2 Interactor Genes in Homo sapiens Based on Gene-NCBI and BioGRID4.4 Databases a

These two databases were accessed on 07/10/2025.
ADP, Adenosine diphosphate; PKA, Protein kinase A; NCBI, National Center for Biotechnology Information; VSX-2, visual system homeobox-2.
Vsx-2 Expressing V2a Interneurons in Embryogenesis
In the process of spinal cord development, various neuronal subtypes are created through the interplay of contrasting morphogenetic gradients along the dorsoventral and rostrocaudal axes of the neural tube. The concentration gradients of various morphogens, such as transforming growth factor beta (TGF-β) and Sonic Hedgehog (Shh), establish positional cues or microenvironments, allowing specific neuronal precursor domains to take shape.25,26 This intricate process is fundamental to dorsal and ventral neurogenesis, with each region containing a number of neuronal groups that originate from distinct precursor domains (Fig. 1).
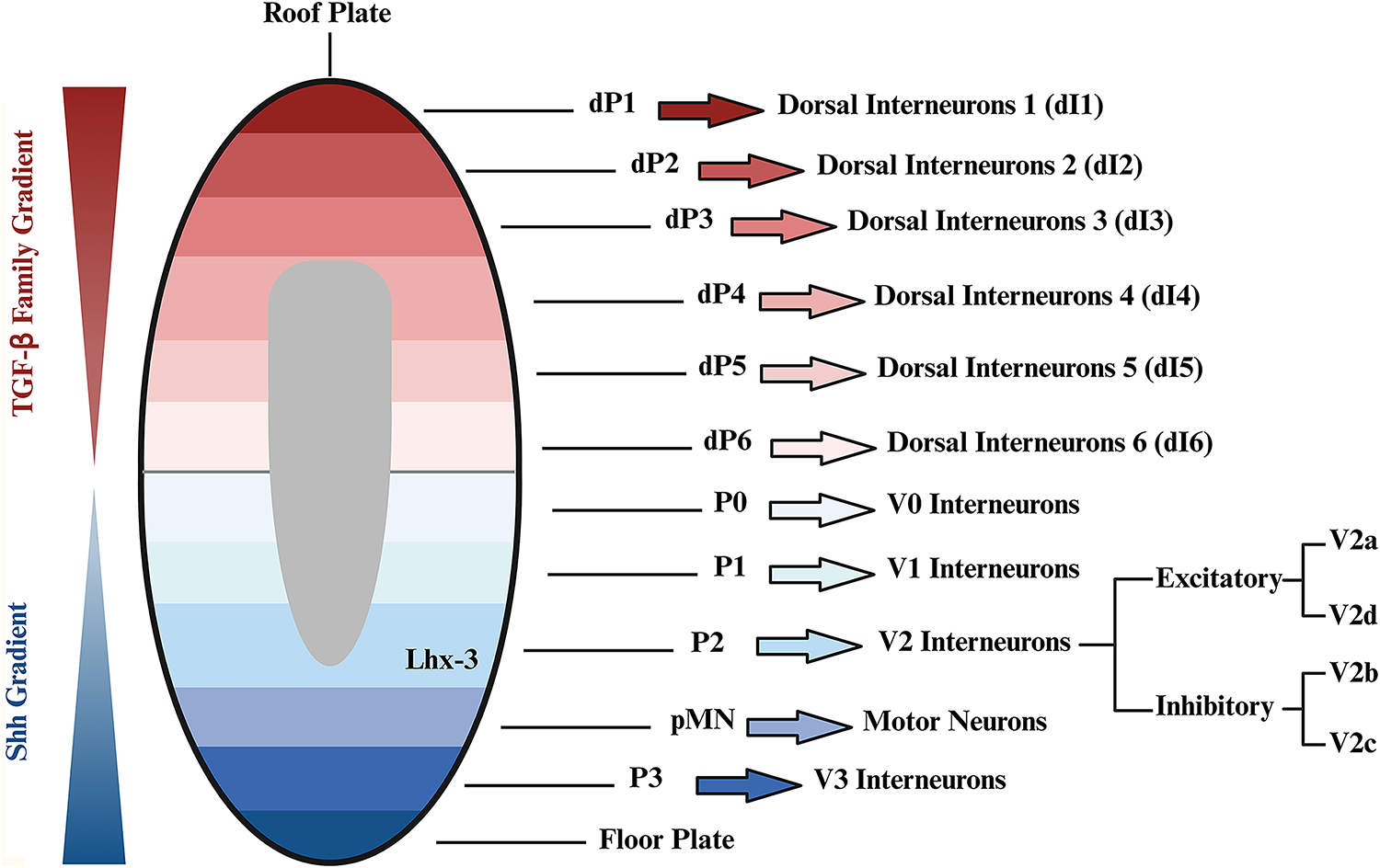
Dorsal and ventral development of spinal cord. During spinal cord development, various neuronal subtypes are formed through the interplay of morphogenetic gradients along the dorsal–ventral axis of the neural tube. The transforming growth factor beta (TGF-β) gradient forms the dorsal spinal cord during embryonic development. The dorsal spinal cord forms into six distinct progenitor domains (dP1 to dP6), which mature into dI1 to dI6. 27 Ventral neuronal patterning is influenced by the secretion of Sonic Hedgehog (Shh) from the notochord and later by the floorplate. The Shh gradient in the ventral spinal cord stratifies the area into five distinct progenitor domains: P0, P1, P2, pMN, and P3, each expressing unique transcription factors, shaping the V0, V1, V2, motor neurons, and V3 interneurons. 28 Interneurons from the V2 domain shape into both excitatory, V2a and V2d, and inhibitory, V2b and V2c, subtypes. Created using BioRender (July 2025).
The TGF-β gradient plays an essential role in forming the dorsal spinal cord during embryonic development, as it establishes the dorsoventral axis and shapes the neural tube. Specifically, the gradient of TGF-β and bone morphogenetic proteins (BMPs) is crucial in defining the nonneural ectoderm and the neural plate border. This gradient is meticulously regulated by various inhibitors, such as chordin and noggin, which modulate BMPs activity to ensure proper neural development.26,29 During embryogenesis, the TGF-β gradient drives the differentiation of neural progenitor cells (NPCs) into specific dorsal cell types, such as sensory neurons and interneurons. 30 Accurate regulation of this gradient is critical for developing a functional spinal cord, as disruptions can lead to developmental abnormalities and neurological disorders.29,30 Studies in model organisms, such as zebrafish and Xenopus, have shown that alterations in TGF-β signaling can result in defects in the development of the dorsal spinal cord, underscoring its importance during vertebrate embryogenesis.31,32 Under proper TGF-β gradient establishment, the dorsal spinal cord develops into six distinct zones, each characterized by specific gene expression patterns influencing the final resident cell fates. These zones, from dorsal to ventral, are dP1–dP6, which further develop into their mature states dI1–dI6 (Fig. 1). 27
Unlike the TGF-β gradient, which dictates the dorsal spinal cord development, the Shh gradient dictates cell fate on the ventral side of the spinal cord. 25 Shh, a protein secreted by the notochord and the floor plate, is recognized as the ventrally secreted signaling protein responsible for providing dorsoventral polarity to the ventral neural tube through its concentration-dependent function. Various concentrations of Shh control the identity and patterning of ventral neural tube cell types through their actions at multiple concentration thresholds.33,34 Shh gradient in the ventral zone plays a crucial role in stratifying this area into five distinct progenitor domains: P0, P1, P2, pMN, and P3 (Fig. 1). These five progenitor domains eventually shape the V0, V1, V2, and V3 interneurons. 28 In addition to the Shh gradient, an E3 ubiquitin ligase, RFN220, and a zinc finger protein, ZC4H2, were found to play a crucial role in the development of the ventral spinal cord. While being co-expressed and interacting with one another, these two proteins regulate the development of the ventral progenitor domains by targeting transcription factors Dbx1/2 and Nkx2.2 to be degraded (Fig. 2). The deletion of the RNF220 in mice resulted in the abnormal development of the ventral progenitor domains, such as loss of the P2 domain, which led to the complete abolition of V2 interneurons. In addition, the knockdown of RFN220 and ZC4H2 in the developing chick spinal cord resulted in the decreased expression of Vsx-2. 35 These findings highlight the importance of RFN220 and ZC4H2 in maintaining a healthy V2 interneuron population during development.
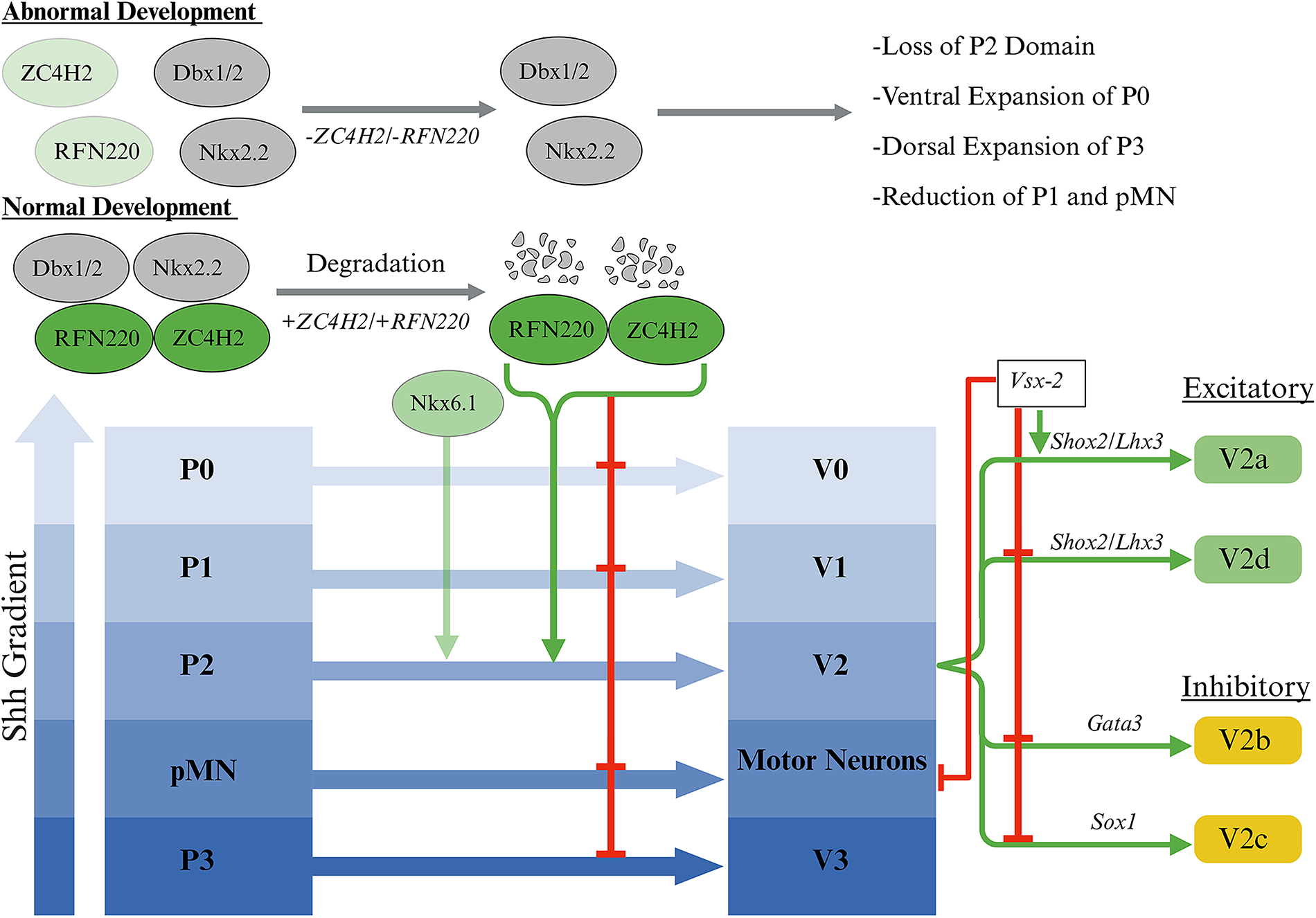
Development and regulation of ventral progenitor domains. An E3 ubiquitin ligase, RFN220, and a zinc finger protein, ZC4H2, are co-expressed and work together to degrade transcription factors Dbx1/2 and Nkx2.2, inhibiting the expansion of the P0 and P3 domains and reduction of the pMN and P1 domains. When co-expressed with the transcription factor Nkx6.1, RFN220 and ZC4H2 facilitate the development of the P2 domain, leading to the generation of Vsx-2-expressing V2a interneurons. 35 During normal development, V2 interneurons differentiate into excitatory (green) and inhibitory (yellow) subtypes. Excitatory subtypes include V2a, marked by the expression of Vsx2, Shox2, and Lhx3, as well as V2d, marked by the expression of just Shox2 and Lhx3. 36 Inhibitory subtypes include V2b, marked by the expression of Gata3, and V2c, marked by the expression of Sox1. 37 In addition, Vsx-2 suppresses the expression of genes associated with non-V2a interneurons and motor neurons. 38 Created using BioRender (July 2025).
The V2 interneuron family originates from p2 neuroepithelial progenitors that display a high degree of heterogeneity, resulting in the development of both excitatory and inhibitory subtypes through Notch1, Foxn4, and Dll4 signaling. V2b and V2c subtypes are inhibitory interneurons, with V2b being identified by Gata3 expression and V2c by Sox1 expression (Fig. 2). 37 On the contrary, V2a and V2d are excitatory interneurons. V2d is identified by Shox2 and Lhx3 expression, while V2a is identified by the expression of Shox2, Lhx3, and Vsx-2. 36 The expression of Vsx-2 also suppresses the expression of genes related to non-V2a interneurons and motor neurons (Fig. 2). 38
V2a Interneuron Subtypes and Their Function
The spinal cord primarily contains two types of neurons: projection neurons and spinal interneurons. The former is classified as neurons whose cell bodies lie within the spinal cord but extend their projections to other regions of the central or peripheral nervous system. In contrast, spinal interneurons are more confined, as their cell bodies and projections remain localized within the CNS. These interneurons can further be classified into different types based on their projection length and direction. Propriospinal interneurons contain ascending or descending long and short projections, whereas local spinal interneurons project their processes ipsilaterally (same side) or commissurally (across the spinal midline). 7
Most V2a interneurons fall within the class of local spinal interneurons, which function as premotor neurons that are responsible for excitatory rhythmic activity. These neurons project ipsilaterally and play a role in three crucial functions: (1) contributing to the proper functioning of the central pattern generator (CPG) by coordinating left-right movements, (2) supporting spinal respiratory circuits, and (3) playing a significant role in promoting neuroplasticity following SCI.2,4,39–41
The crucial role of V2a interneurons within the mammalian CPG was demonstrated by Crone et al. in 2008. 42 When Vsx-2-expressing V2a interneurons were genetically abolished in mice, the spinal cord failed to exhibit proper left and right firing alteration, resulting in disruption of left and right coordination. In addition, these mice displayed increased variability in the periodicity of locomotor-like activity, suggesting these interneurons play a role in stabilizing network activity. This study demonstrates V2a interneurons are responsible for integrating rhythm-generating inputs while also influencing both ipsilateral and contralateral circuitries to facilitate coordinated locomotion. 42 It is important to note the significant differences in CPG organization between swimming and walking animals, as highlighted by Rybak et al. 2015. 43 V2a interneurons in swimming animals, such as zebrafish, cannot initiate rhythmic burst activity as they do in walking animals, primarily due to the complexity of the mammalian CPG. 43
It is also important to emphasize that serotonin (5-HT) plays a critical and well-established role in modulating respiratory function, particularly in the context of SCI. Within the CNS, serotonin acts as a key neuromodulator that supports the excitability and plasticity of motor neurons, including those in the phrenic motor pool that directly innervate the diaphragm. After cervical SCI, descending serotonergic pathways originating in the brainstem are often disrupted, resulting in the loss of excitatory input to the phrenic motor neurons and further limiting the already reduced capacity for spontaneous recovery of breathing. Restoring this serotonergic innervation is therefore essential for reestablishing functional respiratory circuits.44–46 One strategy to address this challenge involves digesting inhibitory molecules, which has been shown to increase the presence of serotonergic fibers surrounding phrenic motor neurons within one week. 47 When combined with a peripheral nerve graft, this permissive environment supports not only local sprouting but also extensive regeneration of serotonergic axons across the injury site. The restoration of these fibers is crucial, as serotonin facilitates synaptic input and provides the excitatory drive within the phrenic motor circuitry necessary for rhythmic and tonic diaphragm activity. 47 Overall, serotonin is essential for reactivating and sustaining respiratory motor output after cervical SCI. V2a interneurons undergo significant depolarization after SCI in response to heightened serotonin sensitivity, which increases their plasticity. 48 Nevertheless, while V2a interneurons play an important role in respiratory motor control, their exact function is not completely understood.
While present throughout the CNS, the function of V2a interneurons differs between regions: hindbrain, cervical, thoracic, and lumbar. From most rostral, there is a network of V2a interneurons in the hindbrain’s medial reticular formation (Fig. 3A) that receive projections from neurons in the pre-Bötzinger complex and play a role in regulating respiratory function, as shown by Crone et al. 2012. 41 When V2a interneurons were genetically removed from the medial reticular formation in newborn mice, death or irregular breathing in surviving mice occurred, demonstrating the hindbrain V2a interneurons importance in regulating respiration in uninjured animals. 41 On the contrary, the population of V2a interneurons in the cervical region of the spinal cord does not have any role in regulating respiratory processes in uninjured animals (Fig. 3B). Instead, this population of interneurons only incorporates itself into the phrenic circuit after high cervical SCI. 4 In addition, cervical V2a interneurons are essential for the neuroplastic activation of accessory respiratory muscles during the progression of amyotrophic lateral sclerosis. 6 These facts indicate cervical V2a interneurons are not essential for respiration in healthy animals but aid in rewiring the circuitry to regain the function if it is lost or under a diseased state.4,6 Taken together, the V2a interneurons in the medial reticular formation of the brainstem are essential for respiration in healthy animals, but cervical spinal V2a interneurons aid in the recovery of respiration after injury or disease.4,6,41
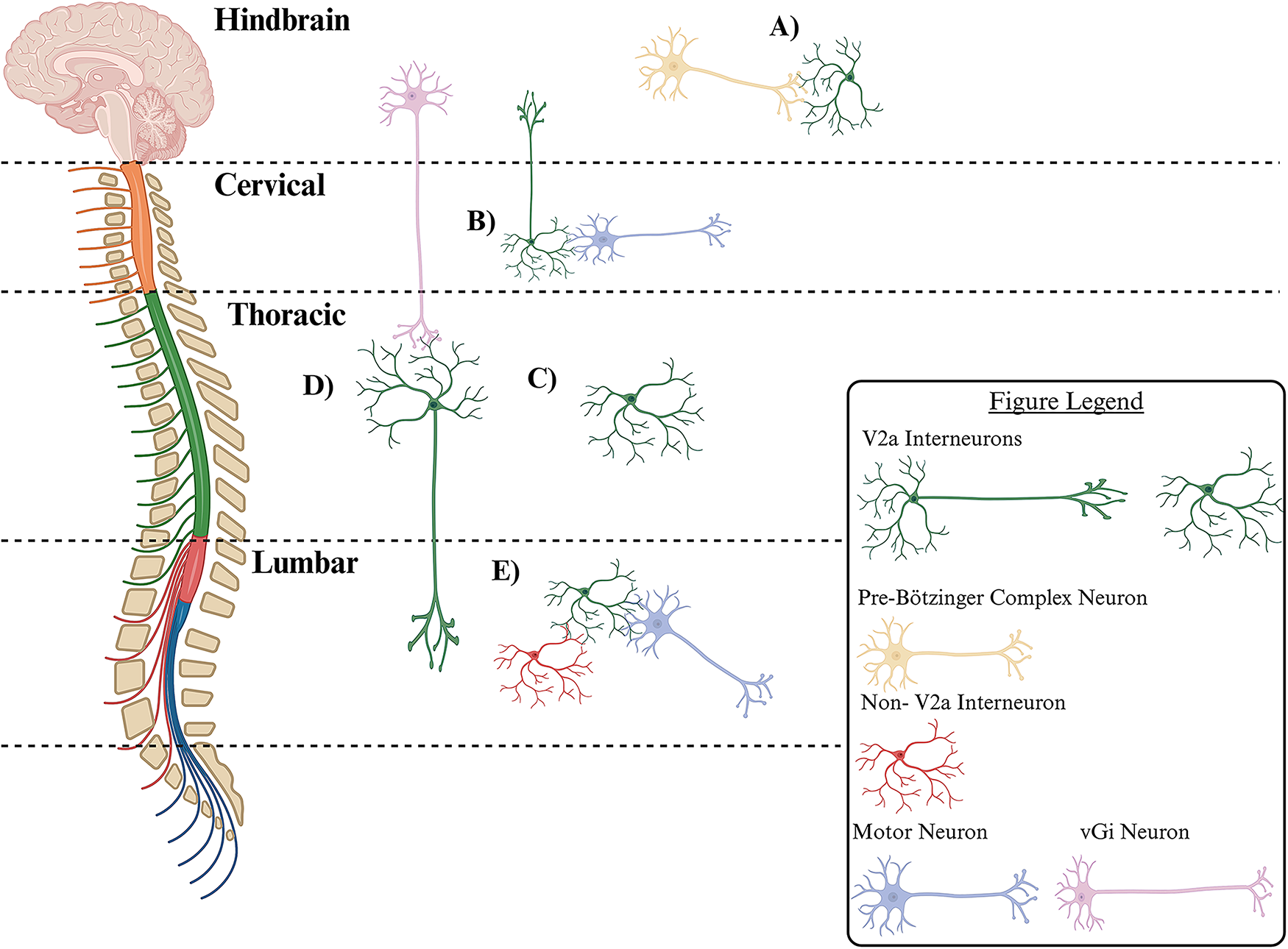
Distribution of V2a interneurons subpopulations in mammalian spinal cord.
Cervical V2a interneurons have also been referred to as branched type 2 V2a interneurons. Hayashi et al. (2018) described type 2 V2a interneurons to be enriched at the cervical level and downregulate the Vsx-2 transcription factor. 49 These type 2 V2a interneurons comprise the C3 to C4 propriospinal neurons, which are characterized by their bifurcating axons that project caudally to forelimb motor neurons and rostrally to the lateral reticular nucleus (Fig. 3B).49,51 Due to their projections, cervical V2a neurons are essential for transmitting descending motor commands to lower cervical levels and sending afferent motor copy signals back to the brain. 51 It is well-established that cervical V2a interneurons contribute to the control of skilled reaching and play a role in respiratory plasticity, as selective elimination of cervical V2a interneurons impairs forelimb reaching movements but does not impact locomotion.4,51
Importantly, Osseward et al. (2021) characterized two subgroups of Vsx-2-expressing V2a interneurons in the midthoracic region of the spinal cord: group-N and group-Z.2,50 Group-N interneurons express NeuroD2 and have short local projections, only extending to the midthoracic spinal cord (Fig. 3C). Group-Z interneurons have long-distance projections and express Zfhx3, receive caudal projections from ventral gigantocellular nuclei (vGi) neurons, and have dense projections extending into the lumbar spine (Fig. 3D).2,50 A recent study by Squair et al. (2023) demonstrated group-Z interneurons play a critical role in aiding mice to recover from SCI. By group-Z interneurons receiving projections from the vGi and possessing long distance projections to the lumbar spinal cord, supraspinal commands bypass the injury site in an incomplete SCI. Comparison between uninjured mice and injured mice who had regained walking function revealed the group-Z interneurons underwent the most transcriptional changes out of all the cells in the thoracic region of the injured spinal cord. Upon further Gene Ontology analysis, the transcriptional changes that occurred in these interneurons are those that are consistent with natural recovery processes, such as upregulation of dendritic spine morphogenesis, synaptic potentiation programs, and actin cytoskeleton reorganization pathways. 2 The most caudal population of V2a interneurons is classified as Type 1 V2a interneurons by Hayashi et al. (2018). These interneurons are reported to be enriched at the lumbar level and maintain expression of the Vsx-2 transcription factor. They have dense local connections that synapse onto other ventral interneurons or motor neurons and are efficient at driving motor neuron firing (Fig. 3E). 49
Other categories of V2a interneuron subpopulations were discovered in zebrafish and are not observed in mammals. Early-born V2a interneurons in zebrafish are known to be activated during intense movements, while later-born V2a interneurons are involved in slower, sustained movements. 52 Some V2a interneurons regulate timing by synapsing with other V2a and V0d interneurons, while others control amplitude by synapsing with local motor neurons. 53 Furthermore, Menelaou and McLean identified that dorsal V2a interneurons project longer distances than ventral ones. 54 Unidirectional V2a interneurons target slow motor neurons, and bidirectional ones target fast motor neurons.55,56 Zhong et al. (2010) identified three electrophysiological types of V2a interneurons—tonic, phasic, and delayed onset—that form synaptic connections with the same type and exhibit electrical coupling. 40 Though Dougherty and Kiehn found no electrical coupling during fictive locomotion, they noted that V2a interneurons impact left–right alternation firing.39,57
V2a Interneurons and Plasticity after Spinal Cord Injury
To date, there are no clinical therapeutic approaches that lead to substantial functional recovery after SCI. Most of the previous research focuses on regenerating or restoring lost pathways, rather than leveraging the body’s intrinsic neuroplasticity. Spinal interneurons are the predominant population of neurons in the spinal cord, playing a crucial role in regulating communication within and between neural networks. 58 Spinal interneurons are not only vital for the normal functioning of spinal networks but also are acknowledged to be vital for the plasticity and functional improvements following SCI. The interneurons achieve this by forming new neuronal pathways, including “bypass” routes, or by reorganizing synapses to reinforce existing pathways.59,60 By fostering spontaneous plasticity after injury, interneurons have the potential to be used therapeutically to improve outcomes after incomplete SCI. The research conducted by Bareyre et al. in 2004 was one of the first to anatomically illustrate that following a partial thoracic SCI, cervical interneurons played a crucial role in transmitting supraspinal signals to the denervated lumbar locomotor networks. After an incomplete SCI, hindlimb corticospinal tract axons formed new contacts with propriospinal interneurons in the grey matter and formed a new functional circuit able to relay supraspinal commands to the hindlimbs. 61 Ever since, interneurons have been targeted, modified, and stimulated to unravel the mechanisms behind their regenerative nature.
Among hundreds of types of interneurons, endogenous spinal V2a interneurons that express Vsx-2 have been identified as crucial components in promoting plasticity and driving therapeutic recovery following SCI (Table 2).2,62 One of the primary changes observed in V2a interneurons after SCI is a 100- to 1,000-fold increase in sensitivity to 5-hydroxytryptamine, commonly known as serotonin. 66 This heightened sensitivity can be attributed to SCI damaging the serotonergic fibers that extend from the brainstem to the spinal cord, resulting in a substantial decrease in serotonin levels caudal to the lesion. 67 As a result, the locomotor networks caudal to the lesion exhibit denervation supersensitivity to serotonin.68,69 Consequentially, V2a interneurons experience significant depolarization in response to serotonin, which increases their mean firing rate, enhancing plasticity after injury. 48
Summary of Key Studies on the Role of V2a Interneurons in the Spinal Cord
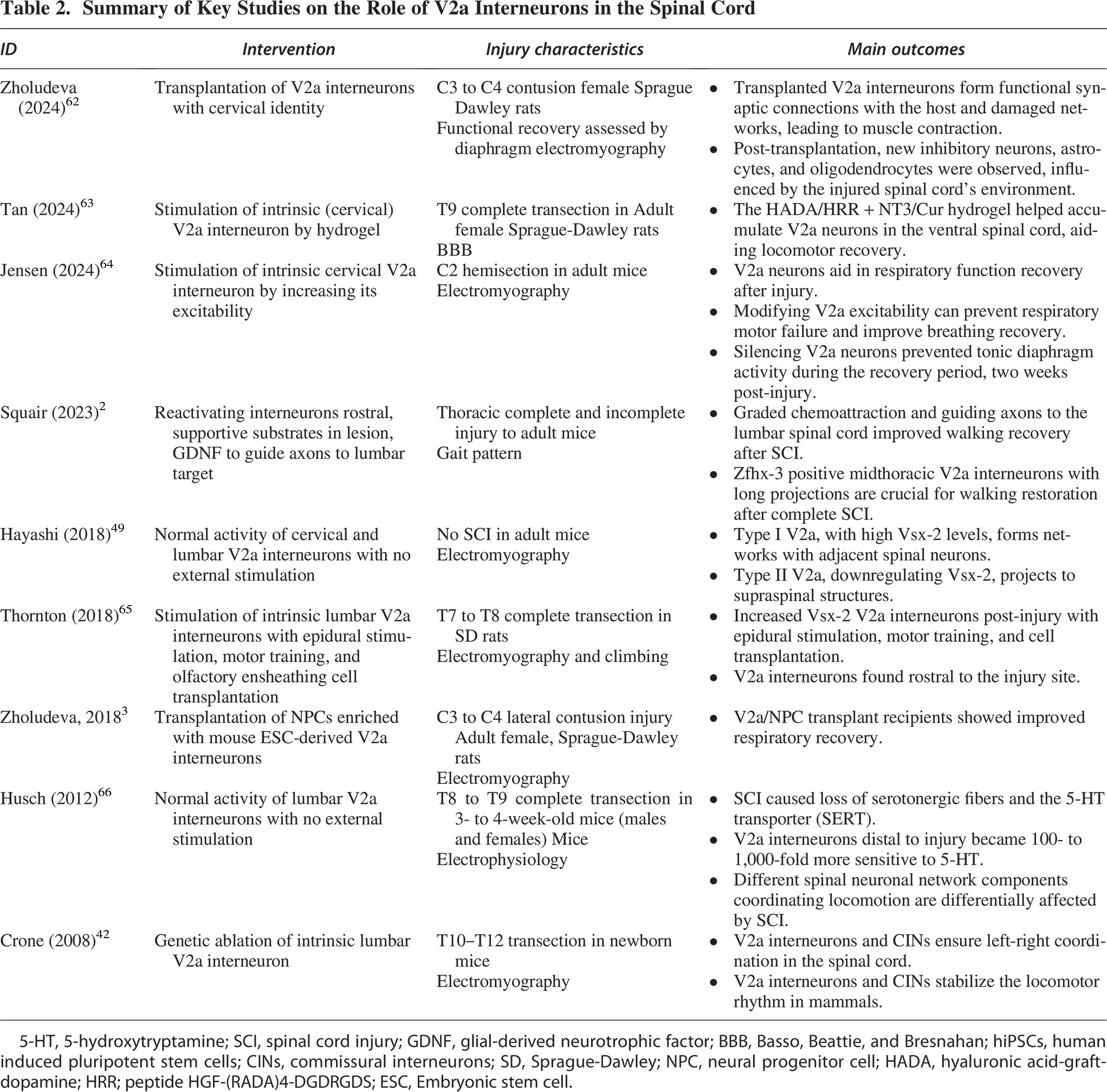
5-HT, 5-hydroxytryptamine; SCI, spinal cord injury; GDNF, glial-derived neurotrophic factor; BBB, Basso, Beattie, and Bresnahan; hiPSCs, human induced pluripotent stem cells; CINs, commissural interneurons; SD, Sprague-Dawley; NPC, neural progenitor cell; HADA, hyaluronic acid-graft-dopamine; HRR; peptide HGF-(RADA)4-DGDRGDS; ESC, Embryonic stem cell.
Two primary strategies are currently employing V2a interneurons to promote plasticity following SCI. The first strategy involves cell therapy of implanting stem cell-derived V2a interneurons into the injury site or promoting differentiation of precursors into V2a interneurons (Fig. 4A). The second strategy utilizes external stimulation to promote axon growth in V2a interneurons around the injured spinal cord (Fig. 4B).
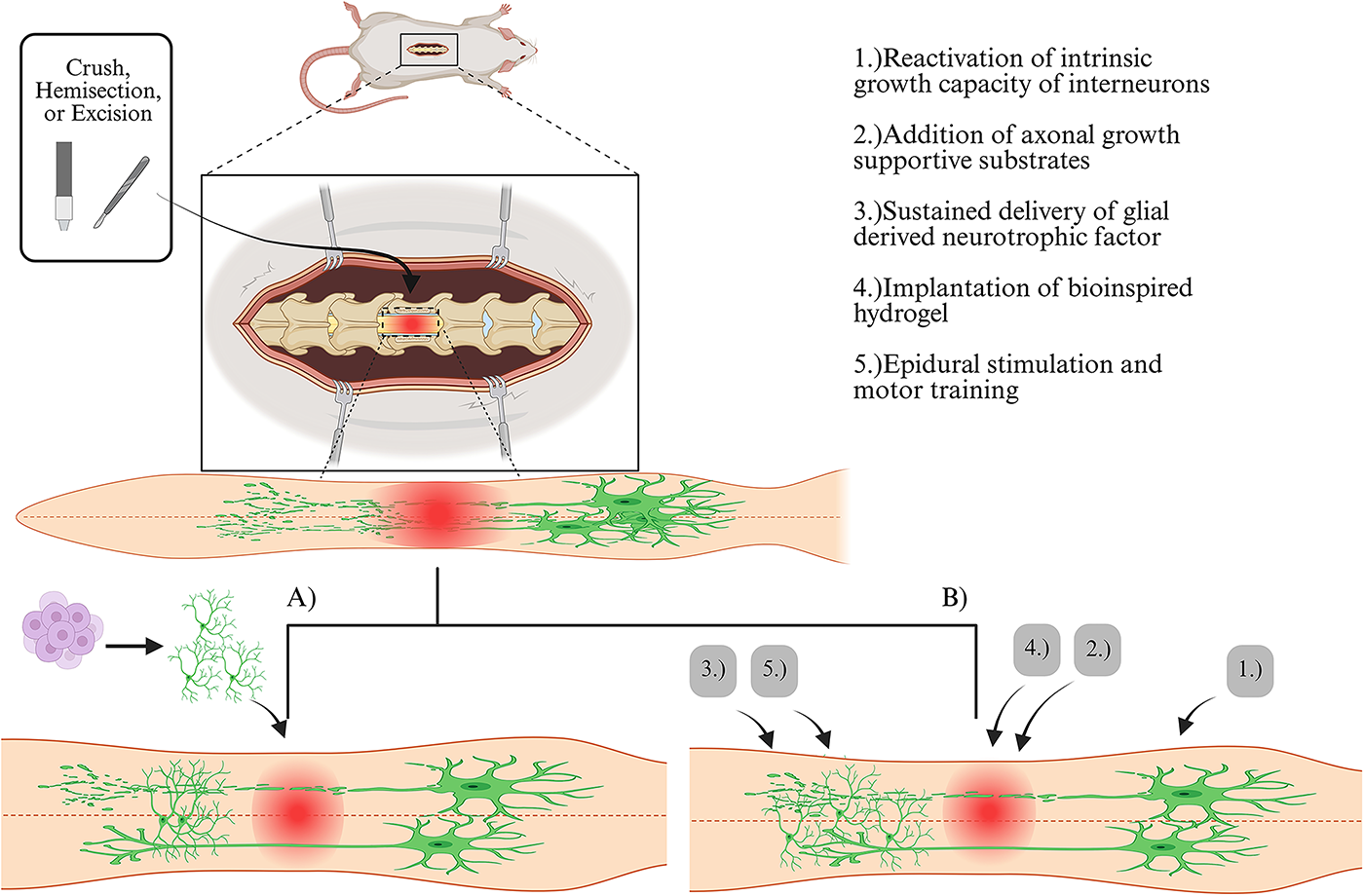
Strategies employing V2a interneurons after spinal cord injury (SCI): There are two main strategies being utilized to harness the potential of V2a interneurons in promoting plasticity after SCI.
The transplantation of excitatory V2a interneurons after cervical SCI significantly enhances respiratory recovery.3,62 Recent work by Zholudeva et al. in 2024 demonstrates that transplanting human V2a interneurons derived from human induced pluripotent stem cells (hiPSCs) with specific rostro-caudal identity into injured mouse spinal cords results in the restoration of injured relay circuits. These transplanted interneurons formed synaptic connections with both the host’s injured circuits and supraspinal pathways, leading to improved diaphragm activity and muscle contractions. In addition, the transplantation led to the emergence of new cell populations, including excitatory and inhibitory interneurons, astrocytes, and some oligodendrocytes. The new emergence of these cell types provides neuronal and glial building blocks necessary to support recovery. 62 Zholudeva et al. also conducted a study back in 2018, transplanting NPCs enriched with V2a interneurons derived from mouse embryonic stem cells into the injured cervical spinal cord. 3 Transplanted cells were observed to migrate, differentiate, and form neurite extensions one-month post-cervical injury. This suggests that the transplanted V2a interneurons can effectively integrate into the host’s neural circuits, facilitating improved connectivity and function in the injured spinal cord. Notably, mice receiving the V2a/NPC transplants showed significantly greater recovery from respiratory challenges than mice who received NPCs alone. This improvement emphasizes the critical role that V2a interneurons play in respiratory function following SCI and highlights the potential of using V2a interneurons to enhance recovery. 3 Recent work by Tucker et al., demonstrated NPC graft-derived V2a interneurons synaptically and functionally integrate into locomotor circuitry after thoracic SCI in mice. Using transsynaptic tracing, it was found that NPC transplantation produced low numbers of synaptic integration into host locomotor circuit. This finding highlights the potential benefit of using NPC-derived V2a interneurons to improve SCI outcomes while also emphasizing the need to enhance graft-to-host integration. 70
While transplanting cells into the injured spinal cord, it is important to consider the microenvironment formed by the glial border, as this has been shown to influence the transplanted cells’ final fate. Patil et al. (2021) demonstrated this when transplanting preoligodendrocyte progenitor cells derived from hiPSC into rats with a chronically injured spinal cord. They found when the glial border was intact, more precursor cells differentiated into V2a interneurons and motor neurons. However, when this procedure was combined with glial scar ablation, it resulted in a decreased yield of both V2a interneurons and motor neurons in the injury site, indicating the microenvironment of the injured spinal cord has significant effects on the fate of transplanted cells. 71
The challenge with the first strategy is that simply transplanting V2a interneurons may not be sufficient to correct the complex circuitry and connectivity in the host spinal cord, especially due to the specific roles of Vsx-2-expressing V2a interneurons in motor command circuits. This limitation arises because these neurons are part of a broader network that includes connections to supraspinal structures, and adding or transplanting individual V2a interneurons alone may not adequately restore this intricate network.49,60 Hence, a more comprehensive approach might be necessary for effective therapy.
Modifying the excitability of cervical V2a interneurons and promoting their accumulation in the ventral spinal cord are promising strategies for enhancing recovery after SCI. Recent studies have demonstrated that these approaches can lead to significant improvements in motor and respiratory functions following SCI. For instance, the implantation of a bioinspired hydrogel, engineered to have sustained release of neurotrophin-3 and curcumin, was shown to promote the accumulation of V2a interneurons in the ventral spinal cord. 63 This intervention resulted in the recovery of locomotor function in rats who underwent T9 spinal cord excision. By promoting the accumulation of specific neuron types, such as V2a interneurons, this approach could offer a promising strategy for restoring function in individuals with SCI. 63 Similarly, Jensen et al. (2024) demonstrated that modifying the excitability of cervical V2a interneurons is a proven strategy to prevent respiratory motor failure and enhance breathing recovery after cervical SCI. 64 Through the implementation of designer receptors exclusively activated by designer drugs, after a cervical hemisection injury in mice, Jensen and colleagues found that increasing the excitability of ipsilaterally projecting V2a interneurons restored rhythmic burst firing of the diaphragm as early as 4 h post-injury, with no adverse effects found after stimulation. Notably, silencing or decreasing the activity of V2a neurons was found to impair diaphragm activity ipsilateral to hemisection injury, thus solidifying the V2a interneurons’ role in promoting respiratory recovery after SCI. These findings suggest that precise control of neuronal excitability in V2a interneurons could be key to successful therapeutic outcomes. 64
Interestingly, Thornton et al. (2018) used a combination of the aforementioned strategies, which further clarified what type of implanted cells lead to the most promising outcomes after SCI. The strategy employed involved the transplantation of olfactory ensheathing cells (OECs) or fibroblasts (FBs) into the site of injury in combination with external stimulation, epidural stimulation, and motor training. Results favored implanting OECs over FB. Rats in the OEC group had more serotoninergic axons that crossed the rostral lesion border and a greater area of neurofilament-positive axons in the injury site compared with rats in FB group. Upon further analysis, thoracic neurons rostral to the injury were predominantly Vsx-2-expressing V2a interneurons, providing evidence that these neurons regenerate synaptic connections across the lesion. 65
In rodent models with lateral hemisection injuries, researchers demonstrate that descending propriospinal neurons spontaneously form new intraspinal circuits and relay functional signals across the lesion.2,73,74 Anderson et al. (2018) used a three-step approach to target descending propriospinal neurons for regeneration across the lesion: (1) reactivating the intrinsic growth capacity of interneurons rostral to the SCI with osteopontin, insulin-like growth factor 1, and ciliary-derived neurotrophic factor; (2) adding a delayed delivery of axon growth—supportive substrates, FGF 2, and epidermal growth factor—within the lesion; and (3) providing sustained delivery of chemoattractive glial-derived neurotrophic factor (GDNF) from a hydrogel depot caudal to the injury. This approach resulted in significantly more descending propriospinal axons growing through a complete crush injury in mice and rats. 72 Squair et al. (2023) used a similar approach with the exception of replacing hydrogel depots with lentivirus-mediated GDNF expression in the lumbar region, which resulted in extensive regrowth of axons to their natural target region and significantly improved hindlimb motor function after a complete spinal cord crush in mice. 2 The authors performed extensive experiments demonstrating Vsx-2+/Zfhx3+ midthoracic V2a interneurons, which have long projections to the lumbar spinal cord, are critical for restoring walking after an anatomically complete SCI.2,50 Therefore, targeting Vsx-2+/Zfhx3+ midthoracic V2a interneurons holds high therapeutic potential, as these interneurons are located close to the injury site and can relay supraspinal inputs across the lesion.
Future Directions
Studies have demonstrated that engineering and transplanting specific subsets of spinal interneurons can further enhance plasticity and functional recovery in individuals with SCI. 60 Recent studies demonstrated that transplanted V2a interneurons are not only connected to the host nervous system and effectively integrated into the host phrenic network but also play a role in enhancing diaphragm function following SCI. 62 Nonetheless, a conspicuous variability in the connectivity between donor V2a interneurons and the host phrenic network indicates required research in this area to broaden our understanding of functional neuronal relays after donor V2a interneuron transplantation. Another proposed field for future research is the specific restoration of 5-HT in V2a interneurons, as research has shown they become supersensitive to 5-HT up to 1,000 times due to the significant decrease in serotonergic concentration as a result of damage to serotonergic fibers projecting from the brainstem to the spinal cord.66,69 It is possible that serotonin restoration in these specific interneuron populations may result in recovery after SCI. In addition, a specific group of midthoracic V2a interneurons (Vsx-2+/Zfhx3+) was identified as critical for restoring walking after a T13 SCI in mice. 2 Therefore, it is likely that there are other V2a subpopulations of neurons within different regions of the spinal cord crucial for regeneration after SCI, which need to be identified. Finally, future studies should consider how to specifically increase synaptic input to these specific V2a subpopulations of interneurons and utilize them as a relay to propel the signal long distances past the injury.
Conclusion
Current therapeutic approaches for SCI have only resulted in minimal functional improvements, which may partially be attributed to researchers tending to concentrate on restoring lost pathways rather than harnessing the body’s innate neuroplasticity. Spinal interneurons play a crucial role in regulating communication between neural networks. Both lumbar and midthoracic V2a interneurons express the Vsx-2, while cervical V2a interneurons do not express Vsx-2. Studies have shown spinal V2a interneurons that express Vsx-2 have been identified as crucial components in promoting plasticity and driving therapeutic recovery following SCI. Vsx-2 interneurons have been leveraged to promote functional recovery by either transplanting V2a interneurons or externally stimulating intrinsic V2a interneurons in the spinal cord. Through exploring techniques to leverage Vsx-2 interneurons’ neuroplastic nature, a new subpopulation was recently identified at the midthoracic level. Zfhx-3 positive midthoracic Vsx-2 V2a interneurons with long projections to the lumbar spine were shown to be critical for restoring walking after SCI. Taken together, this strongly suggests that different subpopulations of Vsx-2 interneurons could be a therapeutic target for developing future treatments to improve motor function after SCI.
Authors’ Contributions
A.K. performed the literature research. Drafting of the article and critical revision of the article were performed by A.K., L.J.B., D.J.H., and A.S.H. All authors read and approved the final article.
Footnotes
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by the National Institutes of Health grant R01 NS136564-01.
Copyright Consideration
All figures included in this article were created using Canvas and BioRender.
Data Availability
This is a review article and does not contain any primary data. All data discussed and analyzed in this review are derived from previously published studies, which are cited throughout the text.