
Abstract
Background
Abdominal Henoch - Schönlein purpura (AHSP), being the most prevalent form of Henoch - Schönlein purpura, has a significant impact on the short - term prognosis of the disease and often involves the kidneys, leading to renal complications that affect children's long - term prognosis. However, the existing early assessment criteria for AHSP and its renal complications are inadequate. The urinary proteome may offer valuable insights.
Objective
To confirm the significance of urinary proteomics in the early detection of AHSP and its renal complications in children.
Methods
The urinary proteome of AHSP patients (with and without renal involvement) was compared with that of healthy controls using liquid chromatography - tandem mass spectrometry (LC - MS/MS) in data - independent acquisition (DIA) mode. Differentially expressed proteins were analyzed through Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis. Mfuzz was employed to analyze the expression levels of proteins related to disease onset and progression. The STRING database was used for protein - protein interaction analysis of relevant biological pathways. Selected differential proteins were verified using parallel reaction monitoring (PRM).
Results
A total of 441 dysregulated differentially expressed proteins (DEPs) were associated with the pathogenesis of AHSP, mainly related to cell adhesion, signal transduction or regulation, and reactions or pathways mediated by inflammatory cells or factors, and predominantly enriched in the lysosomal pathway. A total of 275 DEPs related to renal complications of AHSP were mainly associated with immune processes mediated by immunoglobulins, predominantly enriched in the regulatory pathways of the actin cytoskeleton. Time series clustering analysis identified 10 discrete clusters; three upregulated and two downregulated clusters were chosen to form respective panels. These panels involved various biological processes such as immune and inflammatory processes, lipid metabolism, glycosylation, coagulation, oxidative detoxification processes, and the Wnt signaling pathway, with several important biological pathways being enriched. Protein - protein interaction analysis of key pathways revealed three distinct MCODE networks, mainly involving proteins related to immunity, coagulation, collagen, and integrins. In the validation phase, at least eight urinary proteins useful for diagnosing AHSP or its renal complications were identified, demonstrating good diagnostic performance.
Conclusion
This study offers novel perspectives on the pathogenesis of AHSP and its renal complications in children, and the related proteins may serve as potential biomarkers for diagnosing AHSP and identifying the onset of renal damage. The findings of this study emphasize the importance of urinary proteomics in understanding the disease mechanisms and provide a basis for further research on early diagnosis and treatment.
Keywords
Introduction
Henoch - Schönlein purpura (HSP) is an acute systemic vasculitis mediated by immune complexes, characterized by a classic triad of symptoms: purpuric rash (without thrombocytopenia), arthritis, and abdominal pain. 1 It is the most common form of systemic vasculitis in childhood, with an annual incidence ranging from 10 to 30 cases per 100,000 children under 17 years old, predominantly occurring in children under 10 years of age.2,3 Based on clinical manifestations, HSP can be classified into several subtypes, and the abdominal form is one of the most prevalent, affecting approximately two - thirds of pediatric patients diagnosed with HSP. 4 This subtype presents a range of gastrointestinal symptoms, including abdominal discomfort, flatulence, diarrhea, nausea, and in some cases, gastrointestinal hemorrhage. 5
Abdominal - type HSP is often regarded as an acute and severe condition in pediatric patients. However, the initial symptoms can be nonspecific, making early clinical diagnosis difficult, especially when the symptoms are mild or atypical. 6 A delayed diagnosis can have serious consequences for some patients. In severe cases, abdominal - type HSP can lead to intestinal obstruction and intussusception. 7 In recent years, specific laboratory markers and clinical scoring systems have been used to diagnose and assess the severity of HSP. Additionally, diagnostic modalities such as esophagogastroduodenoscopy and imaging techniques like abdominal ultrasonography and computed tomography have been used to evaluate the extent of gastrointestinal involvement.8–10 Nevertheless, current diagnostic methods have limitations. They may not facilitate early detection or ensure diagnostic accuracy and can be invasive and inconvenient for repeated assessments. Therefore, there is a need to develop and study novel, non - invasive biomarkers for abdominal - type HSP to improve clinical decision - making.
Monitoring for kidney injury as a complication is crucial in the diagnosis and management of abdominal - type HSP. Persistent purpura, recurrent abdominal pain, and gastrointestinal bleeding are significant risk factors for kidney injury in HSP patients.11–14 Renal involvement usually occurs within the first or two months after the onset of HSP, and about 2% to 5% of patients may progress to end - stage renal failure, 15 which can significantly affect the quality of life of affected children. Currently, there is a lack of early pathogenesis and biomarkers for kidney injury in abdominal - type HSP. Although kidney biopsy has been considered the gold standard for diagnosing renal injury in HSP patients, 16 it is often not used to monitor the clinical course or evaluate the response to treatment due to associated risks, patient reluctance, and potential delays.17,18 As a result, the common diagnostic approach for kidney injury in pediatric HSP patients still depends on detecting urinary protein and/or hematuria, which has a relatively low detection rate of 30–50%.17,19,20 Moreover, routine urine analysis may not detect disease progression over time, especially in the early stages of HSP with minimal renal damage. This hinders timely diagnosis, personalized treatment, and preventative measures. Thus, there is an urgent need to find new and clinically relevant biomarkers to improve the prediction of kidney damage in abdominal - type HSP.
We have noted that both serum metabonomics and urine metabonomics studies have provided new insights into the small - molecule metabolic changes associated with the progression from HSP to Henoch - Schönlein Purpura Nephritis (HSPN),21,22 enhancing the predictive ability for kidney injury complications in children with HSP. These studies identified biomarkers that can distinguish between HSPN and HSP without nephritis (HSPWN). However, biomarkers from a small volume of serum in younger patients may be considered invasive. Also, both studies failed to classify HSPWN patients into specific subtypes, which is necessary for a thorough investigation of metabolic changes as kidney injury progresses in different subtypes. Consequently, the accuracy of conclusions about predicting kidney damage in children with specific HSPWN subtypes may be affected.
Urine, being a non - invasive and easily accessible clinical sample, provides valuable insights into disease progression. Alterations in human proteins, especially urinary proteins, are recognized as indicators of pathophysiological changes caused by various diseases. 23 To reduce the interference from differences in the urinary proteome among different HSP subtypes, the abdominal - type HSP, which has a high incidence and affects the short - term prognosis of the disease, was chosen as the study object. This study aims to identify and verify urine markers for abdominal - type HSP and its progression to kidney injury using label - free liquid chromatography - tandem mass spectrometry (LC - MS/MS) and parallel reaction monitoring (PRM) technology.
Materials and methods
Ethics and human subjects
The study was conducted in compliance with the Declaration of Helsinki (as revised in 2013) and received approval from the local ethics committee of Beijing Children's Hospital, Capital Medical University (No. 2020-k-261). Informed consent was obtained from all participants. Informed consent forms for participants under 18 years of age were co-signed by the participants and their guardians. The study cohort consisted of children affected by abdominal-type HSP, with or without kidney injury, and was enrolled from the Traditional Chinese Medicine inpatient department of Beijing Children's Hospital between June 2021 and January 2022. Age- and sex- matched healthy children were recruited from volunteers among the hospital's staff members. Eligible subjects ranged in age from 1 to 18 years. Abdominal-type HSP was diagnosed based on the European League Against Rheumatism/Paediatric Rheumatology International Trials Organisation/Paediatric Rheumatology European Society (EULAR/PRINTO/PRES) criteria 24 : non-thrombocytopenia palpable purpura combined with diffuse abdominal pain. Kidney injury was diagnosed based on the presence of proteinuria (protein concentration >0.3 g/24 h) and/or hematuria (>5 red blood cells per high-power field or ≥2 + on a urinary dipstick test). The exclusion criteria for the study were as follows: (1) participants who could also be diagnosed with joint-type HSP; (2) participants with other forms of vasculitis; (3) participants with other types of rheumatic immune diseases or primary renal conditions.
Sample collection
In the current study, morning urine samples (midstream) from the cases were meticulously collected by attending physicians between 7:00 and 9:00 a.m. on the day of hospital admission, prior to the initiation of any treatment. Concurrently, early-morning spot urine samples from healthy volunteers were gathered during the same timeframe, with collection facilitated by their guardians. Uniformity was maintained across all sample collections by using identical containers of the same brand and type. Subsequent processing of the samples was conducted uniformly in a single laboratory, employing consistent procedures and equipment. For each child, approximately 10 milliliters of midstream, first-void urine was obtained. The urine samples underwent immediate centrifugation at 3000×g for 30 min at a temperature of 4°C to eliminate cellular debris, followed by storage at a temperature of −80 °C.
Sample preparation
Urine samples were first centrifuged at 12,000 × g for 30 min at 4 °C for urinary protein extraction. Then, 4 mL of urine from each sample was reduced with 20 mmol/L dithiothreitol (DTT, Sigma-Aldrich, Cat # D0632) at 95 °C for 10 min and then alkylated with 50 mmol/L iodoacetamide (IAA, Sigma-Aldrich, Cat # I6125) for 45 min in the dark according to the filter-aided sample preparation (FASP) method. 24 After being precipitated with six volumes of ethanol at −20 °C overnight, the pellets were resuspended in lysis buffer (8 mol/L urea (Sigma-Aldrich, Cat # U1250), 2 mol/L thiourea (Sigma-Aldrich, Cat # 88810), 50 mmol/L Tris (Sigma-Aldrich, Cat # 648311), and 25 mmol/L DTT) and measured using the Bradford method.
For protein digestion, a total of 100 μg of protein from each sample was loaded onto a 10 kDa filter device (Pall, Port Washington, NY, USA). After being washed two times with urea buffer (UA, 8 mol/L urea, 0.1 mol/L Tris-HCl, pH 8.5) and 25 mmol/L NH4HCO3 (Sigma-Aldrich, Cat#A6141) solutions, these samples were digested with trypsin (Promega, Cat # v5280) at an enzyme-to-protein ratio of 1:50 at 37 °C for 14 h. The digested peptides were desalted using Oasis HLB cartridges (Waters, Milford, MA, USA) and dried by vacuum evaporation (Thermo Fisher Scientific, Bremen, Germany).
DIA-LC–MS/MS analysis
The indexed retention time (iRT) peptide standards (Biognosys, Switzerland) was added at a ratio of 1:20 v/v to all peptide samples to calibrate the retention time of the extracted peptide peaks. Peptides were separated using the Ultimate 3000 LC-MS/MS system (DIONEX UltiMate 3000 RSLCnano System, Thermo Fisher ScientificTM, San Jose, USA) equipped with a 500 mm × 75 μm fused silica column custom packed with 2 μm 100 Å C18 aqua. Peptides were separated at a flow rate of 1.5μL/min using a 5%–30% linear gradient of buffer B (buffer A: 0.1% formic acid; buffer B: 80% acetonitrile, 0.1% formic acid). The peptides eluted from the analytical columns were ionized at a potential of +2.4 kV and analyzed using the Orbitrap Exploris 480 mass spectrometer (Thermo Fisher ScientificTM, San Jose, USA).
The variable isolation window of DIA method was generated with pooled samples containing all the peptides using mass spectrometry in DDA mode. The LC settings were consistent for both the DDA-MS and DIA-MS modes to ensure the stable retention time. The MS data were acquired in high-sensitivity mode. A full MS scan was acquired within a 350–1500 m/z range with the resolution set to 120,000. The MS/MS scan was acquired in Orbitrap with a resolution of 30,000. The HCD collision energy was set to 30%. The AGC target was set to 5e4, and the maximum injection time was 45 ms.
The individual samples were subjected to analysis in DIA-MS mode. The DIA acquisition employed a variable isolation window with 60 windows. A full scan was conducted with a resolution of 120,000, covering an m/z range from 350 to 1200. The subsequent DIA scan was performed at a resolution of 30,000. The normalized AGC target was set at 300%, and the maximum injection time was set to 50 ms.
MS data analysis
Three DDA raw files were initially searched using Proteome Discoverer (version 2.4; Thermo Scientific). SEQUEST HT was employed to search against the Swissprot human database (released in Jun 2022, containing 20,386 sequences). The search parameters allowed for up to two missed cleavage sites in trypsin digestion. The fixed modification was Carbamidomethyl (C), the variable modification was set as oxidation (M). The parent ion mass tolerances were set at 10 ppm, and the fragment ion mass tolerance was set at 0.02 Da. A Q value (FDR) cutoff of 1% was applied at both the precursor and protein levels.
The DIA raw files were imported into Spectronaut Pulsar (v17.4, Biognosys AG) using a library-free direct DIA analysis strategy. The parameters of the pulsar search engine were set as follows: enzyme/cleavage rule was Trypsin/P, fixed modification was carbamidomethylation; variable modification was oxidation and acetylation of the protein N-terminus; a maximum of 2 missed cleavages; FDR for PSM, peptide and protein group was set as 0.01. DIA analysis was performed with parameters set as follows: the precision iRT was calibrated based on local non-linear regression; the Q-value cutoff of precursor and protein was 0.01; protein label-free quantification (LFQ)method was MaxLFQ; cross-run normalization was performed using local normalization strategy to calibrate the systematic variance of the LC-MS performance; protein inference algorithm was IDPicker; quantify MS level was MS1.
Parallel reaction monitoring analysis
The proteins of interest in the samples were verified using the Parallel Reaction Monitoring (PRM) technique, with each sample undergoing analysis in a scheduled manner. A mixed sample analysis served as a quality control (QC) to assess the stability of the instrument's signal throughout the analysis. The inclusion of iRT standard peptides in each sample allowed for the monitoring of chromatographic retention time stability. To minimize systemic bias, sample groups were analyzed in a randomized sequence for mass spectrometry. Each sample was run on a self-packed C18 reverse-phase capillary LC column (75 μm × 500 mm) with a gradient elution of 5–30% buffer B2 (0.08% formic acid and 80% ACN; flow rate: 1.5 μL/min) over 25 min. An Orbitrap Exploris 480 Mass Spectrometer was employed to analyze the peptides eluted from the liquid chromatography. Mass spectrometry data were collected in high-sensitivity mode with the following settings: PRM mode, full scans at 60,000 resolution and MS/MS scans at 15,000 resolution, rolling collision energy, charge state filtering (including precursors with +2 to +4 charge states), MS/MS scan range of 350–1200 m/z, and a maximum injection time of 20 ms. The Skyline 3.6 software was utilized to process PRM data, with accurate peak selection for all peptides performed manually and then exported. The Progenesis software was used to extract the total ion current (TIC) for the +2 to +5 charge states of each sample. The TIC was applied to normalize peptide abundance and correct for sample loading variations and MS signal intensity. Six fragment ions were selected to calculate peptide intensity. The PRM results were then exported for further analysis.
Statistical analysis
Pattern recognition analysis (principal component analysis, PCA; orthogonal partial least squares discriminant analysis, OPLS-DA) was performed using SIMCA 14.0 (Umetrics, Sweden) software.
Proteomic data was analyzed using non-parametric tests, calculated by the Mann-Whitney U test, and the statistical significance was defined as P-value of less than 0.05. The receiver operating characteristic (ROC) curves were plotted using Metaboanalyst software (http://www.metaboanalyst.ca).
Clinical data were statistically analyzed using SPSS 26.0 software. Measurements that conform to a normal distribution are expressed as mean ± standard deviation (
Bioinformatics analysis
GO and KEGG pathway analysis was conducted by the DAVID (https://david.ncifcrf.gov/) software. Protein-protein interaction (PPI) networks were created for these DEPs by Cytoscape software (version 3.6.1) according to the STRING database (http://string-db.org/). Figures were visualized in GraphPad Prism software (v9; San Diego, CA, USA) and the Wu Kong platform (https://www.omicsolution. org/wkomics/main/).
Results
Study design and clinical characteristics of subjects
In the present study, a total of 172 subjects, including 74 abdominal-type HSP (AHSP) patients, 17 abdominal-type HSP with kidney injury (AHSPKI) patients, and 81 healthy children (HC) were initially recruited. Subquently, a two-stage workflow, encompassing both a discovery and a verification stage, was established as follows. (1) In the discovery stage, subjects were used to identify differentially expressed proteins (DEPs) between groups and explore the possible biological processes and disease pathways by using data-independent acquisition (DIA) quantitative proteomics strategy. (2) In the verification stage, subjects were used to verify the DEPs as biomarkers and construct protein panels by the PRM method. Given the limited number of AHSPKI patients enrolled in this study, we aimed to maximize the utilization of the available urine samples. Consequently, the collected samples from all 74 AHSP and 17 AHSPKI cases were strategically deployed in both the discovery and verification stages. Subsequently, to ensure a robust control group, we conducted two separate random selections, each time choosing 50 individuals from 89 healthy children. These selected individuals were then utilized as controls across in the two exprimental stages, respectively. 25
By comparing the clinical data, we observed that there were 42 males and 32 females with a mean age of 104.11 ± 36.74 months among the AHSP patients. 10 males and 7 females with a mean age of 125.94 ± 27.20 months among the AHSPKI patients. The demographics and clinical characteristics of the three groups were shown in Additional File 1. Table 1 demonstrates the age and sex of the HC, AHSP, and AHSPKI groups and compares the assays inside the ahsp and ahspki groups. As can be seen from the table, AHSPK1 showed statistically significant differences in the clinical parameters of N%, L%, D-D and IgA compared to AHSP, whereas there were no statistically significant differences in the other clinical parameters.
Characteristics of HC, AHSP and AHSPKI groups.
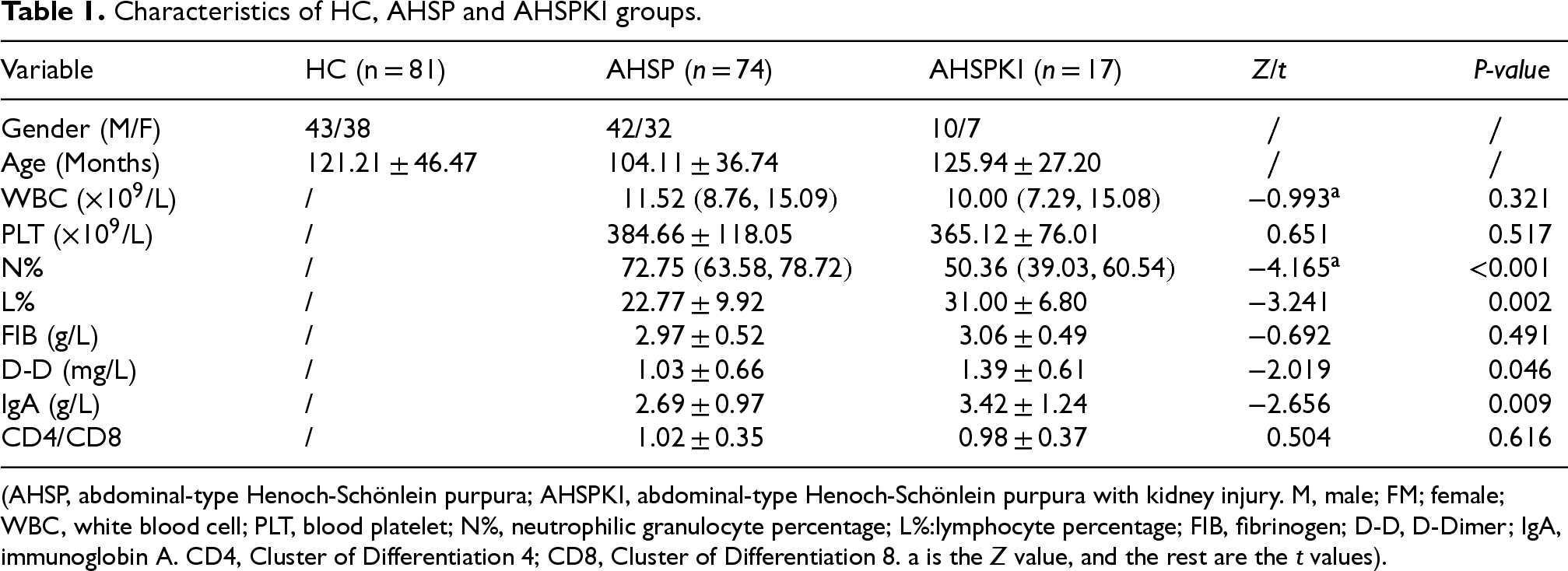
Characteristics of HC, AHSP and AHSPKI groups.
(AHSP, abdominal-type Henoch-Schönlein purpura; AHSPKI, abdominal-type Henoch-Schönlein purpura with kidney injury. M, male; FM; female; WBC, white blood cell; PLT, blood platelet; N%, neutrophilic granulocyte percentage; L%:lymphocyte percentage; FIB, fibrinogen; D-D, D-Dimer; IgA, immunoglobin A. CD4, Cluster of Differentiation 4; CD8, Cluster of Differentiation 8. a is the Z value, and the rest are the t values).
We initiated our study by analyzing the urine proteome of the discovery stage, employing label-free data-independent acquisition liquid chromatography-tandem mass spectrometry (DIA-LC-MS/MS) for quantitation. The average number of identifed proteins in AHSP, AHSPKI, HC, QC-digestion, and QC-DIA groups were 2504, 2328, 2123, 2480, and 2715, respectively (Figure 1A). Further correlation analysis showed that the mean correlation coeffcient of the 9 QC-digestion samples was 0.91 with a median CV value of 0.51 (Figure 1B). Similarily, the 8 QC-DIA samples demonstrated an exceptionally high mean correlation coefficient of 0.99, along with a median CV of 0.32 (Figure 1C). Subsequently, the method of Mann et al. 26 was used to remove samples contaminated by erythrocytes. Samples with relatively low number of identified proteins and those contaminated with erythrocytes, including 3 AHSPKI, 2 AHSP cases and 4 HC subjects, were further excluded (Figure 1D and (E)). To address the issue of missing data, the sequential-KNN method was used to impute proteomic data in there groups separately and remove missing proteins in more than 50% of the samples.
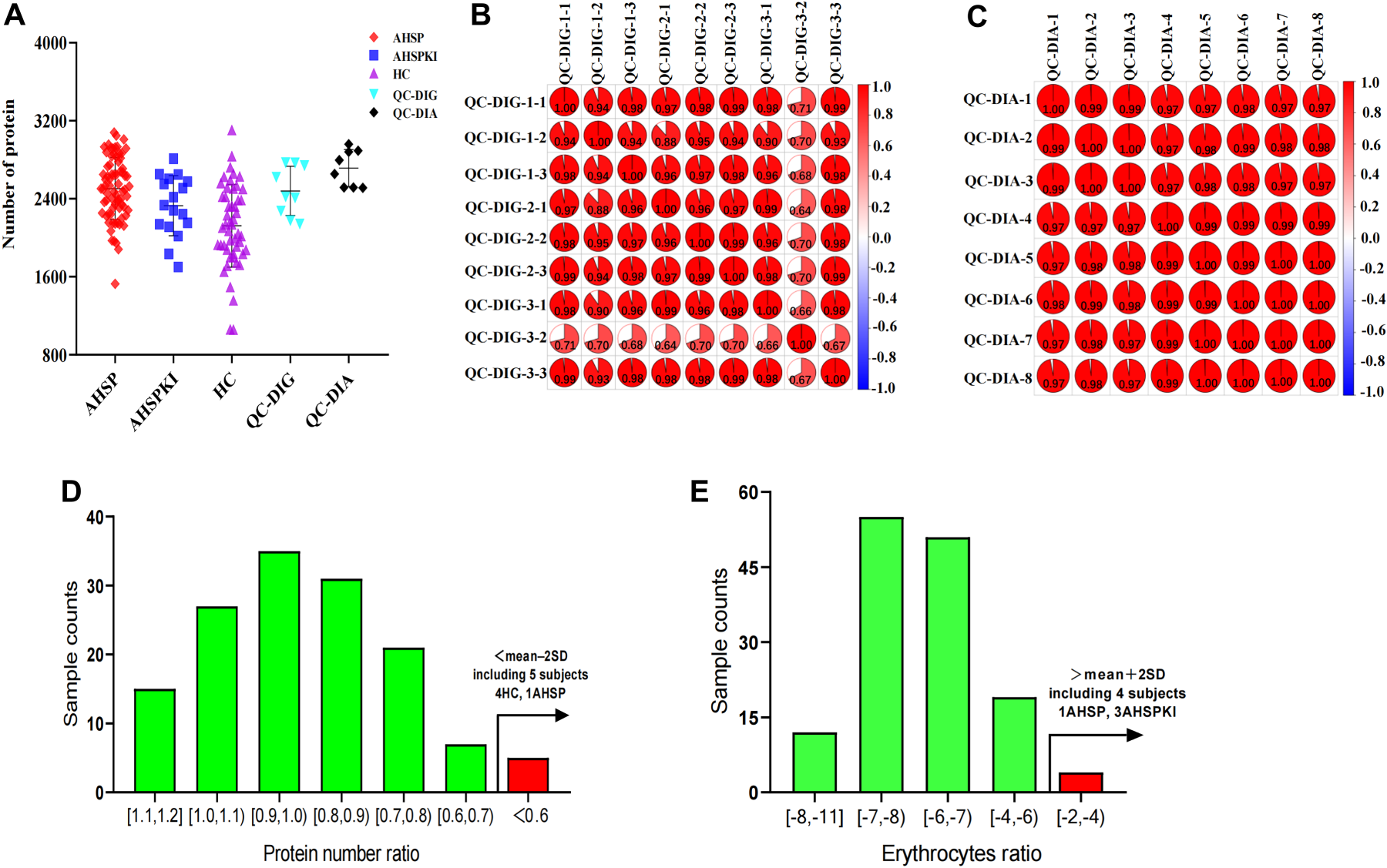
Proteomic characterization of urine samples and quality assessment. (A) The number of identifed proteins in AHSP, AHSPKI, HC, QC-digestion, and QC-DIA groups. (B) The correlation analysis of 9 QC-DIG samples by Pearson correlation coeffcient. (C) The correlation analysis of 8 QC-DIA samples by Pearson correlation coeffcient. (D) Analysis of urine samples with relatively low number of identified proteins. The x-axis represents the ratio of protein number in each sample from the AHSP, AHSPKI, and HC groups to the average protein number in the QC samples, the y-axis represents sample counts. (E) Analysis of urine samples contaminated with erythrocytes. The x-axis represents the log2-transformed ratio of the total quantitative values for erythrocyte contamination proteins to the total quantitative values of all proteins in samples from the AHSP, AHSPKI, and HC groups, the y-axis represents sample counts. AHSP, abdominal-type Henoch-Schönlein purpura; AHSPKI, abdominal-type Henoch-Schönlein purpura with kidney injury; HC, healthy children. QC, quality control; DIG, digestion; DIA, data-independent acquisition.
Initially, to explore the proteomic profiling differences among the three groups, unsupervised PCA analysis was performed. The score plot showed that half of healthy samples in HC group were clustered in a region, and were separated from the two disease groups, while separation trend could also been seen between AHSPKI and AHSP group (Figure 2A). Subquently, OPLS-DA was performed to further determine the proteomic differences among groups. The HC, AHSP and AHSPKI groups could be obviously separated from each other in the OPLS-DA model (Figure 2B). One hundred permutation tests indicated that there was no over-fitting in these models.
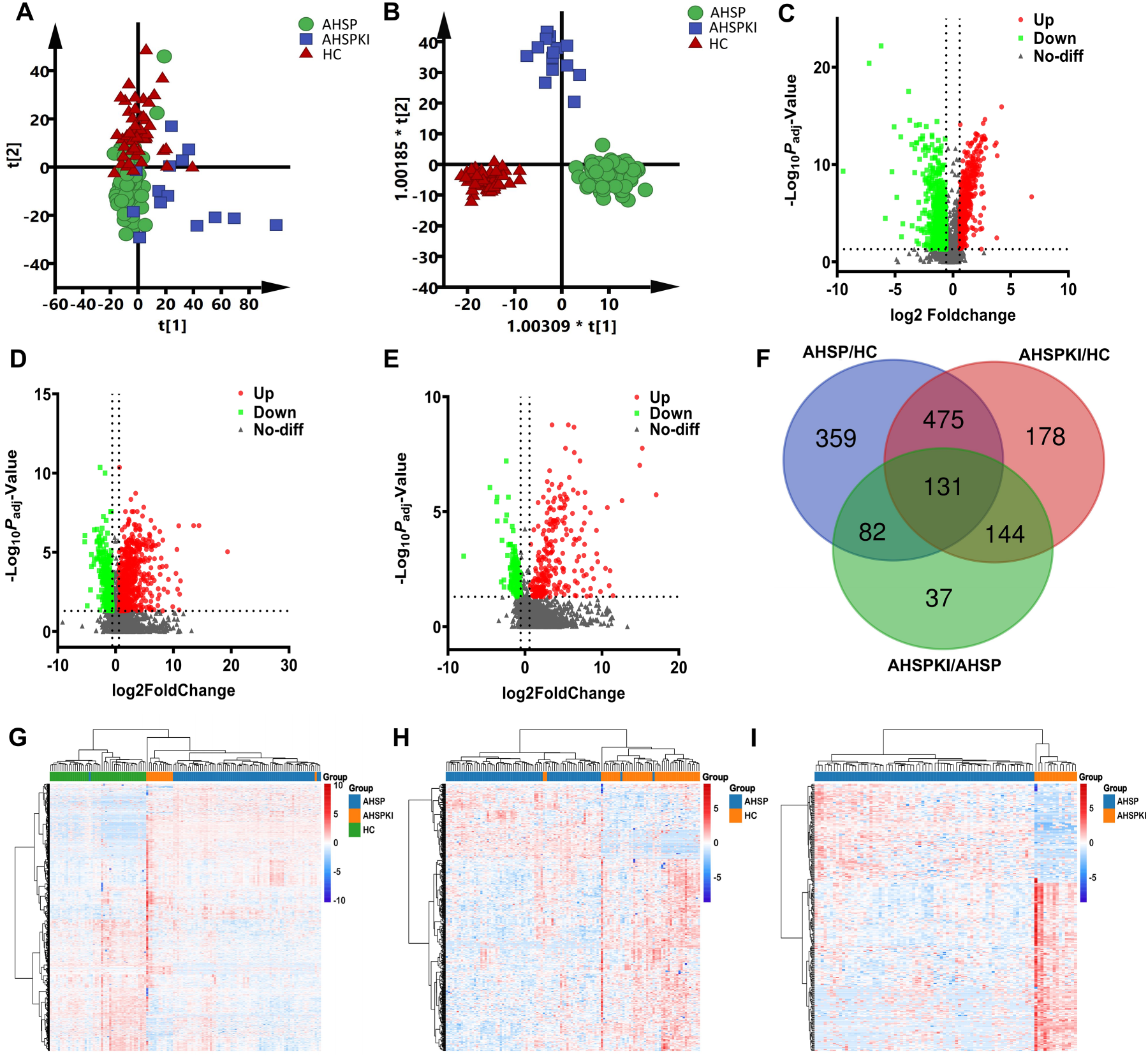
Overview of the urinary proteomes and differentially expressed proteins discovery. (A) Pattern recognition analysis of proteomic profiling differences among groups. (B) Orthogonal partial least squares discriminant analysis of proteomic profiling differences among groups. (C) Volcano plot of differentially expressed proteins between AHSP and HC group. (D) Volcano plot of differentially expressed proteins between AHSPKI and HC group. (E) Volcano plot of differentially expressed proteins between AHSPKI and AHSP group. (F) Venn diagram of differentially expressed proteins between groups. The blue circle represented the 1047 differentially expressed proteins between AHSP and HC group, the red circle represented the 928 differentially expressed proteins between AHSP and HC, the green circle represented the 394 differentially expressed proteins between AHSP and HC. (G) Heatmap of the total 1046 differentially expressed proteins among AHSP, AHSPKI and HC groups. (H) Heatmap of the 441 differentially expressed proteins between AHSP and HC groups. (I) Heatmap of the 275 differentially expressed proteins between AHSPKI and AHSP groups. AHSP, abdominal-type Henoch-Schönlein purpura; AHSPKI, abdominal-type Henoch-Schönlein purpura with kidney injury; HC, healthy children.
To investigate DEPs across groups, the criteria of fold change >1.5 or <0.67 and two-sided adjusted p value <0.05 was employed. A total of 1047 DEPs (445 upregulated and 602 downregulated) were identified between AHSP and HC group, 928 DEPs (619 upregulated and 309 downregulated) were identified between AHSP and HC group, and 394 DEPs (261 upregulated and 133 downregulated) were identifed between AHSPKI and AHSP group. The volcano plots of DEPs when different groups compared were plotted in Figure 2C, D and E, respectively. Furture comparison of the above 1047, 928 and 394 DEPs indicated that total 1406 non-overlapped DEPs were identified by Venn diagram
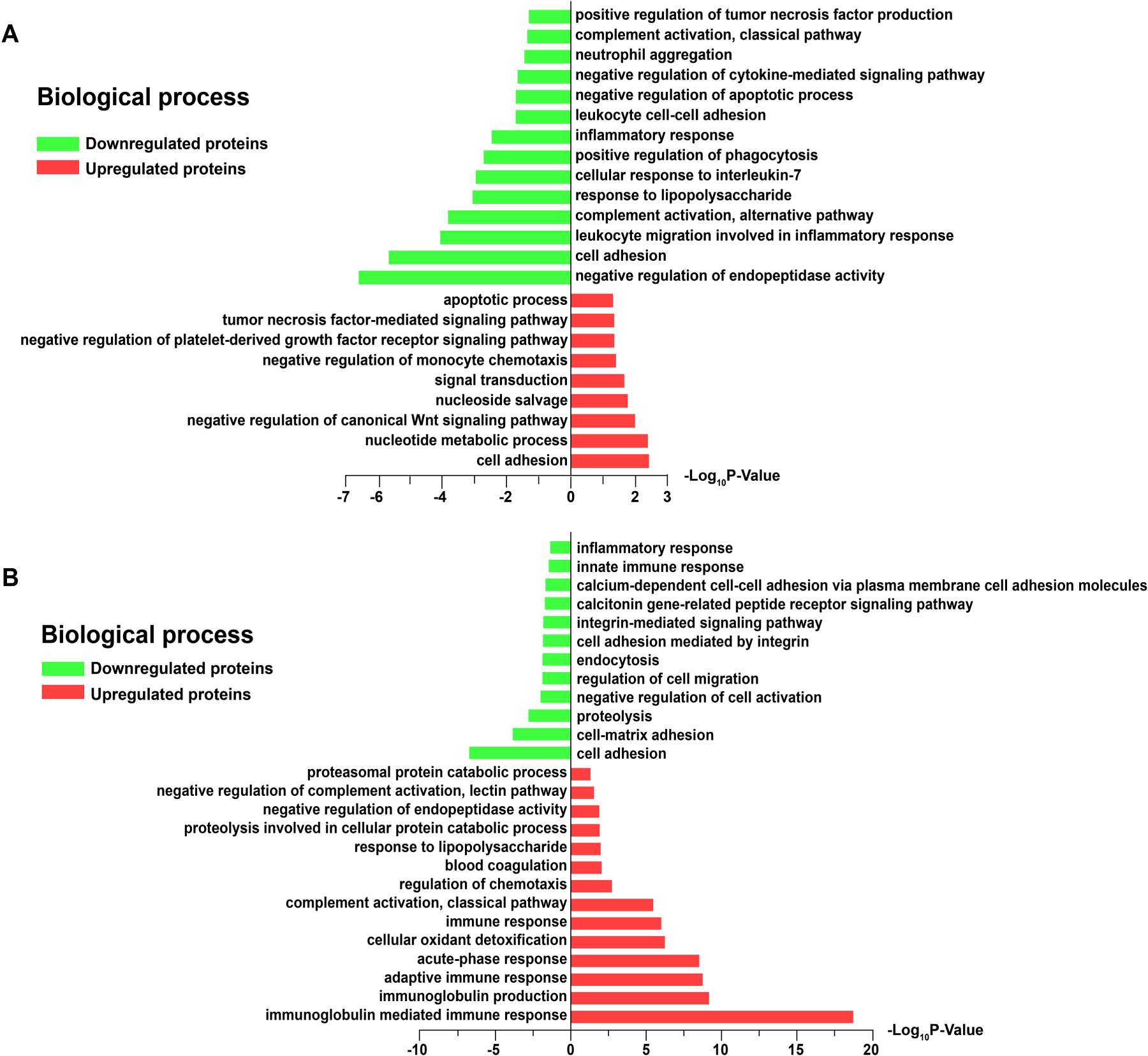
Go enriched analysis of the differentially expressed proteins. (A) Main biological process terms for the 441 differentially expressed proteins between the AHSP and HC groups. (B) Main biological process terms for the 275 differentially expressed proteins between the AHSPKI and AHSP groups.
GO functional enrichment analysis
To further investigate the biological functions of the DEPs identified, GO functional enrichment analysis was performed. Initially, we conducted a GO analysis to enrich and cluster the total of 441 (359 plus 89, 114 upregulated, 327 upregulated) DEPs between AHSP group and HC group. Notably, these 441 DEPs were not overlapped by the DEPs marked within the red circle during the comparison between the AHSPKI group and the HC group (Figure 2F), which aids in elucidating the intrinsic functional characteristics of AHSP itself. The GO annotation analysis of biological processes revealed that the 114 upregulated DEPs were predominantly engaged in processes such as cell adhesion, nucleotide metabolic processes, and multiple factor-mediated signaling pathways. Meanwhile, the 327 downregulated DEPs were largely associated with negative regulation, inflammatory responses, complement activation, the alternative pathway, and responses to lipopolysaccharide
Subquently, we selected another 275 (131 plus 144, 179 upregulated, 96 upregulated) DEPs overlapped between red and green circles to clarify functional characterization of kindey injury complication in AHSP (Figure 2F). The GO annotation enriched analysis of biological processes revealed that the 179 upregulated DEPs were mainly involved in immune or acute-phase response, negative regulation of complement activation, the lectin pathway, and the proteasomal protein catabolic process. In contrast, the 96 downregulated DEPs were involved in cell adhesion, proteolysis, negative regulation of cell activation, endocytosis, and inflammatory response
KEGG pathways enrichment analysis
To explore the potential biological pathways of the 441 DEPs between the AHSP and HC groups, and the 275 DEPs between the AHSPKI and AHSP groups, we performed KEGG pathway analysis. It was observed that the 114 upregulated DEPs between the AHSP and HC groups were mainly associated with five pathways: lysosome, metabolic pathways, the TGF-beta signaling pathway, the intestinal immune network for IgA production, and arginine and proline metabolism. Meanwhile, the 327 downregulated DEPs were found to be significantly enriched in another ten pathways, including neutrophil extracellular trap formation, systemic lupus erythematosus, complement and coagulation cascades, staphylococcus aureus infection, necroptosis, cholesterol metabolism, ATP-dependent chromatin remodeling, Fc gamma R-mediated phagocytosis, focal adhesion, and Rap1 signaling pathway (Figure 4A). Subquently, KEGG pathway analysis was conducted on the 275 DEPs between AHSPKI and AHSP groups. It was observed that the 179 upregulated DEPs were mainly associated with seven pathways: the regulation of actin cytoskeleton, Fc gamma R-mediated phagocytosis, bacterial invasion of epithelial cells, the complement and coagulation cascades, the HIF-1 signaling pathway, tight junction, and neutrophil extracellular trap formation. At the same time, the 96 downregulated DEPs were found to be enriched in another six pathways, including cell adhesion molecules, the PI3K-Akt signaling pathway, ECM-receptor interaction, focal adhesion, human papillomavirus infection, and neuroactive ligand-receptor interaction (Figure 4B).
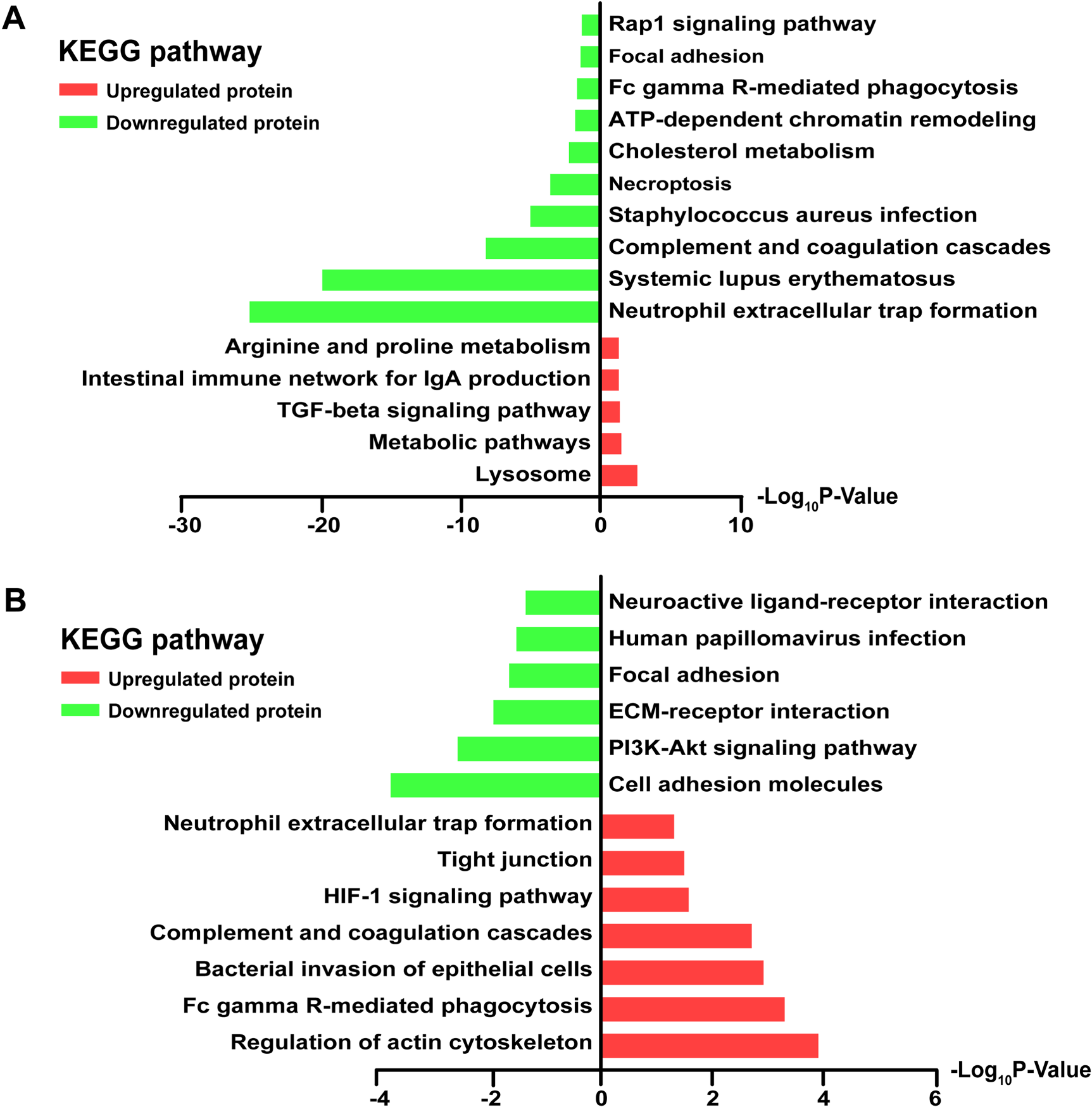
KEGG pathways analysis of differentially expressed proteins. (A) Main KEGG pathways of the 441 differentially expressed proteins between AHSP and HC groups. (B) Main KEGG pathways of the 275 differentially expressed proteins between AHSPKI and AHSP groups.
Based on these findings, pathways such as immune response, phagocytosis, infection, various signaling pathways and receptor interactions may be involved in the biological pathways of AHSP and its kidney injury complication.
Disease occurrence and progression are closely associated with protein expression levels. To select specific proteins and reveal changes in protein abundance among the HC, AHSP, and AHSPKI groups, we applied methods from two previous studies27,28 to cluster the total 1406 identified DEPs from all three groups into ten significant discrete clusters with quantified values using mFuzz
29
(Figure S1). Among these clusters, clusters 4, 7, and 10 exhibited an upregulation trend as one panel
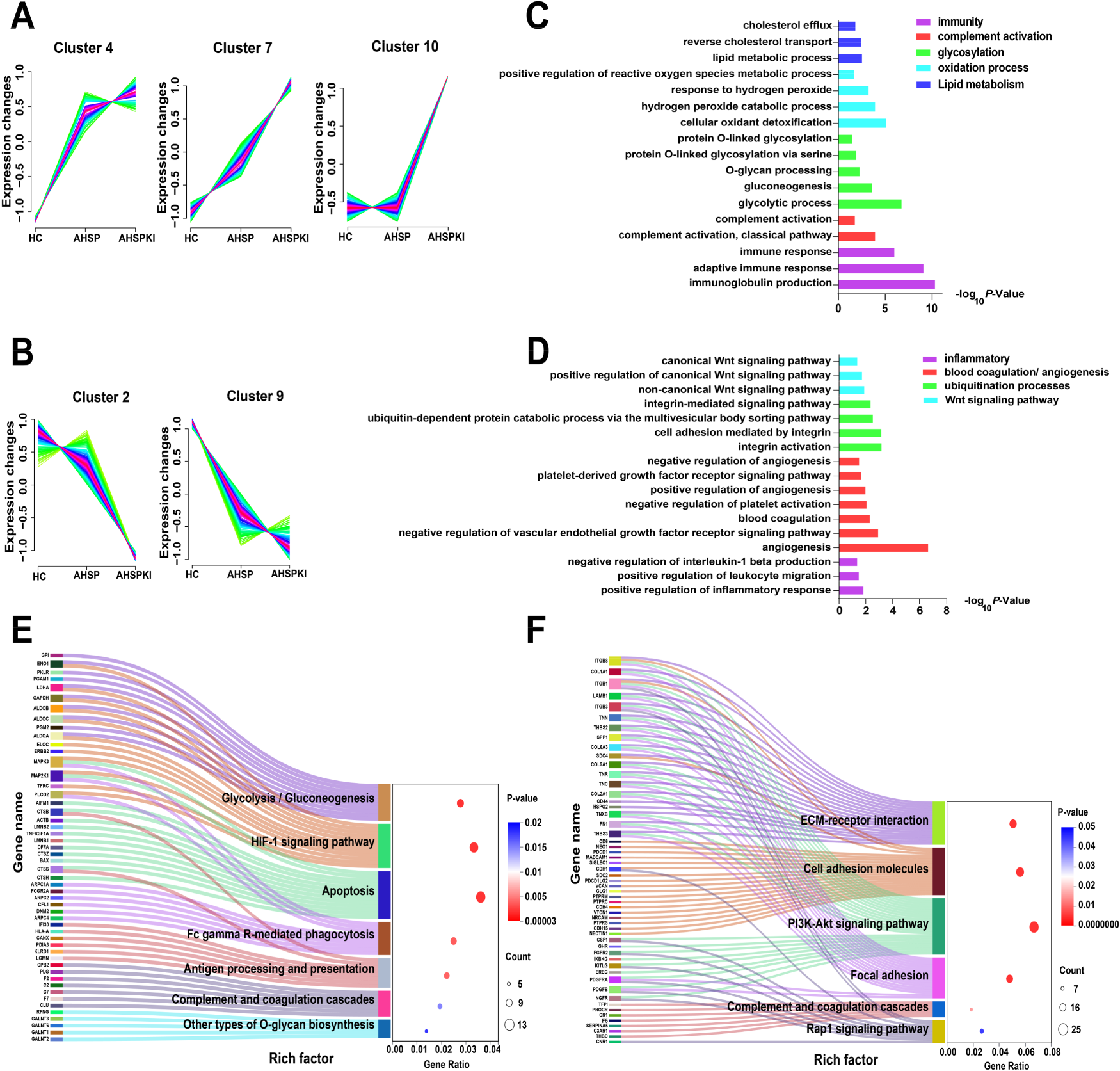
Clustering of the 1406 commonly identified differentially expressed proteins illustrated specific clusters of proteins in HC, AHSP and AHSPKI groups. (A) The cluster 4, 7 and 10 stands for the up-regulated trends uniquely during kidney injury progression in AHSP. (B) The cluster 2 and 9 stands for the down-regulated trends uniquely during kidney injury progression in AHSP. (C) The biological process analysis of the filtered proteins from panel A. (D) The biological process analysis of the filtered proteins from panel B. (E) The biological pathway analysis for filtered proteins from panel A. (F) The pathway analysis for filtered proteins from panel B. AHSP, abdominal-type Henoch-Schönlein purpura; AHSPKI, abdominal-type Henoch-Schönlein purpura with kidney injury; HC, healthy children.
It is worth mentioning that recent reports on three urine proteomics studies of HSPN30–32 align with the identification of key biological pathways, such as focal adhesion, cell adhesion molecules, the PI3K-Akt signaling pathway, ECM receptor interactions, and complement and coagulation cascades. Building on this, we first determined which proteins were mapped to these reported pathways and then carried out a protein-protein interaction enrichment analysis using the STRING database. Meanwhile, the Molecular Complex Detection (MCODE) algorithm was applied to identify densely connected network components. A total of 60 proteins contributed to the five key biological pathways mentioned above (Figure 6A). Protein network analysis showed that 47 proteins formed 2 independent interactions, and 3 MCODE networks were identified (Figure 6B). The first MCODE network consisted of a smaller interaction involving three proteins (VTCN1, PDCD1LG2, and PDCD1), which are highly correlated with immune response. The second MCODE network included six proteins: CPB2, PROCR, TFP1, THBD, F7, and F2. The third network contained nine proteins: COL1A1, COL6A3, COL2A1, ITGB8, ITGB1, ITGB3, THNS2, SPP1, and FN1.
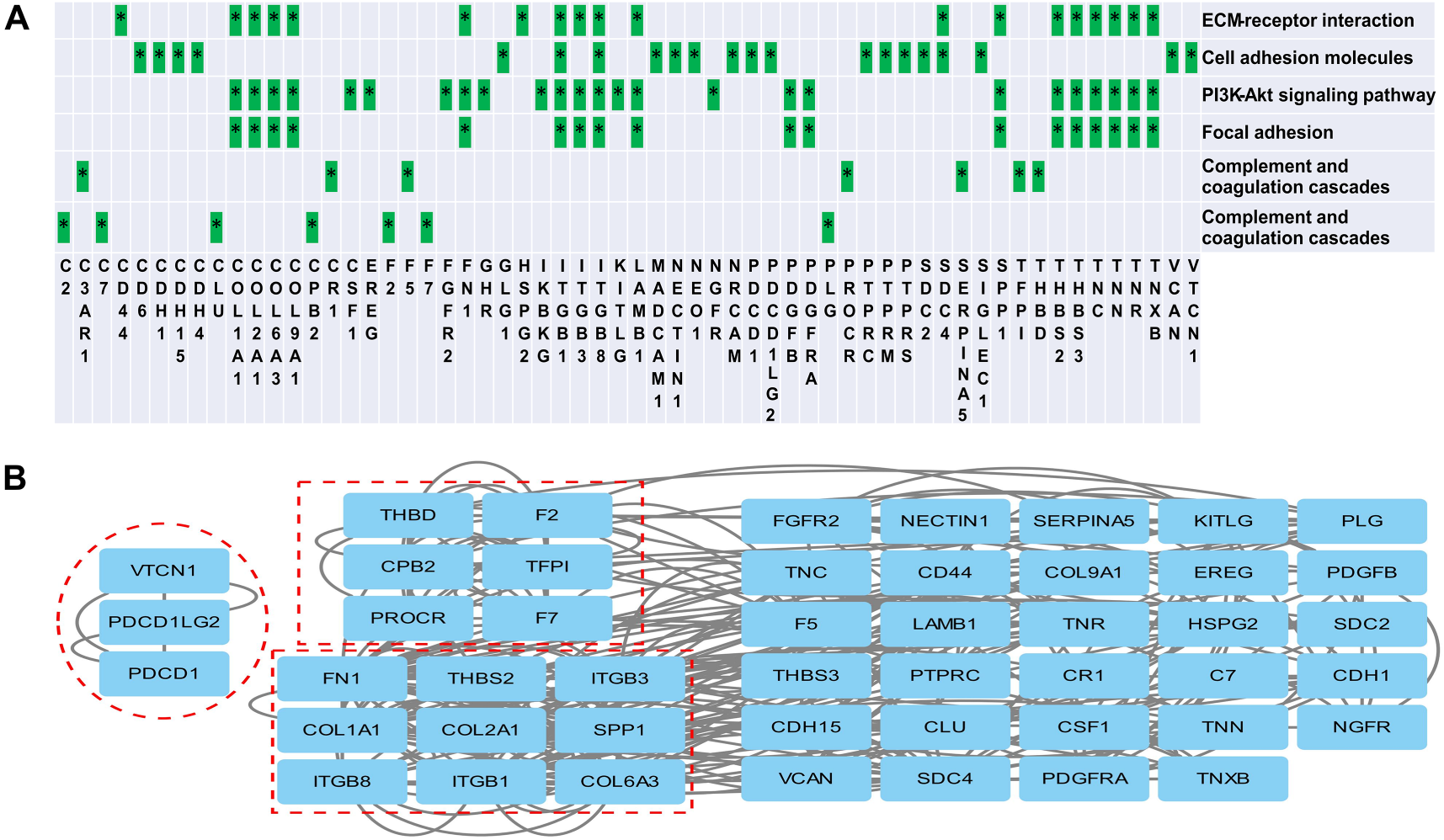
Protein network analysis. (A) Proteins mapped to reported pathways. (B) Network analysis of proteins involved in reported pathway. Three MOCDE networks are outlined by dashed boxes. MOCDE, Molecular Complex Detection.
Given the extensive list of 1406 DEPs identified through DIA, it is not feasible to validate all of them for inter-group comparisons. To address this, we implemented a rigorous selection process to determine an appropriate subset of candidate proteins for PRM analysis. This process involved a stringent screening of DEPs between groups identified during the discovery stage. Initially, we selected DEPs with area under the curve (AUC) values exceeding 0.85, and fold change (FC) values greater than 2.0 or less than 0.5 upon inter-group comparison. Additionally, we ensured that the missing values in proteomic data kept relatively low within each of the two groups being compared.
Through this stringent screening, we successfully narrowed down the initial set of 1406 DEPs across three groups to a refined list of 77 candidate proteins for PRM validation. Of these, 27 proteins remained when comparing the AHSP group to the HC group, 48 proteins when comparing the AHSPKI group to the HC group, and 25 proteins when comparing the AHSPKI group to the AHSP group. The details of AUC, FC, and missing values for these 77 candidate proteins can be found in Additional File 2.
Verification of protein biomarkers based PRM
To maintain system stability and minimize systemic bias during proteomic analysis, a pooled quality control (QC) sample was prepared from all urine samples in the validation stage. Correlation analysis revealed a mean correlation coefficient of 0.98 across 10 QC samples, with a median coefficient of variation (CV) of 0.42, indicative of the stability of the mass spectrometry (MS) platform throughout the analysis.
During the validation stage, a total of 144 peptides corresponding to 67 proteins were identified. Utilizing the same data processing methodology as the discovery stage, we excluded 2 peptide data due to excessive missing values, which were greater than 50% across all three groups. The remaining 142 peptide data were compared between different groups. In total, 87 peptides corresponding to 45 DEPs exhibited consistent changes across both experimental stages when different groups were compared, which included 9 DEPs between the AHSP and HC groups, 32 DEPs between the AHSPKI and HC groups, and 17 DEPs between the AHSPKI and AHSP groups. These consistently changing DEPs can serve as reliable predictive biomarkers of AHSP and its kidney damage complication and also construct protein panels for their clinical diagnosis. Detailed information for the 45 DEPs is presented in Additional File 3.
Evaluation of the protein biomarkers based on ROC analysis
To further evaluate the diagnostic performance of the consistent biomarkers, the 9, 32, and 17 proteins in the veen diagram (Figure 7A) were randomly selected to form the protein panel (compact biomarker combination), and improve diagnostic efficacy between different groups, respectively. The criteria for protein selection for the panel were: (1) the panel should contain no more than 5 constituent proteins; (2) the panel with the largest AUC was selected. Initially, the two disease groups were compared with the HC group, respectively. A panel of 3 urinary proteins (P05090, O14773, P30041) showed excellent sensitivity and specificity for the differentiation of AHSP and the HC group, with an AUC value of 0.942 (Figure 7B). Another panel of 5 proteins, including P00450, P04217, P69905, P98164, and P68871, could significantly distinguish AHSPKI patients from healthy children with an AUC value up to 0.992 (Figure 7C). Subsequently, when the AHSPKI group was compared with the AHSP group, a combination of 5 proteins (P69905, P68871, P02787, P04217, P01859) could differentiate them with an AUC value of 0.919 (Figure 7D).
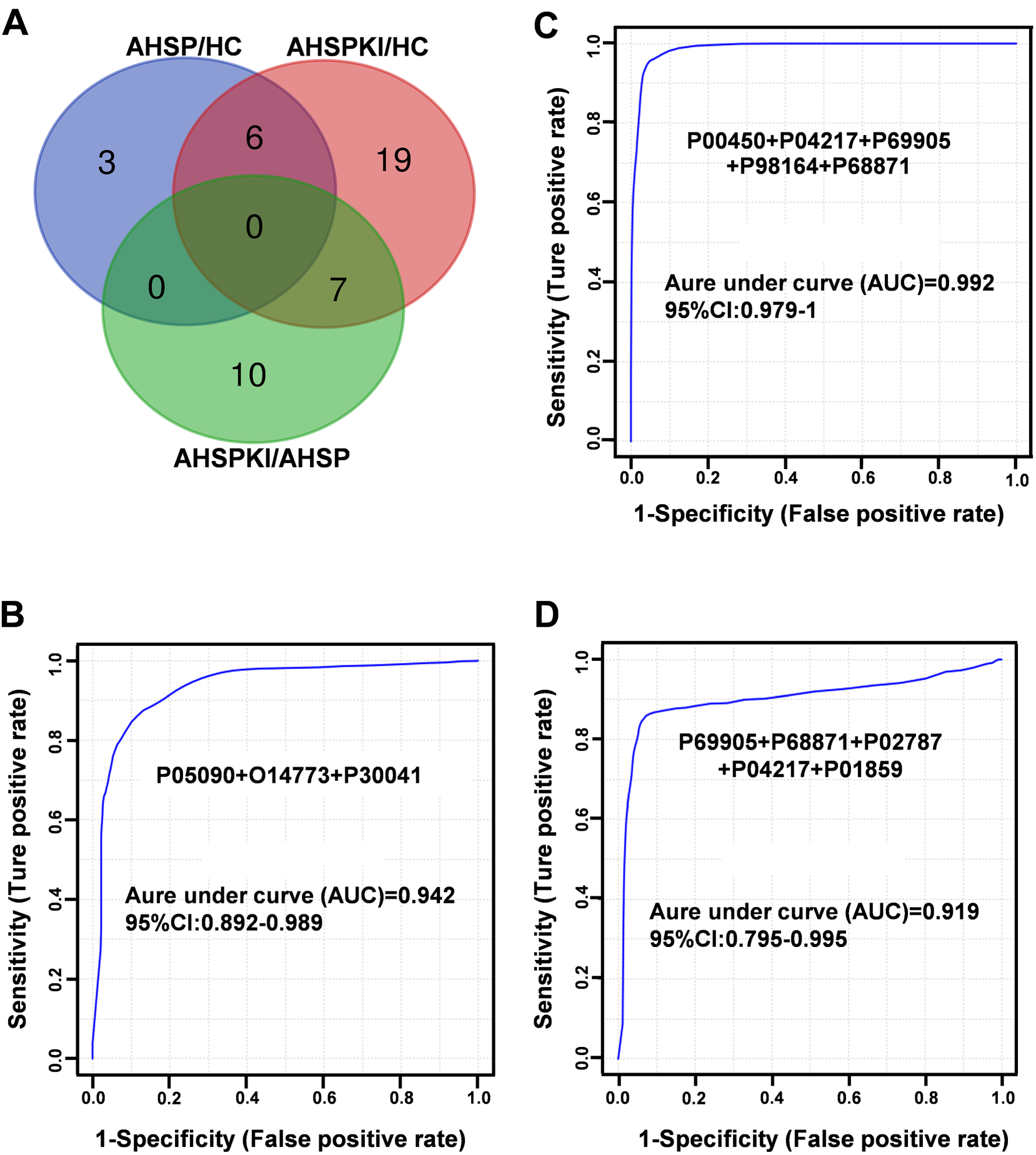
(A) Veen diagram of DEPs with consistent changes in two experiments. (B) Panel for distinguishing AHSP from healthy controls. (C) Panel for distinguishing AHSPKI from healthy controls. (D) Panel for distinguishing AHSPKI and AHSP. The x-axis represents the diagnostic sensitivity of the biomarker panel, and the y-axis represents its diagnostic specificity. P05090: Apolipoprotein D; O14773: Tripeptidyl-peptidase 1; P30041: Peroxiredoxin-6; P00450: Ceruloplasmin; P04217: Alpha-1B-glycoprotein; P69905: Hemoglobin subunit alpha; P98164: Low-density lipoprotein receptor-related protein 2; P68871: Hemoglobin subunit beta; P02787: Serotransferrin; P01859: Immunoglobulin heavy constant gamma 2. AHSP, abdominal-type Henoch-Schönlein purpura; AHSPKI, abdominal-type Henoch-Schönlein purpura with kidney injury; HC, healthy children.
These results indicated that a minimum of 8 urinary proteins (P05090, O14773, P30041, P00450, P04217, P69905, P98164, P68871, P02787, P01859) made contributions to the diagnosis of AHSP or its kidney injury complication with good performance, which is worth forming into a urinary protein panel for further investigation.
AHSP is the most common form of HSP and has a significant impact on the short-term prognosis of affected children. Despite its prognostic implications, the early clinical manifestations in these children are frequently subtle or atypical, which can lead to diagnostic and treatment delays, further affecting the disease's outcome. In our previous study, 33 we adopted the principles of traditional Chinese medicine and collected urine samples from a diverse group of children with AHSP. Utilizing quantitative proteomics, we analyzed and compared the urinary proteome of these patients to that of healthy controls with the goal of identifying whether the urinary proteome profile could differentiate AHSP patients and their respective syndromes. Through this process, we successfully identified differential urinary proteins that have potential as diagnostic and phenotypic markers for AHSP. Initially, our research was primarily focused on the preliminary exploration of urine as a non-invasive biomarker, aiming to support the traditional Chinese medicine approach of ‘diagnosing patterns to identify diseases.’ Consequently, we did not deeply investigate the biological implications of the differential proteins observed between children with AHSP and the healthy control group.
We have recognized that children with AHSP who do not receive a definitive diagnosis and standardized treatment are at a significantly increased risk of kidney injury complications if their abdominal symptoms intensify or worsen. Severe abdominal pain, gastrointestinal bleeding, or persistent symptoms can all increase the likelihood of kidney damage, which in turn affects the long-term prognosis of the disease. Therefore, in contrast to previous studies, in the current research, we initially recruited a larger number of children with AHSP. Building on this, we also included a subset of those with kidney impairment associated with the condition. The aim was to identify urinary proteomic biomarkers for AHSP and its kidney damage, and to provide a detailed explanation of the potential biological processes and pathways they may correspond to. Additionally, we employed PRM to validate the differential proteins between groups, offering assistance in the differential diagnosis of the disease and its renal complications.
During the discovery phase, we identified 1047 DEPs between the AHSP and HC groups, 928 DEPs between the AHSP and HC groups, and 394 DEPs between the AHSPKI and AHSP groups. To perform a biological process and pathway analysis of the DEPs unique to AHSP and its kidney injury complications, we selected 441 DEPs from the comparison between the AHSP group and the HC group (114 upregulated and 327 downregulated), as well as 275 DEPs from the comparison between the AHSPKI group and the AHSP group (179 upregulated and 96 downregulated). This study found that these upregulated or downregulated DEPs are significantly enriched across diverse biological processes and are mapped onto several pivotal pathways. The 441 dysregulated DEPs can represent the pathogenesis of AHSP, mainly related to cell adhesion, signal transduction or regulation, and reactions or pathways mediated by inflammatory cells or factors, such as neutrophils, leukocytes, phagocytes, tumor necrosis factor, interleukins, etc. As a small vessel vasculitis, HSP is associated with an abnormal inflammatory process caused by various antigenic stimuli, which may include bacteria, viruses, or other pathogens. 34 Multiple studies have found that adhesion molecules play a crucial role in the immune-inflammatory response of damaged vascular tissues.35,36 Wang et al. believe that soluble intercellular adhesion molecule-1 (sICAM-1) and vascular cell adhesion molecule-1 (sVCAM-1) can serve as diagnostic markers for HSP. 37 Gattorno et al. suggest that tumor necrosis factor (TNF) and its induced adhesion molecules provide evidence for the pathogenesis of tissue damage in pediatric HSP. 38 In addition, recent proteomic studies have also confirmed the involvement of cell adhesion in the pathogenesis of HSP.30,31 When analyzing 275 dysregulated DEPs related to the pathogenesis of renal complications in HSP, it was found that they are primarily associated with immune processes mediated by immunoglobulins. Furthermore, biological processes such as complement activation via the lectin pathway, and integrin-mediated signaling pathways are also implicated in the occurrence of renal damage complications. Research has found that the activation of the complement system through the alternative or lectin pathways plays a key role in the pathogenesis of HSPN and the associated histological damage.39,40 Fang et al. 31 pointed out that integrins may play a role in pathological changes in the glomeruli, including damage to the glomerular basement membrane and loss of podocytes, which are characteristic pathological changes of HSPN.
Further analysis of the biological pathways of differential proteins during the onset of AHSP showed that upregulated proteins were mainly enriched in the lysosomal pathway, which has been frequently reported in the pathogenesis of systemic lupus erythematosus and lupus nephritis. 41 Hydroxychloroquine, which interferes with lysosomal activity and antigen presentation, inhibits autophagy, and reduces the transcription of pro-inflammatory cytokines, has been selected as an important drug for the treatment of systemic lupus erythematosus and lupus nephritis due to its immunomodulatory, anti-inflammatory, and antithrombotic effects.42,43 A study conducted well before this pointed out that the characteristic clinical manifestation of palpable purpura in HSP may be related to the release of lysosomal enzymes from cells, leading to the destruction of blood vessel walls and leakage of red blood cells into the tissues. 44 In addition, recent research has also indicated that the pathogenesis of HSP may be related to abnormalities in anti-lysosomal membrane proteins.45,46 This alerts us to the possibility that the lysosomal pathway is involved in the pathogenesis of HSP and deserves further in-depth research and exploration. Interestingly, the dysregulated differential proteins were also enriched in pathways such as the intestinal immune network for IgA production and systemic lupus erythematosus, which have been proven to be significantly associated with systemic vasculitis. 47 When analyzing the biological pathways of differential proteins in the complications of AHSP kidney damage, it was found that the dysregulated proteins were mainly enriched in the regulatory pathways of the actin cytoskeleton, which is consistent with recent urinary proteomics studies related to HSPN.31,48 Research has found that the actin cytoskeleton is an important structural and functional element that controls the activity of podocytes and is closely related to kidney damage.
A time series clustering analysis was conducted on 1406 DEPs using mFuzz, revealing 10 significant discrete clusters with quantified values. From these clusters, three clusters showing an upregulation trend and two clusters showing a downregulation trend were selected to form their respective panels. The biological processes involved by DEPs in these panels not only include widely recognized immune and inflammatory processes related to the occurrence and progression of diseases but also include processes such as lipid metabolism, glycosylation, coagulation, oxidative detoxification processes, and the Wnt signaling pathway, which have been reported in previous studies.30,31,48 It is known that lipid metabolism disorders can exacerbate chronic kidney disease, 49 and although there is no direct evidence to suggest that lipid metabolism is related to HSPN, the fact that HSPN is a multifactorial disease indicates that lipid metabolism disorders may be involved in the pathogenesis of the disease. The Wnt signaling pathway is known to regulate various biological processes, including proliferation, migration, invasion, and apoptosis. Extensive research has confirmed that Wnt signaling transduction can continuously exacerbate podocyte damage and proteinuria, leading to the involvement of other pathways in mediating podocyte disease.50,51
Further analysis of the differential proteins corresponding to enriched pathways for interaction network analysis revealed three distinct MCODE networks. The first network consists of three proteins (VTCN1, PDCD1LG2, and PDCD1), where VTCN1 is a T cell regulatory factor of the immunoglobulin superfamily, proven to be highly associated with the occurrence of autoimmune diseases. 52 PDCD1LG2 is related to the induction of immune tolerance under various conditions, 53 and PDCD1 is an important immune checkpoint molecule on activated T cells, playing a significant and diverse role in immune regulation in T cell activation, tolerance, and immune-mediated tissue injury, and has been found to play an important role in the development of postoperative renal injury. 54 Although no studies have directly implicated these proteins in the pathogenesis of HSP and its renal complications, their participation in immune-related functions warrants further investigation. The second MCODE network includes six proteins (CPB2, PROCR, TFP1, THBD, F7, and F2) involved in coagulation. CPB2 is associated with coagulation, thrombosis, inflammation, and innate immunity. 55 PROCR, as an important regulatory factor in the coagulation system of HSP patients, may significantly affect the prognosis of severe cases by causing gastrointestinal bleeding or kidney damage. 56 TFP1, as a marker of endothelial dysfunction, has been proven to be related to thrombosis and bleeding in patients with kidney damage. 57 The protein interaction network suggests an interaction between coagulation-related proteins and the complement system, emphasizing the need for further research into the imbalances and intersections between these systems, consistent with the findings of another proteomics study. 41 The third MCODE network features nine proteins, mainly collagen and integrin proteins. Collagen, a key structural component of the extracellular matrix (ECM), is involved in processes such as cell proliferation, migration, and differentiation, with ECM-receptor interactions implicated in the pathological processes associated with HSP kidney injury.31,58 Abnormal ECM remodeling, a common pathway in glomerular injury and a hallmark of many glomerular diseases, is highlighted. Integrins, cell surface proteins that mediate interactions between cells and the ECM, are implicated in cellular processes including proliferation, migration, differentiation, and apoptosis. In kidney diseases, integrins may contribute to the onset and progression of the disease by influencing ECM deposition, changes in cell phenotype, differentiation, migration, and apoptosis.30,59
In the validation stage, we successfully identified at least eight urinary proteins (P05090, O14773, P30041, P00450, P04217, P69905, P98164, P68871, P02787, P01859) that contribute to the diagnosis of AHSP or its kidney injury complications, exhibiting good diagnostic performance, and are worthy of forming a urinary protein panel for further research.
This study demonstrates the value of urinary proteomics in disease identification. We successfully identified at least eight urinary proteins that are useful in the diagnosis of AHSP or its renal injury complications, providing new diagnostic indicators for the early recognition of the disease and its complications, and a new theoretical basis for clinical diagnosis. Meanwhile, our future research direction is more inclined to analyze these eight kinds of urinary proteins that help to diagnose the disease by composite indexes, so as to improve the accuracy and sensitivity of diagnosis.
This study has several limitations: First, like most proteomics studies, our research is cross-sectional and does not indicate that individuals without renal damage will not develop renal complications in the short term. Second, due to the relatively small number of individuals with existing renal damage, sample acquisition is challenging. We conducted PRM validation for all case samples in the discovery phase, which in fact represents the use of two mass spectrometry methods. It is necessary to further validate in a larger sample size to improve the accuracy of the analysis and reduce its variability. We did not use biological analysis to assess other glomerular diseases.
Conclusions
A total of 441 dysregulated differentially expressed proteins (DEPs) were associated with the pathogenesis of AHSP, mainly related to cell adhesion, signal transduction or regulation, and reactions or pathways mediated by inflammatory cells or factors, and predominantly enriched in the lysosomal pathway. A total of 275 DEPs related to renal complications of AHSP were mainly associated with immune processes mediated by immunoglobulins, predominantly enriched in the regulatory pathways of the actin cytoskeleton.
Time series clustering analysis identified 10 discrete clusters; three upregulated and two downregulated clusters were chosen to form respective panels. These panels involved various biological processes such as immune and inflammatory processes, lipid metabolism, glycosylation, coagulation, oxidative detoxification processes, and the Wnt signaling pathway.
Supplemental Material
sj-xlsx-1-thc-10.1177_09287329251324829 - Supplemental material for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura
Supplemental material, sj-xlsx-1-thc-10.1177_09287329251324829 for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura by Zhongyi Zhu, Jing Wei, Ziyun Guo, Chang Liu, Lulu Jia and Yan Yang in Technology and Health Care
Supplemental Material
sj-xlsx-2-thc-10.1177_09287329251324829 - Supplemental material for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura
Supplemental material, sj-xlsx-2-thc-10.1177_09287329251324829 for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura by Zhongyi Zhu, Jing Wei, Ziyun Guo, Chang Liu, Lulu Jia and Yan Yang in Technology and Health Care
Supplemental Material
sj-xlsx-3-thc-10.1177_09287329251324829 - Supplemental material for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura
Supplemental material, sj-xlsx-3-thc-10.1177_09287329251324829 for Proteomic analysis of urine reveals biomarkers for identification of kidney injury in children's abdominal-type Henoch-Schönlein purpura by Zhongyi Zhu, Jing Wei, Ziyun Guo, Chang Liu, Lulu Jia and Yan Yang in Technology and Health Care
Footnotes
Acknowledgment
The authors have no acknowledgments.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Beijing Natural Science Foundation projects [grant numbers 7172076 and 7212169] and Key Discipline of Traditional Chinese Medicine for Children with Spleen and Stomach Diseases, Beijing Administration of Traditional Chinese Medicine, Beijing Key Program of Traditional Chinese Medicine II-8.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.