
Abstract
According to theories of embodied cognition, understanding a verb like throw involves unconsciously simulating the action of throwing, using areas of the brain that support motor planning. If understanding action words involves mentally simulating one’s own actions, then the neurocognitive representation of word meanings should differ for people with different kinds of bodies, who perform actions in systematically different ways. In a test of the body-specificity hypothesis, we used functional magnetic resonance imaging to compare premotor activity correlated with action verb understanding in right- and left-handers. Right-handers preferentially activated the left premotor cortex during lexical decisions on manual-action verbs (compared with nonmanual-action verbs), whereas left-handers preferentially activated right premotor areas. This finding helps refine theories of embodied semantics, suggesting that implicit mental simulation during language processing is body specific: Right- and left-handers, who perform actions differently, use correspondingly different areas of the brain for representing action verb meanings.
Theories of embodied cognition propose an intimate link between language and bodily experience. In this framework, to understand a word is to create an implicit mental simulation of its referent, using regions of the brain that support perception and action (e.g. Anderson, 2003; Barsalou, 2008; Zwaan, 2004). In keeping with this view, studies show that when participants read action-related verbs like kick, pick, and lick, they activate effector-specific regions of premotor cortex, as when they move the effector most associated with these verbs (i.e., their foot, hand, or tongue; Aziz-Zadeh, Wilson, Rizzolatti, & Iacoboni, 2006; Boulenger, Hauk, & Pulvermüller, 2009; Hauk, Johnsrude, & Pulvermüller, 2004; Tettamanti et al., 2005; but see Kemmerer & Gonzalez-Castillo, in press; Postle, McMahon, Ashton, Meredith, & de Zubicaray, 2008).
The goal of the present study was to refine the notion of implicit mental simulation during language processing by framing experimental predictions in terms of the body-specificity hypothesis (Casasanto, 2008, 2009): If concepts and word meanings are constituted, in part, by implicit simulations of our own perceptions and actions, then their neurocognitive representations should differ for people who perceive and act on the environment in systematically different ways. We investigated whether activity in cortical motor areas during action verb processing reflects the way an individual language user typically performs the action to which the verb refers.
Across neuroimaging studies, activity in cortical motor areas associated with manual-action verbs has been left-lateralized (Aziz-Zadeh et al., 2006; Hauk et al., 2004; Ruschemeyer, Brass, & Friederici, 2007; Tettamanti et al., 2005). This lateralization could be due to the general left-hemisphere dominance for language. Alternatively, it could be a consequence of testing only right-handed participants. We propose that if word meanings are implicit simulations, then understanding words for actions that people typically perform with their dominant hand should involve simulating these actions in contralateral premotor areas (i.e., areas that subserve planning of actions with the dominant hand).
To test this, we used functional magnetic resonance imaging (fMRI) to compare premotor activation in right- and left-handers during a lexical-decision task on manual-action verbs (e.g., grasp or throw) and nonmanual-action verbs (e.g., kneel or giggle). If the motor component of an action verb’s meaning is a body-specific simulation of the action it refers to as the particular language user would be likely to perform it, then activity in premotor cortex during manual-action verb processing should be differently lateralized in right- and left-handers. Each group should preferentially activate premotor areas contralateral to their dominant hand.
The nonmanual-action words served as a control. Finding the predicted difference between right- and left-handers for manual-action words, alone, could be evidence for implicit body-specific simulation of manual actions during word reading. Alternatively, it could be an artifact of differences in language laterality between right- and left-handers more generally. However, this alternative can be ruled out by testing for the predicted interaction of hemisphere (right premotor or left premotor) and handedness (right-handed or left-handed) in voxels that were significantly more active during presentation of manual- compared with nonmanual-action verbs (i.e., by testing for a particular three-way interaction of hemisphere, handedness, and verb type).
As a further control, after completing the lexical-decision task, participants performed a mental-imagery task in which they saw all of the manual- and nonmanual-action verbs again and were asked to consciously imagine performing the actions to which they referred. Comparing results from the two tasks allowed us to determine to what extent activation in motor areas during lexical decision corresponded to verbal semantics, per se, as opposed to explicit mental imagery.
Method
Participants
We tested 32 healthy participants with no known history of neurological problems, dyslexia or other language-related problems, or hearing complaints and with normal or corrected-to-normal vision, all of whom gave informed consent. First, we administered the Dutch version of the Edinburgh Handedness Inventory (EHI; Oldfield, 1971; Van Strien, 1992). Half of the participants were left-handed (n = 16, 12 women, 4 men; mean age: 23.4 years, age range: 19–32 years; EHI score M = –94.3, SD = 8.7, range: –82 to –100, mode = –100). The other half were right-handed (n = 16, 10 women, 6 men; mean age: 23.2 years, range: 20–29 years; EHI score M = 96.6, SD = 7.3, range = 82 to 100, mode = 100). The groups did not differ in age or in absolute EHI value, ts(30) < 1. The Radboud University ethics committee approved the study.
Materials
Stimuli were 96 Dutch verbs expressing concrete actions. Half were related to manual actions, and half were not related to manual actions. This distinction was pretested in a group of raters who did not participate in the fMRI experiment (n = 16), who scored for each verb how much they associated that action with their hand(s). Manual-action words were significantly more associated with hand actions than were nonmanual-action words, t(94) = 23.60, p < .001, p rep > .99. Of the materials that we used, on average, 79% of raters indicated that they tend to perform the action with their dominant hand (SD = 11.8%, Mdn = 81%, mode = 88%). Manual-action and nonmanual-action word lists did not differ in imageability (assessed by the same group of raters), t(94) < 1, number of phonemes, t(94) < 1, lexical frequency (taken from the CELEX database), t(94) < 1), or number of letters, t(94) = 1.51, p = .13, p rep = .78. From the materials that were rejected on the basis of the pretest, 16 filler items were created. Additionally, 16 phonotactically legal pseudowords were created, all with the suffix typical of the regular infinitive form in Dutch (-en). A list of 128 stimuli (48 manual-action words+48 nonmanual-action words+16 fillers+16 nonwords) was created and pseudorandomized with the constraint that the same condition was repeated maximally three times in a row. A mirrored version was presented to half of the participants.
Experimental procedure
Participants performed a lexical-decision task in the MRI scanner. Stimuli were presented using Presentation software (Version 10.2) and were projected from outside of the scanner room onto a mirror attached to the head coil. Words appeared for 1,500 ms in white font, in the middle of a black background. A fixation cross appeared for 500 ms before word presentation, to signal the start of a new trial. In 25% of the trials (i.e., filler and nonword trials), stimulus presentation was followed by a screen with the question of whether the previous stimulus was an existing word or not. Participants were instructed to answer as quickly and accurately as possible by pressing a button with their left or right index finger. No responses were made after critical stimuli (manual-action and nonmanual-action verbs). Response side varied between filler trials, so that sometimes the left button indicated ”yes” and sometimes the right button indicated ”yes.” This was done to prevent biased motor preparation to one response side. Participants had 1,500 ms to respond and got feedback on the screen when they were too slow. Participants were familiarized with the procedure by means of 10 practice trials before the start of the experiment, with stimuli not used in the remainder of the experiment.
In a second task, participants were required to perform explicit motor imagery. We included this task to be able to compare activations in the motor system in response to mere reading of the action verbs and during explicit motor imagery. The same manual-action and nonmanual-action words as in the lexical-decision task were presented. Now participants were instructed to read the word, close their eyes, actively imagine performing the action described by the verb, and open their eyes to have the next trial appear. Closing and opening of the eyes were monitored by an infrared eye tracker and were coded on-line by one of the experimenters. A list of 96 stimuli (manual-action and nonmanual-action words) was created and pseudorandomized with the constraint that the same condition be repeated maximally three times in a row. A mirrored version was presented to half of the participants. Participants were familiarized with the procedure by means of 10 practice items containing different words than used in the remainder of the experiment.
Stimulus onset was effectively jittered with respect to onset of volume acquisition by varying the intertrial interval between 2 and 6 s (M = 4 s) in 250-ms steps (Dale, 1999) in both tasks.
Data acquisition and analysis
Echo-planar imaging (EPI) of the whole brain was conducted with an eight-channel head coil on a Siemens MR system with 3T magnetic field strength, using a repetition time of 2,060 ms, echo time of 30 ms and flip angle of 85°. Thirty-one transversal slices with a 0.5-mm gap between slices were acquired per volume, with a voxel size of 3.5 × 3.5 × 3 mm. Preprocessing involved realignment through rigid body registration to correct for head motion, slice-timing correction to the onset of the first slice, normalization to Montreal Neurological Institute space, interpolation of voxel sizes to 2 × 2 × 2 mm, and spatial smoothing (8-mm full-width/half-maximum kernel). First-level analysis involved a multiple regression analysis with regressors describing the expected hemodynamic responses during observation of manual-action words, nonmanual-action words, filler words, nonwords (durations of 1,500 ms), and responses (button presses; modeled as stick functions). In the imagery task, a model with manual-action and nonmanual-action regressors was created, with durations corresponding to the actual imagery times.
MR disturbances due to small head movements were accounted for by a series of nuisance regressors, namely, the linear and exponential changes in the scan-by-scan estimated head motion, scan-by-scan average signals from outside the brain, white matter, and cerebrospinal fluid (Verhagen, Grol, Dijkerman, & Toni, 2006). Head motion never exceeded 3 mm or 3°.
Given our a priori hypothesis, we created subject-specific 4-mm spherical regions of interest with the maximally activated voxel to the manual-action stimuli (mapwise thresholded at p < .001) as the center coordinate, in cytoarchitectonically defined left and right Brodmann’s area (BA) 6 (premotor cortex; Eickhoff, Heim, Zilles, & Amunts, 2006). In this way, we optimized our analysis for each subject individually while controlling the location of the region of interest (ROI) by means of independent anatomical data (see also Aziz-Zadeh et al., 2006). We ensured that no voxels from the opposite hemisphere were taken into account (which could occur due to the spherical shape of the ROIs). We chose premotor cortex since most earlier studies of action verb processing had found effects here (Aziz-Zadeh et al., 2006; Kemmerer & Gonzalez-Castillo, 2008; Ruschemeyer et al., 2007; Tettamanti et al., 2005). However, as a control, a similar analysis was performed for primary motor cortex by using cytoarchitectonic maps of BA4 (combination of BA4a and BA4p; Geyer et al., 1996). Mean weights from the manual action > nonmanual action, manual action > 0, and nonmanual action > 0 contrasts were taken to a second-level analysis involving repeated measures analysis of variance with hemisphere (left or right) as a within-subjects factor and group (left-handers or right-handers) as a between-subjects factor. Given our a priori hypothesis, we tested a specific interaction direction (i.e., left-hemisphere activation > right-hemisphere activation in right-handers and right-hemisphere activation > left-hemisphere activation in left-handers).
Behavioral data analysis involved independent-samples t tests on percentage of correct trials, late responses (lexical-decision task), and imagery times (imagery task).
Results
Behavioral results
The groups did not differ in percentage of correct responses on the lexical-decision task. The percentage of correct responses for left-handers was 93.9% (SD = 5.5); the percentage for right-handers was 95.5% (SD = 3.6), t(30) < 1. There also was no difference between groups in the percentage of too-late responses. Left-handers responded too late on 0.58% of responses (SD = 1.26), and right-handers responded too late on 0.19% of responses (SD = 0.78), t(30) = 1.05, p = .30, p rep = .64.
In the mental-imagery control task, mean imagining times were 5.63 s (SD = 2.17) for manual-action verbs and 5.55 s (SD = 1.95) for nonmanual-action verbs. Right- and left-handers did not differ in imagining times, either overall or in manual-action or nonmanual-action times separately: for both manual-action and nonmanual-action verbs, t(30) = 1.15, p = .26, p rep = .68; for manual-action verbs only, t(30) = 1.20, p = .24, p rep = .69; for nonmanual-action verbs only, t(30) = 1.08, p = .28, p rep = .66. Neither were there within-group differences: for left-handers, t(15) = 1.64, p = .12, p rep = .79; for right-handers, t < 1.
Neural results
Lexical-decision task
Our main analysis tested for a three-way interaction among handedness, hemisphere, and verb type during a lexical-decision task using manual-action and nonmanual-action verbs, in subject-specific ROIs within premotor cortex (BA6). Results indeed showed a significant Hemisphere × Group interaction in the manual action > nonmanual action contrast values from the subject-specific ROIs, t(30) = 2.14, d = 0.64, p = .04, p rep = .89 (see Fig. 1a). For manual- compared with nonmanual-action verbs, left-handers had consistently stronger activations in right BA6, whereas right-handers showed stronger activation in left BA6 for this contrast, in keeping with predictions based on the body-specificity hypothesis. There were no main effects of hemisphere or group (all Fs < 1).
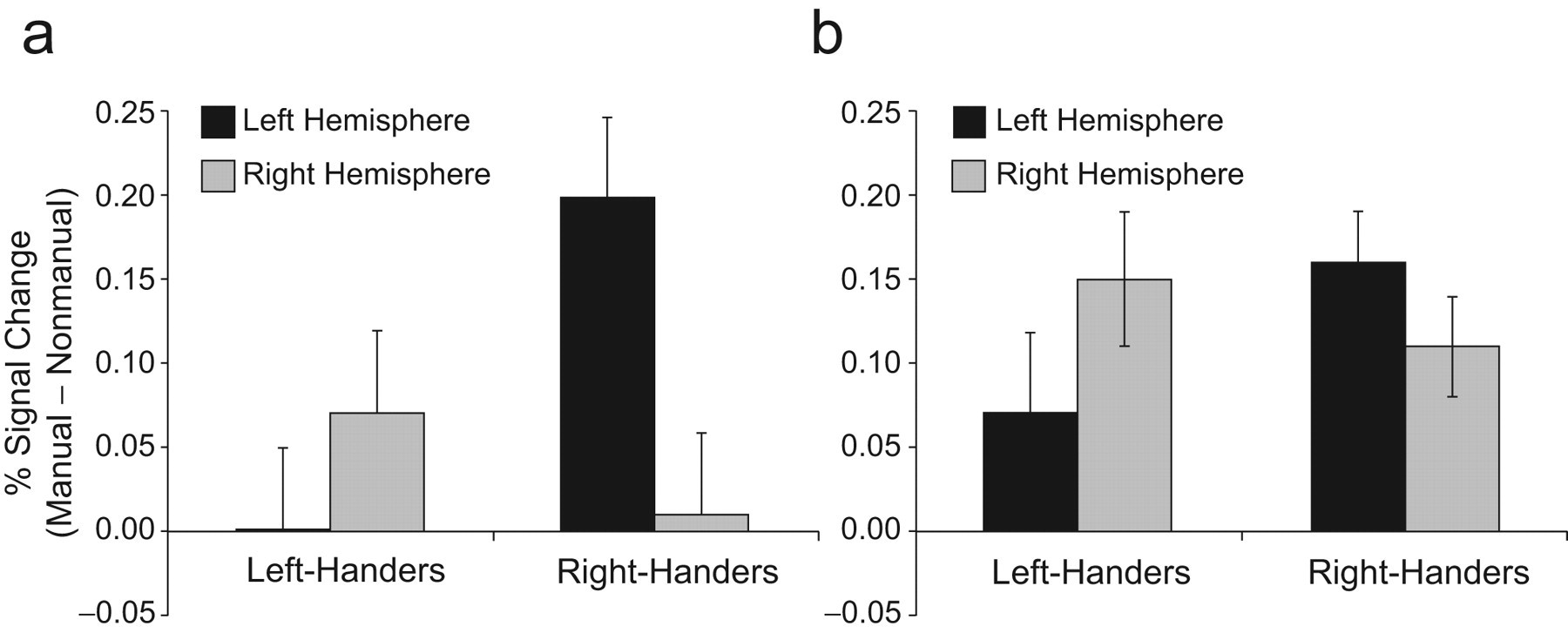
Percentage signal change as a function of handedness and hemisphere in the lexical-decision task. The graphs show the mean difference between manual- and nonmanual-action words for left-handers and right-handers in left-hemisphere and right-hemisphere Brodmann’s area 6 (premotor cortex; BA6). Error bars represent standard errors of the mean. Results are shown separately for (a) a 4-mm subject-specific spherical region of interest around the BA6 voxel maximally activated during manual-action words and (b) all BA6 voxels responsive to manual-action words (thresholded at p < .001).
To confirm that this three-way interaction was driven by the predicted difference in manual-action responses (rather than by incidental differences in nonmanual-action responses), we analyzed manual-action and nonmanual-action responses separately. For both verb types, the main effects of hemisphere and group were significant: manual-action verbs, main effect of hemisphere, F(1, 30) = 4.77, h p 2 = .14, MSE = 0.30, p = .04, p rep = .89; main effect of group, F < 1; nonmanual-action verbs, main effect of hemisphere, F(1, 30) = 2.72, h p 2 = .08, MSE = 0.35, p = .11, p rep = .81; main effect of group, F < 1. These main effects were qualified by the predicted two-way interaction of hemisphere and group in the premotor response to the manual-action stimuli, F(1, 30) = 5.71, h2 = .16, MSE = 0.30, p = .02, p rep = .93, but not to the nonmanual-action stimuli, F(1, 30) = 1.78, h2 = .05, MSE = 0.35, p = .19, p rep = .73.
To corroborate the subject-specific ROI analysis, we tested for the predicted interaction of handedness, hemisphere, and verb type in all voxels that responded to the presentation of manual-action words at p < .001 within our anatomical search region (left and right BA6). A pattern similar to the results of the subject-specific ROI analysis was found (see Fig. 1b), although the interaction was not significant using this more coarsely defined ROI: manual action > nonmanual action Hemisphere × Group interaction, t(30) = 1.59, d = 0.41, p = .12, p rep = .80; F < 1, for the main effect of either hemisphere or group.
Interestingly, there was considerable variation in the location of the subject-specific ROIs within our anatomical search region BA6. Whereas the majority of ROIs were relatively lateral in premotor cortex, the ROIs in a few participants were more medial in pre-supplementary motor area (pre-SMA), another part of BA6 also involved in motor planning (Nachev, Kennard, & Husain, 2008; see Table 1 and Fig. 2). A similar association between handedness and hemisphere was found in pre-SMA as in lateral premotor cortex.
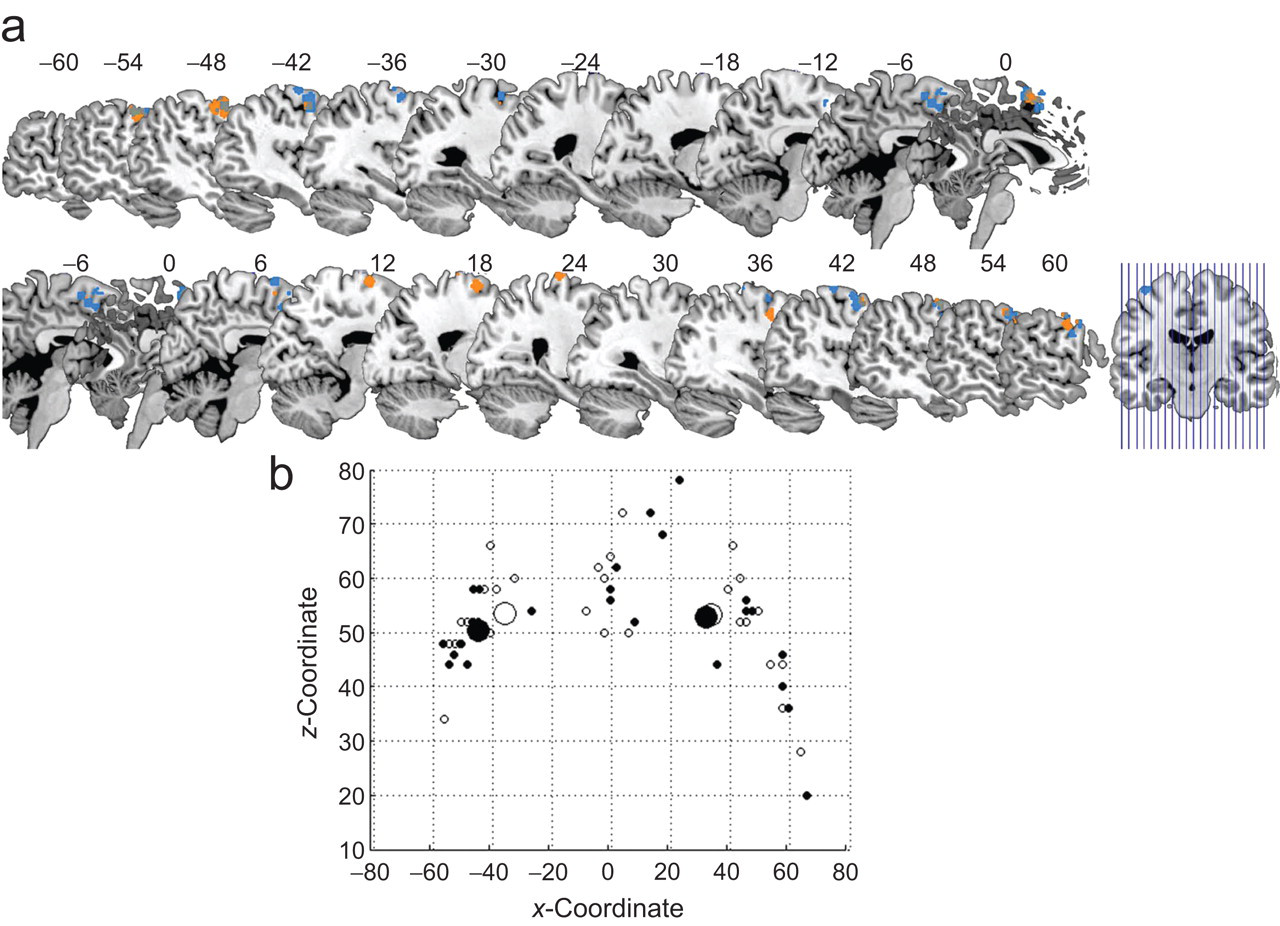
Locations of subject-specific regions of interest in Brodmann’s area 6 (premotor cortex). The images in (a) depict the regions of interest for all left-handers (orange) and right-handers (blue) on multiple sagittal slices (x-coordinates are indicated). The illustration on the right indicates the location of the sagittal slices. The graph in (b) shows the coordinates of peak voxels per region of interest for left-handers (small filled circles) and right-handers (small open circles). Large circles represent the mean coordinates for each group (see Table 1). Negative x-coordinates refer to the left hemisphere; positive x-coordinates refer to the right hemisphere. Coordinates are in Montreal Neurological Institute space.
Mean Coordinates for Subject-Specific Regions of Interest in Left and Right BA6

Note: Coordinates are in millimeters in Montreal Neurological Institute space. Mean locations are represented as the large circles in Figure 2b. Standard deviations are given in parentheses. BA6 = Brodmann’s area 6 (premotor cortex).
No effects of hemisphere or group were observed in BA4 (primary motor cortex), attesting to the specificity of this finding for premotor cortex: manual > nonmanual contrast, Fs(1, 30) < 1, for the main effects of hemisphere and group; manual-only verbs, F(1, 30) = 6.95, h p 2 = .19, MSE = 0.75, p = .01, p rep = .94, for the main effects of hemisphere, and F < 1, for the main effect of group; nonmanual-only verbs, F(1, 30) = 6.25, h p 2 = .17; MSE = 0.67, p = .02, p rep = .93, for the main effects of hemisphere, F < 1, for the main effects of group. The Hemisphere × Group interactions were nonsignificant as well, Fs(1, 30) < 1, for both verb types, and t(30) < 1, for the manual > nonmanual contrast. Similar effects were obtained when taking all responsive voxels in BA4 into account. Likewise, in a whole-brain analysis, no areas were significantly activated by the directional Group × Verb Type interactions.
Mental-imagery task
Data from the control task were analyzed using a similar procedure as for the lexical-decision task (creating subject-specific ROIs in BA6 and BA4 based on MAN < .001 voxels during explicit motor imagery). In the premotor ROI, this yielded a similar Hemisphere × Group interaction during mental imagery of actions, as was found during the reading of action verbs for the manual action > nonmanual-action contast: BA6, Hemisphere × Group interaction, t(30) = 4.04, d = 1.34, p < .001, p rep = .99; main effect of hemisphere, F(1, 30) = 6.42, h p 2 = .18, MSE = 0.023, p = .017, p rep = .93; main effect of group, F(1, 30) = 2.86, h p 2 = .09, MSE = 0.077, p = .10, p rep = .82. A Hemisphere × Group interaction was also found in the primary motor cortex ROI, for BA4: Hemisphere × Group interaction, t(30) = 2.41, d = 0.74, p = .02, p rep = .93; main effect of hemisphere (F < 1); main effect of group, F(1, 30) = 2.24, h p 2 = .07, MSE = 0.048, p = .15, p rep = .77, whereas this pattern of activation was restricted to premotor cortex during lexical decisions.
A comprehensive whole-brain analysis and full discussion of the mental-imagery data are beyond the scope of this report and will be reported elsewhere. Results of the ROI analyses for the imagery task are reported here primarily as a control for activation due to explicit imagery during the lexical-decision task (see the next paragraphs).
Comparison of activation during lexical-decision and mental-imagery tasks
We assessed whether activations during action verbs reading were influenced, or could be caused, by explicit motor imagery. First, we calculated correlations between the manual action > nonmanual action contrast estimates taken from the subject-specific ROIs determined on the lexical-decision data (see earlier discussion), during reading of action verbs (lexical-decision task) and during explicit motor imagery (mental-imagery task). That is, we tested whether subjects who exhibited a particular response on manual action > nonmanual action during lexical decisions would show a similar response during motor imagery in the ROIs in which a Group × Hemisphere interaction was observed during lexical decisions. Positive correlations would be expected if the lexical-decision activations were driven by explicit motor imagery. However, none of these correlations were significant, and all turned out to be negative (but nonsignificant; left BA6, R = –.05, p = .77, p rep = .30; right BA6, R = –.15, p = .41, p rep = .56; left BA4, R = –.25, p = .17, p rep = .75; right BA4, R = –.31, p = .08, p rep = .84).
In a further test of relationships between motor system activation during action word reading and action imagery, we analyzed the degree of spatial overlap in the subject-specific ROIs from the lexical-decision and visual-imagery tasks. The center voxels from the lexical-decision ROIs were spatially separated from the center voxels of the imagery ROIs, and the separation was variable over participants. The average euclidean distance between the peak voxels was 33.4 mm (SD = 21) for left BA4, 37.1 mm (SD = 18) for right BA4, 35.9 mm (SD = 17) for left BA6, 31.1 mm (SD = 21) for right BA6. Moreover, there was hardly any overlap between the lexical-decision and imagery ROIs (2.3% of shared voxels for left BA4, 0% for right BA4, 1.6% for left BA6, and 0.6% for right BA6). So, despite the similar body-specific pattern of responses during lexical decisions and conscious imagery, we found that reading of action verbs and explicit motor imagery engaged nonoverlapping parts of premotor cortex.
Finally, to test for possible overlap in whole BA6/BA4, multivoxel pattern analysis (Peelen, Wiggett, & Downing, 2006) was applied to the statistical maps of the manual action > nonmanual action comparison during lexical decisions and motor imagery. All voxels from BA6 and BA4 were considered, and a correlation was computed between the response pattern during the lexical-decision task and the mental-imagery task. No significant correlations were observed (all ts < 1). To demonstrate the sensitivity of this approach, we calculated correlations between the pattern of responses in left and right BA6 and BA4 within the lexical-decision and motor-imagery data. All within-task analyses yielded strong correlations between the hemispheres (all ps < .002, all p reps > .98), suggesting that the lack of correlation between tasks was not due to a lack of power. Rather, these analyses show that explicit motor imagery was not responsible for the activation we observed during lexical decisions.
Discussion
This study investigated whether the meanings of action verbs are grounded in the ways that particular language users perform the actions to which they refer. During a lexical-decision task, right- and left-handers showed contrasting patterns of activity in cortical motor regions when reading manual-action verbs like grasp and throw, as compared with reading nonmanual-action verbs. Each group preferentially activated premotor areas in the hemisphere contralateral to their dominant hands, suggesting that the motor component of manual-action verb semantics is body specific (Casasanto, 2008, 2009). People with different bodies, who perform actions in systematically different ways, use correspondingly different neural tissues for representing action verb meanings.
During a mental-imagery task, right- and left-handers also showed contrasting patterns of motor activity when imagining manual actions as opposed to nonmanual actions, demonstrating that explicit motor imagery is also body specific. Conscious mental imagery of actions that people usually perform with their dominant hand is subserved primarily by contralateral motor areas involved in planning and executing these actions.
Whereas a body-specific pattern of activation was found in both premotor cortex and primary motor cortex during explicit imagery, it was found only in premotor areas during lexical decisions. This dissociation between the neural substrates of action imagery and action verb semantics is consistent with previous results in right-handers. Tomasino, Werner, Weiss, and Fink (2007) and Tomasino, Fink, Sparing, Dafotakis, and Weiss (2008) found primary motor cortex involvement when participants were encouraged to actively imagine performing actions denoted by verbs, but not when participants read the same verbs in a letter-detection task (see also Aziz-Zadeh et al., 2006; Tettamanti et al., 2005). Although some research has implicated primary motor cortex in action verb semantics (see Pulvermüller, 2005), the majority of fMRI and transcranial magnetic stimulation studies implicate premotor cortex and suggest that primary motor activation during language-processing tasks may be an artifact of conscious imagery inadvertently cued by the stimuli (see Kemmerer & Gonzalez-Castillo, 2008; Papeo, Vallesi, Isaja, & Rumiati, 2009; Tomasino et al., 2007, 2008). Results of the present study support this conclusion.
Importantly, when we compared activity during the lexical-decision and mental-imagery tasks directly, we found no overlap between tasks in either primary or premotor areas and no correlation between activity during the two tasks in subject-specific premotor ROIs. We can therefore conclude with high confidence that the body-specific premotor activity we observed during the lexical-decision task was not due to conscious mental imagery, but rather to lexical processes, per se (see also Tettamanti et al., 2005; Tomasino et al., 2007, 2008).
These findings suggest two refinements to the notion of implicit mental simulation during language processing. First, embodied-cognition researchers sometimes suggest that unconscious simulation and conscious imagery are identical processes (e.g. Gallese & Lakoff, 2005). Indeed, some researchers have stated that they use the terms mental simulation and mental imagery synonymously (e.g., Bergen, Lindsay, Matlock, & Narayanan, 2007, p. 735). Yet, the present data urge caution in equating the notions of implicit simulation during language processing and explicit mental imagery.
Both implicit (unconscious) simulation and explicit (conscious) imagery engage body-specific representations in premotor cortex, but these representations may be different in character. When we read the verb throw, the corresponding premotor activity may constitute a far-less-elaborated action plan than when we consciously imagine throwing. Understanding throw apparently involves an action representation that is specified with respect to which arm we would use. But at least by default, the motor plan activated by a clause like throw the ball may lack the kinds of further specification that would be needed if we were to imagine this action explicitly (e.g., “Is it an underhand or overhand throw?” or “Is it a baseball or a basketball?”). At minimum, premotor representations underlying implicit simulation and explicit imagery must differ quantitatively, in the detail with which action plans are specified. Our data do not rule out the possibility that simulation and imagery could activate overlapping motor areas in certain contexts. But the finding of distinct premotor representations for these processes raises the possibility that they may differ qualitatively, as well as quantitatively.
A second refinement these data suggest concerns the perspective that readers adopt when understanding action words. On the basis of previous studies (e.g., Tomasino et al., 2007), it was not clear to what extent the motor component of action word meanings reflected actions one has observed others performing (allocentric simulation) or actions one has performed (egocentric simulation). The allocentric possibility predicts that neurocognitive representations of manual-action word meanings should be similar in right- and left-handers, since presumably everyone observes about the same proportion of right- and left-handed actions by other people. The discovery that motor activity associated with manual action words is body specific supports the egocentric possibility and suggests that people implicitly simulate their own prior or potential actions when understanding action words.
Is the perspective that people adopt when understanding action words fixed? We suggest that the egocentric simulations participants appear to have created during the lexical-decision task reflect a default interpretation of the words’ meanings, but not the only possible interpretation. Furthermore, these results do not imply that people can only understand language about actions they have performed themselves. Someone who has never used chopsticks could still understand the sentence He picked up the dumpling with chopsticks, plausibly by creating a motor simulation based on visual experiences of other people using chopsticks and on extrapolation from familiar motor actions (eating with a fork, holding a pencil between the fingertips, etc.; see Anderson, 2003). It remains an open question whether people can understand words referring to actions they have never performed as completely as words for actions they have and whether implicit simulations of actions they have never performed are body specific (see Aglioti, Cesari, Romani, & Urgesi, 2008; Calvo-Merino, Grezes, Glaser, Passingham, & Haggard, 2006).
The functional significance of premotor activity for language understanding also remains a question for ongoing research. Due to the correlational nature of fMRI data, it is not possible to infer whether the premotor activity reported here is a necessary part of action verb semantics (Willems & Hagoort, 2007). Although some constituents of word meaning may be abstracted away from our physical experiences (Hagoort, 1998; Mahon & Caramazza, 2008), the present results suggest that at least part of action word meaning consists of implicit mental simulations of actions as a person would perform them with his or her particular body.
Footnotes
Acknowledgements
We thank Martin Laverman, Jacqueline de Nooijer, Daan van Rooij, and Paul Gaalman for their assistance.
The authors declared that they had no conflicts of interests with respect to their authorship and/or the publication of this article.
This research was supported by a National Research Service Award Fellowship (F32MH072502) from the National Institutes of Health to Daniel Casasanto and by a grant from the European Union Joint-Action Science and Technology Project (IST-FP6-003747).