
Abstract
In order to engage in goal-directed behavior, cognitive agents have to control the processing of task-relevant features in their environments. Although cognitive control is critical for performance in unpredictable task environments, it is currently unknown how it affects performance in highly structured and predictable environments. In the present study, we showed that, counterintuitively, top-down control can impair and interfere with the otherwise automatic integration of statistical information in a predictable task environment, and it can render behavior less efficient than it would have been without the attempt to control the flow of information. In other words, less can sometimes be more (in terms of cognitive control), especially if the environment provides sufficient information for the cognitive system to behave on autopilot based on automatic processes alone.
A foundational issue in psychology and cognitive neuroscience concerns how cognitive agents control the processing of features in their environment (Miller, 2000; Shiffrin & Schneider, 1977). Cognitive control is often claimed to be adaptive in the sense that it counteracts or regulates automaticity in order to optimize goal-directed behavior; it thus enhances automatic processes by prioritizing the processing of features that are task relevant (Botvinick, Braver, Barch, Carter, & Cohen, 2001; Ridderinkhof, Ullsperger, Crone, & Nieuwenhuis, 2004). However, as we demonstrated in the present study, there are limits to the usefulness of cognitive control. We showed, in particular, how overcontrol (here induced by task instructions) can prevent the otherwise automatic exploitation of statistical stimulus characteristics needed to optimize behavior.
Although a feature’s relevance is generally construed in terms of an agent’s endogenous goal (Miller, 2000; Ridderinkhof et al., 2004), features can also be relevant because of their diagnosticity (e.g., if a feature value statistically predicts a particular or correct response irrespective of the agent’s conceptualization of the task and his or her attentional set). Previous research has shown that top-down cognitive control is especially important for performance in unpredictable task environments, where there is no simple statistical cue predicting what responses should be made (Shiffrin & Schneider, 1977; Treisman & Gelade, 1980), but it is currently unknown how cognitive control affects performance in structured environments that contain highly predictable exogenous cues. In the present study, we investigated the effect of top-down control in a task containing a predictive feature that was unrelated to the task instructions.
Participants performed a two-alternative forced-choice task on a foveally presented stimulus that could vary on a subset of binary perceptual features, such as color (red, green), shape (diamond, square), size (large, small), topology (open, closed), and location (up, down). Unbeknownst to the participants, we manipulated the statistical informativeness of an additional feature that was not part of the task, such that this feature always predicted the correct response in one condition (the predictive condition) but not in the other condition (the baseline condition). Because the cognitive system is known to exploit statistical stimulus-response contingencies automatically (Shiffrin & Schneider, 1977), performance was expected to be better in the predictive than in the baseline condition.
We embedded these predictive and baseline conditions into two different tasks, which we thought would induce different cognitive-control states. The control task included instructions intended to emphasize the need for top-down control: Participants were instructed to classify the stimulus according to a feature-conjunction rule (e.g., size and topology: left response key for large and open or small and closed shapes, right response key for small and open or large and closed shapes). The automatic task included instructions intended to deemphasize the need for control: Participants were instructed to classify the stimulus according to a single feature (e.g., shape: left response key for a diamond and right response key for a square). In the automatic task, the features were mapped consistently on responses and thus allowed automatic visuomotor translation. In contrast, the stimulus-response mapping in the control task required the attention-demanding integration of two features before the response could be determined (Treisman & Gelade, 1980).
Method
Participants
Thirty-eight Leiden University undergraduates participated for course credit in Experiments 1 (n = 12), 2 (n = 12), 3 (n = 12), and 4 (n = 2).
Procedure
In all four experiments, each trial started with a fixation cross (500 ms), followed by the perceptual stimulus (until response), and performance feedback (500 ms). Each stimulus consisted of a geometric shape (e.g., diamond, square), and stimuli were presented one at a time in the fovea, the area of the eye in which vision is sharpest. The experimental conditions were blocked and alternated throughout each experiment (32 trials per block). Participants performed 48 practice trials prior to each task and 16 practice trials prior to each block. Trial order was randomized for each block.
Experiment 1 had a 2 (task: control vs. automatic) × 2 (feature condition: predictive vs. baseline) design (Fig. 1a). Participants first performed the control task (6 blocks), followed by the automatic task (6 blocks). Stimuli consisted of closed and open diamonds and squares presented in the center of the display. In the control task, participants had to classify the stimulus according to two features—size and topology. Participants were instructed to press one response key if the stimulus was large and open or small and closed, whereas they were told to press the other key if the stimulus was small and open or large and closed. In the automatic task, participants merely had to press one key if the stimulus was a diamond or another key if the stimulus was a square. In the predictive condition, the color of the stimulus predicted the appropriate response key: Each stimulus was either green or red, and the same color was always associated with the same size and topology, and thus the same response key (i.e., green with the left key, red with the right). In the baseline condition, all stimuli were the same color and thus were not predictive.
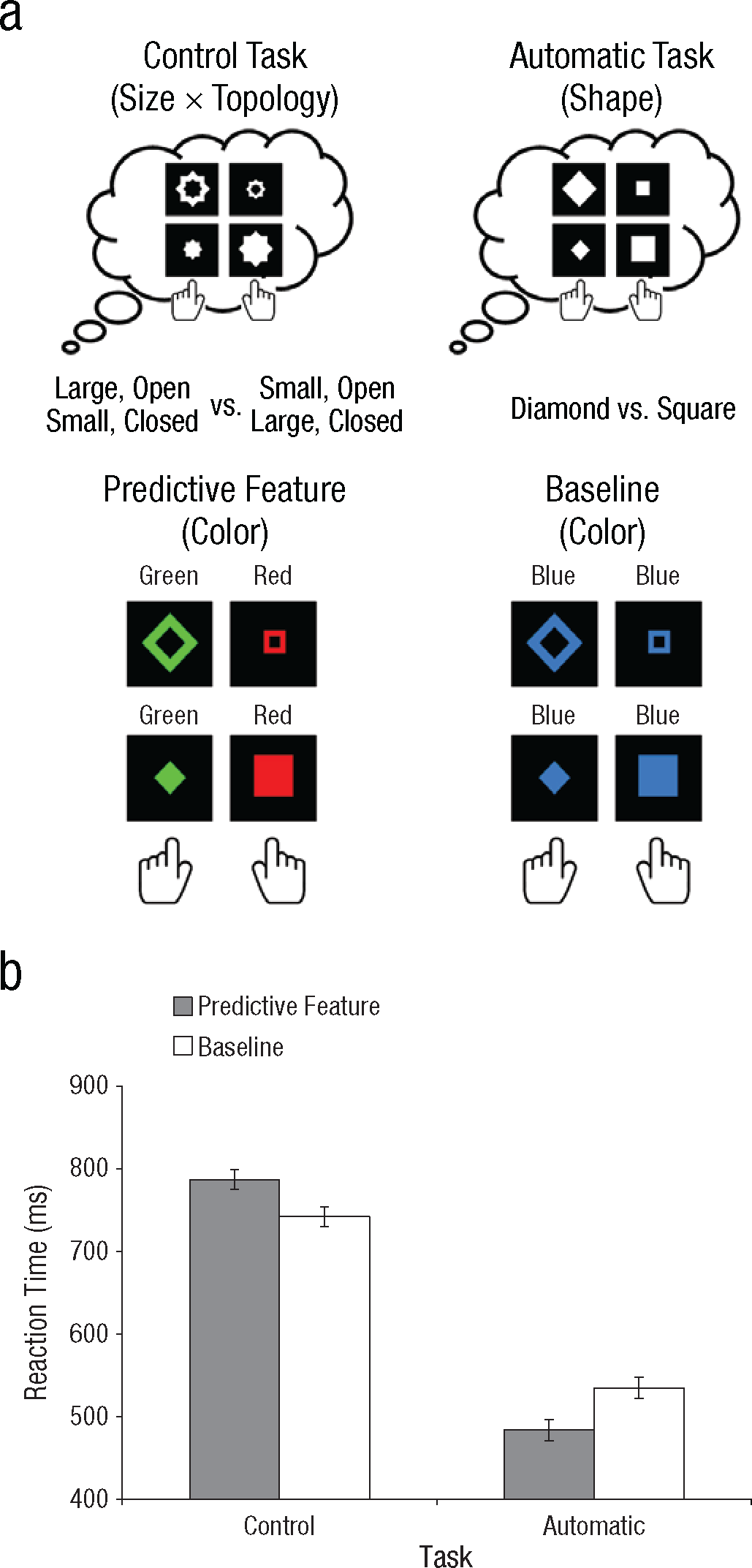
Design (a) and results (b) of Experiment 1. Participants performed two tasks (a; top row). In the control task, they were instructed to classify individual stimuli according to both their size (large vs. small) and their topology (open vs. closed). In the automatic task, participants were instructed to classify stimuli only on the basis of their shape (diamond vs. square). In both tasks, participants completed two conditions (a; bottom row). In the predictive condition, the color of the stimulus (red vs. green) predicted the correct response key used to classify its size and topology. In the baseline condition, all stimuli had the same color, which was therefore not predictive of the correct response key. The graph (b) shows mean reaction time on trials with correct responses as a function of task type and feature condition. Error bars indicate within-subjects standard errors of the mean.
The design of Experiment 2 was similar to that of Experiment 1 (Fig. 2a). As in Experiment 1, participants first performed the control task (6 blocks), followed by the automatic task (6 blocks). Stimuli consisted of filled diamonds and squares. In the control task, participants had to classify stimuli according to two features: size and color. They pressed the left response key for stimuli that were large and red or small and green and the right response key for stimuli that were small and red or large and green. In the automatic task, participants only had to indicate whether each stimulus was a diamond or a square, as in Experiment 1. In the predictive condition, the location of the stimulus predicted the correct response key: Stimuli associated with the left response key were slightly offset toward the top of the display, whereas stimuli associated with the right response key were offset toward the bottom of the display. In the baseline condition, all stimuli appeared in the center of the screen. In both Experiments 1 and 2, task order was not counterbalanced in order to minimize the chance that participants would discover the predictive feature.
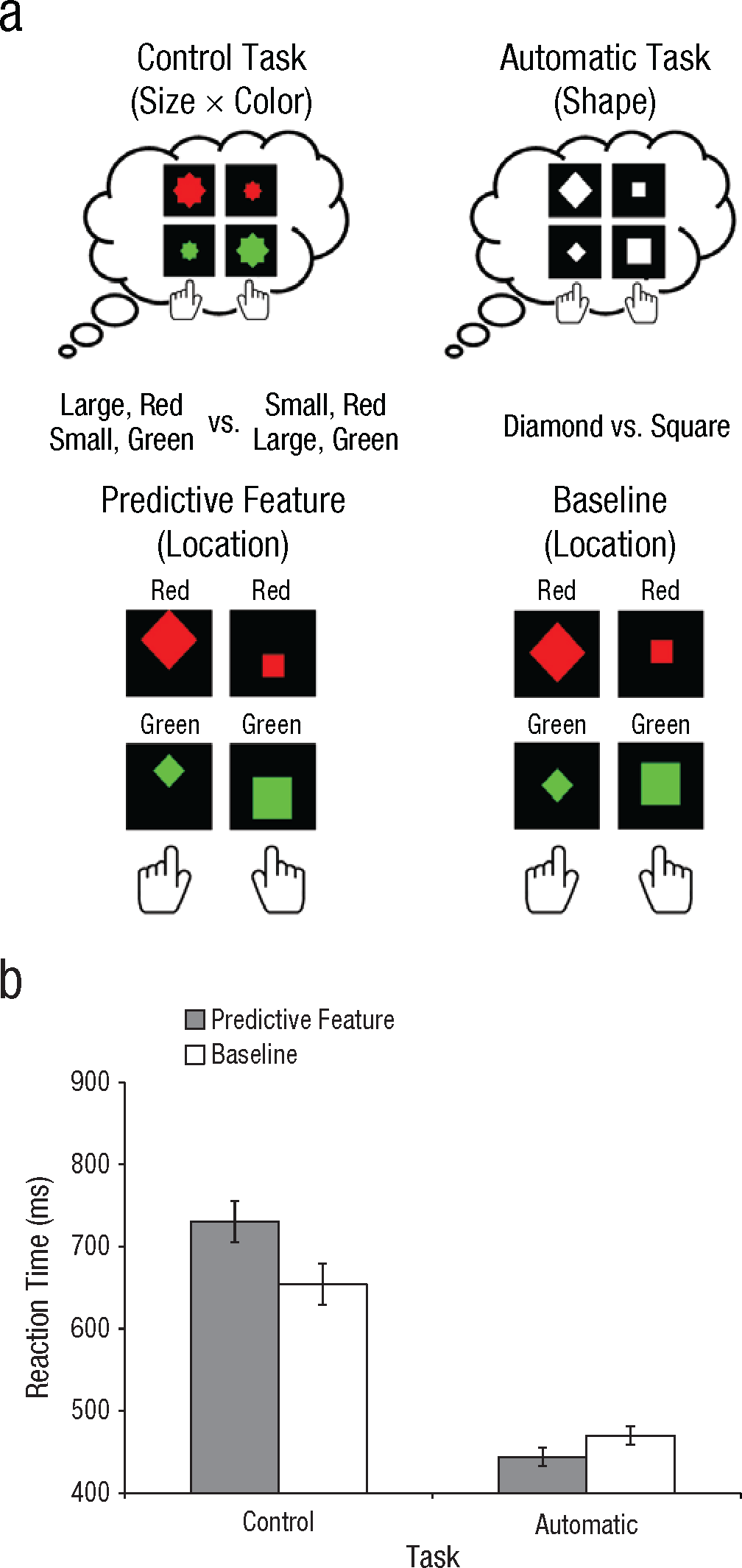
Design (a) and results (b) of Experiment 2. Participants performed two tasks (a; top row). In the control task, they were instructed to classify individual stimuli according to both their size (large vs. small) and their color (red vs. green). In the automatic task, participants were instructed to classify stimuli only on the basis of their shape (diamond vs. square). In both tasks, participants completed two conditions (a; bottom row). In the predictive condition, the location of the stimulus (top vs. bottom) predicted the correct response key used to classify its size and color. In the baseline condition, all stimuli had the same location, which was therefore not predictive of the correct response key. The graph (b) shows mean reaction time on trials with correct responses as a function of task type and feature condition. Error bars indicate within-subjects standard errors of the mean.
In Experiment 3, participants performed only the automatic task (6 blocks; Fig. 3a). Participants indicated whether each stimulus was tilted 2° clockwise (right response key) or 2° counterclockwise (left response key). Stimuli consisted of large and small stars, which were presented both open and closed. In the predictive condition, stimuli tilted clockwise were red, and those tilted counterclockwise were green. Thus, the color always predicted the direction in which the stimulus was tilted and, more important, the correct response. In the baseline condition, all stimuli were the same color (blue), which was therefore not predictive of the correct response.
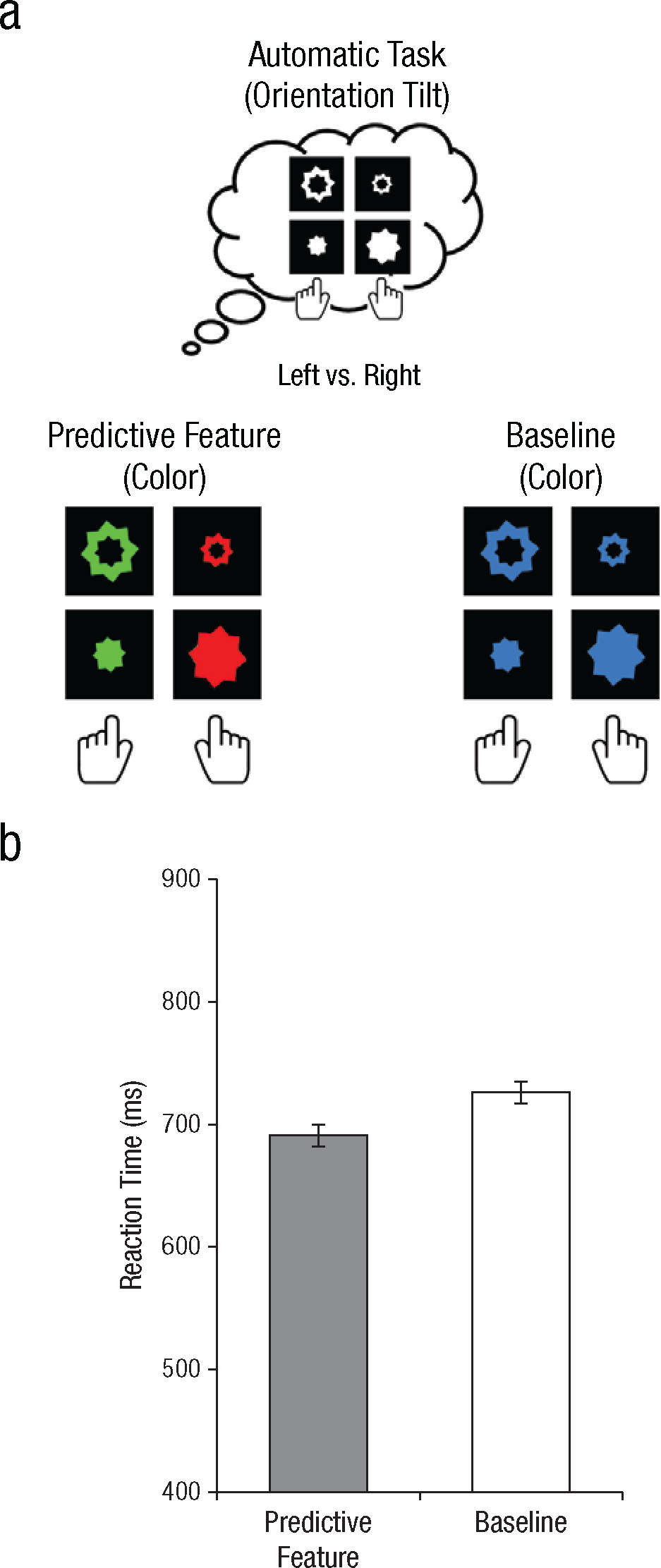
Design (a) and results (b) of Experiment 3. Participants performed only the automatic task (a; top row), in which they were instructed to classify stimuli only on the basis of their orientation (tilted 2° clockwise vs. tilted 2° counterclockwise). Participants completed two conditions (a; bottom row). In the predictive condition, the color of the stimulus (red vs. green) predicted the correct response key used to classify its orientation. In the baseline condition, all stimuli had the same color, which was therefore not predictive of the correct response key. The graph (b) shows mean reaction time on trials with correct responses as a function of feature condition. Error bars indicate within-subjects standard errors of the mean.
In Experiment 4, participants performed 12 different featural variants of the control task (48 blocks). For each variant, participants were instructed to categorize stimuli according to two dimensions. A third dimension either predicted or did not predict the correct response, as it was either correlated or uncorrelated with the correct response, respectively. The dimensions consisted of color (red, green), shape (diamond, square), size (large, small), topology (open, closed), and location (up, down). All 12 possible combinations of dimensions were tested (see Fig. 4).
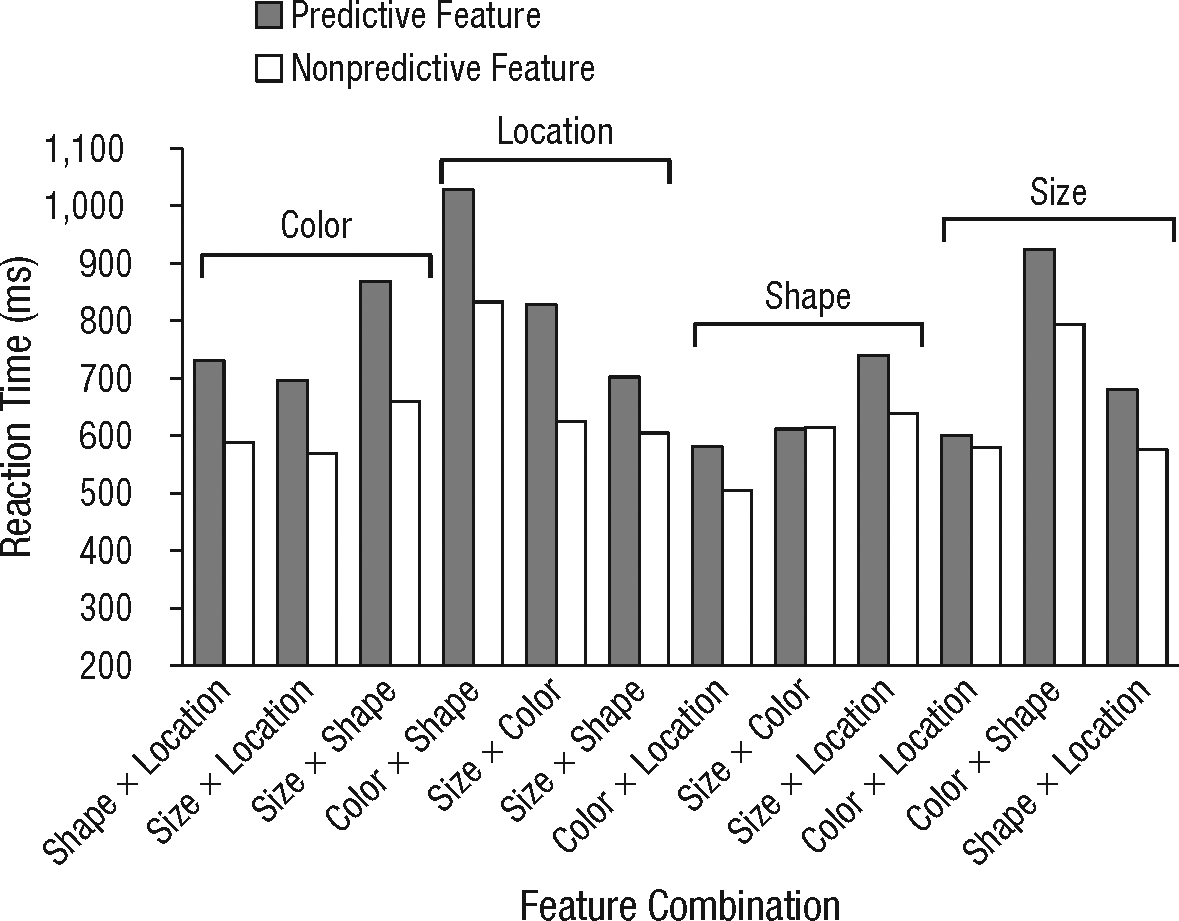
Results from Experiment 4: mean correct reaction time for each variant of the control task as a function of feature condition. For each version of the control task, participants categorized stimuli according to two dimensions (shown along the x-axis) while a third dimension (shown above the data bars) either predicted or did not predict the correct response.
All participants reported after the experiment that they adhered strictly to the classification rule that they were instructed to use. None reported being aware of the predictive nature of the correlated feature in the control task, nor were they aware of the fact that the automatic and control tasks were exactly the same in terms of what stimuli were mapped to what responses.
Data analysis
Reaction times faster than 100 ms and slower than 3,000 ms were discarded (< 1%). Error rates were low (~3–5%) and did not show signs of speed/accuracy trade-offs.
Results
If the automatic pickup of stimulus-response contingencies would be enhanced by exerting strong top-down control, as the adaptive-control assumption suggests, participants should always have shown better performance in the predictive than in the baseline condition and perhaps have exhibited a more pronounced predictive-feature benefit in the control task than in the automatic task. This is not what we observed in Experiment 1, however (see Fig. 1b). The predictive feature improved performance in the automatic task, t(11) = 3.59, p < .01, but it actually impaired performance in the control task, t(11) = 3.58, p < .01. In Experiment 2, we replicated both these effects using a different combination of features (see Fig. 2b): The predictive feature again improved performance in the automatic task, t(11) = 2.33, p < .05, but impaired performance in the control task, t(11) = 2.85, p < .02. Critically, this shows that although participants successfully picked up on the predictive feature, engaging in cognitive control turned this knowledge into a disadvantage.
Although the control and automatic tasks induced different cognitive-control states, they also differed in sheer task difficulty. In Experiment 3, participants performed a perceptually difficult automatic task in which they indicated whether the target was tilted 2° clockwise or 2° counterclockwise. The predictive feature (stimulus color) improved performance despite the high level of task difficulty (Fig. 3b), t(11) = 3.96, p < .01, which indicates that the opposite predictive-feature modulations in the control and automatic tasks (Experiments 1 and 2) cannot be explained by differences in task difficulty.
Was the counterintuitive predictive-feature impairment in the control tasks (Experiments 1 and 2) perhaps due to an increase in featural variability? In Experiment 4, we allowed the predictive feature to vary with the responses in a less deterministic fashion, in which feature and response were either correlated (i.e., predictive) or uncorrelated (i.e., nonpredictive). We observed the same impairment, which indicates that statistical predictability, not featural variability as such, was the critical factor (see Fig. 4). Furthermore, we found that the costs of cognitive control generalized reliably over all possible featural variants of the tasks, t(11) = 6.09, p < .001.
Discussion
In the present study, we investigated the effect of top-down control in a task containing a predictive feature that was unrelated to the task instructions. As expected, the predictive feature improved performance when participants performed the task automatically. Counterintuitively, however, the predictive feature impaired performance when subjects were performing the exact same task in a top-down, controlled manner. It is important to note that we excluded the possibility that these opposite modulations in performance were due to differences in task difficulty between the automatic task and the control task. Furthermore, we showed that the impairment in the control task was due to the statistical informativeness of the predictive feature and could not be attributed to an increase in featural variability. Also, we replicated the performance impairment for various featural variants of the control task, which suggests that the effect generalized to different perceptual features. 1
Cognitive control is commonly claimed to be adaptive because it allows cognitive agents to overcome automatic responses that will lead to disadvantageous actions (Miller & Cohen, 2001). Indeed, in most experimental conflict paradigms, top-down control is clearly adaptive: Participants have to overcome the activation of an erroneous response by a feature that is related to the task set (e.g., when naming the ink color of a word whose letters spell an incompatible color name; see Botvinick et al., 2001). The present study differed in this respect in that we investigated the effect of cognitive control on the activation of a correct response by a feature that is unrelated to the task set. It is precisely because of this difference that our paradigm was able to demonstrate the circumstances under which top-down cognitive control is suboptimal.
One could argue that the suboptimal performance we observed was a consequence of the way our instructions in the control task prevented participants from using the predictive feature. However, the presence of the predictive feature led to a cost in performance in this task, which cannot be explained by a mere failure to process this feature. Instead, this finding suggests that participants successfully picked up on the predictive feature but were unable to use this information to their advantage.
Why did the addition of a predictive feature impair performance in the control task but improve performance in the automatic task? A possible way to explain this interaction is to distinguish between the processes responsible for stimulus-response (S-R) translation and the processes responsible for the ultimate selection of a response (for a broader discussion, see Hommel, 1998). Specifically, we have to assume that (a) S-R translation operates automatically whenever there is a statistical association between a feature and a response (Hommel, 1998) and that (b) response selection can occur in either an automatic parallel fashion or a controlled serial fashion, depending on the way features and responses are mapped onto each other in the task set (Schneider & Shiffrin, 1977; Shiffrin & Schneider, 1977). Controlled serial selection occurs when the task set contains a variable mapping between features and responses, whereas automatic parallel selection occurs when the task set contains a consistent mapping between features and responses.
Under variable mapping conditions, all currently active outcomes of S-R translations are checked serially for whether the eliciting feature matches one of the attended dimensions in the task set—a serial memory search in the sense posited by Sternberg (1969). From this perspective, the deficit in the control task is due to the additional processing time required for the control mechanism to reject the extra outcomes of S-R translations because of the presence of the predictive feature. However, under consistent mapping conditions, all outcomes of S-R translation have parallel access to response selection, which explains the benefit in the automatic task: It is driven by an increase in correct response activation resulting from the presence of extra outcomes of S-R translation. Thus, although multiple S-R translations can occur in parallel, only one of these can be checked at a time against the dimensions in the task set during controlled response selection—a processing characteristic that also accounts for dual-task costs (Hommel, 1998; see also Cohen & Shoup, 2000).
Although our findings suggest important limitations on the usefulness of cognitive control, they need not imply that cognitive agents cannot function adaptively over large time scales: It is quite plausible that, perhaps through trial and error, a momentarily unattended—though informative—stimulus dimension may become increasingly likely to be attended by the agent and thus become exploited advantageously.
Our findings are consistent with recent theoretical proposals that the optimal level of cognitive control depends on the task at hand: High levels of control are best suited for explicit, rule-based, verbal tasks that depend on the capacity limits of working memory, whereas low levels of control are best suited for implicit, reward-based, nonverbal tasks that can be accomplished irrespective of working memory limitations (Chrysikou, Weber, & Thompson-Schill, 2013). Across various cognitive domains, convergent findings suggest that a high level of control may benefit performance if a participant already knows how to perform a task but may not be optimal if the task still has to be learned (Thompson-Schill, Ramscar, & Chrysikou, 2009). A particularly striking example of this phenomenon has recently been reported by Rydell, Shiffrin, Boucher, Van Loo, and Rydell (2010). In their study, a stereotype-threat manipulation caused women to employ a highly controlled task strategy in a visual search paradigm, which prevented them from learning how to automatically allocate attention to the consistently mapped targets. In contrast, women who were not exposed to the stereotype-threat manipulation did successfully learn, presumably because they exerted less top-down control.
The performance cost that we observed in the current study adds to these findings by showing that although participants were able to successfully learn the predictive contingency between the implicit feature and the responses, they had no adaptive control over this information. Thus, the adaptive nature of cognition lies not in the control mechanism itself but in the voluntary regulation of one’s control settings in a way that best suits both the intended goal and the external context—a kind of metacontrol (Memelink & Hommel, 2012).
In conclusion, our findings suggest that automatic processes are less automatic and control processes less adaptive than commonly assumed (Botvinick et al., 2001; Miller, 2000; Ridderinkhof et al., 2004). Apparently, inducing a state of strong top-down control can impair and interfere with the otherwise automatic exploitation of statistical stimulus and response information, and it can render behavior less efficient than it would have been without the attempt to control the flow of information. In other words, less can sometimes be more (in terms of cognitive control), especially if the environment provides sufficient information for the cognitive system to behave on autopilot based on automatic processes alone. Our most important conclusion is that the adaptive function of cognitive control lies not in the workings of the control mechanism itself, but in finding a suitable balance between top-down control (when necessary) and automaticity (when suitable; Cools & D’Esposito, 2011). Hence, cognitive control is adaptive only to the degree that it can restrict its impact to conditions in which it is really required.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.