
Abstract
Normal and abnormal differences in sustained visual attention have long been of interest to scientists, educators, and clinicians. Still lacking, however, is a clear understanding of how sustained visual attention varies across the broad sweep of the human life span. In the present study, we filled this gap in two ways. First, using an unprecedentedly large 10,430-person sample, we modeled age-related differences with substantially greater precision than have prior efforts. Second, using the recently developed gradual-onset continuous performance test (gradCPT), we parsed sustained-attention performance over the life span into its ability and strategy components. We found that after the age of 15 years, the strategy and ability trajectories saliently diverge. Strategy becomes monotonically more conservative with age, whereas ability peaks in the early 40s and is followed by a gradual decline in older adults. These observed life-span trajectories for sustained attention are distinct from results of other life-span studies focusing on fluid and crystallized intelligence.
The ability to sustain attention to a task over seconds to minutes is a core cognitive function that plays a critical role in daily functioning. For example, sustained attention has been linked to academic and employment performance (Kalechstein, Newton, & van Gorp, 2003; Lam & Beale, 1991), and attentional lapses predict driving accidents (Ball, Owsley, & Sloane, 1991; Edkins & Pollock, 1997; Schmidt et al., 2009). Research has also shown that numerous other cognitive processes, such as learning, memory, and executive functions, rely on sustained attention (Barkley, 1997; Sarter, Givens, & Bruno, 2001; Silver & Feldman, 2005). Furthermore, deficits in sustained attention are one of the most pervasive cognitive issues across a wide range of neurological and psychiatric patient populations (Buxbaum et al., 2004; Clark, Iversen, & Goodwin, 2002; Park, Hood, Shah, Fogg, & Wyatt, 2012). Despite numerous investigations of how other cognitive processes (e.g., verbal ability or processing speed) develop across people’s life, life-span changes in sustained attention have not yet been fully characterized. Filling this gap will not only inform cognitive models of human development and aging, but also will help better define and pinpoint the mechanisms of cognitive dysfunction in neurologic and psychiatric populations.
The studies completed thus far on age-related changes in sustained-attention ability have yielded inconsistent results (Staub, Doignon-Camus, Després, & Bonnefond, 2013), including poorer performance in both childhood and aging relative to adulthood (McAvinue et al., 2012), no changes between younger and older adults (Bunce & Sisa, 2002; Staub et al., 2013), and improved functioning in older adults (Carriere, Cheyne, Solman, & Smilek, 2010; Staub, Doignon-Camus, Bacon, & Bonnefond, 2014). One potential cause for these disparate findings is that by focusing on error rates and not comparing hit rates with false alarm rates, previous studies have not dissociated changes in strategy and ability across the life span (Sarter et al., 2001). Moreover, the primary type of errors made (commissions vs. omissions) can vary with the task, which makes comparisons difficult (Staub et al., 2013).
Recently, our laboratory developed the gradual-onset continuous performance task (gradCPT), the aim of which is to better characterize individual differences in sustained attention (Esterman, Noonan, Rosenberg, & DeGutis, 2013; Rosenberg, Noonan, DeGutis, & Esterman, 2013). The gradCPT represents a unique combination of task features in that it both requires frequent overt responses and removes abrupt stimulus onsets that exogenously capture attention. Requiring frequent overt responses is common in continuous performance tasks (e.g., sustained-attention-to-response task, or SART; Robertson, Manly, Andrade, Baddeley, & Yiend, 1997) and allows for reliable analyses of response timing and variability, and accuracy across the whole task as well as within periods of high and low attentional stability (in and out of the zone, respectively). Additionally, the use of gradual stimulus changes makes performance less tied to phasic stimulus onsets and offsets, which better isolates intrinsic sustained-attention abilities. To separate the contribution of strategic changes and ability factors, we have also successfully utilized signal detection analyses of gradCPT (Esterman, Reagan, Liu, Turner, & DeGutis, 2014). Finally, several recent studies suggest that the gradCPT is ecologically valid because a variant of the gradCPT correlates with real-world attentional problems (Rosenberg et al., 2013), and performance on the original version is impaired in patient populations with traditionally poor sustained attention (Auerbach et al., 2014; DeGutis et al., 2015). Together, this suggests the gradCPT is a powerful tool to capture changes in sustained-attention ability throughout the life span.
To help resolve how sustained attention changes across the life span, we tested an unprecedentedly large sample of 10,430 participants between 10 and 70 years old on an adapted 4-min version of the gradCPT (Esterman et al., 2013). This sample was larger than in all previous efforts to model changes in sustained-attention performance during development, aging, or across the life span combined, which allowed us to more precisely model transition periods in performance across the life span using segmented linear regression. We also employed factor analyses and confirmed the existence of two latent, dissociable factors underlying gradCPT performance—the ability to sustain attention (discrimination performance and response time consistency) and the strategic approach (response speed and carefulness). The results show unique patterns in how the ability and strategy factors change across the life span. In addition, they suggest that the life-span trajectory of sustained-attention ability is unique from the trajectories other studies have found for crystallized intelligence (e.g., vocabulary), which continues to improve throughout the life span until the mid-60s, as well as fluid intelligence (e.g., working memory), which peaks in the mid-20s (Craik & Bialystok, 2006; Hartshorne & Germine, 2015).
Method
Participants
We included 10,430 unpaid volunteers between the ages of 10 and 70 years (M = 26.07, SD = 11.77) in the analyses (Fig. 1). Participants were visitors to TestMyBrain.org, a cognitive testing Web site. Data from repeat participation were excluded. Within 7 months, 10,922 people completed the task. Of these participants, 342 were excluded because of missing data or technical problems. Among technical problems, we chose to exclude computers that exceeded 10% error in the average stimulus presentation time, that is, the time it took for a new image to transition from 0% to 100% opacity. This included average stimulus presentation times that were too fast (≤ 720 ms) or too slow (≥ 880 ms). From the remaining 10,580 participants another 150 (1.4%) were excluded for “tune-outs,” defined as intervals of 30 s or more without a response. Of the 10,430 participants that were included, there was a nearly equal ratio of males and females (5,027 males, 5,403 females).
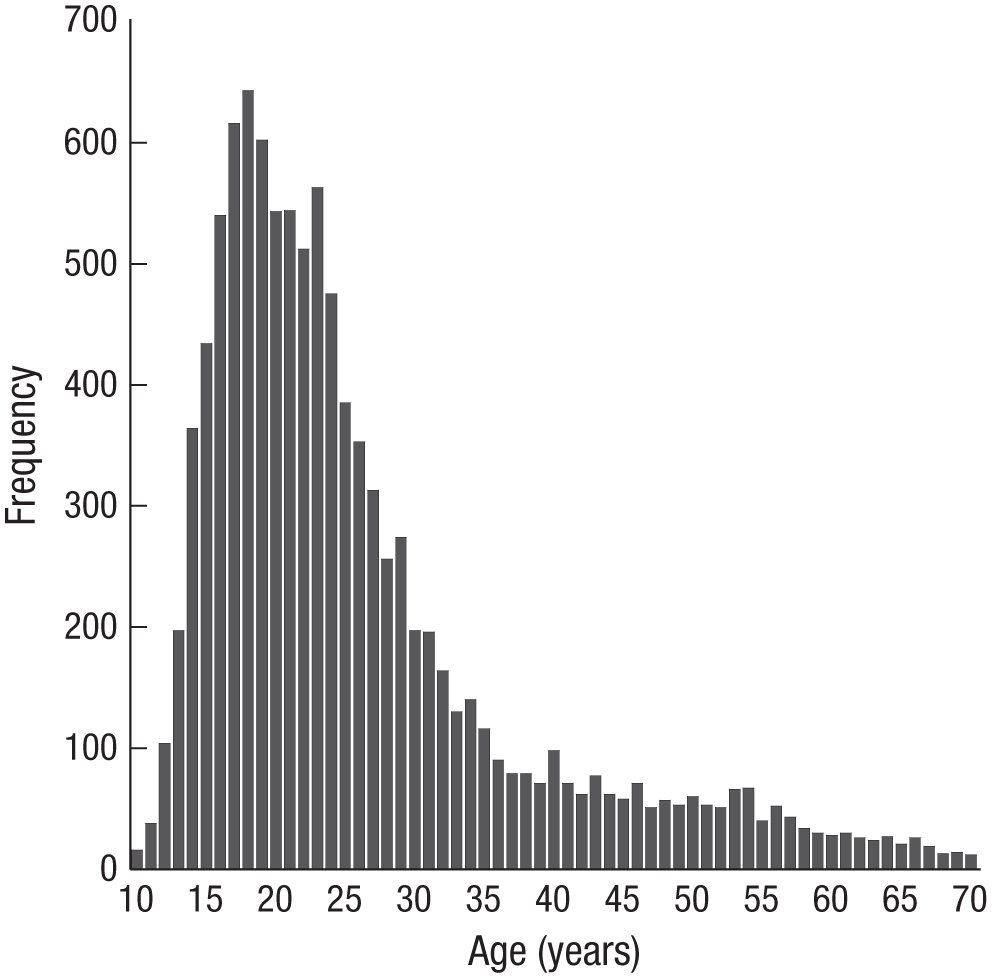
Histogram showing the age distribution of participants.
Task and procedure
Participants visited TestMyBrain.org between March 2014 and September 2014. TestMyBrain.org provides feedback on performance relative to other users. The gradCPT was presented on its home page as a “Continuous Concentration task.” TestMyBrain.org receives traffic mostly from social-networking sites and search engines (Germine et al., 2012). The gradCPT is one test out of several on TestMyBrain.org (others include face recognition and working memory). Participants are free to complete one or more of these tasks; for some participants, the gradCPT was the first or only test they completed, while others may have completed other tasks first. Single-experiment studies on TestMyBrain.org are kept brief (< 10 min) in order to maintain a balance between task completion, participation, and test length. Given the demanding nature of continuous performance tasks, the Web-based gradCPT used a shortened version (4 min vs. 8 min) of the continuous go/no-go task originally reported in Esterman et al. (2013). This test length was chosen because it was sufficiently short that participant attrition rates would be comparable with those of other experiments on the site. The concern with participants dropping out during testing regards a selection bias in which individuals with poorer sustained-attention ability would be less likely to complete the experiment. This led to a total experiment time of approximately 7 min from consent to debriefing.
The stimuli consisted of 20 round (200-pixel diameter), gray-scale photographs of 10 city scenes and 10 mountain scenes. The same trial sequence was used for every participant. This approach is regularly used in individual-differences research (Carlson & Moses, 2001) to eliminate the order of stimuli as a potential source of systematic variation across age groups. The order of scenes was pseudorandom (90% cities and 10% mountains) in that identical scenes did not appear on consecutive trials. Each scene gradually transitioned to the next scene every 800 ms using a linear pixel-by-pixel interpolation, for a total of 299 trials across the 4 min of testing. Additionally, the block began with a fully opaque scrambled city image (i.e., noise image) from which the first trial image transitioned, and the same scrambled image was used at the very end of the block for the last trial image to transition into. Figure 2 illustrates the linear interpolation utilized. At the start of every new trial, the incoming scene had an image opacity of 0% (i.e., it was 100% transparent) and transitioned to 100% opacity by the end of the 800 ms, at which point it fully replaced the previous image. On the following trial, this scene then transitioned from 100% opacity back to 0% opacity while a new image transitioned into view. Thus, on all trials, as the first image faded in clarity, it was gradually replaced by a new image, which transitioned into view and faded into the old image. The new image increased in clarity at the same rate as the old image decreased in clarity. Participants were instructed to press the space bar for city scenes (267 trials) and withhold a response for mountain scenes (32 trials). Thus, the task instructions emphasized accuracy in responses while the continuous nature of the task imposed a limited time within which participants could respond.
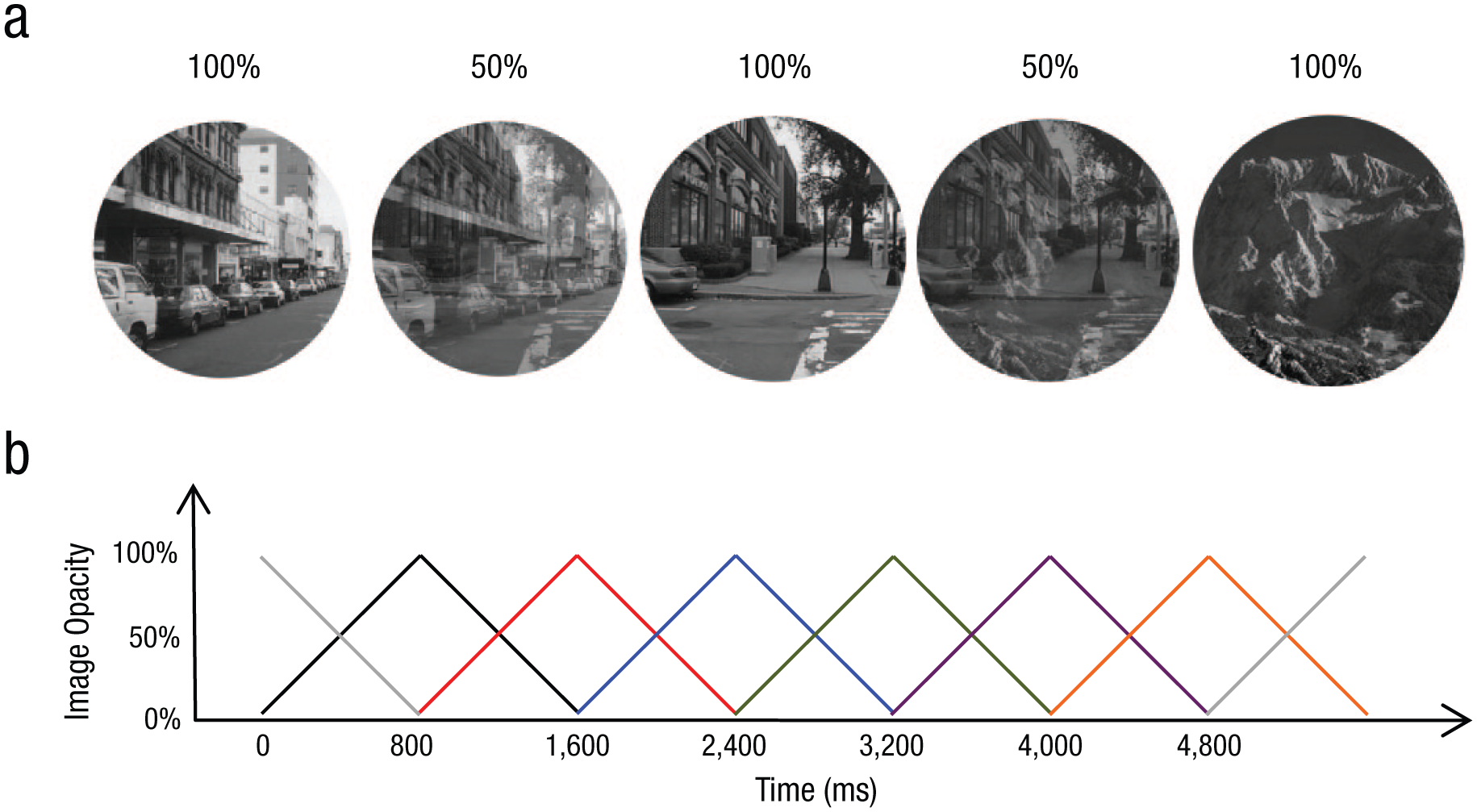
Paradigm of the gradual-onset continuous performance task. The example images (a) show three scenes over two full trials at full (100%) and mixed (50%) opacity. On each trial, a scene appeared at 100% opacity and faded to 0% opacity while a new scene faded in from 0% to 100% opacity. The graph (b) illustrates the linear transition in image opacity from one image to the next. Each colored triangle represents the transition for a single image over time.
Before starting the gradCPT, participants gave informed consent according to the guidelines set by the Committee on the Use of Human Subjects at Harvard University and the Wellesley College Institutional Review Board. Participants were then asked to complete a voluntary demographic survey asking about their age, gender, native language, and ethnicity. Comprehensive instructions as well as three practice sessions (30 s each) were then provided to familiarize participants with the scenes and the task so that it could be completed without assistance from an experimenter. After completing 4 min of the gradCPT, participants were asked if they had cheated or if any problems occurred during the task, and then they were provided with performance feedback. This feedback consisted of a personal score, which was the percentage of correct withholdings to mountains, as well as how this score compared with the average participant.
Analyses
To calculate reaction time measures, we first used an iterative algorithm to assign button presses to specific trials. Button presses were logged relative to the beginning of the experimental block, following the methodology outlined in Esterman et al. (2013). Reaction times were then defined relative to the beginning of each image transition, such that a reaction time of 800 ms indicated a response when the current trial image reached 100% opacity, whereas shorter reaction times indicated that the current trial image was still transitioning when a response was made. All responses were logged throughout the experiment, and the algorithm set a limit of 1,360 ms before a response time was assigned to the next trial (see the Supplemental Material available online for more details).
Using correct responses to cities, we computed the mean reaction time (in milliseconds) and reaction time variability. Reaction time variability was calculated using the coefficient of variation (i.e., the standard deviation of reaction times divided by the mean reaction time for each participant). Commission error rate (responding to a mountain scene) and omission error rate (failing to respond to a city scene) were then calculated.
Additionally, taking a signal detection approach with the hits (correct omissions to mountains) and false alarms (incorrect omissions to cities), we computed d′ (a measure of discrimination ability) and criterion (a measure of strategy or willingness to respond in the case of uncertainty). We used standard procedures to correct for cases in which hit rates were 100% or false alarm rates were 0%, with one-half error deducted or added on the basis of the number of target or nontarget trials presented, respectively (see the Supplemental Material). Factor analyses were completed with a direct oblimin rotation, which uses an oblique rotation that is more accurate than an orthogonal rotation and provides a more optimal solution when the resulting factors are correlated. Commission and omission errors were not used in the factor analysis because they were nonindependent of the signal detection variables.
An additional feature of the gradCPT is that when analyzing the results, the duration of the task is split into states of low and high variability (“in the zone” vs. “out of the zone”). Although this version of the task was abbreviated to 4 min (and thus limited in- vs. out-of-the-zone epochs to 2 min each), we nonetheless explored performance during each of these states using variance-time-course (VTC) analysis (Esterman et al., 2013). Specifically, the reaction times for correct responses were converted to absolute z scores, and values for correct omissions and trials with errors were linearly interpolated using the average of the two neighboring trials. The VTC was then smoothed using a Gaussian kernel with a 20-trial window and an 8-s full width at half maximum. From the smoothed VTC for each participant, two attentional states were defined using a median split to separate in-the-zone and out-of-the-zone epochs. Thus, in-the-zone epochs contained trials on which reaction times were closest to the mean of the run, whereas out-of-the-zone epochs contained trials with the most deviant reaction times, including both the fastest and the slowest responses.
Results
The following analyses focus on four dependent measures: mean reaction time, reaction time variability, d′, and criterion (see Fig. S3 in the Supplemental Material for commission and omission error results).
Reliability of dependent measures
Reliability measurements were obtained using Spearman-Brown-corrected split-half correlations comparing the averaged performance of the 1st and 3rd minutes with the averaged performance of the 2nd and 4th minutes. All four dependent measures showed acceptable to high internal reliability: .94 for mean reaction time, .90 for reaction time variability, .78 for d′, and .80 for criterion.
Comparison of lab-based and Web-based measures
To test whether the gradCPT yielded similar results when performed over the Web as it does in the lab, we selected the subset of 6,290 participants between the ages of 18 to 34 years and compared their performance to the first 4 min of gradCPT data in 17 age-matched participants previously collected in a controlled laboratory setting (Esterman et al., 2013; see Fig. S1 in the Supplemental Material). Independent samples t tests assuming unequal variance showed no difference in accuracy (d′), criterion, or commission and omission error rates (ps ≥ .29). Mean reaction time was slightly slower in the present sample (mean difference = 81 ms), t(6305) = 5.43, p < .001, d = 1.20, which may be because of technical or hardware differences in response collection over the Internet versus in the laboratory (McGraw, Tew, & Williams, 2000). Additionally, reaction time variability was lower in the Web-based than in the lab-based sample (mean difference = −0.029), t(6305) = 2.44, p = .01, d = 0.56. Notably, however, the differences in reaction time measures did not accompany differences in accuracy (d′, commission and omission error rates) or criterion. This supports alignment between the performance of young adults in controlled laboratory settings with the Web-based participants in the present study and is consistent with the findings of previous Web-based studies showing comparable performance levels with lab-based studies of perception, attention, and working memory (Germine et al., 2012; Halberda, Ly, Wilmer, Naiman, & Germine, 2012; Hartshorne & Germine, 2015; McGraw et al., 2000).
Modeling life-span changes in sustained-attention performance
To achieve our goal of understanding the mechanisms of sustained-attention changes across the life span, we separately modeled each of our four key sustained-attention measures across the 10- to 70-year age range (see Fig. 3). At a descriptive level, one could expect three types of processes across time: a growth in capability, a plateau where capability is maintained, and declines in capability. To uncover these possible changes, we modeled the data using the average data for each age. Because the age distribution of the participants was skewed, we further binned the youngest and oldest participants into age bins, with the constraint that at least 100 participants fall into each bin. For all ages between 12 and 33, there were enough participants that each age was binned separately. To obtain at least 100 participants for the other ages, we binned ages as follows: 10–12, 33–34, 35–36, 37–38, 39–40, 41–43, 44–46, 47–49, 50–53, 54–57, 58–61, and 62–70 years. The mean of each age bin was then used in the following analyses. Very similar results were found when the age means were modeled without binning and with the raw individual participant data (see the Supplemental Material). We first applied a standard polynomial approach previously used to model life-span changes in sustained attention (McAvinue et al., 2012). Polynomial modeling revealed significant nonlinearity for all four measures (cubic trend ps ≤ .0003).
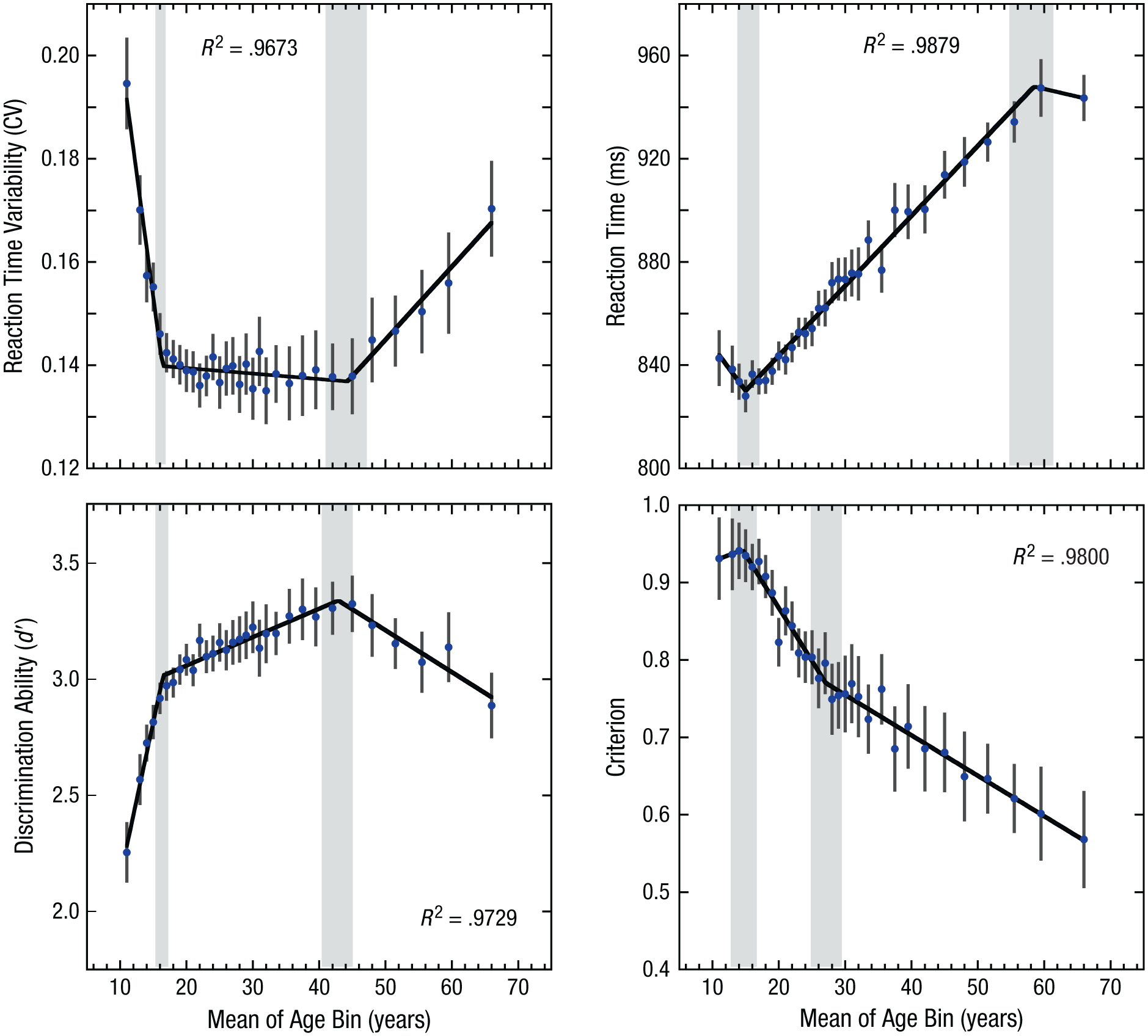
Changes in sustained-attention performance for each of the age bins. Results are shown separately for variability in reaction time, mean reaction time, discrimination ability (d′), and criterion. Error bars show 95% confidence intervals. Solid lines show the best-fitting functions from the hierarchical regression analysis. Gray bars show the 95% confidence intervals for the estimated breakpoints. Reaction time variability is defined as the coefficient of variation (CV; the standard deviation divided by the mean reaction time).
To quantify the nature and timing of performance changes across the life span, we next employed hierarchical regression analyses utilizing segmented linear functions (i.e., piecewise regression). The benefit of these segmented linear functions is that they allow one to capture multiple combinations of changes (growth, plateau, decay) with no assumption of symmetry such as that implicitly assumed when utilizing quadratic functions in trend analyses. Moreover, the breakpoint between the two linear segments and the confidence intervals around this breakpoint provide a direct estimate of a transition zone, or an age range in which transitions are most likely to occur. Segmented linear functions have been successfully used to model life-span changes in white matter tract integrity (Yeatman, Wandell, & Mezer, 2014). However, to date, such models have not been applied to life-span changes in cognition. In the present study, linear functions were compared with a two-phase segmented linear function with one transition (breakpoint). If the two-phase model provided a significantly better fit, we then compared two-phase functions with three-phase functions (one vs. two transitions).
Changes in ability across the life span
As Figure 3 shows, d′ and reaction time variability exhibited a similar pattern across the life span, with both being fit best by three-phase linear functions (see Table 1 for model comparisons). Both measures showed rapid development in sustained-attention ability between 10 and 16 years of age, then a period of relative stability until approximately 43 years of age, and finally a decline in ability across old age. In particular, the estimated breakpoints for d′ occurred at 16.5 years (95% confidence interval, or CI = [15.9, 17.1]) and 42.9 years (95% CI = [40.3, 45.5]), which are notably similar to the reaction time variability breakpoints at 16.4 years (95% CI = [15.9, 16.9]) and 44.3 years (95% CI = [41.2, 47.4]).
Results of Hierarchical Regressions Used for Model Selection

Note: Each analysis determined whether the second, more complicated model (the alternative) explained significantly more variance than the first, null model.
Examining the slopes, or rate of change during each phase, we found that discrimination ability (d′) rapidly improved between 10 and 16 years (0.13 per year, 95% CI = [0.11, 0.16]). This was followed by a period of relative stability with modest but significant increases in discrimination ability between 17 and 43 years of age (0.012 per year, 95% CI = [0.0097, 0.015]), t(26) = 9.71, p < .0001. At approximately 43 years, d′ peaked and then gradually declined (−0.018 per year, 95% CI = [−0.023, −0.013]). Similarly, for reaction time variability, the slope patterns highlighted a rapid performance improvement (decrease in response variability) between 10 and 16 years of age (decrease in coefficient of variation by −0.01 ms per year, 95% CI = [−0.011, −0.008]). This was followed by a period of stability between 16 and 44 years of age, with small but unreliable decreases in reaction time variability (−0.0001 per year, 95% CI = [−0.0003, 0.00006]), t(26) = 1.35, p = .19. Performance then declined beyond age 44, with reaction time variability increasing at a rate of 0.0014 per year (95% CI = [0.001, 0.002]). For both measures, the estimated slopes during older adulthood were less than one fifth those observed during the childhood development period, which suggests that while participants showed a decline in task ability as they got older, the rate of decline observed was not nearly as great as the rate of increase in task ability seen during development. Notably, beyond the rapid development observed between 10- and 16-year-olds, the results show a continued though slowed increase in discrimination ability (d′), with maximum sustained-attention ability occurring far later in life in the mid-40s. This result highlights a unique life-span trajectory for sustained-attention ability compared with other cognitive abilities, such as fluid intelligence, which has been shown to peak in the mid-20s (Craik & Bialystok, 2006; Hartshorne & Germine, 2015).
Changes in strategy across the life span
The criterion and mean reaction time measures showed markedly distinct life-span patterns from d′ and reaction time variability (see Fig. 3), though these measures were also best fit by the three-phase linear functions (see Table 1 for model comparisons). In particular, mean reaction time and criterion showed a period of rapid development followed by two distinct but monotonically changing shifts in strategy across the latter years. For mean reaction time, the estimated breakpoints occurred at 15.0 years (95% CI = [14.0, 16.6]) and 58.5 years (95% CI = [55.1, 61.9]). The slope patterns highlighted a trend toward a speeding of reaction time between 10 and 15 years of age (at −3.45 ms per year, 95% CI = [−7.1, 0.1]), t(26) = 1.97, p = .06. A reversal was observed following this period, with reaction times slowing between 15 and 58 years of age at a rate of 2.7 ms per year (95% CI = [2.6, 2.9]). After the second transition point, reaction times flattened, and the slope estimate showed an unreliable speeding rate of −0.6 ms per year (95% CI = [−2.5, 1.3]), t(26) = 0.60, p = .52. The slowing of reaction times from 16 to 29 years of age likely reflects a strategic shift, since choice reaction time has been shown to speed up in this range (Williams, Hultsch, Strauss, Hunter, & Tannock, 2005). The monotonic increase in reaction times observed from 30 to 58 years of age could also reflect a strategic shift, though it is also consistent with a general age-related slowing of reaction times (Ratcliff, Thapar, & McKoon, 2001). The change in slope observed beyond the second breakpoint may be related to the experiment’s design and the implicit reaction time limitations inherent in continuous performance tasks (i.e., reaction time ceiling effect). Specifically, the algorithm used to assign reaction times has an implicit maximum of 1,360 ms before responses are assigned to the next trial. While no individual participant showed mean reaction times close to or above this ceiling (see the Supplemental Material), it is possible that more reaction times above this maximum occurred in participants over 58 years of age, leading to an increase in reaction time variability and a flattening of the calculated mean reaction times.
The pattern of change observed for the criterion measure showed similarities to that observed for the reaction time measure. First, there was a slight upward slope toward a greater bias to respond (a more impulsive strategy) in the first phase. The initial breakpoint was at 14.5 years (95% CI = [13.0, 16.9]), though this slope was not significantly greater than zero (slope = 0.003, 95% CI = [−0.01, 0.02]), t(26) = 0.43, p = .67.
After the initial transition point, which indicates the least cautious approach to the task, a change in the pattern occurred, and (similar to the reaction time measure) a monotonic trend was observed across the rest of the age groups. This trend suggests a continuous shift toward a more cautious approach to the task, as reflected in a reduced bias to respond on a given trial. In contrast to the other performance measures, the second transition point was seen earlier at 27.1 years (95% CI = [24.4, 29.8]). Between 15 and 27 years, a reliable shift toward a more conservative approach was observed, with a slope of −0.01 per year (95% CI = [−0.02, −0.01]). Following the second transition zone, the rate of change slowed but continued toward a decreased bias to respond during a given trial (slope = −0.0052 per year, 95% CI = [−0.006, −0.004]).
Collectively, these measures indicate that beyond approximately 15 years of age, participants show a gradual slowing of reaction times and a more careful approach to the task with age. This is in line with previous studies demonstrating more impulsivity in adolescence than in adulthood (Galvan et al., 2006), and the observed slowing of reaction time from 15 to 30 years old when reaction time abilities are typically still improving (Williams et al., 2005) is suggestive of strategic slowing. In contrast, the slowing of reaction times from ages 30 to 58 likely reflects a combination of decreasing simple target-detection reaction time (Ratcliff et al., 2001) and a more cautious strategy, which has previously been demonstrated in older adults (Deakin, Aitken, Robbins, & Sahakian, 2004).
To further investigate and provide converging evidence for this strategic shift across the life span, we calculated the degree to which individuals slowed down responses following an incorrect response. This phenomenon, referred to as posterror slowing, is thought to reflect error monitoring (Dutilh et al., 2012). To better isolate posterror slowing, we regressed out the mean reaction time for correct responses immediately preceding erroneous responses from the mean reaction times for correct responses immediately following erroneous responses. As Figure 4 shows, posterror slowing showed a very similar life-span pattern to criterion and reaction time, consistent with the interpretation that the three measures are driven by strategic shifts toward cautiousness. The pattern of posterror-slowing data was best fit by a two-phase segmented function (see Table 1). Similar to the criterion and reaction time results, the parameters of the two-phase model showed a transition zone at 15.0 years (95% CI = [13.3, 16.8]), which represents the lowest error monitoring in early adolescence. As with the criterion measure, a negative but unreliable slope was observed in the first phase (slope = −4.36, 95% CI = [−9.9, 1.1]), t(28) = −1.62, p = .12. However, following the transition zone, there was a consistent increase in posterror slowing of 3.5 ms per year (95% CI = [3.2, 3.7]). This result suggests a gradual shift toward greater error monitoring across the life span above and beyond that explained by slower reaction times due to changes in central nervous functioning in older adults (Ratcliff et al., 2001). The increase in posterror slowing is also consistent with previous self-reports by older participants of less mind wandering during task completion and greater intrinsic motivation to perform well on similar tasks (Staub et al., 2014).
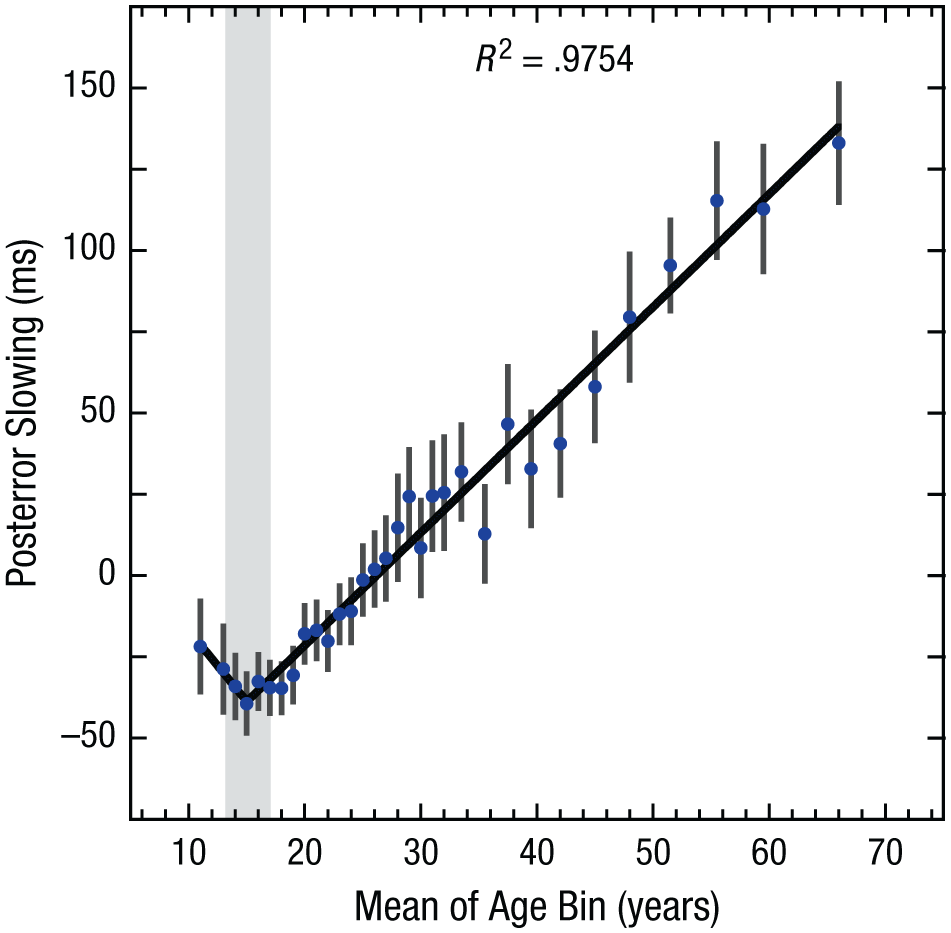
Posterror slowing for each of the age bins. Error bars show 95% confidence intervals, and the solid line represents the best-fitting two-phase segmented linear function. The gray bar shows the 95% confidence interval for the estimated breakpoint.
Collectively, then, the mean reaction time, criterion, and posterror slowing measurements all showed a monotonic trend across participants 15 years and older, which represented a strategic shift toward a slower, more cautious approach to the task that diverged significantly from the pattern observed in the measures of d′ and reaction time variability.
Factor analysis of latent measures in sustained-attention performance
To further confirm the dissociation between sustained-attention ability and strategy, we conducted exploratory factor analyses on the four primary variables (mean reaction time, reaction time variability, d′, criterion). Two factors emerged with eigenvalues over Kaiser’s criterion of 1: one with high loadings for reaction time variability and d′, and the other with high loadings for mean reaction time and criterion (see Fig. 5a and Table 2). Together these components explained 85.21% of the variance. Given the transition zones detected in sustained-attention ability across the life span, we also conducted the factor analysis for each age group based on the breakpoints observed for reaction time variability and d′ (10–16, 17–43, 44–70). The same variables cluster together across all three age ranges, which provides further support for the hypothesis that two latent variables relate sustained-attention strategy and ability to task performance and changes in task performance across the life span.
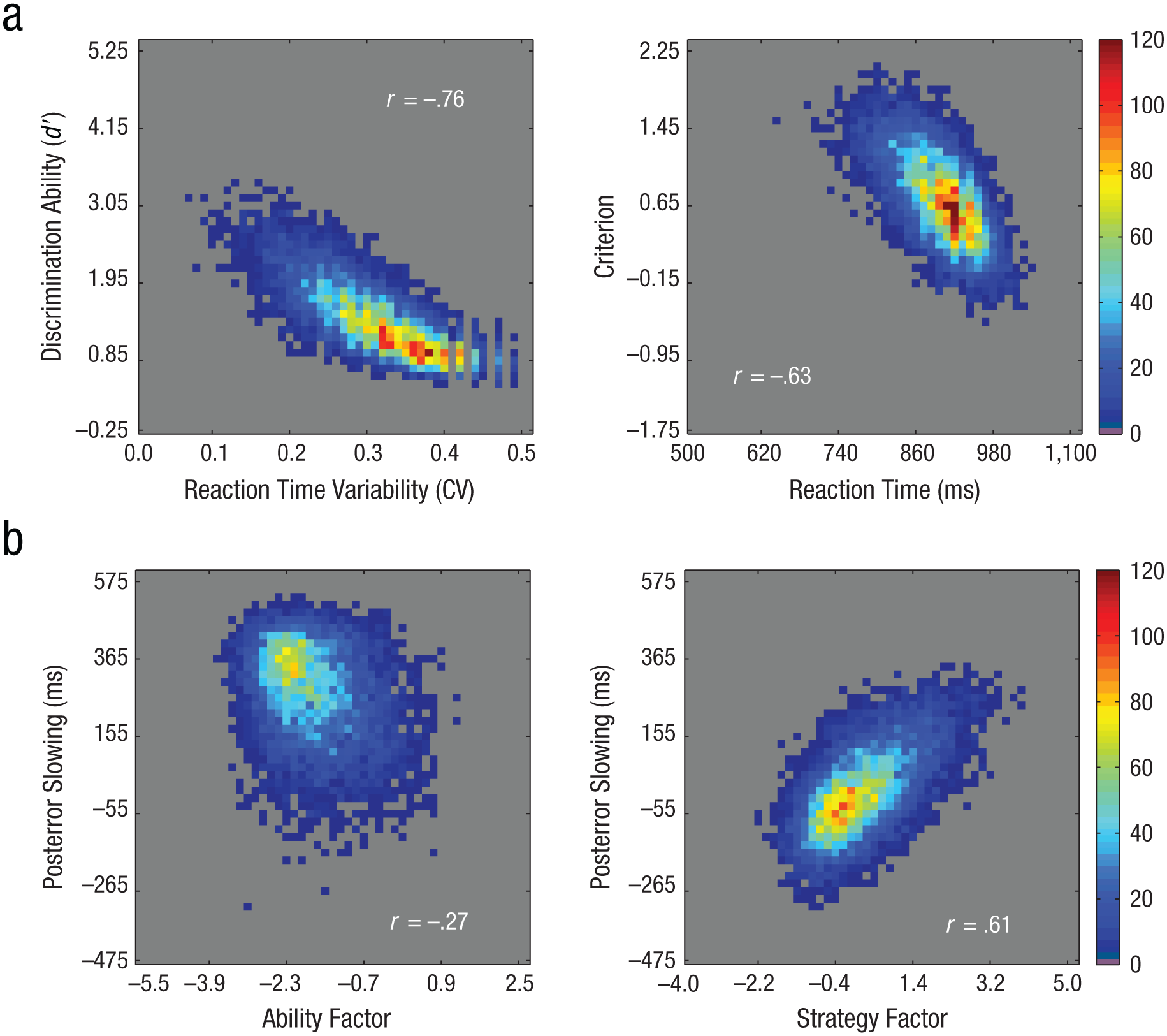
Density scatterplots showing correlations across all participants for measures of sustained-attention ability and sustained-attention strategy. The top row (a) shows correlations between measures related to ability (left) and strategy (right). The bottom row (b) illustrates the relationships between posterror slowing and the calculated factor score related to ability (left) and the calculated factor score related to strategy (right). For all plots, the colors indicate the number of participants represented at a given location. Reaction time variability is defined as the coefficient of variation (CV; the standard deviation divided by the mean reaction time).
Loadings of the Primary Variables on the Two Factors Identified in the Factor Analysis
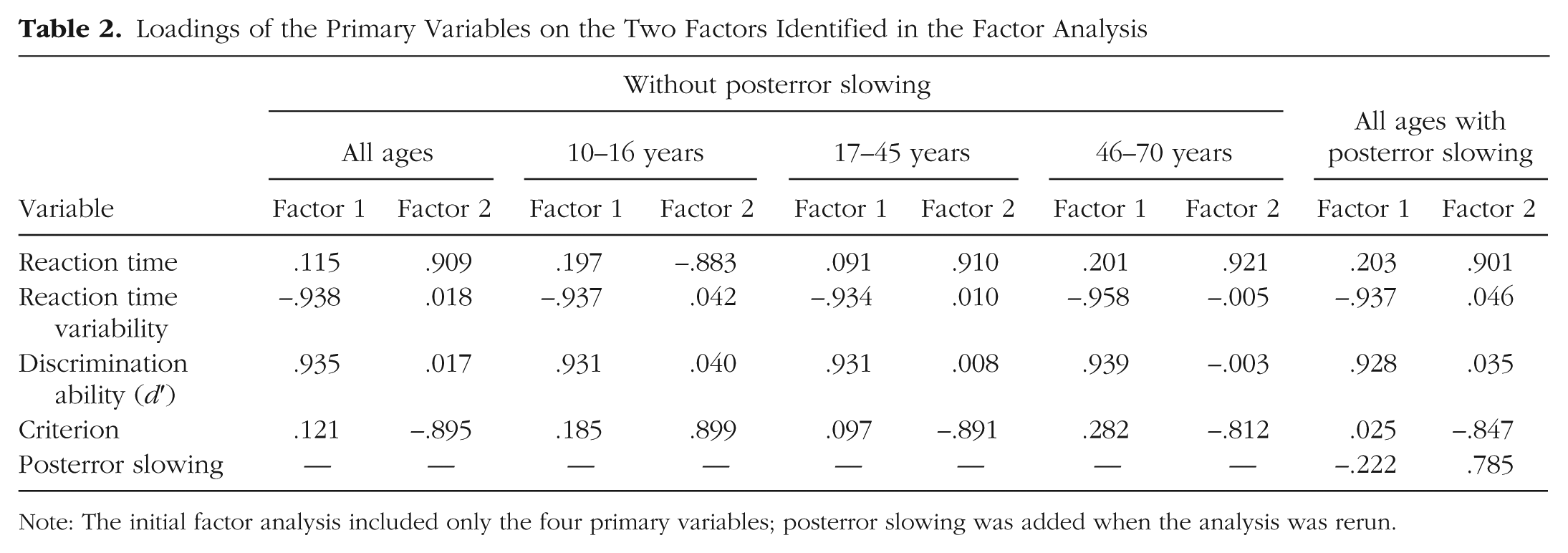
Note: The initial factor analysis included only the four primary variables; posterror slowing was added when the analysis was rerun.
Finally, rerunning the factor analysis when including posterror slowing showed very similar results, with posterror slowing clustering with reaction time and criterion, while d′ and reaction time variability loaded onto a separate component (Fig. 5b). Together, these results further confirmed that d′ and reaction time variability both measured a similar latent variable (i.e., sustained-attention ability), which is dissociable from the latent variable that criterion, reaction time, and posterror slowing measured (i.e., sustained-attention strategy).
Comparison of life-span changes in and out of the zone
In our final analysis, we explored whether the observed age-related changes in the task as a whole were consistent across states of high and low attentional stability (in and out of the zone, respectively). During in-the-zone and out-of-the-zone epochs, we compared two variants of the three-phase segmented linear functions used in the previous analyses. In this analysis, we modeled a simple main effect of zone as a shift in the intercept parameter (i.e., baseline shift) across the two zone conditions, while an interaction was reflected in a shift in any of the other five parameters (two breakpoints, three slopes). Thus, the null model was one in which the intercept parameters were free to vary across the two zone conditions, while the other five parameters were shared. In the alternative model, all six parameters were free to vary across the two zone conditions. Qualitatively, the developmental trajectories were similar across attentional states, with overlapping transition points and the same slope directions when participants were in the zone and out of the zone. However, results of the model comparison showed that only the criterion measure was not significantly better fit by the alternative model, F(5, 52) = 1.42, p = .23, which indicates that the change in criterion when participants were in the zone versus out of the zone was well modeled by an additive shift of −0.095 for all ages (see Fig. 6 and Table 3). In contrast, all three of the remaining performance variables were better fit by the alternative model, which indicates some differences in life-span trends across the zone conditions—mean reaction time: F(5, 52) = 3.83, p = .005; reaction time variability: F(5, 52) = 27.50, p < .0001; d′: F(5, 52) = 3.32, p = .01.
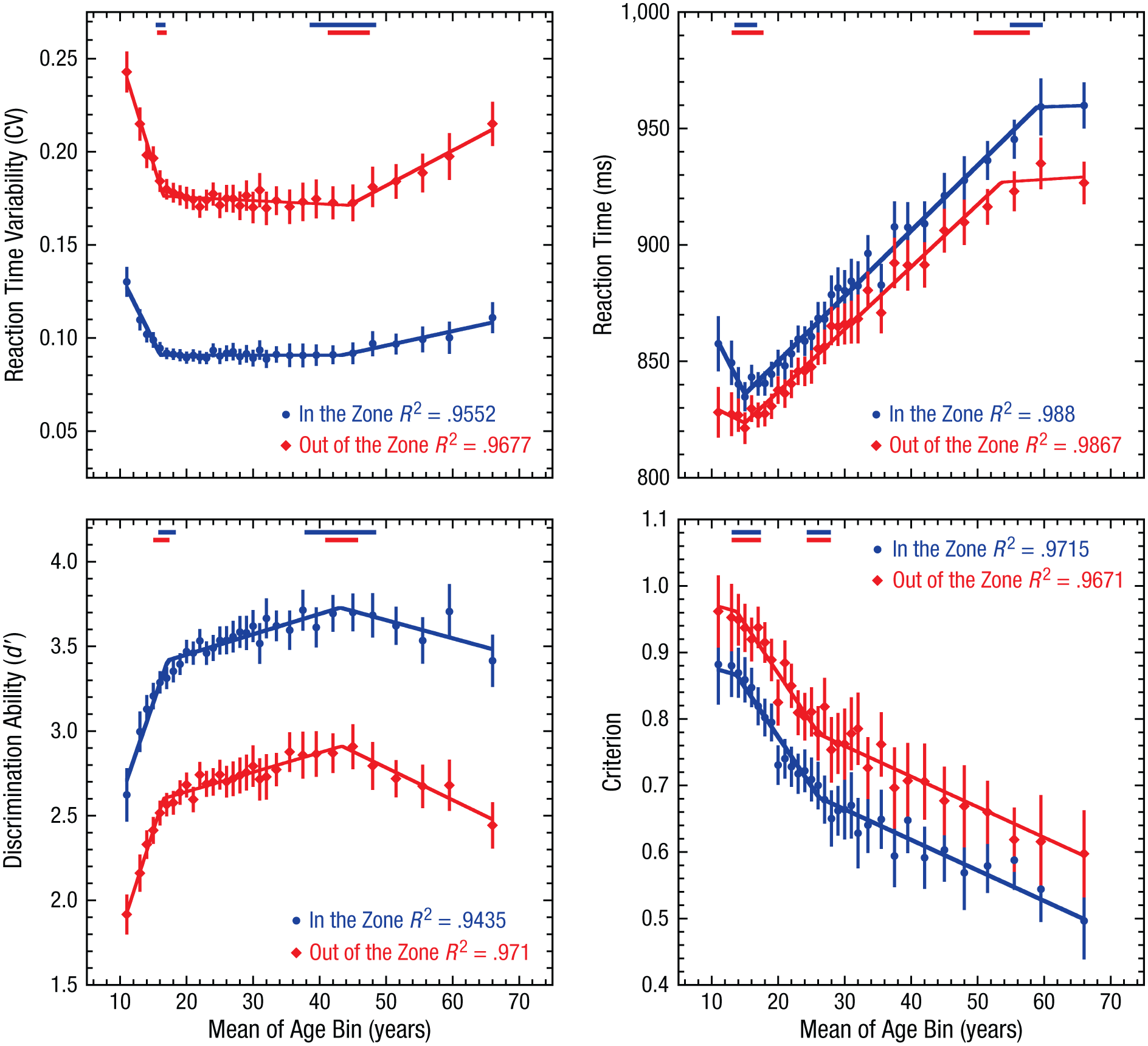
Changes in sustained-attention performance for each age bin and zone condition. Results are shown separately for reaction time variability, mean reaction time, discrimination ability (d′), and criterion. Error bars show 95% confidence intervals. Blue and red lines show the best-fitting functions from the hierarchical regression analysis for each condition. Red and blue dashes at the top of each graph show the 95% confidence intervals for the estimated breakpoints. Reaction time variability is defined as the coefficient of variation (CV; the standard deviation divided by the mean reaction time).
Parameter Estimates for Performance Measures for the Two Zone Conditions
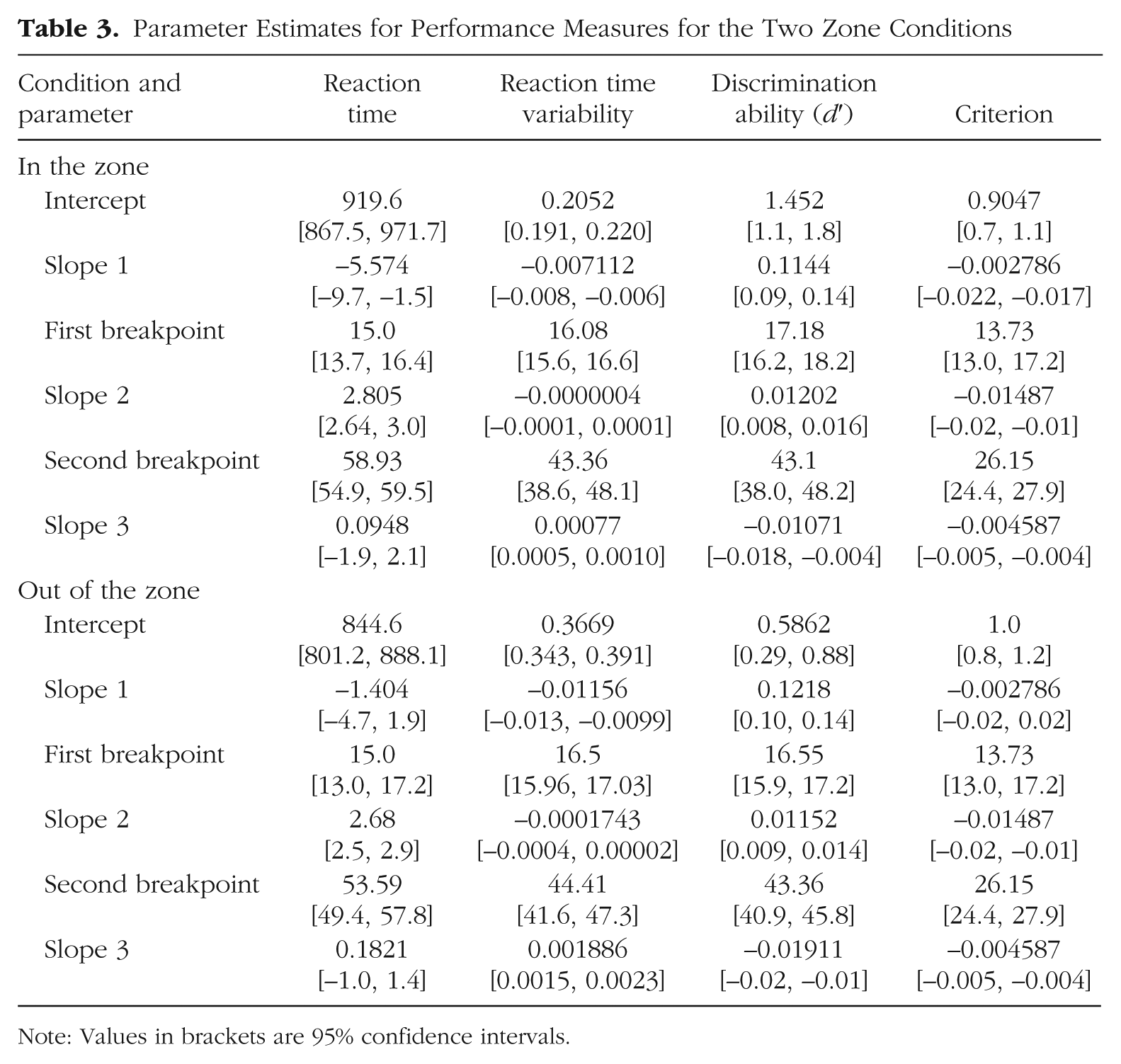
Note: Values in brackets are 95% confidence intervals.
Figure 6 and Table 3 show that while these three performance variables had similar trajectories across the age ranges tested, the most prominent differences occurred during the development and aging phases. In particular, the d′ slope for participants over the age of 43 years increased by approximately 78% when they were out of the zone compared with in the zone, which indicates that age-related declines in task sensitivity are more prominent during times of greater attentional instability. Similarly, a twofold increase was seen in the slope parameter for reaction time variability in participants over the age of 44 years, which indicates that age-related increases in reaction time variability were more prominent when participants were out of than in the zone. Additionally, the differences in the initial slope parameters for reaction time and reaction time variability suggest that differential developmental changes may occur in response generation across periods of low and high attentional stability.
Discussion
In the present study, we used an unprecedented sample size and novel methods to explore sustained attention across a 60-year life span. We demonstrated that two distinct underlying processes contribute to sustained-attention performance: (a) the ability to maintain consistent and accurate performance and (b) the strategy of going faster with a bias to respond or slower with a bias to withhold.
The modeling results show that these two underlying factors have differential life-span trajectories with critical transitional phases. All performance measures suggest an initial period of development with a rapid increase in task ability and a shift toward a faster, less cautious strategy across the youngest age groups, with early transition zones in adolescence around 14 to 17 years of age. However, beyond this initial transition period, important dissociations in life-span trajectories were observed, with task-ability parameters (d′ and reaction time variability) showing evidence for continued improvement through adulthood and marked decreases in ability beyond 43 years of age. In contrast, adult participants showed a monotonic trend in strategy toward a slower and more conservative approach to the task. Different strategy life phases were best characterized by changes in the magnitude of the slope parameters (i.e., the rate of change) but not in the direction. This shift in strategy was further supported by age-related changes in posterror slowing, which again highlights a monotonic shift toward increased error monitoring beyond age 15. While simple reaction times are known to slow in older adults because of primary sensorimotor changes (Ratcliff et al., 2001), the continuous nature of the changes observed in reaction time and criterion measures from as young as 16 years of age, coupled with the same pattern observed in posterror slowing when we controlled for overall reaction time, suggest that the changes across the full age range are best accounted for by changes in task strategy. This does not discount any role for sensorimotor changes in influencing these measures. One intriguing possibility is that the relative influence of such factors increases with age and ultimately limits the benefit of strategic slowing in the older participants with regard to task accuracy.
The dissociable life-span trajectories between sustained-attention strategy and ability were also evident both during participants’ relative best (in the zone) and worst (out of the zone) periods of performance. The most notable difference between attentional states was that the decline in the ability parameters after approximately the age of 43 was markedly steeper during out-of-the-zone than in-the-zone periods. Such periods are thought to reflect the most taxing intervals of the task (Esterman, Rosenberg, & Noonan, 2014), and they suggest more pronounced age-related decline in more challenging tasks. The results of factor analyses further demonstrated that the dissociation between strategy and ability was evident across the sample as a whole, as well as within each of the three specific age-ranges tested.
The use of segmented linear regression analyses points to important regions of time that we have labeled transition zones. While the class of functions we utilized here defines a specific breakpoint for each zone, changes are likely to occur gradually over a period of time in individuals. One particularly useful aspect of these functions is that they provide explicit estimates of transition periods in the parameter estimates which, when interpreted with confidence intervals, provide a likely time window across which transition zones may be expected and developmentally appropriate. Such normative data could provide a basis for potentially revealing abnormal life-span trajectories, such as those associated with developmental disorders (e.g., attention-deficit/hyperactivity disorder) and pathological aging (e.g., dementia).
More broadly, the results also suggest that sustained-attention ability peaks far later in life than other visual and cognitive-processing mechanisms. Specifically, the results show that despite a slowing in growth during adulthood, sustained-attention ability (i.e., d′) peaks at 43 years of age. This is far later than other cognitive abilities, such as those related to fluid intelligence. Studies have shown that sensory and cognitive-processing abilities, including visual processing speed and working memory, peak before age 30 and decline thereafter (Baltes & Lindenberger, 1997; Germine, Duchaine, & Nakayama, 2011; Halberda et al., 2012; Hartshorne & Germine, 2015; Owsley, 2011). Conversely, we found that sustained-attention ability shows an earlier peak than has been measured for simple knowledge accumulation related to crystallized intelligence (Craik & Bialystok, 2006; Hartshorne & Germine, 2015). This suggests that sustained-attention ability represents an important, distinct mechanism that contributes to an individual’s ability to process information and interact with the world. While young adults may surpass people of other ages in the speed and flexibility of information processing, and older adults may possess the most stored knowledge regarding the world, we found that middle-aged adults have the greatest capacity to remain attentive. One explanation for sustained-attention ability peaking at age 43 is that attention is highly trainable (DeGutis & Van Vleet, 2010), and practice focusing attention throughout adulthood may further hone this skill.
Sustained attention peaking in one’s 40s is also consistent with recent studies of white matter and prefrontal cortex integrity across the life span (Hedden & Gabrieli, 2004; Yeatman et al., 2014). One recent study has shown asymmetrical maturation and degeneration processes in frontal white matter tract integrity across the life span, which qualitatively matches the pattern observed in our ability factor (Yeatman et al., 2014). Sustained attention activates a large-scale network of cortical and subcortical regions, including areas in the frontal lobe (Esterman et al., 2013). Thus, changes in frontal white matter tract integrity over time may significantly affect many cognitive functions, including sustained-attention ability.
There are several limitations of the present study that future studies may address. First, we used a cross-sectional design, which prevents assessment of individual life-span trajectories. Longitudinal studies would provide more useful data for investigating the pattern of changes within these transition zones. Second, participants were free to potentially complete multiple experiments before participating in the current experiment. This most likely introduced interparticipant variability rather than bias across the age ranges, but future studies may wish to account for this potential issue. Third, the present sample was skewed, with a relative undersampling of the youngest and oldest participants. As the modeling results were consistent regardless of which data were used (binned or unbinned age means or individual data), this is unlikely to have greatly affected the results: The smallest binned age group still had 122 participants. A related issue is the potential underestimation of age-related declines. Given the use of Internet-based volunteers, it is possible that there was a selection bias in the older participants toward higher-functioning older adults. While other TestMyBrain.org studies have replicated life-span trends in cognition compared with traditionally collected and nationally representative U.S. samples (Hartshorne & Germine, 2015), future research obtaining greater demographic and educational details, along with extending the sample age range, would help to validate the current findings.
Despite these limitations, the current results provide important information that will help to fill the gaps in understanding normative changes in sustained attention across the life span. Utilizing factor analyses and a novel regression approach that highlighted important transition periods in sustained-attention ability and the strategies utilized by participants, we provided a new foundation for future research on sustained attention as well as studies on a range of neurocognitive functions that depend on sustained attention.
Footnotes
Acknowledgements
The contents of this article do not represent the views of the Department of Veterans Affairs or the U.S. government.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was partially supported by the Department of Veterans Affairs. F. C. Fortenbaugh has an Advanced Geriatric Fellowship from the Department of Veterans Affairs. M. Esterman has a Career Development award from the Department of Veterans Affairs Clinical Sciences Research and Development Service (1IK2CX000706-01A2). J. B. Wilmer has a Brachman Hoffman Fellowship.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.