
Abstract
Jealousy may have evolved to protect valuable social bonds from interlopers, but some researchers have suggested that it is linked to self-awareness and theory of mind, leading to claims that it is unique to humans. We presented dogs (N = 18; 11 females; age: M = 4.6 years, SD = 1.9) with situations in which they could observe an out-of-sight social interaction between their owner and a fake dog or between their owner and a fleece cylinder. We found evidence for three signatures of jealous behavior in dogs: (a) Jealousy emerged only when the dog’s owner interacted with a perceived social rival, (b) it occurred as a consequence of that interaction and not because of the mere presence of a conspecific, and (c) it emerged even for an out-of-sight interaction between the dog’s owner and a social rival. These results support claims that dogs display jealous behavior, and they provide the first evidence that dogs can mentally represent jealousy-inducing social interactions.
In humans, jealousy may have evolved to protect valuable social bonds that confer emotional or material benefits to an individual (White, 1991) and can emerge even as a consequence of imagined interactions between two people (Leary, 2003; Lewis, 2002). It has been argued that jealousy, as a secondary emotion, may require a concept of self, which allows the individual to perceive a threat to their bond with a social partner (Lewis, 2010). Several authors have posited that individuals must have a sense of self to experience jealousy and therefore may possess other complex social cognitions, such as self-awareness and theory-of-mind capabilities (Draghi-Lorenz, 1998; Leary, 2003; Lewis, 2003). This has led to claims that jealousy is unique to humans (Leary, 2003; Lewis, 2000; Panksepp, 2005; Ramachandran & Jalal, 2017) and emerges as a direct consequence of these cognitive capacities (Leary, 2003; Panksepp, 2010). However, nonhuman animals also experience rich social lives and so could benefit from such an emotion to protect valuable relationships from interlopers (Serpell, 1996; Topál et al., 1998). Dogs are an excellent model species for testing this hypothesis, given that the social bond between dogs and their owners shows similarities to the relationship between mothers and infants in humans (Palmer & Custance, 2008; Romero et al., 2014), and many dog owners attribute jealousy more frequently to their pets than other secondary emotions (Morris et al., 2008).
To date, a number of studies have attempted to test whether dogs show jealous behavior. In these studies, researchers examined how dogs react when observing their owners petting or feeding a potential social rival (either a real dog or a fake dog), compared with an inanimate object. Four studies have found differences in dogs’ reactions to these two events—including in the amount of time spent looking at the owner (Abdai et al., 2018; Prato-Previde, Nicotra, Fusar Poli, et al., 2018), snapping (Harris & Prouvost, 2014), whining (Harris & Prouvost, 2014), and moving in between or pushing the owner and the rival (Abdai et al., 2018; Harris & Prouvost, 2014)—and in brain-activation patterns (Cook et al., 2018), whereas one study found no behavioral differences (Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018). The literature to date therefore suggests that dogs might react to jealousy-inducing situations as humans do, with a constellation of different behaviors that vary among individuals (Hart et al., 1998; Hart, 2016; Mize & Jones, 2012). Current research, however, does not provide conclusive evidence that these differences in behavior are due to jealousy because no study has unambiguously mapped jealous behavior to jealousy-inducing situations. If these changes in behavior are a consequence of jealousy, they should emerge only when the owner interacts with the social rival, rather than as a consequence of the mere presence of a conspecific. Factors other than jealousy, such as arousal, fear, aggression, territoriality, or play directed toward a perceived conspecific, might be driving the changes to dog behavior seen between these two conditions (Abdai & Miklósi, 2018; Serpell, 2018; Vonk, 2018). It is also possible that owners might consciously or subconsciously behave differently toward the object they are petting, given that they have much more experience and comfort petting a dog or fake dog, compared with some of the objects used in these studies, such as a jack-o’-lantern pail.
Given the issues with interpreting past results on dog jealous behavior, and the more general difficulties of investigating emotions in other animals, we sought to identify parallels between the patterns of behavior around human jealousy and the contexts that might elicit jealous behavior in dogs. If multiple human signatures of an emotional-cognitive process are observed in other animals, we can be more certain that a similar process is present as well (Bastos & Taylor, 2020; Taylor, 2014).
Here, we investigated whether three human signatures of jealous behavior are seen in dogs. First, just as human infants are jealous of their mothers interacting with a potential rival but not an inanimate object (Hart et al., 1998; Mize et al., 2014), jealous behavior should emerge only when a dog’s relationship with its owner is threatened by a social rival, rather than when the owner interacts with a nonsocial object (Signature 1). Second, humans react specifically to jealousy-inducing interactions rather than to the presence or absence of a potential rival (Masciuch & Kienapple, 1993; Salovey & Rodin, 1986), so this behavior should emerge in dogs specifically as a consequence of the owner’s interaction with a social rival, rather than the mere presence of the owner and a conspecific with whom the owner does not interact (Signature 2). Finally, as shown in human adults (Buss et al., 1992; Harris, 2000), jealous behavior should emerge even when the interaction between the dog’s owner and the rival has to be mentally represented by the subject, rather than only when the interaction is directly visible (Signature 3). The presence of this third signature in dogs would be highly exciting because it would provide the first evidence that dogs have the capacity to mentally represent jealousy-inducing social interactions, a capacity that has been argued as uniquely human (Leary, 2003).
Statement of Relevance
Can nonhuman animals experience rich inner lives and feel complex secondary emotions? A promising avenue through which to investigate this question is jealous behavior in dogs. Previous research suggesting that dogs show jealous behavior has not been able to exclude alternative hypotheses for their reactions to jealousy-inducing situations. Our study provides a new quantitative measure that can be used to study jealous behavior in dogs—their approach response—and found evidence that dogs’ behavior is consistent with human patterns of jealous behavior. Dogs showed jealous behavior specifically toward jealousy-inducing interactions between their owners and a social rival, but not an inanimate object, even when these interactions were not directly visible to them.
To test for these three signatures, we measured the most consistent jealous behavior observed across different individuals and age groups in infants, a desire to approach (Mize et al., 2014; Mize & Jones, 2012), as a proxy for subjects’ eagerness to reach a social interaction taking place between either their owner and a realistic-looking fake dog or between their owner and a large fleece cylinder. In humans, jealous responses trigger electroencephalogram activity in the left prefrontal cortex, the same area associated with approach actions (Harmon-Jones et al., 2009; Mize et al., 2014; Mize & Jones, 2012).
Unlike researchers in previous studies (Abdai et al., 2018; Cook et al., 2018; Harris & Prouvost, 2014; Prato-Previde, Nicotra, Fusar Poli, et al., 2018; Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018), we ensured that the social rival was present in all conditions, kept owners blind to condition during test trials, and occluded actual contact between the owner and the potential social rival, thus requiring dogs to infer the occurrence of this interaction via mental representation.
Method
Ethics statement
This research was conducted in accordance with the Animal Welfare Act of 1999 and received ethics approval from The University of Auckland Animal Ethics Committee (Reference No. R001826) and The University of Auckland Human Ethics Committee (Reference No. R018410). Owners provided written informed consent prior to participating in the study.
Subjects
We tested 21 dogs, three of which were excluded from the study because of experimenter error. This resulted in a final sample size of 18 dogs (11 females; age: M = 4.6 years, SD = 1.9; see Table 1 for subject information). We used a Bayesian stopping rule to determine our sample size. We included dogs until we reached a Bayes factor (BF) greater than 3 or less than 0.33 (Wagenmakers et al., 2018), had tested at least 10 dogs, and had fully counterbalanced trial orders across subjects. All dogs were family pets recruited through an online survey and were brought in for testing sessions at the Clever Canine Lab, which is based at The University of Auckland. Subjects could participate in the study if they (a) had been in the owner’s household for a minimum of 6 months; (b) were not aggressive toward other dogs; (c) had never seen large, realistic-looking fake dogs at home; and (d) showed no signs of discomfort when attached to a tethered leash by an experimenter. These criteria ensured that subjects were likely bonded to their owner, were comfortable with the experimental setup, and would not pull toward a large, realistic-looking fake dog because of fear or aggression. Owners were not told the purpose of the study until all trials were concluded.
Demographic Information for All Subjects
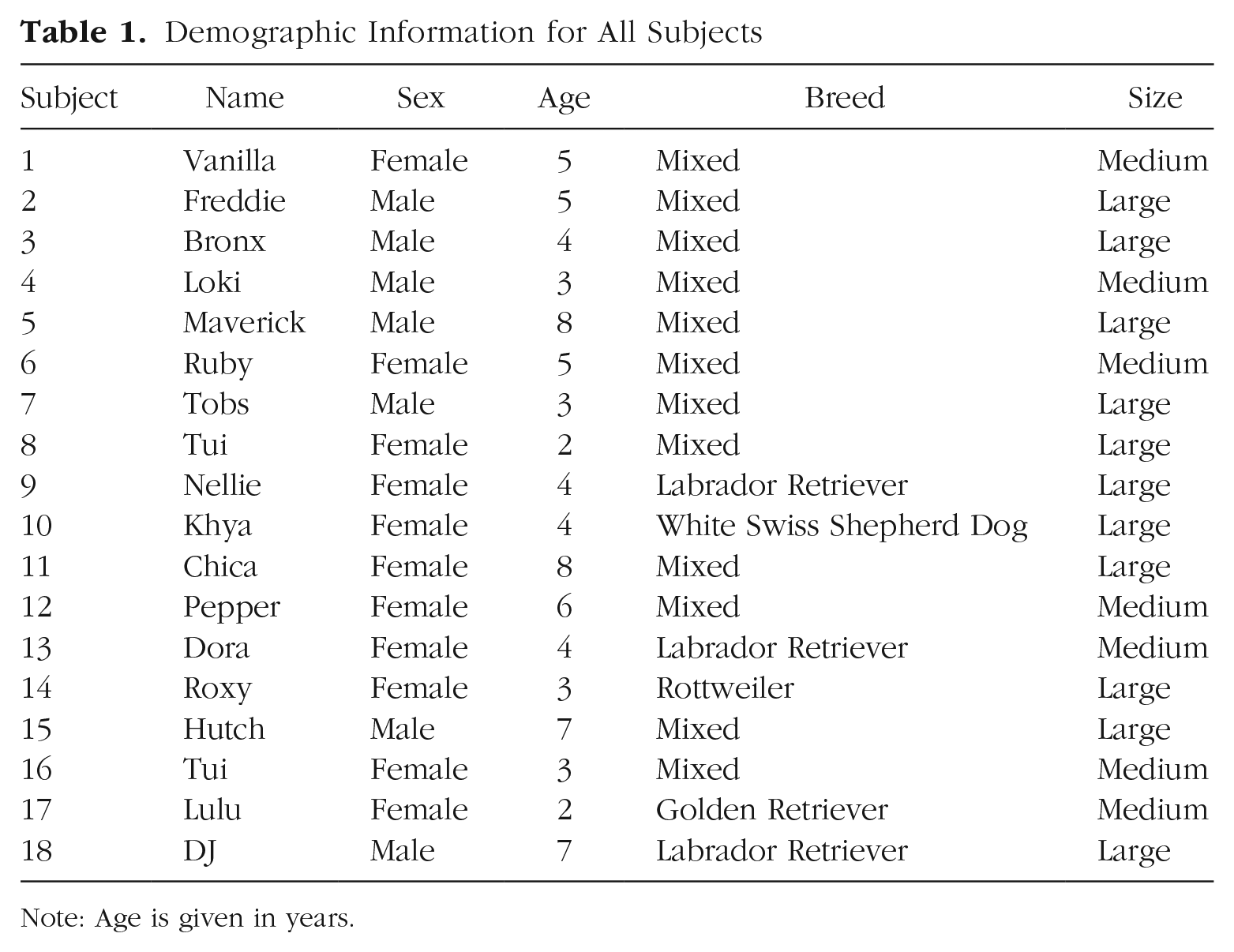
Note: Age is given in years.
Experimental setup
At the start of each trial, dogs were taken into a small room (3.6 m × 3.4 m) by an experimenter. They were held in the room until the trial had been set up in the main testing area. The dog’s peak pulling force (N) across each 30-s trial was measured using a WSDFG50K force gauge (Wedderburn, Sydney, New South Wales, Australia).
For each trial, the dog was centrally positioned 5 m away from a large barrier (2.90 m × 0.86 m), and a smaller barrier (0.86 m × 1.2 m) was placed 1.5 m to one side of the large barrier. The location of the small barrier (to the right or to the left of the large barrier) was randomized across subjects. Photographs of both barriers are provided in Figure 1c. Throughout all trials, the owner sat in a chair placed centrally behind the large barrier. Owners wore opaque goggles and noise-canceling headphones playing white noise throughout each trial. The large barrier was equipped with wheels and could be slid to reveal the owner behind it and any objects placed beside the owner. The small barrier was lifted and removed at the start of each trial and replaced at the end of each trial.

Stimuli and apparatus used in testing. Photographs of the fake dogs used in (a) the dog and cylinder conditions and (b) the confirmation trial. Photographs of the small and large barriers used to occlude objects throughout testing are shown in (c).
When designing our study, we took past research in this area into account. We used large, realistic-looking fake dogs with fur-like texture, unlike some past studies (Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018), in which models were made from plastic or had unusual proportions. We also used a greater distance, presenting the fake realistic-looking dogs 5 m away. There is evidence in the literature that large plastic models are convincing at a distance of 3 m (Barnard et al., 2012), suggesting that a realistic fake dog could be mistaken for a real social rival at a distance of 5 m. Fake dogs with fur-like texture, such as the ones used in our study, appear to be more convincing than plastic models. Shelter dogs react similarly to real dogs and stuffed toy fake dogs at a distance of 1 m, and stuffed toy fake dogs can elicit normal responses from real dogs in a playful context (Leaver & Reimchen, 2008). We opted to use a fake dog to ensure that olfactory, auditory, and visual cues remained as similar as possible across conditions. A real dog may have reacted to being spoken to by wagging its tail, creating a potential confounding factor (air movement or sound) that could have led subjects to react differently in the two conditions.
Procedure
All subjects experienced two test conditions, the dog condition and the cylinder condition, presented in a counterbalanced order across subjects. Dogs were first habituated to the force gauge and to the sight of their owners wearing goggles and headphones during a brief interaction prior to the first trial.
In the dog condition, owners first sat behind the large barrier. Experimenter 1 then placed a large, realistic-looking fake dog (60 cm × 22 cm × 60 cm) that resembled an American Foxhound next to them. The position of the fake dog (to the left or to the right of the owner) was randomized across subjects. A fleece cylinder was positioned in front of the barrier on the opposite side from the fake dog. After these objects were set up, Experimenter 2 (hereafter, referred to as the handler) brought the subject out of the small room and tethered it to a metal door frame by a leash. This leash was attached to the force gauge. Experimenter 1 then walked across the room to move the small barrier to one side of the large barrier. There were no objects behind the small barrier to reveal in this condition. Experimenter 1 then walked over to the large barrier and slid it across to reveal the fake dog sitting next to the owner while the owner faced straight ahead. The barrier was kept open for 5 s. If the dog looked away from the barrier during that time, the handler alerted Experimenter 1, who tapped the barrier to call the dog’s attention. The dog was then given 5 s to view the fake dog (Fig. 1a). The barrier was then closed again, concealing the fake dog. Experimenter 1 then opened a small fleece-covered shelf beside the owner, quietly tipped the fake dog over, and placed the shelf in the same location where the fake dog had originally stood. The owner was tapped on the shoulder as a signal to initiate petting and talking to the shelf as if it were a real dog. The use of the shelf ensured that the owner was provided with no tactile information about the dog that they were supposedly petting, ensuring that the owner could not provide unconscious cues to the dog on how to react to the interaction. Therefore, from the subjects’ perspectives, their owners appeared to be petting the previously seen fake dog, but the owners themselves were blind to experimental conditions. At the end of 30 s, the experimenter tapped the owner on the shoulder again. The owner then stopped the interaction, faced forward again, and placed their hands on their lap. The handler obtained the peak pulling force from the force gauge and then took the subject back into the small room.
The second test condition (the cylinder condition) was identical to the dog condition, but this time, the realistic-looking fake dog was placed behind the small barrier at a distance of 1.5 m away from the large barrier and facing the owner. It was therefore made visible for the full duration of the trial, unlike in the dog condition, where it remained hidden behind the large barrier. Instead of petting and speaking to the shelf behind the barrier, owners were now instructed to pet and speak to a fleece cylinder (22 cm × 22 cm × 60 cm) placed in front of the barrier. Owners were asked to speak to the fleece cylinder as if it were a real dog. However, the cylinder remained static and visible to the subjects to ensure that it was perceived as an inanimate object.
Regardless of whether they were interacting with the fake dog or the cylinder, owners were instructed not to say their dogs’ names, any names of dogs they might know, training commands, or words that might cause their dog to get excited (e.g., “treats,” “balls,” “walkies”). Instead, owners repeated a few generic phrases (e.g., “What a good boy!” “You are such a good dog!” “What a clever girl!”) several times for the duration of the trial. Owners rehearsed each condition while their dog was kept in a soundproof corridor before the start of each test trial.
Following the two test conditions, dogs experienced a final confirmation trial, which served as a manipulation check for our mentally represented interaction in the dog condition. It ensured that we were testing dogs who reacted to a visible jealousy-inducing condition, that is, they would pull at least 10 N over the course of 30 s while watching a directly observable jealousy-inducing interaction. In this trial, the experimenter slid the large barrier to reveal a fake dog behind it. Subjects then directly observed their owners, now not wearing headphones or goggles, petting and speaking to a realistic-looking fake dog for 30 s. One of two different fake dogs, resembling either a Doberman Pinscher or a German Shepherd Dog, both measuring 55 cm × 18 cm × 50 cm, was used in this interaction (Fig. 1b). New fake dogs were used in this trial in case subjects had realized by then that the previously encountered fake dog was not a real social rival, as it remained motionless for the duration of both test trials. The fake dogs used were randomly assigned and counterbalanced across subjects, unless owners mentioned that their dogs had had a negative experience with either breed, in which case the other one was used in the final trial. This reduced the risk of subjects’ individual experiences affecting their pulling force in the confirmation trial for reasons unrelated to jealous behavior. All tested dogs pulled at or above 10 N in this final trial (M = 112.15 N, 95% credible interval = [51.17, 173.13]); therefore, no subject was excluded from our analyses because it did not react appropriately when it could visibly see its owner petting a fake dog.
Analyses
We used the subject’s peak pulling force by the end of each 30-s trial as a proxy for dogs’ desire to approach their owner in each of the three trials. We predicted that if dogs showed signs of jealous behavior only when their owners interacted with a social rival, they should pull harder in the dog condition than in the cylinder condition. The two conditions were compared using a Bayesian paired-samples t test. The prior distribution used for the alternative hypothesis was a half-Cauchy distribution (r = .707) centered on an effect size of 0. All analyses were carried out using the BayesFactor package (Version 0.9.12-4.2; Morey & Rouder, 2018) in the R programming environment (Version 3.5.3; R Core Team, 2019). All data and analysis code are provided in the Supplemental Material available online.
Results
In the dog condition, subjects first observed their owner sitting next to a realistic-looking fake dog. The fake dog was then occluded by a barrier, and the owner petted and spoke to a fleece-covered shelf behind the barrier while a fleece cylinder remained in full view of the subject. In the cylinder condition, the owner petted and spoke to a visible fleece cylinder of approximately the same texture and size as the fake dog while the fake dog remained in full view of the subject. The order of the dog condition and cylinder condition was counterbalanced across subjects. In both conditions, we measured subjects’ desire to approach the scene by recording their peak pulling force over a 30-s interaction, that is, the hardest pull recorded over the course of the 30 s. We predicted that if dogs showed all three signatures of jealous behavior, they would pull harder in the dog condition than in the cylinder condition.
Dogs’ average peak pulling force in the dog condition was 158.03 N (95% credible interval = [66.79, 249.27]), whereas in the cylinder condition, it was 73.51 N (95% credible interval = [36.65, 110.37]). Dogs pulled significantly harder in the dog condition than in the cylinder condition (Fig. 2; Bayesian paired-samples t test: BF = 4.438 ± < 0.01% error).
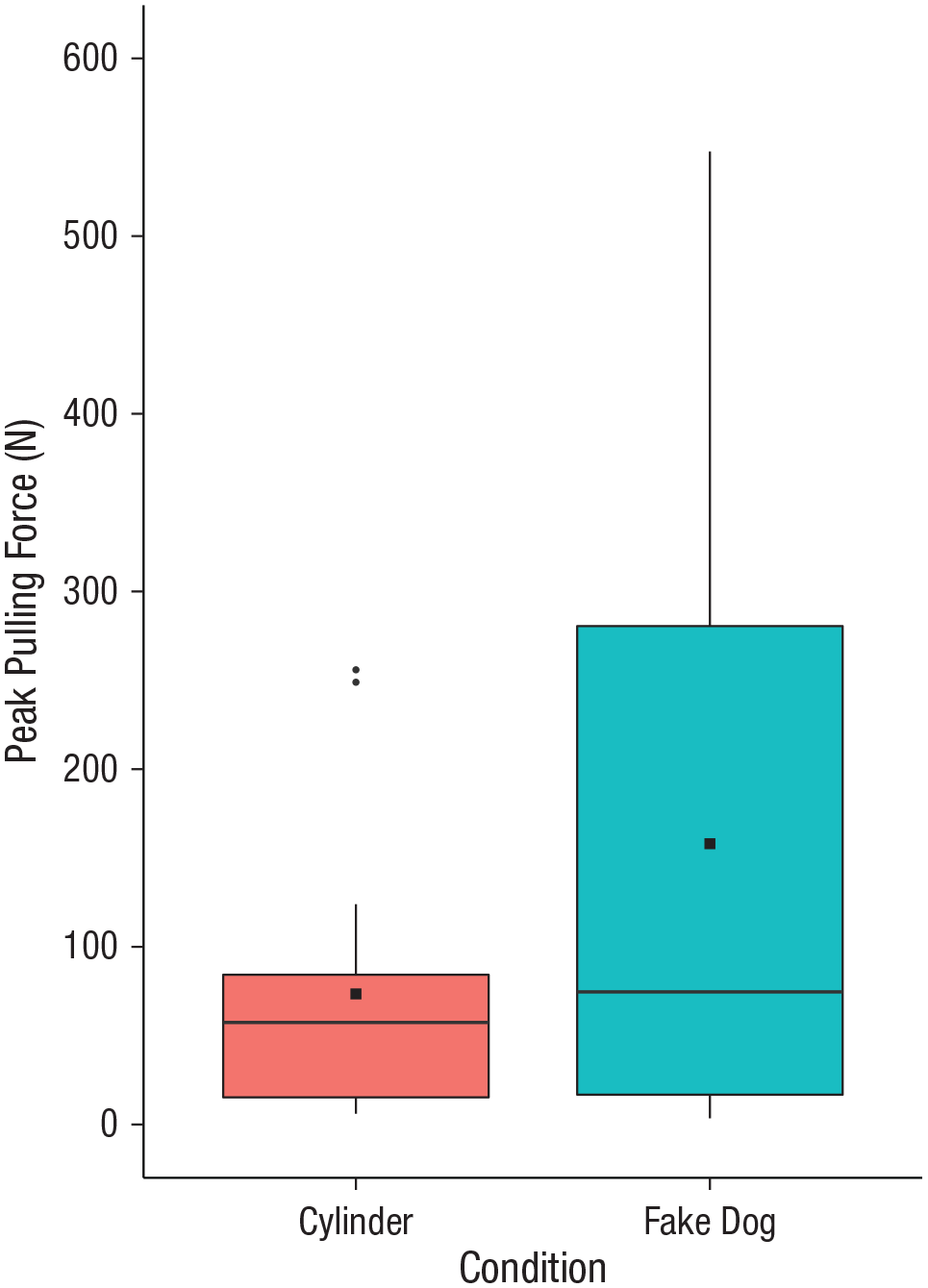
Dogs’ peak pulling force in the two test conditions (N = 18). In each boxplot, the horizontal bar indicates the median, and the square indicate the mean; whiskers mark the 25th and 75th percentiles, and the boxes include the data between the lower and upper quartiles. Outliers are indicated by round dots.
We compared dogs’ pulling force in our final confirmation trial, designed as a manipulation check, with their pulling force in the dog condition to exclude two potential alternative hypotheses. Dogs might have pulled more strongly in the dog condition than in the cylinder condition because they were attempting to gain visual access to the interaction (the visual-access hypothesis). Alternatively, they might have pulled less strongly in the cylinder condition than in the dog condition because the social rival, being fully visible, was more distracting and so elicited less pulling in the cylinder condition (the distraction hypothesis). Both these alternative hypotheses predict that dogs should have pulled harder in the dog condition, in which the fake dog was behind a barrier, than in the confirmation trial, in which the fake dog was fully visible. A Bayesian paired-samples t test revealed substantial evidence that there was no difference between the two conditions (BF = 0.121 ± < 0.01% error), suggesting that dogs were not simply seeking to gain visual access to a hidden interaction in the dog condition and that they were not more distracted when the fake dog was fully visible in the cylinder condition. This test supports our hypothesis that dogs’ pattern of behavior was consistent with jealous behavior and not a consequence of either of these alternative hypotheses.
We also gathered video data from 15 subjects after the confirmation trial. This allowed us to test whether subjects perceived the fake dog as a real conspecific, in which case we expected to see conspecific-directed behaviors after reaching the fake dog. We coded these videos for anogenital and facial sniffing. Fourteen out of 15 subjects (93.3% of dogs) performed conspecific-directed sniffing behaviors. These behaviors suggest that subjects perceived the fake dogs as real throughout the study.
Discussion
Our study shows that dogs pulled harder toward their owners when their owners appeared to interact with an out-of-sight fake dog than with a fleece cylinder. This occurred despite both the fake dog and fleece cylinder being present in both of these conditions and owners being blind to the object they were petting and speaking to. These results provide evidence that dogs show three signatures of human jealous behavior: (a) Jealous behavior emerged in response to a social partner attending to a social rival but not to a nonsocial object, (b) it emerged specifically in response to this interaction and not merely because of the presence of a conspecific, and (c) it emerged even though the interaction itself took place outside of the subject’s view.
These results provide strong support for past research (Abdai et al., 2018; Cook et al., 2018; Harris & Prouvost, 2014; Prato-Previde, Nicotra, Fusar Poli, et al., 2018; Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018) that has suggested that dogs behave differently in response to jealousy-inducing and nonjealousy-inducing contexts because they are experiencing some form of jealousy (or proto-jealousy; Harris & Prouvost, 2014; Prato-Previde, Nicotra, Fusar Poli, et al., 2018; Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018).
Although an approach behavior can serve various purposes in dogs, our results allow us to exclude alternative explanations for these past results, such as arousal, interest, fear, aggression, territoriality, or play directed toward their owner or a perceived conspecific (Bräuer & Amici, 2018; Serpell, 2018; Vonk, 2018). This is because our findings show that dogs differentiated between interactions involving a perceived social rival and an inanimate object even when the fake dog was present in both conditions. Had dogs’ reactions been a consequence of other factors, pulling force should not be expected to differ between the dog condition and cylinder condition, given that in both conditions, the social rival and the nonsocial object were presented, their owners behaved identically, and condition order was counterbalanced across subjects. The crucial difference between the two conditions was therefore what object the owners directed their attention to—either a nonsocial object or a perceived social rival. Furthermore, owners were blind in both the dog and cylinder conditions. Therefore, we can also exclude the hypothesis that owners might have consciously or subconsciously behaved differently toward the object they were petting, which could have cued dogs’ distinct reactions in the two conditions.
Importantly, our results suggest that dogs showed jealous behavior rather than envious behavior. Although these two emotions are similar in many ways, jealousy is concerned with protecting a social bond from an interloper, and envy is a desire for a resource that the subject cannot access (Parrot & Smith, 1993). Our study excludes envious behavior as a possible alternative explanation by providing the same social resources (the owner’s attention) to a social rival in the dog condition and to a nonsocial object in the cylinder condition. Had dogs simply desired the social resource of their owner’s attention, they should have pulled just as strongly in the control cylinder condition as they did when their owner directed that attention toward a perceived social rival in the test condition (dog condition). However, dogs pulled more strongly in the dog condition than they did in the cylinder condition, indicating that the reason for this behavior was that their owner’s attention was directed toward a social rival—and not the lack of attention in and of itself.
These findings also suggest that dogs may display more than a form of proto-jealousy, as has been suggested in the past (Harris & Prouvost, 2014; Prato-Previde, Nicotra, Fusar Poli, et al., 2018; Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018). In our study, dogs selectively displayed jealous behavior in response to a hidden social interaction rather than one that was directly observed. Although previous research has shown that dogs are capable of mentally representing hidden objects (Bräuer & Call, 2011), this ability has never been extended to mental representations of jealousy-inducing social interactions. In fact, this human signature of jealousy has, to date, been demonstrated only in adult humans (Buss et al., 1992; Harris, 2000) and has been claimed to be unique to our species (Leary, 2003). Dogs’ ability to “connect the dots” and infer that their owners’ actions were directed toward a hidden rival suggests that dogs not only are capable of mentally representing social interactions but also specifically do so when interpreting interactions that might threaten the social bond they have with their owners. This has implications above and beyond the similarities between jealous behavior in dogs and jealousy in adult humans. Our study provides evidence that dogs can mentally represent social interactions, which in itself is a complex cognitive ability. This finding therefore raises the question of how widely dogs might use mental representation when thinking about the world around them.
Darwin (1871/2004) was one of the first researchers to suggest that dogs show jealous behavior, stating that “everyone has seen how jealous a dog is of his master’s affection, if lavished on any other creature” (p. 92). The results here, in showing that dogs display three human signatures of jealous behavior, provide strong support both for this statement and for claims in more recent research that dogs differentiate between jealousy-inducing and nonjealousy-inducing interactions (Abdai et al., 2018; Cook et al., 2018; Harris, 2000; Prato-Previde, Nicotra, Fusar Poli, et al., 2018; Prato-Previde, Nicotra, Pelosi, & Valsecchi, 2018). Given our findings, it seems unlikely that past results are due to other factors, such as arousal, fear, aggression, territoriality, or play (Abdai & Miklósi, 2018; Serpell, 2018; Vonk, 2018). Our results also represent, to the best of our knowledge, the first evidence that jealous behavior in dogs may be more cognitively complex than originally thought, in that they show a signature of jealousy present in adult humans. Dogs still reacted with jealous behavior when they had to mentally represent the social interaction between their owner and a social rival, an ability that has so far been shown only in adult humans (Buss et al., 1992; Harris, 2000). This finding has important implications for dog welfare, given that it suggests that dogs are capable of inferring that unseen social interactions can threaten their bond with their owner. It is impossible to know whether dogs’ display of jealous behavior correlates with a human-like subjective experience of jealousy (Nagel, 1974). However, these results suggest that dogs may live much richer inner lives than we often give them credit for (Webb & de Waal, 2018) because they add strong support for the growing evidence that dogs may experience a secondary emotion.
Supplemental Material
sj-csv-1-pss-10.1177_0956797620979149 – Supplemental material for Dogs Mentally Represent Jealousy-Inducing Social Interactions
Supplemental material, sj-csv-1-pss-10.1177_0956797620979149 for Dogs Mentally Represent Jealousy-Inducing Social Interactions by Amalia P. M. Bastos, Patrick D. Neilands, Rebecca S. Hassall, Byung C. Lim and Alex H. Taylor in Psychological Science
Supplemental Material
sj-html-2-pss-10.1177_0956797620979149 – Supplemental material for Dogs Mentally Represent Jealousy-Inducing Social Interactions
Supplemental material, sj-html-2-pss-10.1177_0956797620979149 for Dogs Mentally Represent Jealousy-Inducing Social Interactions by Amalia P. M. Bastos, Patrick D. Neilands, Rebecca S. Hassall, Byung C. Lim and Alex H. Taylor in Psychological Science
Footnotes
Acknowledgements
We thank all dogs and their owners for their participation in this study and the volunteers for their assistance with data collection. We also thank Patrick Wood for blind-coding our video data and providing assistance with our figures.
Transparency
Action Editor: Eddie Harmon-Jones
Editor: Patricia J. Bauer
Author Contributions
All the authors designed the experiment. A. P. M. Bastos, P. D. Neilands, R. S. Hassall, and B. C. Lim carried out data collection. A. P. M. Bastos and P. D. Neilands analyzed the data. All the authors wrote the manuscript and approved the final version for submission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.