
Abstract
Rewards exert a deep influence on our cognition and behavior. Here, we used a paradigm in which reward information was provided at either encoding or retrieval of a brief, masked stimulus to show that reward can also rapidly modulate perceptual encoding of visual information. Experiment 1 (n = 30 adults) showed that participants’ response accuracy was enhanced when a to-be-encoded grating signaled high reward relative to low reward, but only when the grating was presented very briefly and participants reported that they were not consciously aware of it. Experiment 2 (n = 29 adults) showed that there was no difference in participants’ response accuracy when reward information was instead provided at the stage of retrieval, ruling out an explanation of the reward-modulation effect in terms of differences in motivated retrieval. Taken together, our findings provide behavioral evidence consistent with a rapid reward modulation of visual perception, which may not require consciousness.
The effect of reward on human behavior has often been studied in terms of its ability to motivate overt responses (Custers & Aarts, 2010). Recent research shows that reward also exerts an influence prior to overt choice by modulating cognitive processing (Pessoa & Engelmann, 2010). Particularly, reward influences the likelihood that stimuli will command attentional resources—regardless of whether the reward-signaling information is relevant to task goals (Hickey et al., 2010; Maunsell, 2004) or not (Le Pelley et al., 2015; Pearson et al., 2020)—enhancing cognitive processing at the location at which reward-signaling information appears.
Here, we investigated whether the reward effect can be observed even earlier in the stream of visual information processing, through modulating early perceptual processes. This possibility is suggested by studies showing that reward modulates neural gain in early sensory areas (e.g., primary visual cortex) that encode low-level visual features in animals (Shuler & Bear, 2006; Stănişor et al., 2013) and humans (Seitz et al., 2009; Serences, 2008). This raises the intriguing idea that encoding of visual stimuli might be modulated by their reward status: Establishing a neutral stimulus as a signal of high reward, relative to a signal of low reward, might enhance the speed and accuracy of encoding that stimulus. If participants become better at seeing high-reward signals, it would suggest that fundamental aspects of our phenomenological experience of the world are shaped by previous experiences of reward.
Consistent with the idea that reward-related information can be perceived rapidly, previous findings have shown that subliminally presented (and subjectively invisible) reward cues can elevate motivation and influence behavior. For example, a subliminal image of a high-value coin, relative to a low-value coin, can enhance cognitive performance (Bijleveld et al., 2009; Capa et al., 2013) and physical effort for a subsequent task (Bourgeois et al., 2016; Pessiglione et al., 2007). However, although this prior research shows that subliminal reward cues can be encoded and influence ongoing behavior, it does not establish whether perception of the cues themselves is influenced by their reward-signaling status (i.e., whether high-reward coins are more likely to be encoded than low-reward coins). Instead, previously observed effects may reflect downstream influences of reward on later cognitive processing (high- and low-reward coins are encoded equally, but high-reward coins provide greater motivation for subsequent behavior). The current study goes beyond prior research by investigating whether reward enhances perception such that high-reward signals are encoded faster or more precisely than low-reward signals.
To this end, we developed a novel adaptation of a task used by Soto et al. (2011) to include a manipulation of reward. On each trial (Fig. 1), a cue stimulus—a Gabor grating of a particular orientation—appeared for either 17 ms (short cue) or 200 ms (long cue) and was followed by a 100-ms pattern mask. This masking procedure rendered the cue subjectively invisible to participants on short-cue trials yet clearly visible on long-cue trials. The orientation of the cue (the to-be-remembered information) varied unpredictably on each trial. After a 2,000-ms interval, a test stimulus—a second Gabor grating with an orientation differing by 30° from the cue—appeared. Participants’ task was to respond according to whether the cue stimulus required rotation clockwise or counterclockwise to match the test stimulus. After making this orientation response, participants reported their subjective perception of the cue on a 4-point scale. This self-report approach reliably tracks the amount of relevant information that can be decoded from cortical activity (Dehaene & Changeux, 2011).
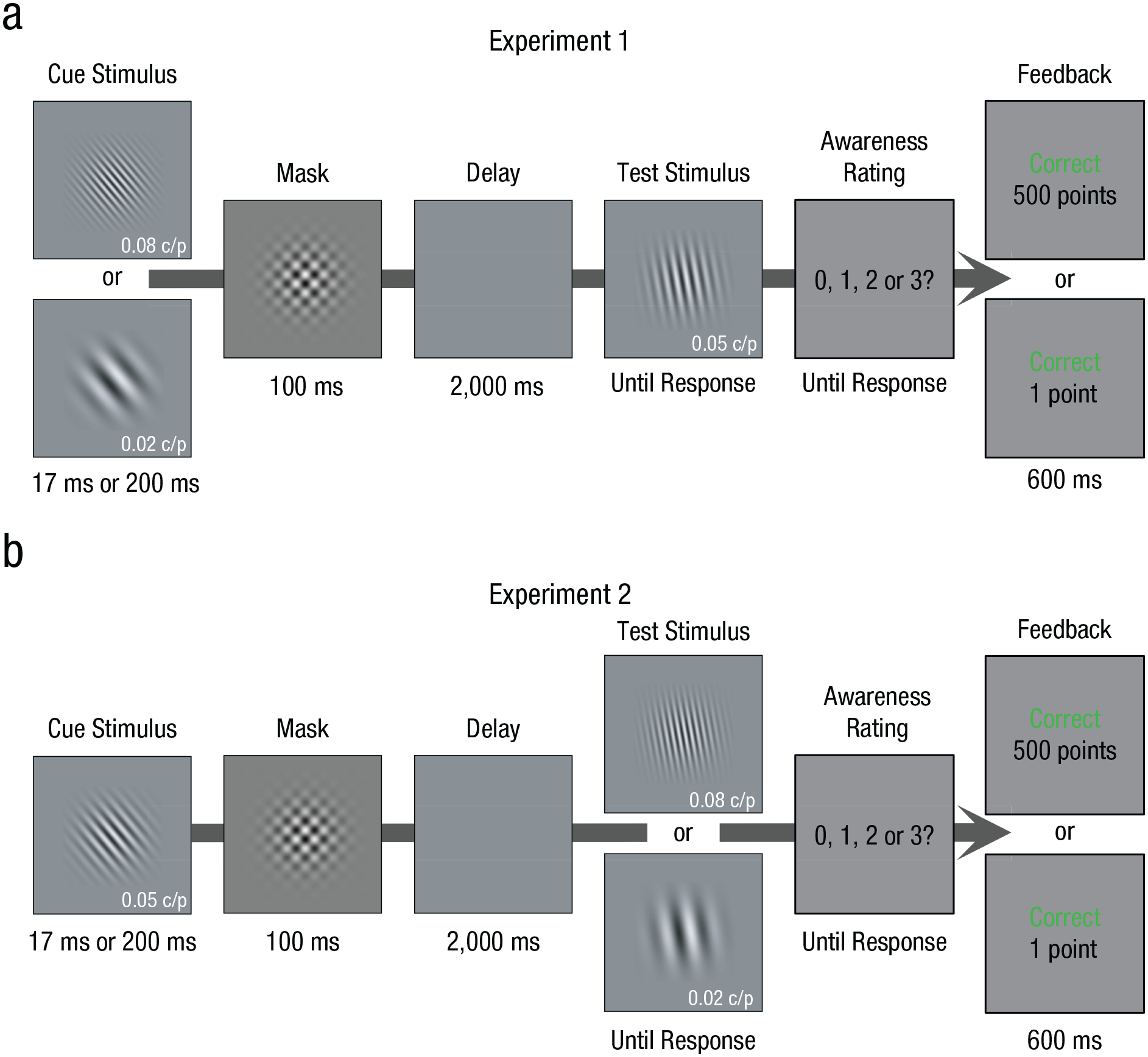
Experimental design. On each trial, a cue stimulus with an unpredictable orientation was presented for 17 ms or 200 ms and then masked. After a 2,000-ms delay, a test stimulus appeared: Participants’ task was to indicate (via pressing a button) whether the orientation of the test stimulus was clockwise or counterclockwise. They then rated their awareness of the cue (0 = did not see anything, 1 = maybe saw the cue, 2 = saw the cue but not its orientation, 3 = saw the cue and its orientation). Feedback then showed both perceptual-response accuracy (correct or incorrect) and the reward earned for that response. In Experiment 1 (a), the spatial frequency of the cue (0.02 or 0.08 cycles/pixel [c/p]) signaled whether a correct response would earn high or low reward (500 points or 1 point; points were later converted into money). In Experiment 2 (b), the spatial frequency of the test stimulus signaled whether the reward was high or low. The relationship between spatial frequency and reward was counterbalanced across participants. Note that the contrast of the cue and test stimuli are increased here for readability.
The critical aspect of the current task was the manipulation of reward. In Experiment 1, the spatial frequency of the cue signaled whether a correct response would earn high reward or low reward (Fig. 1a). Importantly, the cue’s reward-related information (spatial frequency) was unrelated to its response-related information (orientation), so the reward-signaling feature was not predictive of the correct response. If reward-signaling information influenced rapid encoding of the cue, we should observe greater response accuracy for high-reward than low-reward cues in the short-cue condition, when rapid encoding of the cue is required. Furthermore, a low awareness level of the cue necessitates rapid encoding of its weak sensory signals, rendering the reward effect most likely to manifest.
In Experiment 2, all cue stimuli had the same spatial frequency, and instead, the spatial frequency of the test stimulus signaled whether the reward was high or low (Fig. 1b). This allowed us to establish whether reward influenced the retrieval of information that had previously been encoded: Because encoding would be similar across all trials—because cues were equivalent—any performance enhancement on trials with a high-reward-signaling test stimulus must reflect participants’ greater motivation to retrieve the cue on these trials.
Statement of Relevance
Reward is deeply involved in the brain mechanisms responsible for how we see. Here, we show that the effect of reward can be detected even when the reward information appears so briefly that it is not consciously perceived, suggesting that reward has an impact on the very earliest stages of our brain’s information-processing stream. Specifically, we demonstrated that reward can enhance perception of visual information, an effect happening so fast that consciousness lags behind it. These findings indicate that our perception of the world is shaped by our previous experiences at a fundamental level, consistent with the idea that reward can change sensory gain at early stages of the visual system. More generally, our research suggests a deep and adaptive link between motivation and perception.
Method
Participants
Sixty-eight participants (23 male; age: M = 20.72 years, SEM = 0.37) took part in exchange for course credit or $20 (Australian); all reported normal or corrected-to-normal vision. We removed nine participants because their accuracy was low (< 60%) in the long-cue condition, in which the cue stimulus was clearly visible and so achieving high accuracy should have been easy. The final sample comprised 59 participants (Experiment 1: n = 30; Experiment 2: n = 29). For both experiments, we used pilot data to simulate the sample size sufficient for greater than 80% power. All participants received an additional, performance-based monetary reward (M = $4.92, SEM = $0.15). Experiment scripts and raw data are publicly available via OSF (https://osf.io/kp2qx/). This study was approved by the University of New South Wales Sydney Human Research Ethics Advisory Panel (Psychology).
Apparatus
Participants sat approximately 60 cm from a 23-in. monitor (resolution: 1,980 × 1,080 pixels; refresh rate: 120 Hz). Stimuli were presented using MATLAB (The MathWorks, Natick, MA) with Psychophysics Toolbox Version 3 extensions (Kleiner et al., 2007).
Stimuli
The cue and test stimuli (Fig. 1a) were Gabor gratings with a spatial frequency of 0.02, 0.05, or 0.08 cycles per pixel, centrally presented on a midgray background (RGB value: 128, 128, 128). The mask was a plaid comprising two overlaid gratings (spatial frequency = 0.05 cycles/pixel) oriented at 45° and 135°. Michelson contrast was 0.1 for cue and test gratings and 0.8 for the mask. The cue and test stimuli had a diameter subtending 4.9° of visual angle, and the mask had a diameter of 6.9°. The orientation of the cue stimulus was either 10°, 40°, 70°, 100°, 130°, or 160° (measured clockwise from vertical). The orientation on any given trial was randomly selected from these options, ensuring that it was unpredictable and varied. The orientation of the test stimulus was always 30° different from the cue, either clockwise or counterclockwise.
Design and procedure
Each trial began with a central fixation cross for 500 ms, followed by presentation of the cue stimulus for either 17 ms (short-cue condition) or 200 ms (long-cue condition). In Experiment 1, the spatial frequency of the cue was either 0.02 or 0.08 cycles per pixel. In Experiment 2, the cue had a spatial frequency of 0.05 cycles per pixel on all trials. The mask appeared immediately after the cue stimulus for 100 ms. This was followed by a blank delay of 2,000 ms, after which the test stimulus appeared. The spatial frequency of this test grating was 0.05 cycles per pixel in Experiment 1 and either 0.02 or 0.08 cycles per pixel in Experiment 2. Participants’ task was to press a key to indicate whether the cue stimulus would have to be rotated clockwise or counterclockwise to match the orientation of the test stimulus. The test stimulus remained visible until participants made their orientation response or until trial time-out after 10 s. Participants then rated their subjective awareness of the cue on a scale ranging from 0 to 3 (0 = did not see anything, 1 = maybe saw the cue, 2 = saw the cue but not its orientation, 3 = saw the cue and its orientation). Participants were instructed to use the 0 rating only if they felt they had not seen the cue at all (i.e., they had not seen its orientation, spatial frequency, or any other indication). After giving their awareness rating, participants received on-screen feedback stating their orientation-response accuracy and the corresponding reward that they had earned. The reward feedback ensured that participants received ongoing experience of the relationship between spatial frequency and reward magnitude. The intertrial interval was 500 ms.
In Experiment 1, the spatial frequency of the cue signaled that a reward would be given for a correct orientation response (500 points or 1 point, depending on the cue’s frequency; errors earned no points). In Experiment 2, the spatial frequency of the test stimulus signaled that a reward would be given for a correct response. The relationship between spatial frequency (0.02 cycles/pixel, 0.08 cycles/pixel) and reward (high, low) was counterbalanced across participants. Participants were informed of the relationship between spatial frequency and reward at the start of the experiment and were told that the points would later be converted into cash, which was paid to them at the end of the experiment. This approach of combining initial instruction of reward relationships with ongoing experience (via trial-by-trial reward feedback) follows previous research investigating the effects of reward associations on visual processing (e.g., Donohue et al., 2016; Failing & Theeuwes, 2018; Hopf et al., 2015; Watson, Pearson, Most, et al., 2019).
Each participant performed 24 practice trials before completing 240 trials in the main phase of the experiment, consisting of five repetitions of each combination of reward (high, low), cue duration (short, long), cue orientation (10°, 40°, 70°, 100°, 130°, 160°), and test-stimulus tilt (clockwise, counterclockwise), in random order. This meant that the to-be-remembered information (the orientation of the cue) varied randomly and unpredictably from trial to trial, independently of the reward information that the cue’s spatial frequency provided. Participants took a self-paced break after every 60 trials.
Data analysis
We excluded trials in which no orientation response was made before time-out after 10 s (0.21% of trials). Within the bound of this 10-s time-out limit, this was not a speeded task; consequently, our analyses focused on response accuracy.
The long-cue condition (200-ms cue presentation) should have been straightforward for participants. This condition was included for two reasons: (a) to ensure that participants received regular, clear exposure to the contingencies between spatial frequency and reward through trial-by-trial feedback and (b) so that some trials were relatively easy, to maintain motivation. Subsequently, our analyses focused on the short-cue (17-ms presentation) condition, because in this condition, rapid perceptual processing was critical for accurate response (Dehaene & Changeux, 2011). In both experiments, we compared overall accuracy in the short-cue condition between high-reward and low-reward trials and also compared each of these values with chance performance (50% correct). We then analyzed accuracy in high- and low-reward trials as a function of participants’ self-reported awareness of the cue on each trial. For this analysis, we divided trials into two types: those in which participants indicated no awareness of the cue (awareness rating of 0) and those in which participants indicated some awareness of the cue (trials with awareness rating of 1, 2, or 3, combined). We excluded participants from this analysis who had fewer than 10 trials for a given awareness level to ensure a reliable estimate of performance. In the Supplemental Material available online, we present accuracy results at each awareness level separately.
Unless otherwise stated, analyses were two tailed. The 95% confidence intervals (CIs) reported below are for the mean difference between relevant conditions. Bayes factors (BFs) were calculated for nonsignificant results (p > .05) to distinguish between insufficient sensitivity and true evidence for the null hypothesis (Dienes, 2014; Wagenmakers, 2007). We used a Cauchy prior with a width of 0.707 (Jeffreys, 1961), and following Jeffreys (1961; Rouder et al., 2009), we considered a BF (BF10) as providing substantial evidence in favor of the null hypothesis if it was less than 1/3.
Results
Experiment 1
Figure 2a shows the accuracy of orientation responses in Experiment 1 as a function of the cue’s duration (short: 17 ms; long: 200 ms) and the reward that it signaled. As anticipated, participants found the task relatively straightforward in the long-cue condition: Mean accuracy was high (> 85% correct) and did not differ significantly as a function of reward, t(29) = 0.17, p = .869, dz = 0.03, 95% CI = [–0.03, 0.03], BF10 = 0.20. Performance was poorer in the short-cue condition than the long-cue condition, but one-tailed t tests confirmed that short-cue accuracy was above chance (50% correct) for both the high-reward condition, t(29) = 5.40, p < .001, dz = 0.99, 95% CI = [0.60, ∞], and low-reward condition, t(29) = 3.71, p < .001, dz = 0.68, 95% CI = [0.55, ∞], indicating that the masked 17-ms cue was encoded on at least some trials. Critically, performance in the short-cue condition differed as a function of reward: Accuracy was significantly greater for high-reward than low-reward cues, t(29) = 2.38, p = .012, dz = 0.43, 95% CI = [0.01, ∞], consistent with the idea of perceptual enhancement for high-reward cues.
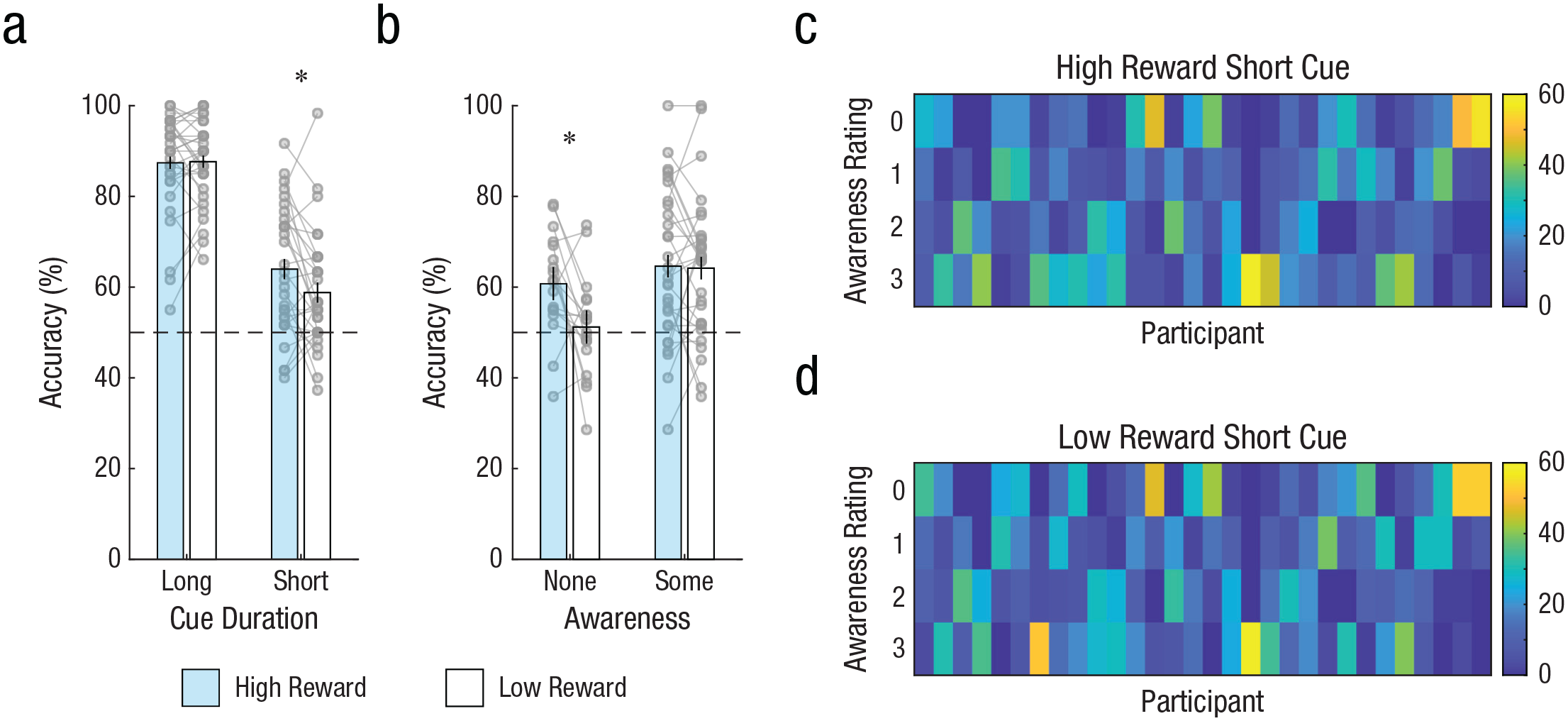
Results from Experiment 1. Mean accuracy for trials with a long cue duration (200 ms) and a short cue duration (17 ms) is shown (a) as a function of whether the cue signaled availability of high reward or low reward. Mean accuracy for short-cue trials in which participants reported having no awareness or some awareness of the cue is shown (b) as a function of reward availability. Data bars show group means; error bars represent within-subjects 95% confidence intervals. Gray circles show individual participant means; lines linking gray circles show performance of individual participants in each reward condition. Dashed gray lines represent chance-level performance (50% correct). Asterisks indicate significant differences between reward conditions (p < .05). The heat maps on the right show the number of ratings made by each participant at each awareness level, separately for short-cue trials in which (c) the cue signaled high reward and (d) the cue signaled low reward.
Next, we considered participants’ self-reported awareness of the cue. Figures 2c and 2d show the distribution of awareness ratings (0–3) across trials with high-reward and low-reward cues, respectively. If participants were aware of any reward-related enhancement in perceptual encoding, we would observe a difference in the pattern of awareness ratings between reward conditions (with greater awareness for high-reward cues). However, a multinomial mixed-effects model revealed that the pattern of awareness ratings did not differ significantly between reward conditions, F(3, 232) = 1.76, p = .157, adjusted R2 = .01. Similarly, comparing the proportion of high-reward and low-reward trials on which participants gave a zero rating (indicating no awareness of the cue) again yielded no significant effect of reward condition, t(29) = 1.19, p = .242, dz = 0.22, 95% CI = [–0.07, 0.02], BF10 = 0.37. The BF suggests that the reward-signaling status of the cue did not alter participants’ subjective experience of that cue (although the evidence only moderately favored the null hypothesis).
Finally, we examined whether the reward-related enhancement observed in orientation responses differed as a function of participants’ awareness of the cue (Fig. 2b). For this analysis, we divided trials into two types: those in which participants indicated no awareness of the cue (i.e., trials in which participants gave an awareness rating of 0, indicating that they had seen neither the spatial frequency nor the orientation of the cue) and those in which participants indicated some awareness of the cue (combined data from trials with awareness ratings of 1, 2, or 3; in the Supplemental Material, we present results at each awareness level separately). We excluded participants with fewer than 10 trials for a given awareness level to ensure reliable estimates of performance. This left a sample of 18 participants for analysis of the within-subjects effect of reward on no-awareness trials and 29 participants for the corresponding analysis on some-awareness trials.
For no-awareness trials, response accuracy was significantly greater than chance for high-reward cues, t(17) = 4.08, p < .001, one tailed (because below-chance performance would not be meaningful here), dz = 0.96, 95% CI = [0.56, ∞], but not for low-reward cues, t(17) = 0.46, p = .327, one tailed, dz = 0.11, 95% CI = [0.47, ∞], BF10 = 0.14. The BF in the latter case suggests substantial evidence in favor of the null hypothesis. Notably, accuracy was significantly greater for high-reward than low-reward cues for trials in which participants reported no conscious awareness of the cue, t(17) = 2.76, p = .007, one tailed, dz = 0.65, 95% CI = [0.04, ∞], demonstrating a reward-related performance enhancement in the absence of awareness.
For some-awareness trials, performance was significantly above chance for both high-reward cues, t(28) = 4.64, p < .001, one tailed, dz = 0.86, 95% CI = [0.59, ∞], and low-reward cues, t(28) = 4.79, p < .001, one tailed, dz = 0.89, 95% CI = [0.59, ∞], and there was no significant difference between reward conditions, t(28) = 0.19, p = .853, dz = 0.03, 95% CI = [–0.04, 0.05], BF10 = 0.20. This suggests that when participants had at least partial awareness of the cue, the reward-signaling status of that cue had no effect on response accuracy.
In summary, Experiment 1 found a reward-related performance benefit under the most challenging condition: when cue presentation was very brief, and participants reported having no awareness of the cue. This could reflect enhanced neural encoding of high-reward cues relative to low-reward cues. An alternative possibility is that encoding was similar for both cue types but that participants were more motivated to retrieve information about the high-reward cue when presented with the test stimulus because they could earn a larger reward for a correct response on high-reward trials. Experiment 2 probed this idea by testing for an influence of reward on motivated retrieval of cue information.
Experiment 2
In Experiment 2, the cue stimulus had the same (medium) spatial frequency on every trial and, hence, provided no reward information (Fig. 1b). Instead, the spatial frequency of the test stimulus signaled whether high or low reward was available for a correct response. This test stimulus was presented until response, so reward information was clearly visible to participants on every trial. If the reward-related performance enhancement observed in Experiment 1 reflected greater motivation to retrieve the cue when high reward was at stake, then we expected to observe a similar effect in Experiment 2 (or a larger effect, because reward information was now more visible). By contrast, if the performance enhancement in Experiment 1 reflected an influence of reward on encoding of the cue stimulus, then there should be no corresponding effect in Experiment 2 because encoding of the cue occurred before any reward information was provided.
Figure 3a shows accuracy of orientation responses in Experiment 2 as a function of cue duration and the reward signaled by the test stimulus. As in Experiment 1, accuracy on long-cue trials was generally high (> 85% correct), but somewhat counterintuitively, accuracy was significantly lower for high-reward trials than low-reward trials, t(28) = 2.60, p = .015, dz = 0.48, 95% CI = [–0.05, –0.01]. The reason for this is unclear. One possibility is that the representation of the high-reward test stimulus—presented until response and so clearly visible—interfered with memory of the cue, which was relatively weak because the cue was presented for only 200 ms. Future studies could assess this (post hoc) account by examining the effect of increasing the cue-exposure time in the long-cue condition. In the short-cue condition, accuracy was above chance for both high-reward trials, t(28) = 3.70, p < .001, one tailed, dz = 0.69, 95% CI = [0.54, ∞], and low-reward trials, t(28) = 3.84, p < .001, one tailed, dz = 0.71, 95% CI = [0.55, ∞]. Critically, and unlike in Experiment 1, there was no enhancement of performance on high-reward relative to low-reward trials, t(28) = 0.73, p = .765, one tailed (because we tested for a difference in the same direction as in Experiment 1), dz = 0.14, 95% CI = [–0.04, ∞], BF10 = 0.13, and the BF indicated substantial evidence for the null hypothesis. The latter finding suggests that performance on short-cue trials was unaffected by differences in reward-motivated retrieval.
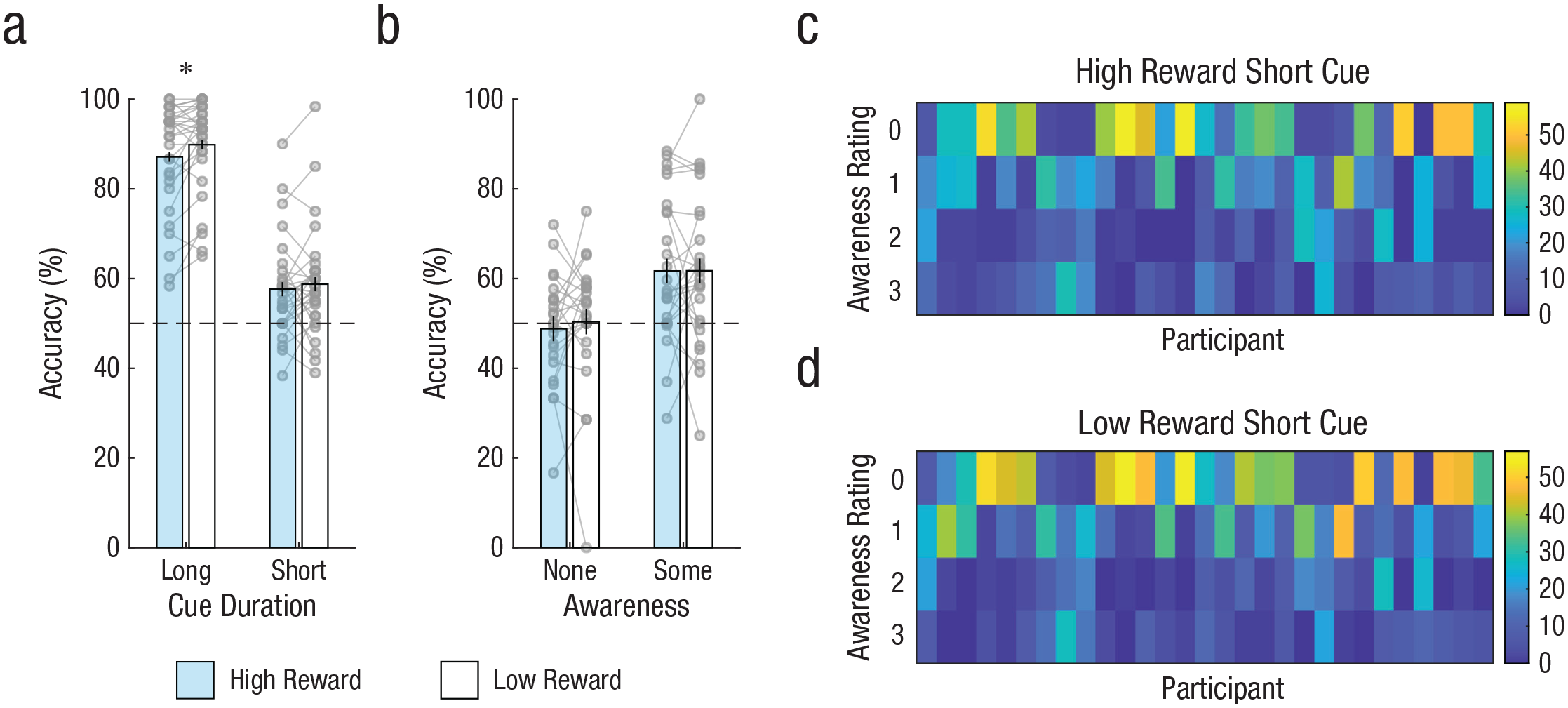
Results from Experiment 2. Mean accuracy for trials with a long cue duration (200 ms) and a short cue duration (17 ms) is shown (a) as a function of whether the test stimulus signaled availability of high reward or low reward. Mean accuracy for short-cue trials in which participants reported having no awareness or some awareness of the cue is shown (b) as a function of reward availability. Data bars show group means; error bars represent within-subjects 95% confidence intervals. Gray circles show individual participant means; lines linking gray circles show performance of individual participants in each reward condition. Dashed gray lines represent chance-level performance (50% correct). The asterisk indicates a significant difference between reward conditions (p < .05). The heat maps on the right show the number of ratings made by each participant at each awareness level, separately for short-cue trials in which (c) the cue signaled high reward and (d) the cue signaled low reward.
Figures 3c and 3d show the distribution of awareness ratings for short-cue trials on which reward was high and low, respectively, in Experiment 2. The overall pattern of awareness ratings did not differ significantly between reward conditions, F(3, 224) = 2.03, p = .111, adjusted R2 = .28, and neither did the proportion of no-awareness (0-ratings) trials, t(28) = 1.30, p = .205, dz = 0.24, 95% CI = [–0.04, 0.01], BF10 = 0.42. Thus, reward did not appear to change subjective awareness of the cue, although the latter BF suggests only moderate support for the null hypothesis.
Figure 3b shows response accuracy for short-cue trials in Experiment 2 in which participants reported no awareness of the cue (ratings of 0) or some awareness of the cue (ratings of 1–3). Removing participants with fewer than 10 trials for a given awareness level left 24 and 26 participants for analysis of no-awareness and some-awareness trials, respectively. For no-awareness trials, one-tailed tests revealed that accuracy was not greater than chance for high-reward trials, t(23) = 0.49, p = .684, dz = 0.10, 95% CI = [0.45, ∞], BF10 = 0.13, or low-reward trials, t(23) = 0.11, p = .456, dz = 0.02, 95% CI = [0.45, ∞], BF10 = 0.11. Notably, and unlike in Experiment 1, there was no evidence of greater accuracy for high-reward than low-reward trials, t(23) = 0.57, p = .714, one tailed (because we tested for a difference in the same direction as in Experiment 1), dz = 0.12, 95% CI = [–0.06, ∞], BF10 = 0.13. For some-awareness trials, accuracy was above chance for both high-reward trials, t(25) = 3.73, p < .001, dz = 0.73, 95% CI = [0.56, ∞], and low-reward trials, t(25) = 3.50, p < .001, dz = 0.69, 95% CI = [0.56, ∞], and did not differ between reward conditions, t(25) < 0.01, p = .500, dz < 0.01, 95% CI = [–0.04, ∞], BF10 = 0.11. Consistent with the analysis collapsing over all short-cue trials of Experiment 2, these findings suggest that reward did not influence performance as a consequence of differences in motivated retrieval.
Discussion
This study investigated the influence of reward on rapid visual perception. In Experiment 1, the cue contained both reward-related and response-related information. We found a reward-related increase in accuracy when the cue appeared briefly (for 17 ms). In Experiment 2, the test stimulus signaled reward magnitude: The cue provided no reward information, and hence, a reward could not influence perceptual encoding of this cue. Under these conditions, there was no evidence of greater accuracy on high-reward than low-reward short-cue trials, and a Bayesian analysis provided substantial support for the null hypothesis. Notably, a direct comparison of accuracy in the critical short-cue (17-ms) trials across experiments—using a binomial linear mixed-effects model—highlighted a difference in the pattern of performance when reward information was presented at the time of encoding (Experiment 1) or retrieval (Experiment 2). Specifically, a two-way analysis of variance revealed an interaction between the effect of experiment (Experiment 1, Experiment 2) and reward magnitude (high, low), F(1, 7892) = 5.20, p = .023, adjusted R2 = .058, and a significantly greater effect of reward on accuracy when information was placed at encoding rather than retrieval. Taken together, these findings suggest that the reward-related advantage observed in Experiment 1 did not reflect a difference in motivated retrieval of cue information, because if this were occurring, we should have seen a similar (or larger) effect in Experiment 2—whereas in fact the opposite was the case. Instead, our results suggest that the reward advantage observed in Experiment 1 stems from the presence of reward information at the time of encoding of the cue, despite it being masked from consciousness.
One interpretation of these findings is that reward changes encoding: The visual system becomes more sharply “tuned” to high-reward than low-reward information; high-reward stimuli undergo rapid encoding with higher fidelity. Consequently, the perceptual attributes of high-reward-signaling cues would be encoded with greater precision in a limited time, and this would include the to-be-remembered attribute of orientation—even though orientation was actually independent of reward status in our experiments. This possibility of enhanced encoding of high-reward cues fits with neuroimaging findings showing that reward modulates neural gain in early visual areas that encode low-level features (Serences, 2008; Stănişor et al., 2013). Consistent with this account, Experiment 1 found a reward advantage only when the cue appeared very briefly (17 ms) and not when it appeared for longer (200 ms). The long-cue condition provided ample time to encode the cue so reward modulation of encoding would have no detectable effect. In the short-cue conditions, however, little time was available, so differences in encoding speed for high- and low-reward cues would have a more noticeable effect on performance.
An alternative (but related) interpretation of the current reward benefit is that high- and low-reward cues are encoded equivalently, but this representation is better propagated over the 2-s delay between cue and test for the high-reward stimulus. This account posits that reward influences the maintenance of a perceptual representation. Our finding of reward modulation for no-awareness trials seems contrary to traditional views of working memory, which hold that active maintenance requires consciousness (Baddeley, 1992). However, recent research indicates that briefly presented visual information can be maintained even when subjectively invisible (King et al., 2016; Soto & Silvanto, 2014). Thus, beyond modulating encoding, reward might have facilitated the sustained processing of weak visual signals until test, when they could influence performance. Future studies could further investigate the processing stage at which reward operates using neuroimaging with high temporal resolution (electroencephalography, magnetoencephalography) to track changes in neural information over time.
Deeper analysis of short-cue trials of Experiment 1 showed a significant influence of reward on accuracy for trials in which participants reported no conscious awareness of the cue, but not when they reported at least partial awareness. The no-awareness trials are those on which neural encoding of the cue was presumably most limited, rendering these trials most likely to reveal a rapid reward modulation of performance. This finding also suggests that the reward information is extracted at a very early stage of information processing, raising the possibility that the underlying neural mechanisms might involve subcortical reward-processing regions distinct from the consciousness-generating network (Gottfried et al., 2003; McFadyen et al., 2020). According to this account, the lack of reward-modulation effect on “aware” trials may suggest that extensive processing triggered by conscious awareness swamps the reward-modulation effect, consistent with previous evidence that conscious information produces greater and more widespread neural activity than unconscious information (Dehaene & Changeux, 2011; Yi et al., 2004). Although reward influenced the accuracy of orientation responses, it did not significantly alter the proportion of short-cue trials on which participants reported no awareness as opposed to some awareness of the cue. Thus, enhanced perception of the cue in the high-reward condition did not seem to increase its subjective visibility, consistent with the idea that reward modulation operates outside awareness, although Bayesian analyses indicated only moderate support for the null hypothesis.
Regardless, the involvement of consciousness is of secondary importance here: The key finding of the current study is that perceptual encoding was enhanced for briefly presented, reward-signaling stimuli, demonstrating that the underlying perceptual processes are malleable and can be shaped by our experience of reward. In effect, the perceptual system enhances rapid processing of high-reward information. This research raises further important questions. For example, future studies could further probe the source of the reward effect. One possibility is that this effect reflects top-down modulation, whereby participants develop a greater expectancy of high-reward stimuli—because these stimuli are worth more—which leads to upweighting of sensory gain for the corresponding spatial frequency (Kaiser et al., 2019; Yon et al., 2019). This influence of expectancy may be mediated by attention; that is, expectancy-driven attentional prioritization of high-reward stimuli may enhance perceptual encoding of these stimuli. This possibility is consistent with previous findings showing attentional enhancement for detecting (and learning) a briefly presented feature when it is task irrelevant (for reviews, see Seitz & Watanabe, 2005; Shibata et al., 2014; Watanabe & Sasaki, 2015). However, the current finding that reward enhanced performance only when participants were less aware of the reward cue would then suggest that any such experience-driven upweighting operates outside of consciousness. An alternative possibility is that the reward effect reflects a more automatic influence, whereby experience of stimulus-reward pairings conditions the sensitivity of feature detectors irrespective of expectancies. Another open question is whether the perceptual system adapts to increase sensitivity to reward specifically or also to other motivationally significant outcomes, such as punishment (Watson, Pearson, Wiers, et al., 2019).
The influence of reward on perception demonstrated here may be implicated in previous demonstrations of reward modulation of cognitive functions. For example, high-reward-signaling stimuli are more likely than low-reward-signaling stimuli to capture visual attention (Anderson et al., 2011; Le Pelley et al., 2017), indicating that reward influences salience on an attentional priority map (Belopolsky, 2015; Failing & Theeuwes, 2018). The current effect of reward on perception may contribute to this reward-modulated attention: If high-reward stimuli are encoded more rapidly, they can exert an earlier influence on the priority map, driving more rapid capture of attention. In situations involving multiple stimuli competing for cognitive resources, reward modulation of perception may have knock-on effects that last beyond the brief temporal window investigated in the current experiments.
Beyond its theoretical significance, the idea of perceptual enhancement for reward-related information may have clinical implications. For example, drugs of abuse produce potent neural reward signals (Berridge et al., 2009; Robinson & Berridge, 2001), and recent work shows that addictive behaviors are associated with individual differences in reward-related attentional biases (Albertella et al., 2019; Anderson et al., 2013). The current findings suggest that reward can exert even earlier influences on information processing, raising the possibility of reward-related perceptual differences implicated in addiction.
We should add a note of caution in drawing conclusions regarding the conscious (or unconscious) nature of the perceptual effect observed here. We used a subjective measure of awareness on the assumption that conscious states are characterized by an individual’s ability to report them (Lau & Rosenthal, 2011). However, this approach is susceptible to response bias (Persuh, 2017): We cannot be sure of participants’ internal criteria. There may be trials where participants rated awareness as zero yet had conscious access to (partial) cue information. Participants were not incentivized to make accurate awareness ratings (because the experimenter could not know the “correct” answer) and may not have reflected carefully on their level of awareness. Such concerns limit our confidence in conclusions relating to absolute levels of consciousness, an issue that is not always acknowledged in work using subjective measures (Tsuchiya et al., 2015). We are safer in drawing inferences regarding relative levels of consciousness and conclude that Experiment 1 demonstrated a reward enhancement when participants were less aware of the cue.
In conclusion, the present study provides behavioral evidence consistent with a rapid, reward-related modulation of visual perception. Our findings suggest that reward is fundamentally implicated in visual information processing, and the perceptual system becomes sensitized for detection and encoding of reward-related stimuli in a way that does not require conscious awareness.
Supplemental Material
sj-pdf-1-pss-10.1177_09567976211021843 – Supplemental material for Reward Rapidly Enhances Visual Perception
Supplemental material, sj-pdf-1-pss-10.1177_09567976211021843 for Reward Rapidly Enhances Visual Perception by Phillip (Xin) Cheng, Anina N. Rich and Mike E. Le Pelley in Psychological Science
Footnotes
Acknowledgements
We thank Peter Humburg for helpful advice regarding statistical analyses.
Transparency
Action Editor: M. Natasha Rajah
Editor: Patricia J. Bauer
Author Contributions
All the authors conceptualized and designed the study. P. (X.) Cheng programmed the experiment and collected and analyzed the data. M. E. Le Pelley supervised the research. P. (X.) Cheng drafted the manuscript. M. E. Le Pelley provided critical revisions to all drafts. A. N. Rich provided critical comments on the final draft. All the authors approved the final manuscript for submission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.