
Abstract
This study uses palynologic data to determine the effects of regional climate variability and human activity on the formation and development of tree islands during the last ~4000 years. Although prolonged periods of aridity have been invoked as one mechanism for their formation, Native American land use has also been hypothesized as a driver of tree island development. Using pollen assemblages from head and near tail sediments collected on two tree islands and documented archeological data, the relative roles of Native Americans, climate variability, and recent water-management practices in forming and structuring Everglades tree islands are examined. The timing of changes recorded in the pollen record indicates that tree islands developed from sawgrass marshes ~3800 cal. yr BP, prior to human occupation. Major tree island expansion, recorded near tail sediments, occurred ~1000 years after initial tree island formation. Comparison of the timing of pollen assemblages with other proxy records indicates that tree island expansion is related to regional and global aridity correlated with southward migration of the Intertropical Convergence Zone. Local fire associated with droughts may also have influenced tree island expansion. This work suggests that Native American occupation did not significantly influence tree island formation and that the most important factors governing tree island expansion are extreme hydrologic events due to droughts and intense twentieth century water management.
Introduction
Climate processes associated with deglaciation, insolation, and natural climate variability played a primary role in determining the spatial distribution of vegetation on local, regional, and global scales. These vegetation patterns have been altered to varying degrees by human activities such as land clearance, burning, agriculture and selective planting (Abrams and Nowacki, 2008; Day, 1953; Delcourt and Delcourt, 2004; Denevan, 1992; Heckenberger et al., 2007; McLauchlan, 2003; Meadows and Baxter, 2001; Vale, 1982; Williams, 2000). However, in the Americas for example, the spatial scale of these human-modified landscapes by pre-Columbian people is thought to be exaggerated and limited to localized areas (Bush et al., 2007). Regardless, since their arrival in the New World, European settlers have continually modified the landscape of eastern North America vegetation resulting in the highly altered vegetational landscape of the twenty-first century (Brush and Davis, 1984).
Native Americans in South Florida lived primarily along the coast of southern Florida, utilizing tree islands of the Everglades as temporary fishing and hunting camps during the winter (Griffin, 1988). One proposed mechanism of tree island formation involves Native American modification of the landscape (Carr, 2002). Deposition of thick (10–30 cm) faunal middens and the collapse of wooden platforms have been hypothesized as loci for sediment accumulation, raising the elevation of the tree island surface above the adjacent marsh and initiating the establishment of woody taxa (Carr, 2002).
However, another hypothesis suggests that tree islands formed over topographic highs in the underlying limestone bedrock and developed through positive biogeochemical feedback (Ross et al., 2006). This hypothesis is consistent with previous paleoecological studies indicating that tree islands formed during periods of regional aridity (Willard et al., 2006), possibly tied to southward displacement of the mean position of the Intertropical Convergence Zone (ITCZ). The ITCZ is a low latitude belt of convective storms circling the Earth, whose northerly or southerly position is driven by changes in insolation. The relative strengths of the North Atlantic Oscillation (NAO) and the El Niño Southern Oscillation (ENSO) have also been invoked as important mechanisms controlling precipitation in the circum-Caribbean and the Everglades (Cronin et al., 2002; Haug et al., 2001, 2003). Proxy records show that prolonged changes in precipitation during the late Holocene drove changes in the wetland plant assemblages throughout the Everglades (Bernhardt and Willard, 2009; Donders et al., 2005; Willard et al., 2006). These extreme hydrologic events are thought to be more important than average annual conditions in shaping tree island vegetation (Wetzel et al., 2008).
Here, paleoecological analysis is used to quantify the relative roles of humans and climate as drivers of the formation and succession of tree islands. The following questions are addressed: (1) what was the pre-tree island vegetation on the elevated heads and near tails; (2) did Native American activity initiate tree island formation or modify the structure of previously established tree islands; and (3) how did regional climate affect tree islands and Native American cultures? Detailed records from tree island heads and near tails are used to clarify the timing of tree island formation and integrate the resulting data with archeological analyses from the same tree islands. Previous research (e.g. Willard et al., 2006) focused almost exclusively on sedimentary sequences collected in the near tails of tree islands. Although near tail sediments have well-dated pollen sequences detailing tree island development, it is unknown how they compare to sediment cores collected on tree island heads. This study compares pollen records from tree island heads and near tails to further refine the patterns and timing of tree island formation.
Regional overview
The Everglades presently covers an area of ~ 6000 km2 and is a phosphorus-limited, precipitation-driven, subtropical wetland system (Figure 1; Davis et al., 1994). The natural hydrologic system covered an area of ~12 000 km2 (Davis et al., 1994) and reflected the seasonal precipitation pattern. The distribution of Everglades wetland plant communities is controlled by water depth, hydroperiod, substrate type, and fire regime (Kushlan, 1990). Tree islands comprise approximately 14% of the Everglades (Sklar and van der Valk, 2002). Fixed tree islands are teardrop-shaped features, oriented parallel to flow, formed over topographic highs in the underling Pliocene and Pleistocene limestone bedrock (Gleason and Stone, 1994; Heisler et al., 2002; Loveless, 1959). The elevated dry head serves as a refuge for flora and fauna that cannot tolerate prolonged flooding and consists primarily of subtropical hardwoods, shrubs, and ferns (Heisleret al., 2002). General examples of woody vegetation found on elevated, teardrop-shaped, tree islands include Bursera simaruba, Rivina humilis, Eugenia axillaris, Annona glabra, Chrysobalanus icaco, Persea borbonia, Ilex sp., Salix carolina, Myrsine floridana, and Morella cerifera (Heisler et al., 2002). The near tail lies downstream of the head (Figure 1) and experiences longer hydroperiods and water depths. Accordingly, the vegetation is composed of plants with a greater tolerance for flooding, including water-tolerant hardwoods (Morella and Cephalanthus), ferns (Osmunda regalis), sedges (e.g. Cladium), and other marsh taxa (e.g. Sagittaria and Pontederia) (Heisler et al., 2002).
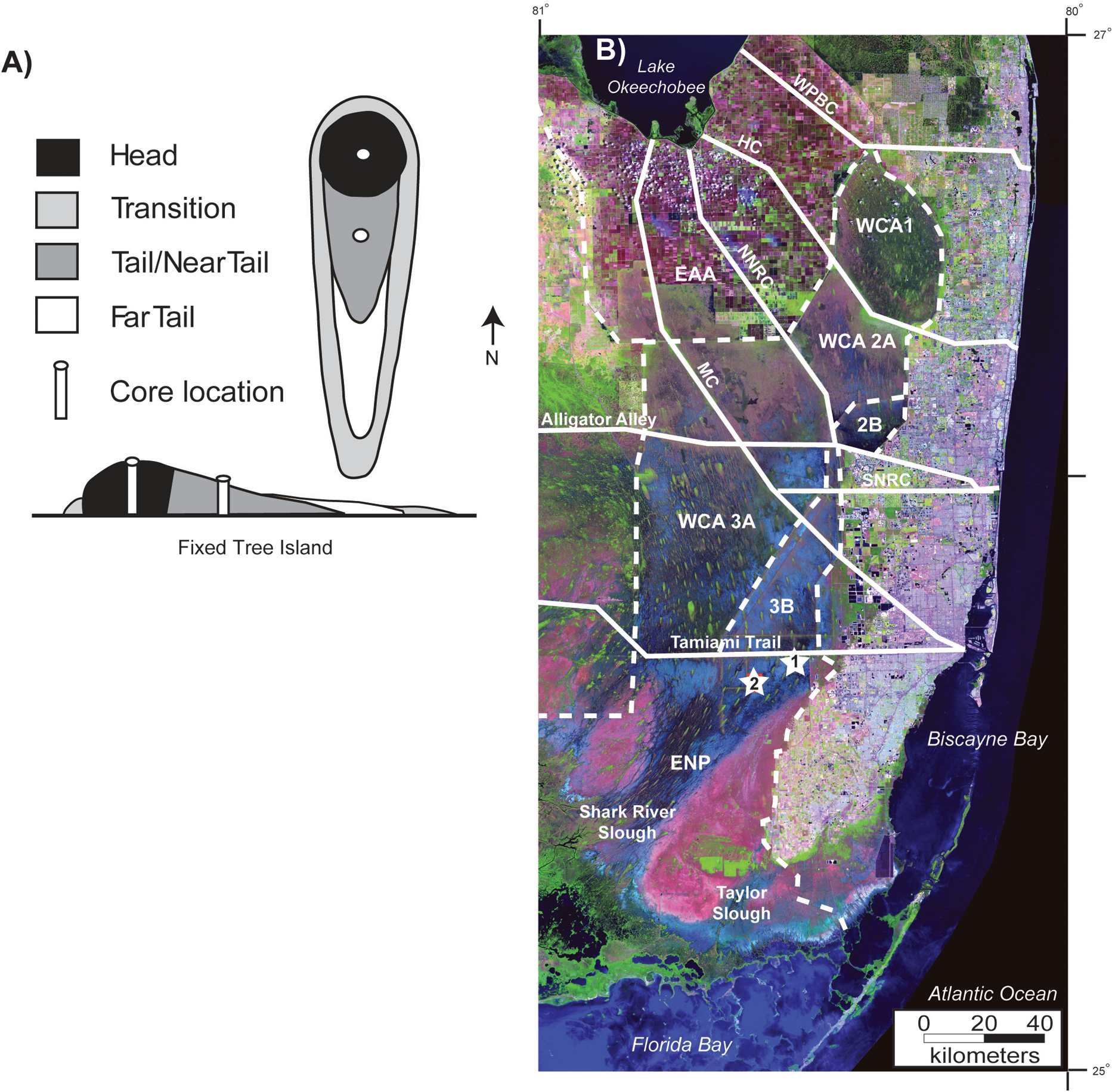
Location map. (A) Idealized illustration of the structure and vegetation types of a typical fixed, or teardrop-shaped, tree island. The figure is modified after Willard et al. (2006). (B) Satellite image of Florida Everglades with core locations (Image modified from South Florida Water Management District, 2009). Solid white lines note major canals and roads. WPBC, West Palm Beach Canal; HC, Hillsborough Canal; NNRC, New North River Canal; SNRC, South North River Canal; and MC, Miami Canal. Dotted lines mark the boundaries of Water conservations areas and Everglades National Park. WCA, Water Conservation Area; ENP, Everglades National Park; EAA, Everglades Agricultural Area. White stars with numbers indicate cores examined in this study. 1, Heartleaf Hammock; and 2, Duck Club Hammock
During the late nineteenth and early twentieth century European-American settlers constructed fishing and hunting cabins and some permanent domiciles on tree islands of the Everglades, as well as drainage efforts rendering parts of the Everglades usable for agricultural and urban development (Light and Dineen, 1994). The first major drainage phase began in the
Climate and hydrology
The Everglades is characterized by seasonal precipitation, with most rain (70%) falling during the wet season from May through November (average 86 cm; range 58–135 cm), resulting in high Everglades water levels in the summer and fall (Duever et al., 1994). An average of 43 cm of rain (17–78 cm) falls during the remainder of the year (McPherson and Halley, 1996). This seasonal precipitation pattern is driven by migration of the Bermuda High (BH). In the wet season (summer–fall), the system is located near Bermuda, and in the dry season, the BH is located near the Azores (Stahle and Cleaveland, 1992). The El Niño-Southern Oscillation influences interannual climate variability, with heavy winter rainfall during large El Niño warm events and low winter precipitation, severe lightening strikes, and increased fire during La Niña events (Beckage et al., 2003; Kiladis and Diaz, 1989).
In the natural Everglades system, water flowed southward during the wet season along a gentle slope of 3 cm/km (Kushlan, 1990) from Lake Okeechobee toward Florida Bay and the Gulf of Mexico through Shark River Slough and, to a lesser extent, Taylor Slough. Water depth and hydroperiod were controlled by a combination of the regional trends in precipitation and substrate elevation. The latter is influenced by bedrock topography, sediment accumulation rates (which also are influenced by vegetation type), and sediment loss from exposure or fire.
Settlement history of two Everglades tree islands
Two teardrop-shaped tree islands with archeological evidence of Native American activity were chosen for this study (Table 1). Heartleaf Hammock (also known as Billboard Hammock; site 1, Figure 1) is approximately 0.8 km long by 0.3 km wide. Archeological test pits along with radiocarbon dating of artifacts indicate human activity since the late Archaic (5000–2500 calibrated years before present or cal. yr BP). The earliest dated artifact is a marine shell dated at 4520–4250 cal. yr BP (Schwadron, 2009). Because this shell could have been transported from the coast at any time since that date, it does not provide definitive evidence for the onset of human occupation. However, archeological evidence does indicate that tree island settlement still could be much older than originally thought (~2500 cal. yr BP; after Widmer, 1988) for the interior of the Everglades (Schwadron, 2006). Near continuous occupation persisted until ~500 cal. yr BP, with a hiatus, or lack of artifacts, during Glades IIIc (900–800 cal. yr BP).
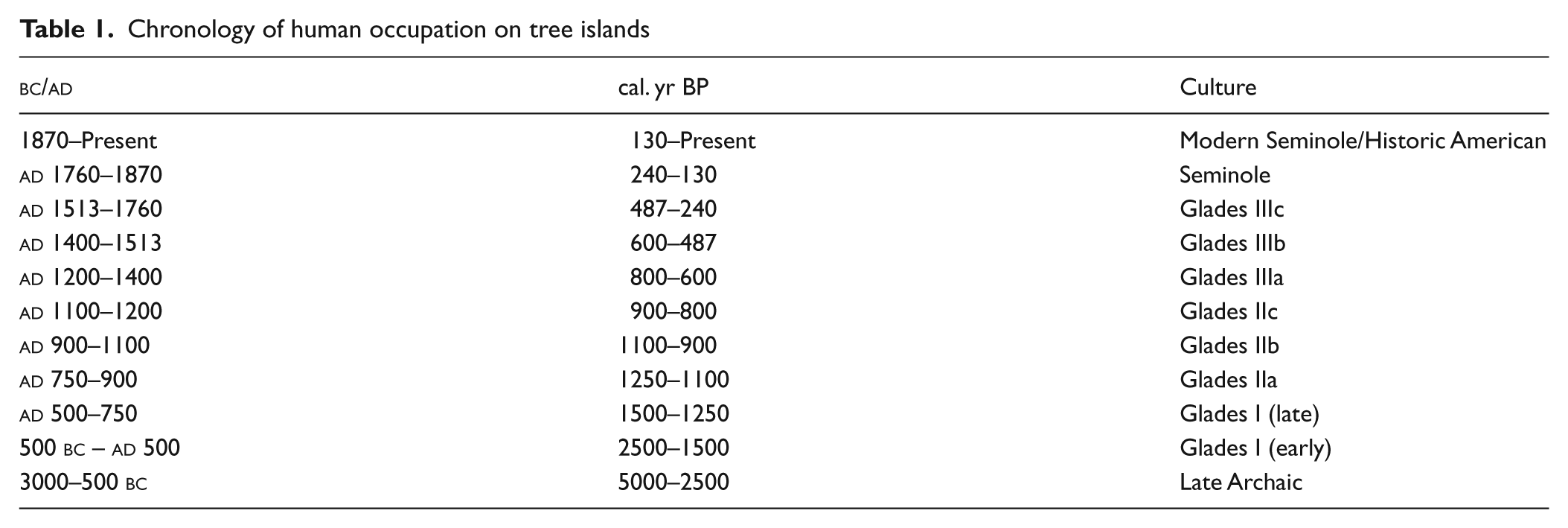
Chronology of human occupation on tree islands
Duck Club Hammock (also known as Sour Orange Hammock; site 2, Figure 1) is approximately 2.4 km long by 0.5 km wide. Human activity dates from the late Archaic to historical time (5000–~50 cal. yr BP). Evidence of human activity is based on archeological material such as middens (Archaic), a kaolin pipe and a glass button (Seminole and historic), and more recently a hunting cabin and picnic tables (nineteenth to twentieth centuries) (Schwadron, 2009).
Methods
Core collection and sampling
A 4 cm diameter vibracore was used to penetrate dry, mineral sediments on the elevated head and reach the underlying bedrock. Cores in near tail sediments were collected using a piston corer with a 10 cm diameter barrel. For all cores, sediment lithology was described, and sediment was sampled for pollen and microscopic charcoal in 1 cm increments for the top 20 cm and in 2 cm increments from 20 cm to the base of the core. All samples were dried in an oven at 50°C.
Geochronology
Chronology of these cores is based on radiocarbon dates (14C) and pollen biostratigraphy. Radiocarbon dates were obtained on bulk sediments by Beta Analytic and calibrated to calendar years using the Pretoria Calibration Procedure (Stuiver et al., 1998; Talma and Vogel, 1993). Dates in Table 2 are presented as calibrated years before
Radiocarbon dates a
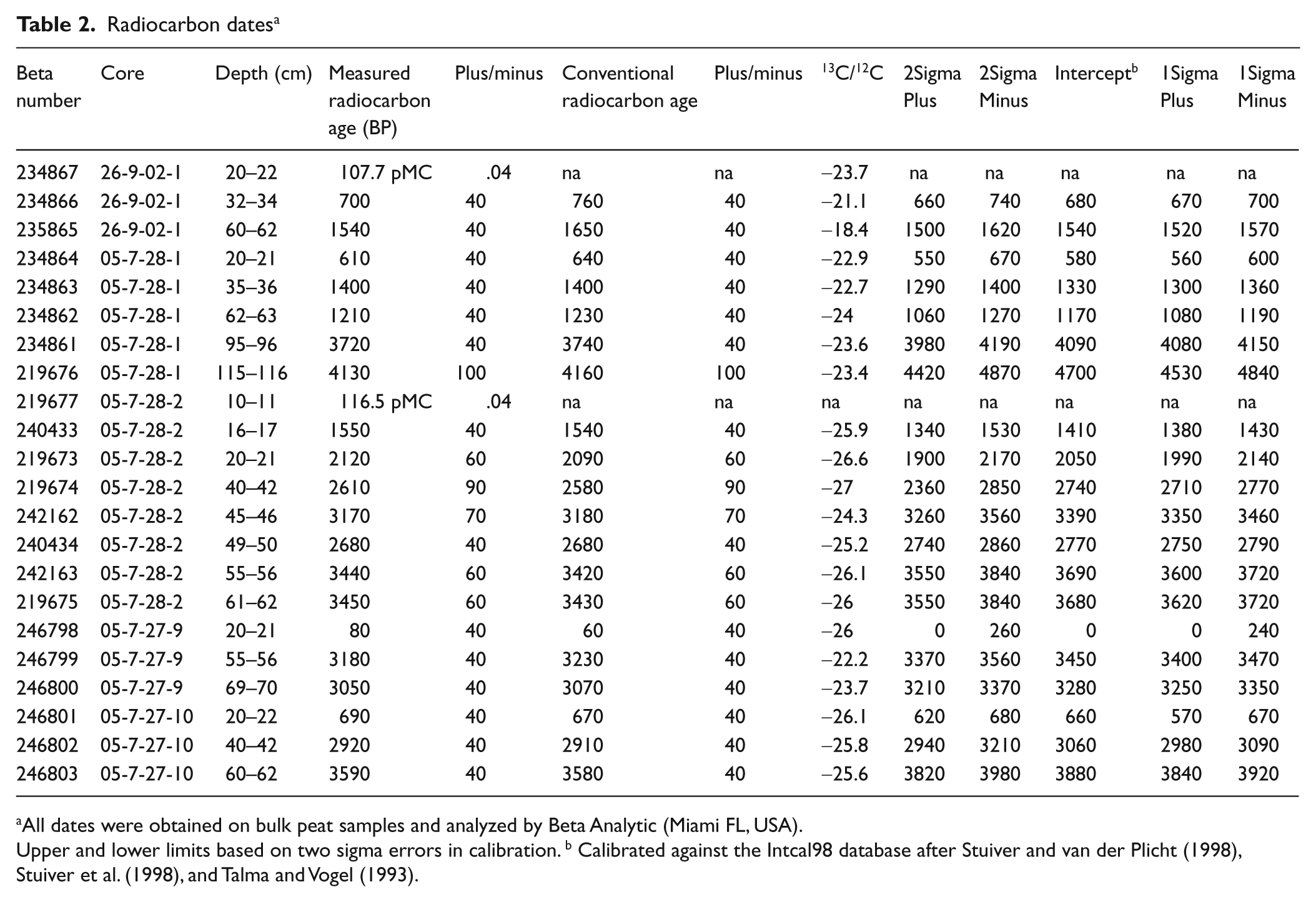
All dates were obtained on bulk peat samples and analyzed by Beta Analytic (Miami FL, USA).
Upper and lower limits based on two sigma errors in calibration. b Calibrated against the Intcal98 database after Stuiver and van der Plicht (1998), Stuiver et al. (1998), and Talma and Vogel (1993).
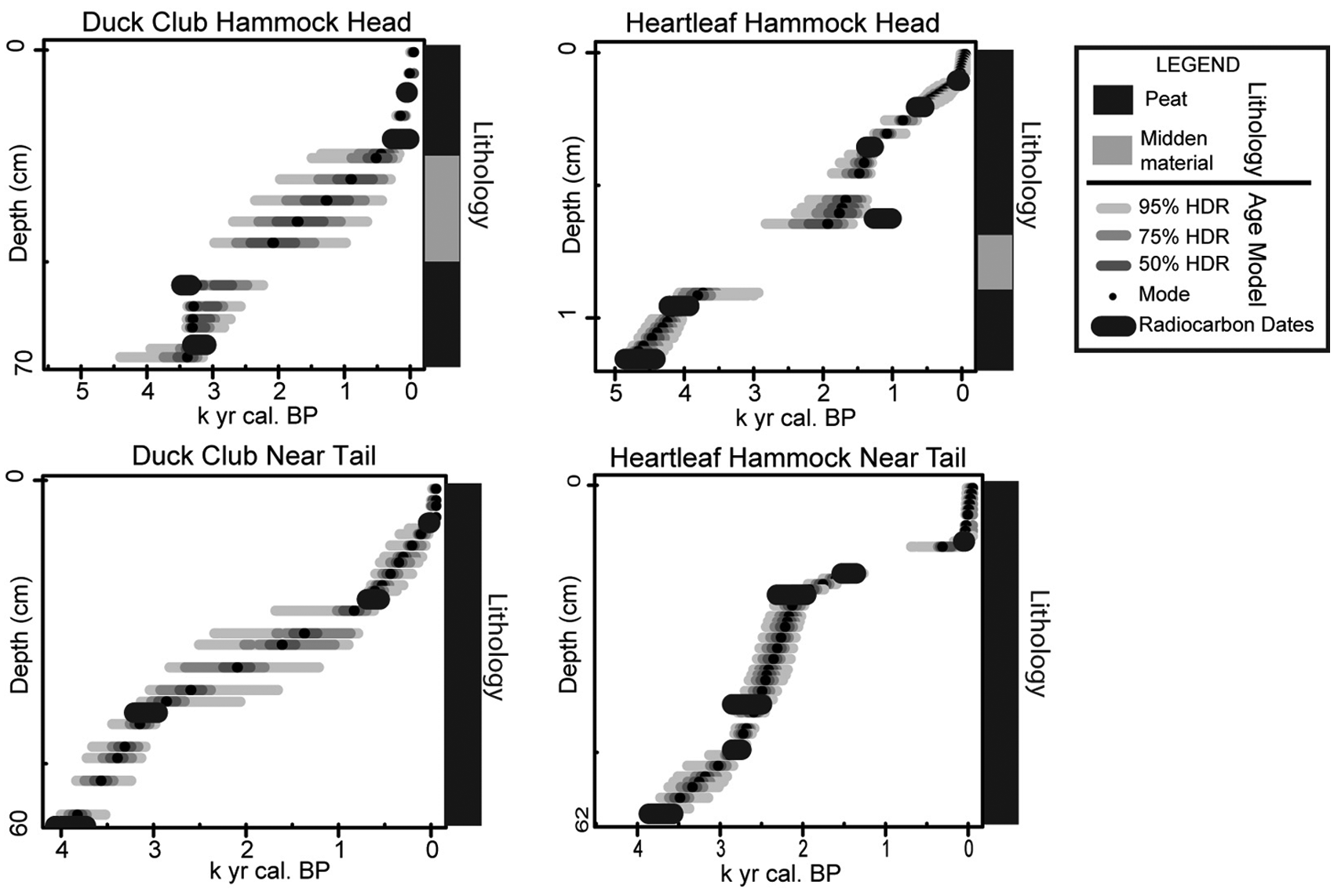
A e models show age plotted in calibrated years before present (
Pollen analysis
Palynomorphs (pollen and fern spores) were isolated using standard palynological preparation techniques (Traverse, 2007; Willard et al., 2001). For each sample, one tablet of Lycopodium spores was added to between 0.5 and 1.5 g dry sediment to calculate palynomorph concentration (grains/g). Samples were processed with HCl and HF to remove carbonates and silicates, respectively, acetolyzed (1 part sulfuric acid:9 parts acetic anhydride) in a boiling water bath for 10 min, neutralized, and treated with 10% KOH for 10 min in a water bath at 70°C. After neutralization, residues were sieved with 149 µm and 10 µm nylon mesh to remove the coarse and clay fractions, respectively. When necessary, samples were swirled in a watch glass to remove mineral matter. After staining with Bismarck Brown, palynomorph residues were mounted on microscope slides in glycerin jelly. In general, at least 300 pollen grains and spores were counted from each sample to determine percent abundance and concentration of palynomorphs. Identification was aided by slides from the United States Geological Survey (Reston, Virginia) reference collection and Willard et al. (2004). Pollen data are archived on the SOFIA (South Florida Information Access http://sofia.usgs.gov) and North American Pollen Database at the World Data Center for Paleoclimatology in Boulder, CO (http://www.ngdc.noaa.gov/paleo/pollen.html).
Charcoal analysis
The metric of charcoal area per pollen grain (C/P) was used to quantify changes in local fire regime. The C/P ratio has been successfully used in other wetlands to reconstruct fire history (see Pederson et al., 2005). Microscopic charcoal particles greater than 50 by 10 µm (500 µm2) were counted on slides made for pollen analysis. This size fraction is considered to represent local fire events within 20 km (Clark, 1988; Pederson et al., 2005). Several samples were barren of pollen and the C/P method could not be used, therefore the absolute concentration of charcoal fragments (greater than 500 µm2) was calculated and compared with the C/P ratio (Figure 3). Charcoal concentration and C/P show similar patterns and therefore only the C/P data are presented with pollen data (Figures 4 and 5).

Microscopic charcoal from tree island sediments. The concentration of microscopic charcoal (fragments ≥500 µm/g sediment) is plotted against C/P (µm2/pollen grain; methodology Pederson et al., 2005). C/P is the dashed line with open squares and concentration is solid black line
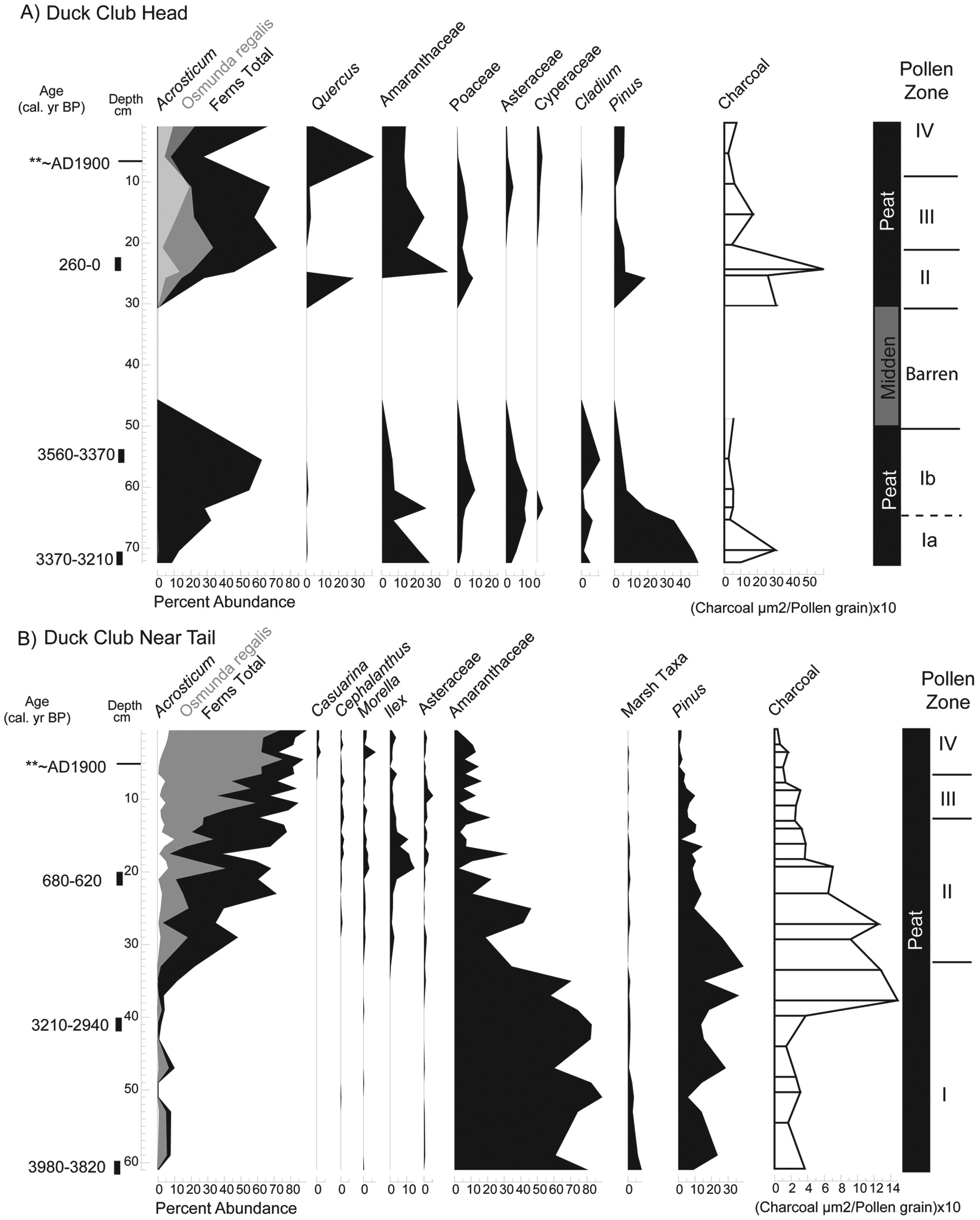
(A) Pollen assemblage from Duck Club Hammock head. (B) Pollen assemblage from Duck Club Hammock near tail. Pollen assemblages are shown for species with greater than 2% abundance. Radiocarbon dates are given as the intercept point and 2-sigma range as calibrated years before 1950 (cal. yr BP), see Table 2. Pollen zone I is analogous to sawgrass marsh/ridge vegetation; Pollen zones II–IV are analogous to tree island vegetation
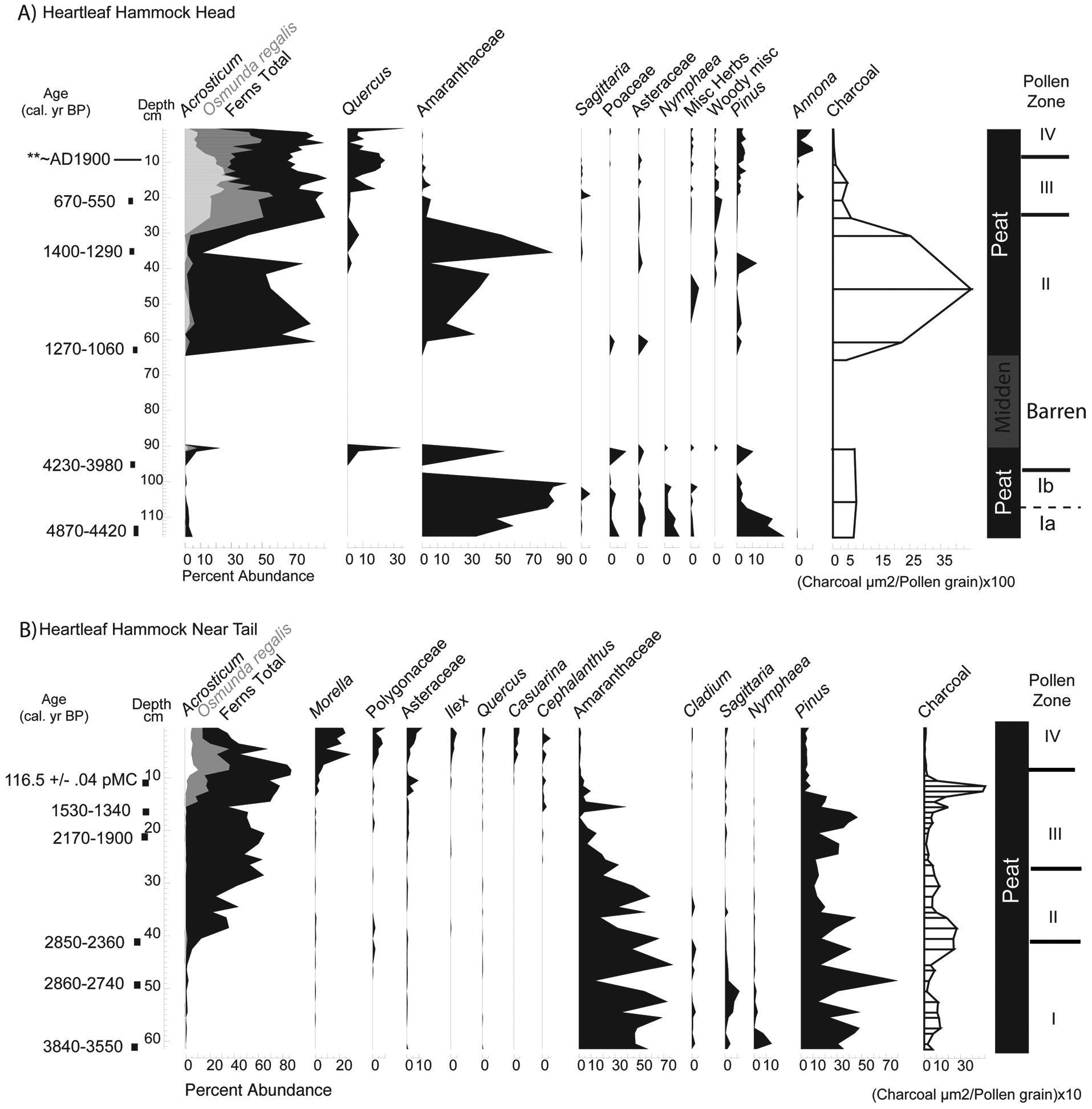
(A) Pollen assemblage from Heartleaf Hammock head. (B) Pollen assemblage from Heartleaf Hammock near tail. Pollen assemblages are shown for species with greater than 2% abundance. Radiocarbon dates are given as the intercept point and 2-sigma range as calibrated years before
Statistical analysis
Reconstruction of past plant communities was based on statistical comparison of fossil and modern assemblages from different wetland communities throughout the Everglades (Willard et al., 2001). Using the modern analog technique (Overpeck et al., 1985), we calculated squared chord distance between down-core pollen assemblages and surface samples collected throughout the Everglades (Willard et al., 2001, 2006; this research) to define the dissimilarity between each fossil and modern assemblage. Squared chord distance was chosen because it is not influenced by rare pollen taxa in the data set (Overpeck et al., 1985). Down-core samples with dissimilarity coefficients equal to or less than the critical value 0.15 are considered to share similar vegetation and environmental parameters with their modern analog (Willard et al., 2001).
Pollen zones were based on a combination of ecological reasoning, visual inspection, modern analogs, and objective zonation using the program CONISS (Grimm, 1987). Zonation is used to assist description and correlation of down-core pollen assemblages within and between cores. The method used to define zones in this study is constrained cluster analysis by method of incremental sum of squares (CONISS; after Grimm, 1987). In this method of numerical zonation, clusters are constrained to only have stratigraphically adjacent samples. Resultant zones are defined and plotted next to pollen diagrams.
Pollen assemblages and source vegetation
Tree island vegetation is characterized by high abundance of fern spores (>40%). Acrostichum spores, as well as Blechnum and Thelypteris, typically dominate head assemblages, whereas tail assemblages are characterized by high abundances of Osmunda spores (Willard et al., 2006). Amaranthaceae (>20%), Cyperaceae (5–30%), Nymphaea (>10%) pollen and low fern spore abundances (<30%) characterize marsh/ridge-slough vegetation. Pinus pollen is transported in from the surrounding areas. Morella, Asteraceae, Cephalanthus, Quercus, and Ilex are taxa indicative of drier conditions (Bernhardt and Willard, 2009; Willard et al., 2001, 2006).
Results
Duck Club Hammock
A vibracore recovered from the elevated dry head is 73 cm long and has a chronology provided by three radiocarbon dates and the first occurrence of Casuarina pollen at 5 cm (Figure 4A and Table 2). The base of the core dates to 3370–3210 cal. yr BP. On Duck Club Hammock, previous studies identified a calcrete layer at approximately 50 cm (Schwadron, 2009), but this core did penetrate 100 cm of sediment before hitting bedrock. The sediment consists of 30 cm of peat overlain by an organic, sand-sized faunal midden layer from 30 to 47 cm (Figure 4A). The midden material is primarily charred animal bone. No seeds or macroscopic plant material are present and this interval is barren of palynomorphs. The upper 30 cm of the core, directly above the midden layer, consists entirely of black peat. A peak in microscopic charcoal (500 µm2/pollen grain) is present above the midden layer at 29–25 cm. Pollen is well preserved in all samples except those in the midden layer, which were barren.
A piston core recovered from the near tail is 60 cm long and composed entirely of peat (Figure 4B). Three radiocarbon dates (Table 2) and the first occurrence of Casuarina pollen at 5 cm provide the chronology. The base of the core dates to 3980–3820 cal. yr BP. A microscopic charcoal peak (145 µm2/pollen grain) is recorded at 39 cm above a radiocarbon date of 3210–2940 cal. yr BP. The amount of microscopic charcoal steadily declines upcore. Pollen is well preserved throughout the core.
Heartleaf Hammock
A vibracore recovered from the elevated, dry head is 116 cm long and composed of black peat and a sandier midden layer (Figure 5A). Chronology is based on five radiocarbon dates (Table 2) and the first occurrence of Casuarina at 10 cm. The midden was deposited sometime between 4230–3980 cal. yr BP and 1270–1060 cal. yr BP. The faunal midden of sand-sized charred animal fragments occurs at 73 to 90 cm. This material is primarily composed of broken and burnt animal bone, turtle shell, and animal teeth, and it is barren of pollen. No seeds or macroscopic plant material were found in either core. The basal sample dates to 4870–4420 cal. yr BP. Microscopic charcoal is most abundant above the barren interval/midden layer (~4000 µm2/pollen grain). Pollen was well preserved except in the midden layer.
A piston core from Heartleaf Hammock near the tail is 62 cm long and comprised entirely of peat. Six radiocarbon dates and the first occurrence of Casuarina at 13 cm provide the chronology (Figure 5B). The base of the core dates to 3840–3550 cal. yr BP. A hiatus in deposition corresponding to a peak in microscopic charcoal is indicated at 13 cm (480 µm2/pollen grain) dating to within the last century of deposition (presence of Casuarina pollen). The hiatus is at least 1000 years. Another peak in microscopic charcoal (~240 µm2/pollen grain) around 40 cm dates to 2860–2740 cal. yr BP. Pollen is well preserved throughout the core.
Pollen zones
Pollen assemblages for all cores (Figures 4–5), have four distinct assemblage zones. Basal pollen zone Ia on the heads of Duck Club and Heartleaf Hammock is analogous to sawgrass marsh/ridge vegetation, characterized the common occurrence (combined percentages >10%) of Nymphaea, Cyperaceae (including Cladium), Poaceae, and Sagittaria pollen. Zone Ib is also analogous to marsh vegetation but contains a rise in fern spores. Basal near tail sediments also are analogous to sawgrass marsh sites. Near tail assemblages in zone I of Duck Club and Heartleaf Hammocks are dominated by Amaranthaceae pollen (70–90%) and Pinus.
In cores from heads of Heartleaf and Duck Club Hammock, sediments deposited between pollen zones I and II are completely barren of pollen. The barren units are ~25 cm thick and are composed of sand-sized faunal midden material. In pollen zone II on tree island heads, fern spores dominate assemblages (~60–70%) and are analogous to tree island vegetation. Amaranthaceae pollen typically comprises ~20% of assemblages, except in Heartleaf Hammock head where there is one peak (90%) at 35 cm. Peat accumulation was continuous across the zone I to II transition in near tail cores and is dated ~2800 cal. yr BP. Amaranthaceae pollen decreases below 30% in near tail sediments, and pollen of typical marsh plants (Sagittaria, Nymphaea, and Cyperaceae) is nearly absent. One difference between near tail sites is the presence of Ilex after ~600 cal. yr BP in Duck Club. Pollen zone II in Heartleaf and Duck Club Hammocks represents initial occurrence of tree island vegetation.
Zone III on both tree island heads is characterized by increases in Osmunda regalis and Acrostichum and by fern spores in general in near tails of all tree islands. Assemblages are analogous to mature tree island tails and are dominated by Osmunda spores (>70%).
Zone IV includes sediments deposited during the twentieth century, based on the occurrence of modern carbon and Casuarina pollen. Osmunda increases in all sites during by the late twentieth century. Woody vegetation such as Cephalanthus, Annona, Quercus and Morella reach their highest abundances in zone IV.
Discussion
Evolution of tree island vegetation
This study documents pre-tree island vegetation from modern tree island heads, evaluates the timing of tree island formation, and confirms earlier interpretations based on near tail sediments (Willard et al., 2006). Pollen assemblages from basal sediments recovered from tree island heads and near tails pre-date or are concurrent with earliest evidence for human occupation (Schwadron, 2009). These pre-occupation records lack abundant fern spores characteristic of tree islands and are dominated by taxa indicative of sawgrass marsh/ridge vegetation, including Amaranthaceae, Cyperaceae and Nymphaea pollen (Figures 4 and 5). The near absence of fern spores in near tail sediments suggest that the vegetation preceding the formation of the tree island was marsh-like and is consistent with results from previous studies (Willard et al., 2006).
Both Heartleaf and Duck Club Hammock begin to develop around 3800 cal. yr BP (Figures 4–6). Age estimates of tree island initiation and expansion are based on calculating the approximate age at which fern spores abruptly increase for all cores (Figure 6). A radiocarbon date on a marine shell on Heartleaf Hammock head, 4520–4250 cal. yr BP, indicate Native American activities on the islands after or concurrently with the initial formation of these islands (Schwadron, 2009). Faunal middens, barren of pollen, were deposited on Duck Club Hammock after initiation of tree island communities, and the dates of midden accumulation on Heartleaf Hammock are unclear. Microscopic charcoal peaks are recorded above each midden layer. Intense local burning and disturbance of the soil oxidized pollen and destroyed the record of community composition during this period of occupation. Pollen is readily destroyed by oxidation (Traverse, 2007). Middens may have further raised the surface elevation above the adjacent marshes, increasing colonization by woody tree island vegetation. However, the absence of pollen on the heads leaves us unable to address this hypothesis. In cores from the near tails, sedimentation appears to have been continuous and indicates tree island establishment in the near tails by ~2800–2600 cal. yr BP for Duck Club and Heartleaf Hammocks (Figure 5). However, the timing of this change is not uniform across the Everglades. A well-dated tree island examined by Willard et al. (2006) indicates tree island vegetation was established ~1000 cal. yr BP for Manatee Hammock. Comparison of head and near tail records from Duck Club Hammock indicate that establishment of tree island vegetation in the near tail lagged that on the head by 1000 years (Figure 5). Archeological excavations on both Duck Club and Heartleaf Hammock record continuous Native American occupation during this time period (zone III) (Schwadron 2009; Figures 3–5).
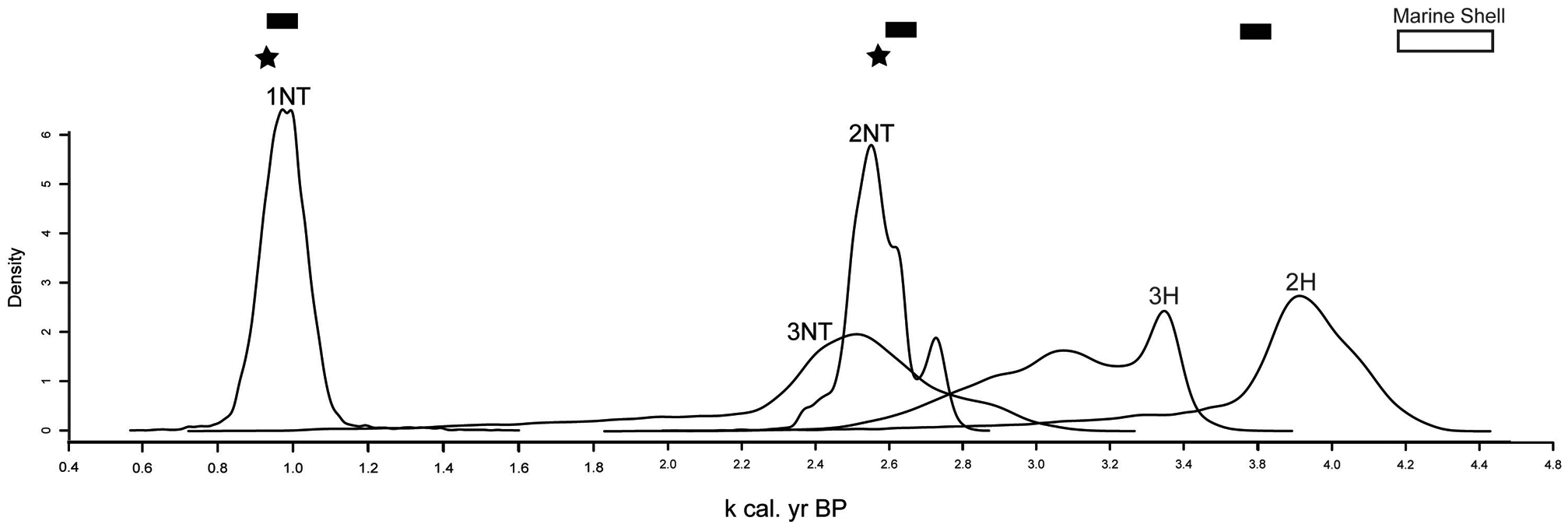
Probability distributions (posterior distribution) for the age of tree island initiation and expansion. Initiation and expansion of tree islands is based on the rise in fern spores. 1NT, Manatee Hammock near tail (Willard et al., 2006); 2NT, Heartleaf Hammock near tail; 3NT, Duck Club Hammock near tail; 2H, Heartleaf Hammock head; 3H, Duck Club head. Bars represent intervals of circum-Caribbean drought (Curtis and Hodell, 1993; Hodell et al., 2001; Polk et al., 2007). Stars represent time periods where shifts in Native American cultures in the Everglades are attributed to climate (Griffin, 1988). A white rectangle represents the calibrated age range of a marine shell interpreted as an archeological artifact recovered from Heartleaf Hammock (Schwadron, 2009)
Tree island head vegetation changes are recorded ~1000 cal. yr BP on Duck Club and Heartleaf Hammock (Figures 3 and 4). Both Acrostichum and Osmunda increase in both head and near tail sediments (Figures 3 and 4). This corresponds to a period of regional tree island maturation and changes in ridge and slough vegetation (Bernhardt and Willard, 2009; Willard et al., 2006).
Climate as a driver of vegetation change
Based on the timing of changes in pollen assemblages, this study suggests that that climate was a major factor initiating tree island growth and expansion. Wetzel et al. (2008) hypothesize that extreme hydrologic events such as drought are more important than average annual conditions in shaping tree island vegetation, whereas short-term (<10 years) flooding or perturbations do not alter vegetation composition (Zaffke, 1983). During dry season and droughts, nutrients, especially phosphorus, are redistributed from the adjacent marshes to the island through increased evapotranspiration by trees on the island (Ross et al., 2006; Wetzel et al., 2005). Increased soil phosphorus concentrations enhance peat accumulation rates, increasing tree island elevation relative to the adjacent marsh (Ross et al., 2006; Wetzel et al., 2008). Drought also allows recruitment of woody vegetation from the seed bank (Wetzel, 2002). Establishment of woody vegetation attracts more birds and other animals to the islands and further increases and concentrates phosphorus on the island (Orem et al., 2002). Overall, this positive feedback mechanism, driven by climate, enhances tree island growth and expansion. Fire is also a key factor in shaping tree island vegetation (Wetzel, 2002), and the occurrence and intensity of fires in the Everglades would increase as a result of periodic climate-driven droughts (Beckage et al., 2003). This study finds microscopic charcoal peaks correspond to periods of tree island expansion (Figures 5 and 6). It is probable that local fires induced by these prolonged periods of aridity further facilitated tree island expansion.
Initiation of tree island growth (Figure 6) is concomitant with regional periods of aridity at ~3800, ~2800, and ~1000 cal. yr BP in the circum-Caribbean (Curtis and Hodell, 1993; Hodell et al., 2001; Polk et al., 2007) and in South Florida (Alvarez Zarikian et al., 2005). Extended periods of aridity are likely influenced by the mean southward migration of the ITCZ (Haug et al., 2001). Precipitation through the late Holocene on the Florida peninsula is also influenced by changes in intensity in the North Atlantic Oscillation (NAO) and the El Niño Southern Oscillation (ENSO) (Cronin et al., 2002; van Beynen et al., 2007, 2008). Tree island expansion occurs during periods of regional aridity centered around ~ 2800 and 1000 cal. yr BP. Microscopic charcoal increases during these periods of regional aridity and tree island expansion. Tree island vegetation matures around the ~1000 cal. yr BP aridity event, which is consistent with regional trends in other tree islands and Everglades’ communities (Bernhardt and Willard, 2009; Willard et al., 2006).
Potential effects of climate variability on Native Americans and tree island vegetation
Cultural shifts in various civilizations around the globe have been linked to periods of extreme aridity (Brooks, 2006). For example, the collapse of the Mayan civilization (~1200 years ago) is attributed to extended periods of drought and changes in the mean position of the ITCZ (Haug et al., 2003; Hodell et al., 1995). Intervals of regional aridity have influenced Native American occupation patterns in Florida as far back as ~12 000 cal. yr BP (Faught and Carter, 1998). In the Everglades, the transition between Native American cultures has been indirectly related to both climate stability and change (Griffin, 1988). The cultural transition between the Archaic and Glades cultures (~2500 cal. yr BP) occurred during a period of regional and global aridity as well as changes in tree island vegetation (Figure 5).
Anomalous tree island abandonment during Glades IIc (~800–900 cal. yr BP) based on the absence of artifacts from 56 of 62 sites has been interpreted as a human response to changes in regional in climate (Griffin, 1988). The lack of artifacts has also been interpreted as a change in the cultural development and introduction of new peoples into the region (Carr, 2002). This period of cultural change coincides with tree island initiation and expansion throughout the Everglades, as driven by regional climate (this study; Willard et al., 2006). Speleothem records from Central Florida document a severe drought during this time period (van Beynen et al., 2008). Tree islands were used for camps primarily during the winter. Changes in the strength of the North Atlantic Oscillation and El Niño, which influence winter precipitation in South Florida (Swart et al., 1996), could have made conditions unfavorable for tree island occupation. These data leave open the possibility for climate fluctuations playing a role in the cultural shifts recorded in the Everglades.
Native American and modern land use as a driver of tree island vegetation
Native American activities have been proposed as mechanisms of tree island initiation and growth (Carr, 2002). Deposition of thick (10–30 cm) faunal middens and the collapse of wooden platforms have been hypothesized as loci for sediment accumulation, raising the elevation of the surface above the adjacent marsh, and initiating the establishment of woody taxa (Carr, 2002). Native camps also may have altered the canopy structure on the islands, similar to reductions documented on sites of modern camps (Heisler et al., 2002). Further, Native Americans are thought to have used low intensity fires to modify the landscape (Robertson, 1953). Pollen evidence indicates that, at least at these sites, tree island vegetation pre-dates the appearance of Native American artifacts. However, this hypothesis requires further coordinated sampling for both pollen and artifacts with precise dating of both sediments and artifacts.
The change in vegetation during the first millennium of Native American occupation (for example the period between ~3800 and 2600 cal. yr BP) appears to be minor compared with rapid changes documented in the last century of land use (Figures 3 and 4). Because humans did not permanently occupy tree islands, only during winters as temporary fishing and hunting camps (Carr, 2002), the overall development of the island does not appear to have been drastically altered.
The compartmentalization and alteration of the natural hydrology in the twentieth century had a larger impact on tree island vegetation in a shorter time period than the collective impacts of the occupation history of Native Americans. The effects of twentieth-century water management on tree islands are well documented (Heisler et al., 2002; Wetzel, 2002, 2008; Willard et al., 2001, 2006) and the construction of roads perpendicular to flow, including Alligator Alley and Tamiami Trail, compounded the alterations to the natural hydrology. The associated prolonged flooding and deeper water depths in parts of the system (particularly WCA 2A and southernmost 3A, 3B) killed subtropical hardwood trees, creating many ‘ghost tree islands’. In contrast, shorter hydroperiods increase the tree island’s vulnerability to fire (Sklar and van der Valk, 2002). Vegetation on Heartleaf Hammock reflects these drier conditions, as seen by the increase in Morella, Quercus, Asteraceae, and Ilex. Fire is important in shaping tree island vegetation (Wetzel, 2002), and fire suppression could be a factor in the distribution of modern vegetation. Three of the four sediment cores examined for this study indicate a reduction in fire as evidenced by the overall decrease in microscopic charcoal abundance during the twentieth century. The increase in concentration of charcoal on Duck Club during the twentieth century (Figure 3) could be due to the fact that people have turned this island into a recreation site, as seen by the cabin and manicured landscape on the head.
Conclusion
Pollen analysis of tree island sediments indicates that the tree island vegetation developed from a marsh environment on the heads. Initiation of head vegetation on these two tree islands is ~ 3800 cal. yr BP. Near tail sediments indicate a ~1000 year lag, after the formation of the initial tree island, in the development of neat tail vegetation. Native American occupation of tree islands occurred after and/or concurrent with the onset of tree island formation, during a period of regional aridity. Faunal middens as thick as ~27 cm on tree island heads appear to post-date the initiation of tree island vegetation, and their presence could have had an influence on the elevation of head sediments in relation to the adjacent marsh. Because of (poor) pollen preservation in midden sediments, it is difficult to identify vegetation assemblages during the accumulation of these sediments. Near tail sediments do not record any changes in vegetation during the same interval of occupation. Tree island expansion occurred during a period of regional aridity throughout the circum-Caribbean thought to be driven by the mean southward position of the ITCZ. During the onset of twentieth-century water management further changes in tree island vegetation are recorded. Changes in tree island vegetation, associated with climate variability ~2600 and 800 cal. yr BP, may have influenced cultural change in the Everglades. In order to further clarify the role of Native Americans on tree island vegetation, more accurate dating of artifacts is needed to correlate changes in vegetation with human activities.
Footnotes
Acknowledgements
This research is supported by the USGS Greater Everglades Priority Ecosystem Studies Program and was carried out in partial fulfillment of a Ph.D. at the University of Pennsylvania, Department of Earth and Environmental Sciences. The author is extremely grateful to the Everglades National Park for site access. Margo Schwadron provided key guidance in site selection as well as field assistance. Thomas Sheehan, Bryan Landacre, and Adam Bentham assisted with field and laboratory work. Drs D. Willard and B. Horton helped guide this research. Drs L. Wingard, T. Cronin, F. Scatena, and H. Pfefferkorn and two anonymous reviewers provided invaluable comments that greatly improved the quality of this manuscript.