
Abstract
Coastal dune fields and dune-body associated lakes were studied through sedimentological, malacological and palynological analyses. Lake deposits and palaeosoil horizons gave new information about phases of aeolian activity and dune building, coastline and sea level, vegetation history and cultural impact. After a period of intense aeolian activity (Late Pleistocene) that limited soil-forming processes, a wet aeolian field, recorded by podzolized sands and interdune wetland deposits, was present in the region. Around 5000 to 4000 yr BP sea level reached the current level and the coastline was situated c. 1 km further inland compared with the present situation. Subsequently lagoon-barrier island systems developed c. 4000 yr BP. Vegetation in the region during this time was a pine/oak forest. Between 4000 and 1600 yr BP the forest was replaced by a semi-natural heathland. Later the heathland was degraded, allowing dune migration and finally formation of a desert-like landscape. This process was probably a consequence of a combination of deteriorating climate during the ‘Little Ice Age’ (LIA) and overgrazing. To counteract sand invasion, reforestation efforts were initiated in the region c. 300 years ago and locally along the coastal margin between 1924 and 1940. Formation of the row of freshwater lakes bordering the dune field at present was contemporaneous with the initial reforestation phase. A close relation between lake formation and accumulation of sand dunes was found. Various types of dunes were identified, truncated dunes, parabolic dunes and transverse/oblique dunes. Dune migration caused an eastward displacement of lakes.
Introduction
An extensive coastal dune field occupies the zone between Quiaios and Tocha in central littoral Portugal. The development of this dune field was episodic and influenced by conditions related to climate, sea level and sediment input, vegetation cover and human activity. Stratigraphy of the dunes records environmental and cultural changes during their evolution. Several sandy layers from periods of sand drift are interleaved with wetland deposits and palaeosols formed under more stable conditions.
A previous investigation from the region has indicated that aeolian activity in western Iberia seems to be out of phase with the rest of Europe (Clarke and Rendell, 2006). Sand invasion and dune accretion is linked with storminess, which is thought to be influenced by the North Atlantic Oscillation (NAO) (Clarke and Rendell, 2009; O’Connor et al., 2011). North of Iberia positive NAO winters cause northward displacement of westerly storms and bring wet and windy weather. On the contrary, rainy westerlies over the Iberian Peninsula and the western Mediterranean seem to be caused by negative NAO (Clarke and Rendell, 2006; Luterbacher et al., 2002). High aeolian activity is hence to be expected in littoral Portugal during periods of low activity in the rest of western Europe.
Episodic sand invasions have caused recurring problems for human activity and agriculture along the Portuguese littoral and stabilization measures to reduce this threat initiated already in the thirteenth century when a royal decree ordered the planting of forests of maritime pine (Pinus pinaster). The remedy was effective in stabilizing the sand dunes and littoral Portugal nowadays is characterized by planted pine forests. Little is known, however, about past natural vegetation cover and dynamics of vegetation history in the region.
The main objective of this investigation is to disclose the genesis and evolution of dune-related Holocene deposits of the coastal Quiaios–Tocha region. Lithofacies, based on texture, composition and geometry of the sediment bodies is associated with fossil content (pollen and molluscs). The palaeoenvironmental conditions and evolution are inferred from these sets of data.
Study site
The investigated area is located in central Portugal, north of Serra da Boa Viagem (Figure 1A), a coastal mountain with Jurassic carbonate and siliciclastic rocks. The coastal margin to the north is covered by Late Pleistocene and Holocene coastal deposits. The Holocene dunes extend up to 7 km inland from the coastline. A row of shallow lakes is found at the border between these dunes and an older and more abraded dune field, the so-called Gândara dunes (Carvalho, 1964). The younger dunes morphology is still preserved in many cases. The foredune is a fairly continuous ridge whose growth was enhanced, through the setting up of a wooden fence along the whole extension of the coastline. This intervention occurred during the 1920s when sand traps were used to prevent the burying of local pine plantations by aeolian sand (Almeida, 1995; Rei, 1940). Inland, behind the foredune, the most common dune forms are elongated dunes classified both as oblique (Almeida, 1995) and transverse (Clarke and Rendell, 2006; Noivo, 1996;). The best-defined forms are aligned almost E–W, can reach lengths of more than 1 km and are spaced approximately 200 m apart (Figure 1B). These dunes were still active at the beginning of the twentieth century and they were stabilized by reforestation between 1924 and 1940 (Pestana, 1926; Rei, 1940). At present they sustain a pine forest dominated by Pinus pinaster. The present-day parallel fluvial drainage pattern outlines and is influenced by the orientation of these dune forms. Throughout the dune field several seasonally inundated pools can be found in lower areas between the dune ridges. Furthermore, sectors with parabolic dunes are found scattered in the region (Almeida, 1995; Clarke and Rendell, 2006; Noivo, 1996). The parabolic dunes are located in the vicinity of Lake Braças and Lake Vela and are also found immediately north of the village of Quiaios (Figure 1) (Almeida, 1995; Barbosa et al., 1988). These dunes show indications of belonging to an earlier dune generation than the oblique/transverse dunes, not only on account of their dune form but moreover because of their further developed and acid soils. This permitted the development of a heathland vegetation community comprising Calluna vulgaris, Erica spp. and Lavandula pedunculata, an undercover vegetation type practically absent from the more recent dunes which are dominated by Corema album and Halimium sp. (H. halimifolium and H. calycinum).
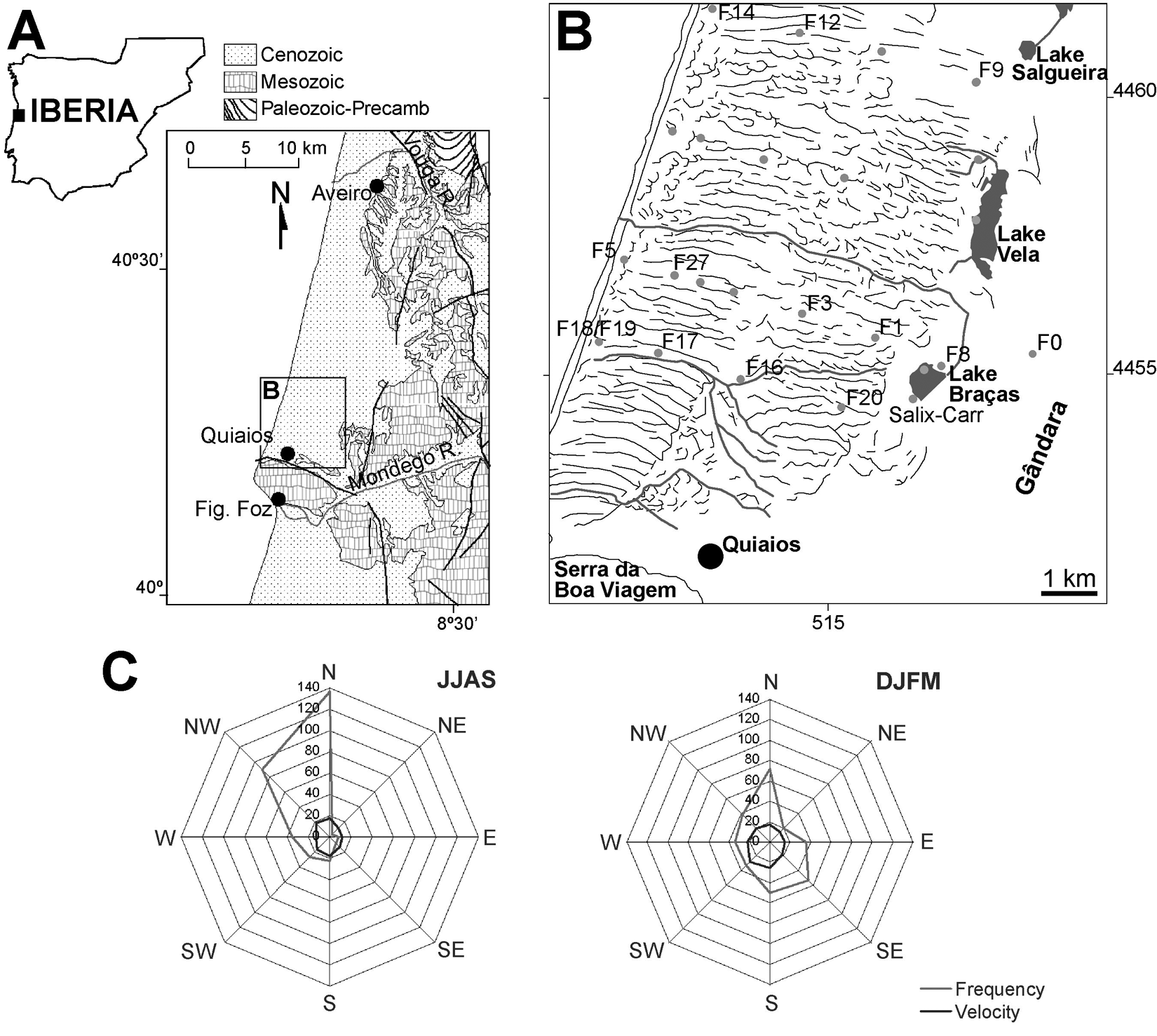
Geological and geographic setting of the studied area. (A) Location in West Iberia. (B) Representation of the dune ridges (black lines), streams (grey lines) and position of studied boreholes and cores. (C) Wind roses of S. Jacinto (W of Aveiro) for summer (JJAS) and winter (DJFM) months. Frequency is the number of records and velocity in km/h (2× exaggerated) (source: Meteorological Institute, Portugal)
The region is dominated by Mediterranean climatic characteristics envisaged by monthly variations of both precipitation and temperature, comprising three hot and dry months (June, July and August). Annual precipitation is around 1000 mm, however, with considerable irregularity from year to year. Relative air humidity is always higher than 74%. The maritime influence causes mild winters and cool summers with mean annual temperature maxima of 19.5°C and minima of 9.2°C. North to northwest winds are most frequent especially in the hottest months (Figure 1C). In the colder months winds from the southwestern, southern and western quadrants are the strongest and most frequent.
Methods
Fieldwork
A geophysical survey was carried out prior to the drilling phase (Castilho et al., 2007a). The electromagnetic data facilitated the choice of borehole sites along selected transects approximately perpendicular to the coastline (Figure 1B). The drilling method chosen (cable tool manual percussion method) is suitable for unconsolidated sediments and can operate above and beneath the water-table with little chance of contamination (Driscoll, 1986). The borehole is stabilized by a casing that is previously driven down (1 m each time) and a sand bailer (suction bailer) that operates inside the casing to cut the borehole and recover the sediments. The operator is able to maintain close contact with the drilling process and the sediments may be continuously assessed. Sediment samples were collected systematically every 40 cm for sedimentological and palynological analysis or whenever the operator noticed changes in the encountered material. Additionally, cores from two lakes (Lake Vela and Lake Braças) were extracted with a 1 m long Livingstone piston corer with 6 cm diameter. The lake cores were respectively 84 cm and 68 cm long (see Figure 5). Deeper coring was obstructed by abundance of sand.
Chronology
Thirteen samples were selected for 14C dating (Table 1). Four were from the lake cores and the remaining ones were from the borehole samples. Dating was performed by Nasjonallaboratoriet, NTNU, Norway (TUa), BETA Analytic Inc., USA (Beta) and Instituto Tecnológico e Nuclear, ITN, Portugal (Sac). Dates were calibrated with the CALIB 5.0.1 software package (Stuiver and Reimer, 1993) using the INTCAL04 data set (Reimer et al., 2004). For brackish shells the Mixed marine NoHem (Hughen et al., 2004) was used.
14C datings
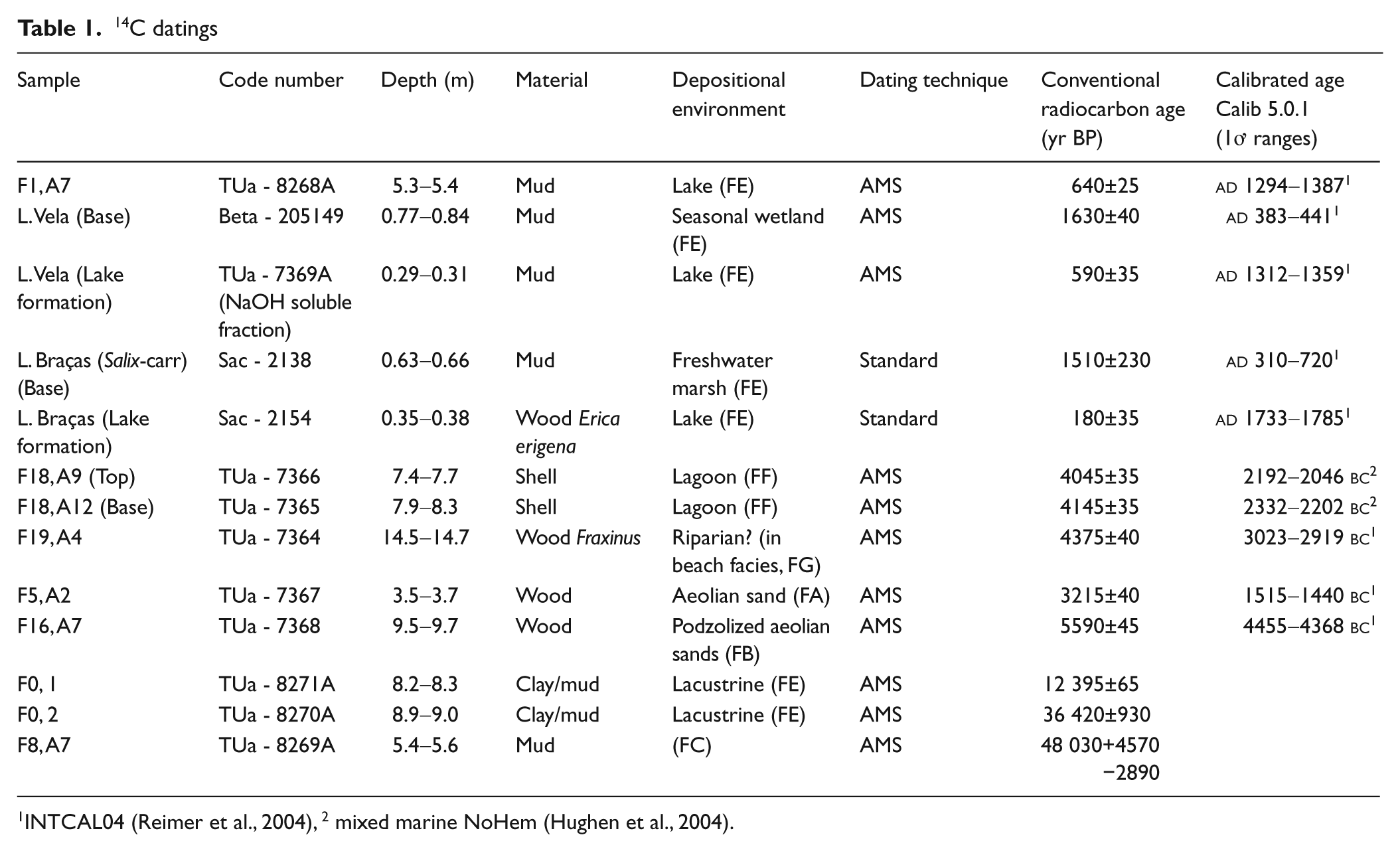
INTCAL04 (Reimer et al., 2004), 2 mixed marine NoHem (Hughen et al., 2004).
Sedimentology
Because sedimentary structure is not preserved in the collected samples, facies definition is based on sediment texture, biological content (pollen and molluscs), thickness and colour. The grain-size of a total of 181 samples from the boreholes was determined by laser diffraction using a Coulter LS 230 equipment. Sediments with more than 5% of particles coarser than 2 mm were also sieved. The bulk grain-size results obtained by both sieving and laser diffraction, were integrated on a conceptual scale with 1/2Ø intervals based on a logarithmic transformation of the particle diameter (D), in mm (Ø=−log2D) (Krumbein, 1934; Wentworth, 1922). The colour of air dried sediment samples was measured by comparison with the Munsell Soil Color Charts (Munsell, 2000). The nomenclature used indicates the colour name and its Munsell notation.
Palynology
Pollen samples were extracted from organic deposits. Nine were from organic layers in the boreholes, and 62 from the two lake cores. Samples from the lake cores were collected with c. 2–5 cm intervals. This accuracy was not possible for the borehole samples. Owing to methodological limitations during drilling of the boreholes, each sample contained large amounts of sediment (20–40 cm) and hence a maximum of two pollen samples was retrieved from each layer (see Figure 3). The pollen samples were subjected to standard laboratory treatment including HF exposure for 48 h (Fægri and Iversen, 1975).
Pollen and microfossil data of the samples are presented in percentage pollen diagrams. Minimum sum of regional pollen was 250 whenever possible. All data were calculated as percentages of total regional pollen (TRP) including pollen of dry-soil taxa with the exception of Pinus and taxa occupying the lake margins. Pollen of Pinus, local taxa (mainly hygrophilous) and non-pollen palynomorphs (NPP) are expressed as percentages of TRP. The pollen diagrams were constructed using Tilia and Tilia graph (Grimm, 1991–1993) with five times exaggeration curves. Stratigraphy of the lake core is presented using the Troels-Smith (1955) classification system.
Organic facies contained pollen, spores and other microfossils and the assemblages of indicator species suggest different palaeoenvironments as indicated in the diagrams. The diagrams from the lake cores reveal the same regional vegetation changes and the diagram from Lake Vela was chosen to represent the region.
Results
Sedimentology
The studied sediments have seven different facies defined by multiple criteria (Figure 2). Grain-size distributions and biological content are the fundamental attributes for the facies definition.
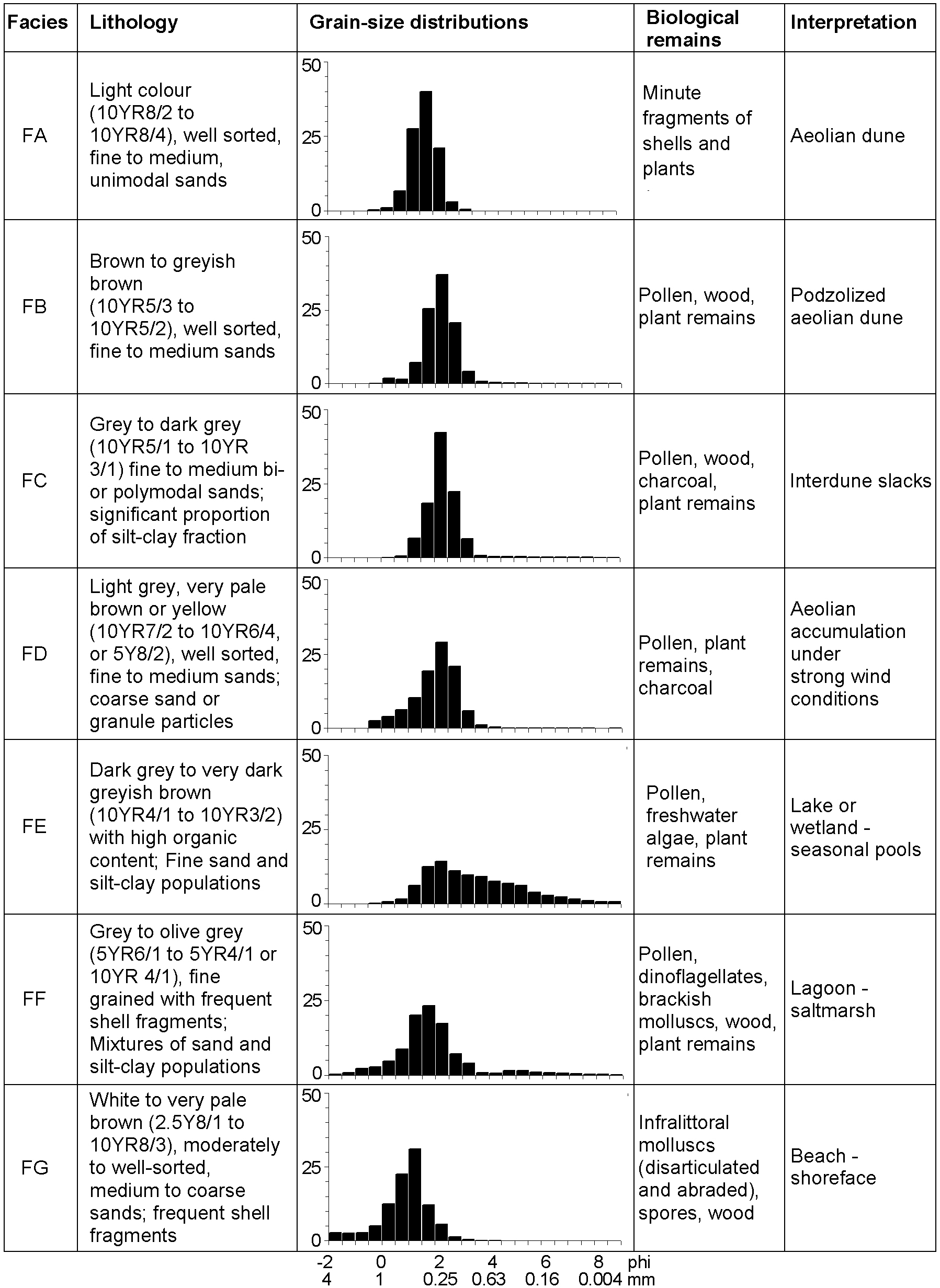
Sedimentary facies characteristics
Facies FA–FD
The most frequent facies (FA, FB, FC and FD) are characterized by well or very well sorted sandy sediments with limited amounts of gravel, silt and clay and unimodal grain-size distributions. The modal size and mean are always in fine to medium sand grain-size classes (0.15–0.5 mm). Facies FA consists of light grey (in particular close to the topographic surface) or very pale brown (in lower stratigraphic levels) sands. It is found widespread along the studied coastal margin, constituting the most common sediment. In some locations the entire Quaternary record shows this facies. However, they are frequently intercalated with facies FB, FC and FD.
Darker, brown or greyish brown, podzolized sands of facies FB show just slight grain-size distribution differences from facies FA, such as higher proportion of silt-clay-size fraction, and finer mean. The podzols are usually incipient and these sediments still tend to be friable. Up to two levels of facies FB are observed in some locations.
Facies FC is distinguished by the presence of significant proportion of a silt-clay size fraction (up to 6%) and its darker greyish colour because of the presence of organic matter. They define relatively thin (a few cm) beds and are frequently associated with podzolized sands (FB). These sediments contain a diversified pollen association that includes wetland species (see below).
Facies FD contains higher amounts of coarse sand and gravel-size particles (up to 19%, but usually below 2%), sometimes assuming a bimodal or polymodal grain-size distribution. Facies FD occasionally has organic-rich intraclasts with a dimension of a few centimetres.
Grain-size distribution of facies FA to FD (in particular facies FA) is very similar to present-day coastal dune sediments, indicating that these facies are primarily related to aeolian deposition. The podzolized sediments of facies FB are explained by the development of pedogenetic processes during periods of relatively low aeolian activity or in sectors protected from wind action. The significant proportion of fine-grained particles and presence of vegetal remains in facies FC are attributed to deposition in interdune areas relatively close to the water-table. The pollen element indicates the presence of humid dune slacks. The coarser-grained character of facies FD suggests that it may be associated with earlier phases of dune accumulation under extreme wind conditions or when the vegetation cover was scarce. The intraclasts in FD must be related to wind deflation.
Facies FE
Facies FE is distinguished by its polymodal character, with modes of very fine to medium sand and silt-size, and the significant proportion of silt-clay grain-size fraction (up to 28%). Internally its grain-size distribution is highly variable. They define beds of diverse thickness (a few centimetres to 1 m). These sediments are always very rich in organic material, including algal remains and pollen of aquatic plants (see below).
The polymodal grain-size distribution, with relatively high proportion of silt-clay particles in mixture with a sand fraction together with the high organic content and the presence of pollen of hygrophilous taxa and algae, indicate palustrine or shallow lacustrine environments in the dune field.
Facies FF
Muddy sand sediments with bivalve and gastropod shells that are common in brackish environments (Loripes lucinalis, Cerastoderma edule, Peringia ulvae and Bittium reticulatum are the most frequent) define facies FF. The grain-size distribution of these sediments is quite heterogeneous, with varying silt-clay fraction (1–83%). The fraction coarser than 2 mm is composed mainly of fragments of shells. The grain-size distribution is polymodal, including modes of fine to medium sand, medium to coarse silt and fine to very fine silt. The sand population is very similar to facies FA. Facies FF constitute sediment beds of thicknesses of up to 2 m.
The mollusc association point to a brackish lagoon environment with sandy mud substrate and algal mats. This association was adapted to low-energy, rather restricted, ecological conditions, but always with significant influx of sea water. This fact seems to be confirmed by the absence of freshwater or low salinity mollusc species. The presence of a sand population similar to the characteristic of the aeolian facies suggests that wind-blown sand was an important contribution to the infilling of the lagoon.
Facies FG
Facies FG is always found below facies FF. It consists of medium to coarse sands of light colours with abundant shell fragments and limited silt-clay fraction (<1%). The total thickness of these sediments could not be measured because the boreholes never reached the lower sediments, but in places it was found to be c. 30 m. The shell fragments are frequently disarticulated and abraded. They include the shallow marine species Glycymeris glycymeris, Spisula solida, Donax vittatus and Nassarius reticulatus.
The textural features and the mollusc association indicate that these sediments were deposited in coastal, beach to shoreface environment.
Facies distribution
Three sectors, with different facies and facies sequences, may be recognized along an east–west transect (Castilho et al., 2007b). Two transects, representative for the region, are presented in Figure 3. The western sector shows a marine influenced succession. Here, the Holocene sequence comprises, from base to top, beach sediments (facies FG), brackish lagoon sediments (facies FF) and aeolian sands (facies FA). Its eastern limit (0.8–1 km from shoreline) coincides with the eastern limit of the lagoon and beach facies. In the transitional sector (between c. 1 and 3 km from shoreline) aeolian dune sands (facies FA) dominate the sediment record. The podzolized aeolian sands and interdune facies are poorly represented in this part of the investigated succession. In some boreholes facies FA is found until a depth of around 12 m. In the eastern sector the aeolian dune sands (facies FA–FD) are intervened with wetland and freshwater lake deposits (facies FE) and it is possible to identify one or more levels with podzol (facies FB) and interdune sediments (facies FC) at depths greater than 2 m. A recurring association of facies FB, FC and FE occurs between upper and lower zones where facies FA is dominant (Figure 3). In this sector the aeolian succession frequently starts out with the coarser-grained facies FD.
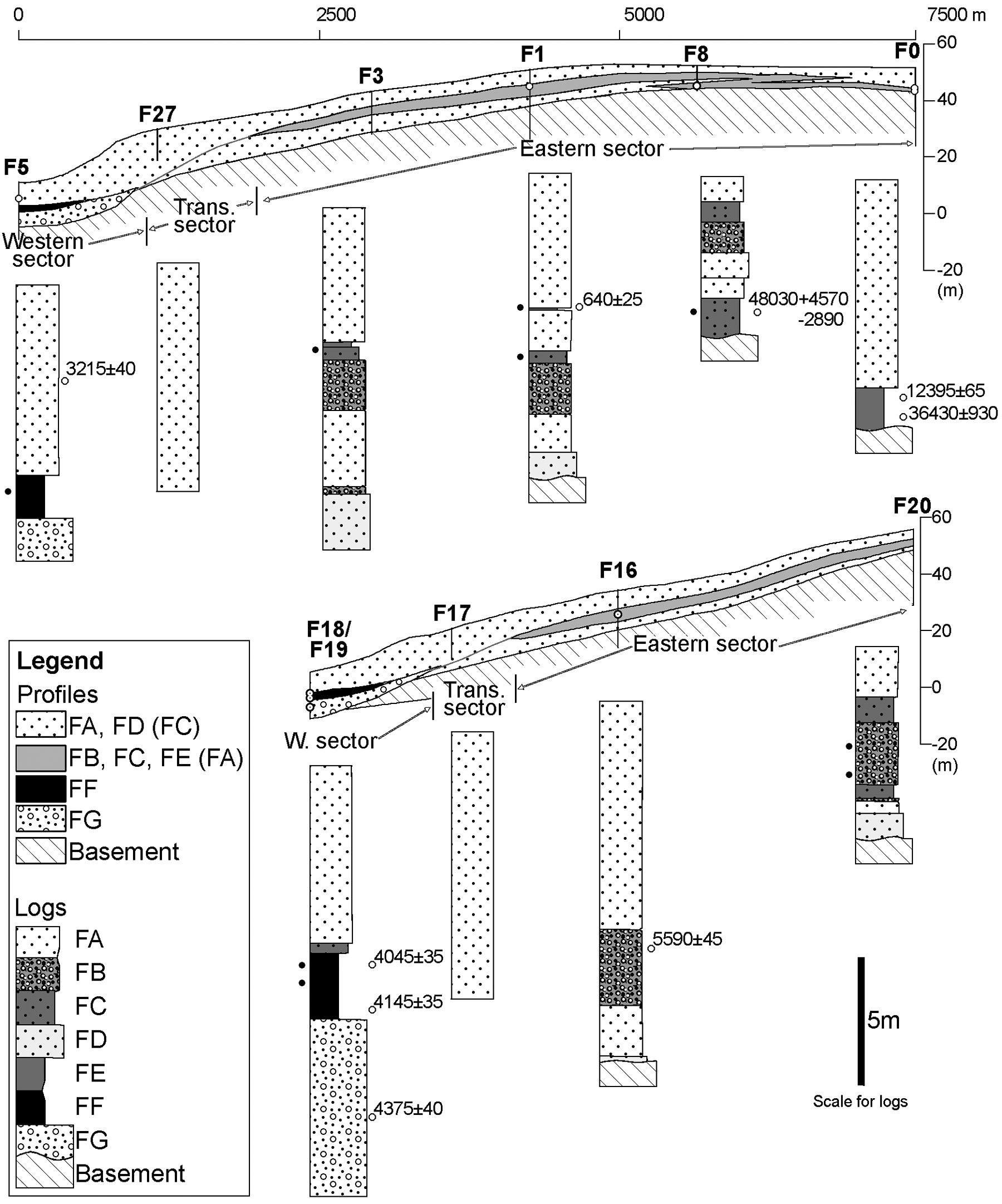
Facies distribution in the Quiaios–Tocha dune field. Facies between brackets in profile legends correspond to less common sediments. Black dots at the left of logs indicate palynological samples
Chronology
The bases of the lake cores were dated as well as the levels of lake formation (Table 1). Datings from the borehole samples were chosen with the aim to determine the time period of lagoon existence (F18, F5), the time of a maximum inland position of the coastline (F19), the phase of lake existence west of the present lakes (F1), the period of lake presence east of existing lakes with subsequent sand invasion (F0) and the moment of formation of a deposit with a uniquely different pollen assemblage (F8). Dates quoted in this article as yr BP are conventional (non-calibrated) 14C years.
Palynology
Only the western and eastern sectors had pollen containing organic deposits (Table 2). All depths of boreholes are given in metres below surface.
Pollen/NPP content
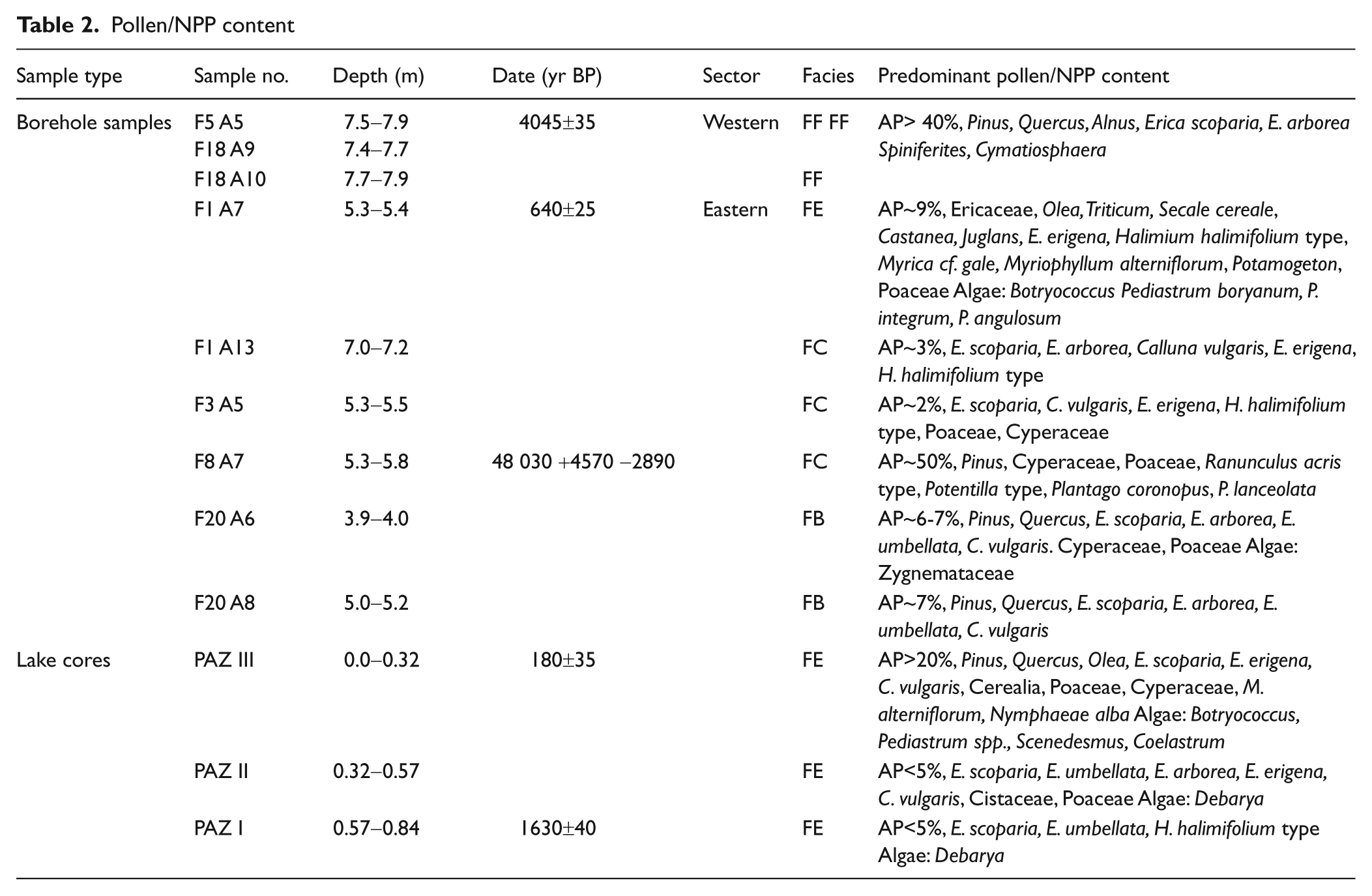
Borehole samples
Pollen and NPP content of the borehole samples are presented in Figure 4.
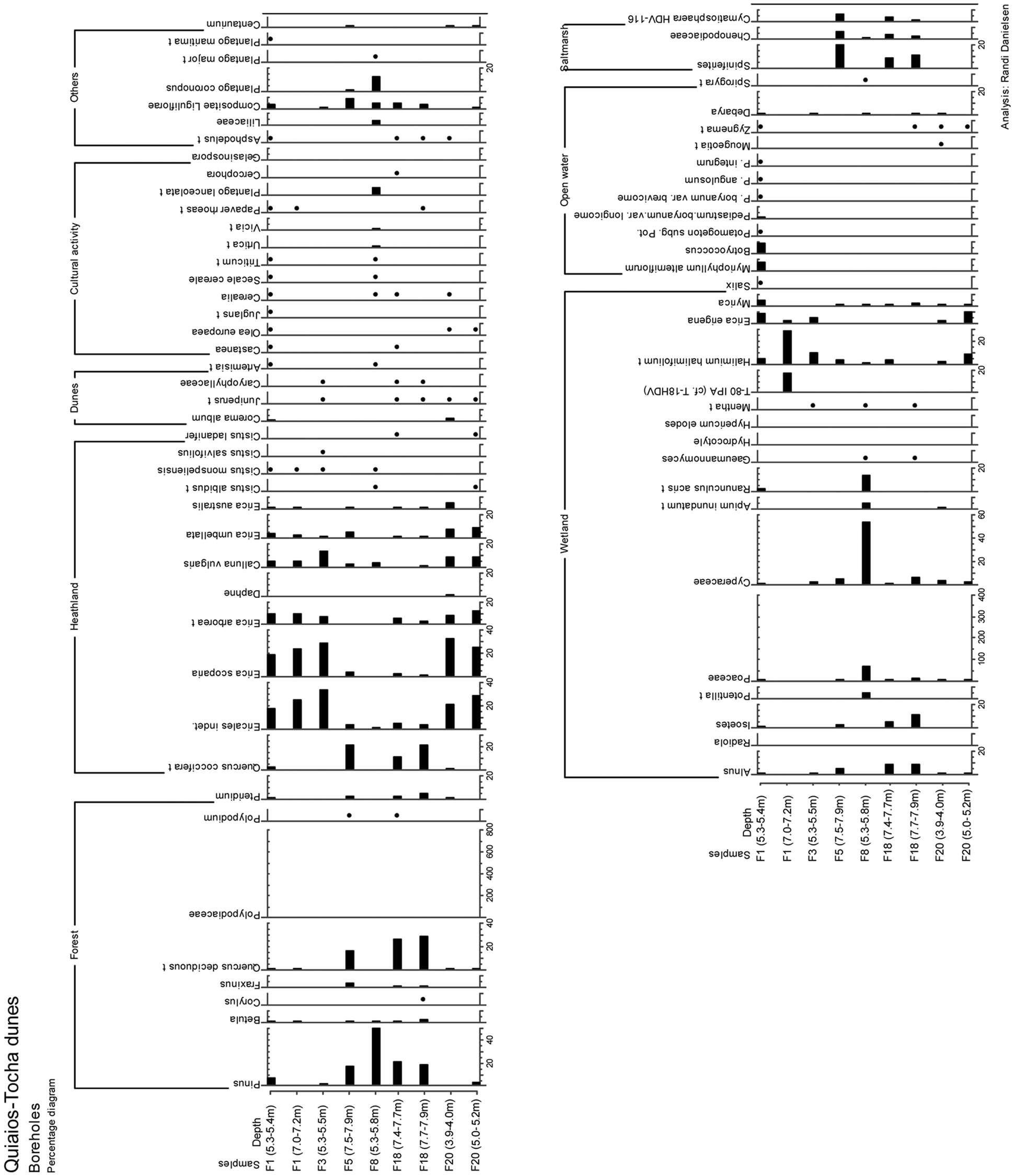
Pollen diagram Boreholes
Western sector – F5 and F18
The boreholes from the western sector penetrated a lagoonal deposit rich in mollusc fragments and plant remains including pollen. The deposit is situated between 1 m above and 2.6 m below present Mean Sea Level (MSL).
Pollen content of both F5 and F18 (facies FF) register salt marsh vegetation. The dominance of pollen from pine and oak indicate the proximity of a mixed forest of these taxa. Pollen of Chenopodiaceae suggest a salt marsh environment as do the presence of the dinoflagellate Spiniferites. Also the green algal genus Cymatiosphaera is considered an indicator of marine influence (Bakker and van Smeerdijk, 1982; Mateus, 1992). A humid Alnus carr with grass and sedges seem to have fringed the lagoon inland. The brackish lagoon was dated in F18 (4145±35 yr BP and 4045±35 yr BP) (Figure 3).
The younger part of the lagoonal deposit shows continuing presence of mixed oak/pine forest. The occurrence of large pollen of Poaceae (possibly Cerealia) may indicate agriculture and human presence in the near vicinity of the lagoon in this more recent deposit although we cannot discard the inflection of Cerealia type produced by salt marsh/dune Poaceae (Mateus, 1992).
Eastern sector – F1, F3, F8, F20
Two samples were collected from F1, a lower deposit found in podzolized sand (facies FC) and an upper organic deposit (facies FE). Heathland dominated by Ericales characterized the vegetation in the region during the formation of both deposits. Arboreal pollen have very low percentages so eventual trees must have been scattered or distant. In the sample from the lower unit, the local element of the pollen was deposited in a wetland area or at the edge of a lake, possibly seasonally inundated.
The upper muddy layer, however, was deposited in a lake environment and lake conditions persisted throughout the year. The abundance of various species of algae and aquatic plants confirm this freshwater lake environment. Lake conditions at F1 have been dated to 640±25 yr BP. Anthropogenic activities in the near vicinity are demonstrated by the presence of pollen of agricultural plants such as Triticum and Secale, as well as Castanea, Olea and Juglans.
The sample at F3 (facies FC) has a pollen content that indicates a regional vegetation of heathland type. Pollen of hygrophilous plants such as E. erigena, Halimium halimifolium type (including H. lasianthum) and Cyperaceae suggest that the local environment contained humid dune slacks.
The mud nodule from borehole F8 (facies FC) contains a pollen assemblage totally unlike the assemblages observed in the other samples. Complete dominance of pollen of Pinus demonstrates a regional pine forest. The local environment is represented by marsh/sedge vegetation with high pollen values of Cyperaceae, Poaceae, Ranunculus, Potentilla and of various species of Plantago. The sample was dated
Both samples from F20 (facies FB) suggest a regional vegetation of open heathland with varying degree of local soil humidity. The lower sample was probably a humid heathland while the upper sample suggests higher soil water content in a grass/sedge marsh. Microfossil content of both samples suggests a location at the margin of a lake or in a dune slack.
Lake cores
The pollen assemblages found in the lake cores (facies FE) represented by the diagram from Lake Vela (Figure 5), could be divided into three pollen assemblage zones (PAZ) recording the regional vegetation through time (solid lines). Local vegetation changes in hygrophilous taxa and algae were divided into local zones (interrupted lines). A more explicit analysis of the lake cores are presented in Danielsen (2008, 2010). A summarized version of the vegetation development, as deduced from the lake samples, is offered here:
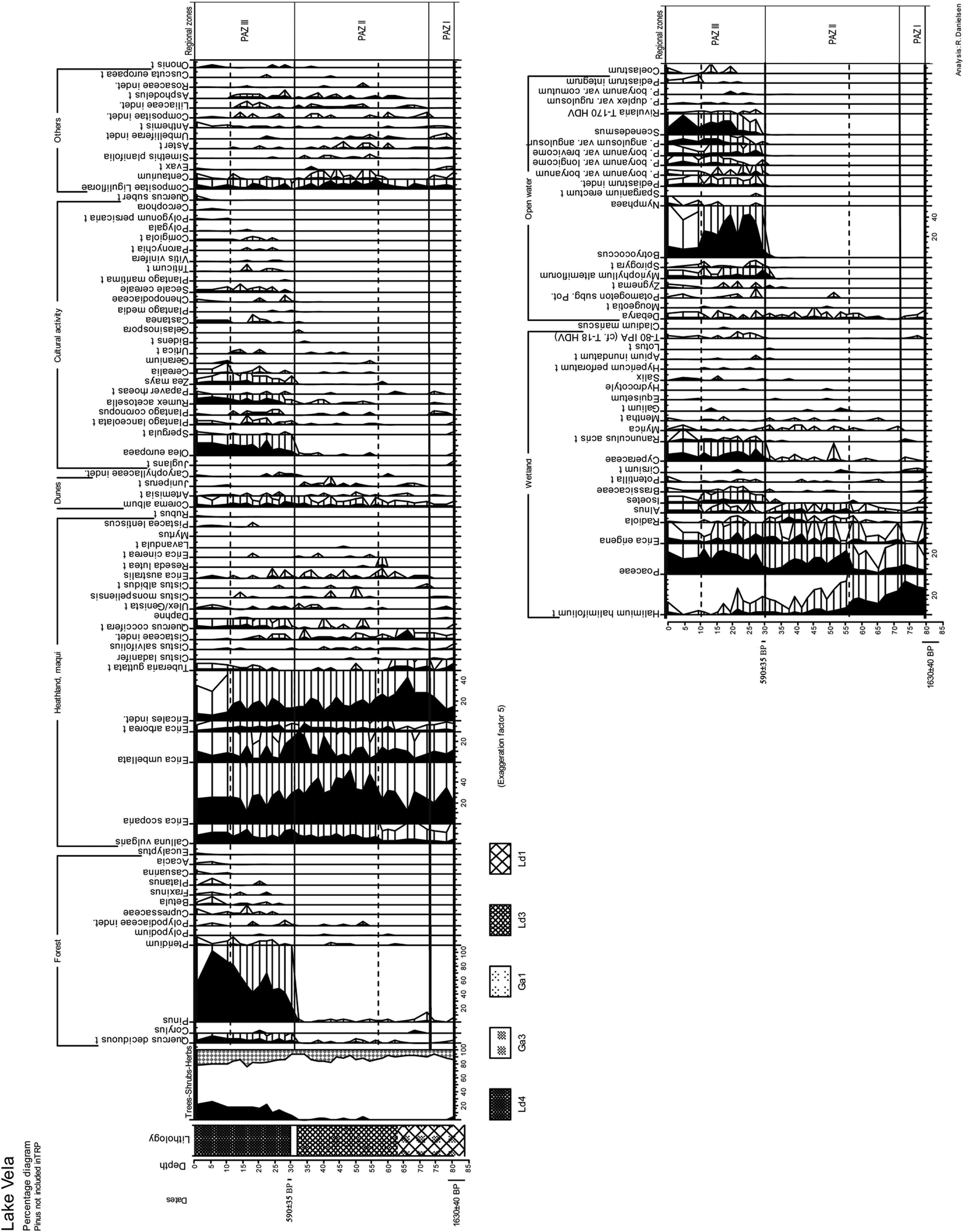
Pollen diagram Lake Vela
PAZ I
Sediment accumulation at the lake site started 1630±40 yr BP. This initial zone is characterized by low values of arboreal pollen (< 5%) and high values of heathland and wetland plants. The upland vegetation registered in this zone is interpreted as treeless open heathland vegetation with E. scoparia and E. umbellata. Wetland vegetation with H. lasianthum and E. erigena dominated locally representing a terrestrial hydroseral stage.
PAZ II
In this zone a continuous low representation of forest vegetation prevailed concomitant with the dominance of pollen of heathland species such as E. scoparia. Pollen of Olea, Cerealia and Zea mays appear in this zone suggesting agricultural activity. Indicators of grazing also increased from this zone onwards. The local dominance of H. lasianthum was gradually substituted by E. erigena and Poaceae possibly due to higher humidity or water-level of the soil.
PAZ III
Reforestation of the region is recognized in this zone. Pollen of Pinus was most important but pollen of Quercus also increased. Lakes developed at the onset of this phase as seen by the large increase in quantity and variety of algal remains. Indicators of agriculture and grazing attained high values. The transition between PAZ II and PAZ III has been dated in both lakes. In Lake Vela the mud fraction at the transition was dated to 590±35 yr BP whereas in Lake Braças stumps of E. erigena, from the moment of inundation and death of this species, gave the more recent date of 180±35 yr BP. The regional pollen curves indicate that lakes formed at the same time at the two sites, apparently at the moment when arboreal pollen curves increase abruptly. The dating from Lake Vela is therefore presumed to be too old as it may contain abraded organic sediment deposited together with aeolian sand at the period before lake formation (see discussion below). The dating from Lake Braças is young and inconclusive but on the basis of old maps in combination with the dating, it is concluded that the time of formation was in the time period 1723–1866 (Danielsen, 2008).
The local reforestation (1924–1940) is identified in the upper part of the zone by the emergence of pollen of exotic taxa (Eucalyptus, Acacia, Platanus and Casuarina).
Discussion
Late-Pleistocene environment
The oldest organic deposit retrieved in this investigation was dated 48 000 yr BP but as it attains the limit of the range of 14C dating, may be older. This sample originated from F8 west of Lake Braças at c. 5.5 m depth. The pine forest represented in the pollen assemblage may reflect the reported Würm (Marine Isotope Stages 3–2) forests of Pinus gr. sylvestris-nigra species group (including Pinus sylvestris, P. nigra and P. uncinata) (Figueiral, 1995; Figueiral and Carcaillet 2005, García-Amorena et al., 2007). Studies from North Portugal, have registered several finds of trunks of pine along the coast (García-Amorena et al., 2007; Granja, 1999; Granja and Carvalho, 1995). At the S. Pedro de Maceda and Esmoriz beaches c. 70 km towards north, numerous in situ pine trunks were dated between 33 960±800 yr BP and 19 910±260 yr BP (García-Amorena et al., 2007).
The truncated dunes of the Gândara plain, east of Lake Vela and Lake Braças, are thought to be of Pleistocene age, possibly related to the last glaciation, since they are associated with wind-worn pebbles and Palaeolithic artefacts, as well as periglacial deposits (Carvalho, 1964). More recently the dunes are placed in a wider time frame ranging from the Würm to the Holocene (Granja and Carvalho, 1992, Granja et al., 2008). This investigation showed that between 36 420±930 yr BP and 12 395±65 yr BP a lake existed in the easterly location of F0. The top of the lake sediment in F0 is covered by these truncated dunes indicating an origin of the dunes synchronous with the climatic degradation during the Younger Dryas. According to the sea level curve for Portugal (Dias et al., 1997, 2000) sea level fell c. 20 m between 12 000 and 10 000 yr BP, a scenario of exposure of large amounts of beach sand to aeolian activity. Similar dune fields exist in southwestern France, the Landes dunes, in the Aquitaine region. The French dunes have been compared with the Portuguese dunes in various publications (Carvalho, 1954; Paskoff, 2001) and analogies between them in what concern a Pleistocene origin, have been described.
Holocene environment
Forests and coastal lagoons
The oldest Holocene samples were obtained from boreholes F16 and F19. In borehole F16, at 9.5 m depth, the layer was characterized by large quantities of wooden remains indicating a local forest and gave an age of 5590±45 yr BP. In F19 at 14.5 m many fragments of Fraxinus were found in beach to shoreface sediments and one fragment was dated to 4375±40 yr BP. Fraxinus is common in freshwater zones and not in beach environments, but the fragments probably originated from overwashed habitats near the coast or were transported from riverbanks to the sea.
At the end of the Flandrian transgression 5000–4000 yr BP the coastline reached its maximum Holocene inland position (Figure 6). Large amounts of beach to shoreface sediments (facies FG), assigned to this period, were identified c. 1 km east of the present coastline. Beach or marine deposits were not found further towards the east and this disproves the hypothesis of several investigators of a coastline reaching the area of the present row of freshwater lakes 6–7 km inland around 3000 yr BP (Abecassis, 1961; Dias et al., 1997; Figueiredo, 1987; Noivo, 1996).
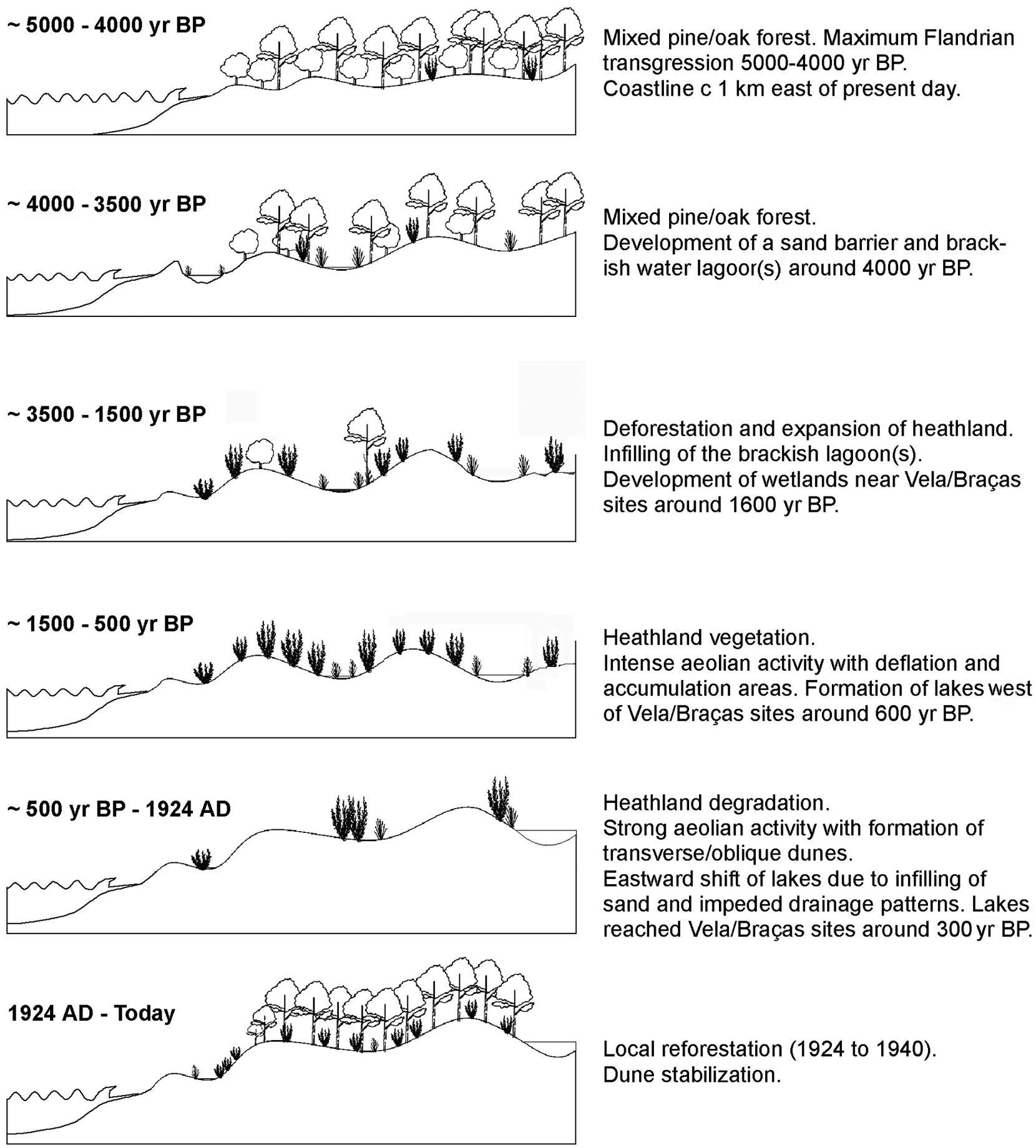
Mid- to late-Holocene evolution phases of the Quiaios–Tocha littoral dune fields
Lagoonal deposits were encountered along the coastal fringe at elevations between 1 m above and 2.6 m below present MSL. 14C datings of shells from these deposits in F18 gave respectively 4145±35 yr BP and 4045±35 yr BP. This corresponds well with the sea level curve for Portugal (Dias et al., 1997, 2000) which shows that between 5000 and 3000 yr BP sea level reached more or less its current position. The lagoon(s) is thought to have formed as a consequence of decelerating rate of sea level rise, causing formation of a sand barrier off the Quiaios–Tocha coast. Dating the base and top of the lagoonal deposit in F18 (Figure 3), indicate only 100 years existence of the lagoon. This is probably not correct and most likely caused by sediment mixing at the time of closure of the lagoon. Planned future investigation including survey with Ground Penetrating Radar (GPR) may solve questions concerning dimension, number and lifetime of lagoon(s). Pollen and macrofossil remains from the intercepted lagoonal deposits at F5 and F18 depict vegetation dominated by a mixed forest of Pinus and Quercus with Alnus near coastal marshes and Fraxinus in freshwater areas. All along the coast of Portugal similar lagoonal deposits have been identified and they are invariably associated with lagoon-barrier island systems (Alday et al., 2006; Bernardes and Rocha, 2007; Bernardes et al., 2001; Callapez et al., 2005; Cearreta et al., 2002; Dinis et al., 2006; Freitas et al., 2002; Granja, 1999; Granja and de Groot, 1996; Granja et al., 2010; Henriques and Neto, 2002; Mateus, 1992). A comparable system is also reported from the Galician coast at this time (Bao et al., 2007).
Deforestation, heathland expansion and parabolic dune formation
Deforestation of the region followed and heathland vegetation expanded. It is probable that forests disappeared as a result of a combination of negative impact from climate and human activity surpassing threshold values for the existence of such forests. An element pointing towards anthropogenic involvement in the deforestation process is that it is not contemporaneous in the different regions of Portugal. The amount of settlements from the Chalcolithic, the Iron Age and Roman times registered in the hills bordering the Quiaios–Tocha region (Mesquita de Figueiredo, 1943; Guerra and Ferreira, 1971, 1974; Santos Rocha, 1888–1900, 1908) as well as the presence of indicators of grazing and agriculture in the deposits formed during the heathland phase, suggest human activity as a plausible main cause of the deforestation process. The exact time of deforestation is not yet established. We are merely able to say that it happened sometime between 4000 and 1600 yr BP. The latter date is based on the fact that at the time of the initial sediment accumulation at the lake sites (PAZ I) there are no indications of forests in the diagrams (Danielsen, 2008). In F5 a layer very rich in wooden fragments was dated to 3215±40 yr BP. This may have resulted from a phase of forest destruction, but more investigation is needed to confirm this.
During the heathland phase, a vegetation mosaic of dry heathland and wetland zones in dune slacks, characterized the region. Both lake cores (PAZ I and II) and also boreholes F3, F1 and F20 record this phase. The heathland was dominated by the tall heather Erica scoparia together with other Ericales such as E. umbellata, E. australis and Calluna vulgaris and with species of the Cistaceae. Dune slacks and wetland zones were characterized by Halimium lasianthum and E. erigena and by algal remains of the Zygnemataceae (Debarya, Zygnema type and Mougeotia type) indicative of seasonally inundated pools (van Geel et al., 1989).
Heathland is a vegetation type that during millennia existed along the entire European Atlantic coast (Haaland, 2002) preserved by anthropogenic action (grazing, cutting and burning) and hence considered semi-natural. Holocene deforestation and heathland development has also been reported from palynological investigations of other parts of Portugal; northwest Alentejo (Mateus, 1989, 1992; Queiroz, 1989, 1999; Queiroz and Mateus, 1994), the Tagus valley, about 75 km NE of Lisbon (van Leeuwaarden and Janssen, 1985), Nazaré (Gomes, 2007), Furadouro- Esposende region (Gómez-Orellana et al., 2001), Serra da Estrela (van der Knaap and van Leeuwen, 1995) and Viseu (López-Sáez et al., 2001).
After deforestation and opening up of the landscape, the environment became more vulnerable to climate change and susceptible to aeolian activity. The parabolic dunes are remnants of a period of sand migration when the vegetation cover was sufficiently uninterrupted to anchor ‘arms’ of the dunes (Durán and Herrmann, 2006; Tsoar and Blumberg, 2002) and yet discontinued enough to allow aeolian activity to occur. They are preserved in areas where vegetation cover protected them against subsequent sand invasions such as in the triangular forest called ‘Pinhal do Povo’ planted prior to 1773 (Serra et al., 1773) to protect the village of Quiaios against invading sand. The great majority of the parabolic dunes point towards southeast and east indicating the existence of dominant northwesterly winds during the time of formation. Parabolic dunes at some stage may have formed throughout the whole extension of the dune field but turned into transverse dunes, in places without vegetation cover, during the following aeolian phases.
Climate deterioration, formation of transverse/oblique dunes and reforestation
Around 300 years ago the region’s heathland was strongly influenced by pastures and by the climatic degradation during the ‘Little Ice Age’ (LIA) (Clarke and Rendell, 2006; Danielsen, 2008). Aeolian activity and sand drift was facilitated by discontinuous vegetation cover and possibly enhanced by harsh climatic conditions. Dune migration shifted large quantities of sand across the coastal plain and apparently led to relocation of wetland areas and lakes. Holocene dune slack and wetland deposits are found in facies FB, FC and FE in boreholes F1, F3, F20 and zones PAZ I and II of the lake cores and the characteristic vegetation is described above. Lake deposits are registered in the upper sample of the organic deposit in F1 and in PAZ III of the lake cores. The lake environment was characterized by algal remains of the genus Pediastrum and of aquatic plants like Myriophyllum alterniflorum, Nymphaea and Potamogeton. The lake phase in F1 was dated 640±25 yr BP. Formation of Lake Vela and Lake Braças as permanent open water lakes, occurred between 1723 and 1866 (Danielsen, 2008). The existence of a lake in F1 during the heathland phase, later covered by aeolian sand and the subsequent origin of lakes in the Vela and Braças sites c. 1 km further east around 300 years ago (Danielsen, 2008), suggest an eastward dislocation of the lacustrine systems (Figure 6). Similar drainage impediment by sand drift and resulting lake development has been reported from a palaeolake in the Netherlands (Bos et al., 2006).
The transverse/oblique dunes (Figure 1) formed during LIA (Clarke and Rendell, 2006; Danielsen, 2008; Noivo, 1996) a period of increased storminess and sand mobilisation throughout Europe as summarized in Clarke and Rendell (2009). The dunes are mainly oblique as their crests are uniformly sloping on both sides indicating that they were formed by winds of two opposite quadrants. In fact, the present-day wind roses (Figure 1C) show that in spite of the general dominance of winds from north and northwest, the southwest winds are stronger in the humid months. The transverse dunes, with a steeper inclination on the leeward side of the dune crest, are exclusively found in a limited area in the southeastern part of the investigated region. These dunes solely formed by the action of winds from north to northwest showing that they were protected by the winds from the south because of the existence of a forest (‘Pinhal do Povo’?).
Lack of vegetation or other obstruction of dune movement in the Quiaios–Tocha region, led to an advance of dunes inland of many kilometres length. Sand ridges of 15–20 m accumulated at the edge of patches of forests. The magnitude of sand displacement during LIA may, however, better be illustrated by comparison with a similar area c. 50 km south of the Quiaios–Tocha region where a large pine forest (Pinhal do Rei), planted on the order of king Dom Dinis (1261–1325), existed. Dunes accumulated at the western edge of the forest and the outline of the dune ridge follows the ancient forest edge and reaches elevations of 70 m (André, 1996). At the site, an in situ fossil pine trunk at the base of a 40 m dune ridge was dated 370±40 yr BP, demonstrating the quantity of sand deposited there in a relatively short period of time (André, 1996).
Reforestation measures were implemented in order to protect agricultural fields from migrating dunes. Initial reforestation is detected in the pollen diagrams of the lake cores (PAZ III) around 300 years ago by a large and rapid increase in arboreal pollen values, particularly in the ones of Pinus. Local planting in the Quiaios dunes themselves was carried out between 1924 and 1940 (Rei, 1940) and is recorded in the diagrams as a large increase in Pinus pollen values and appearance of pollen of exotic species planted in the region such as Acacia, Eucalyptus, Platanus and Casuarina. The success of reforestation is confirmed by the unbroken coastal forest subsisting in the region to this day.
Conclusions
The investigation gives insight into the landscape development of the Quiaios–Tocha dunes during the mid to late Holocene in particular (Figure 6), but with some isolated signals from earlier periods. During the Würm glaciation the region probably had a Pinus sylvestris forest. Around 12 000 yr BP aeolian activity created sand dunes reaching far inland. This is an indication of harsh climate conditions and scattered vegetation at the time. Later a mixed oak/pine forest seems to have predominated in the region as was the situation when sea level reached its current position 5000–4000 yr BP and the coastline reached its maximum inland position about 1 km east of the present one. Around 4000 yr BP barrier protected coastal lagoons formed, and were later infilled with sand. During the forest phase the existing dunes were vegetated and some podzol horizons formed. The forest persisted for some time but was eventually replaced by a semi-natural heathland before 1600 yr BP. The water-table seems to have been high and wetland environments developed locally. Cultural activity including forest clearance, pastures and agriculture may have caused the deforestation and contributed to the maintenance of the open heathland.
After deforestation the landscape was especially vulnerable to climate change and cultural activity and two periods of major sand mobilization can be detected. The parabolic dunes formed during the first period when the region still had a relatively uninterrupted vegetation cover. During a following period of sand mobilization, oblique/transverse dunes formed in unvegetated and barren parts of the coastal plain. Deteriorating climatic conditions during LIA may have intensified sand drift from the west. The most recent phase resulted in the counteractive reforestation of the last centuries. Dunes invaded existing lakes and wetlands, disrupting the drainage patterns in the coastal plain and eventually relocated the lakes eastwards to their present location around 300 years ago.
Footnotes
Acknowledgements
The authors would like to thank the Earth Science Department and the Centre of Geosciences, Coimbra University; the Laboratory of Archaeosciences of IGESPAR (Instituto de Gestão do Património Arquitectónico e Arquelógico), Lisbon; Águas da Figueira, S. A., DGF, CCDRC and the local authorities of Bom-Sucesso and Tocha. Special thanks to Dr José Mateus and Dr Paula Queiroz (Terra Scenica, Lisboa) for their invaluable advice on palynology of Portugal. The manuscript was improved thanks to valuable comments by anonymous reviewers.
This research work was co-funded by the Portuguese Foundation of Science and Technology (FCT) of the Portuguese MCTES and POCI 2010, through Project EcOwet (POCI/CTE-GEX/58951/2004), and by POCI 2010 and FSE through the scholarship SFRH/BD/11100/2002.