
Abstract
Pollen, non-pollen palynomorph, macrofossil, and lithological analyses of mid-Holocene deposits from a small peat bog in northern Poland provide new proxy data on vegetation dynamics and climate between c. 9500 and 4600 cal. BP. The most prominent wetter climate phases occurred between c. 9500 and 9280±160 and between c. 5000 and 4600 cal. BP while a hot and dry period between 8185±145 and 5299±195 cal. BP corresponds with the Holocene Thermal Maximum (HTM). The HTM phase has been characterized by absolute dominance of Tilia cordata and the presence of T. platyphyllos beyond its current geographical range limit and a c. 2700 yr long, high incidence of the parasite fungus Kretzschmaria deusta in the local forest stand, concurrent with a dry phase on the mire. For the HTM, we suggest mean summer temperatures at least 3°C higher than today’s values. These data are in accordance with the geographic pattern of the HTM temperature anomalies in northern Europe, including the Baltic region, which suggest a gradient of positive anomalies from north to south and from west to east. The data also illustrate the effect of climate warming on the mid-Holocene pathogen outbreak and may indicate intensive vegetative reproduction as a Tilia strategy for population survival. Summer drought was probably important for high success of K. deusta in infecting weakened Tilia trees.
Keywords
Introduction
The mid-Holocene climate has been one of the most thoroughly debated topics since publication of the first results in Quaternary palaeoecology. Although in recent years, the number of interdisciplinary projects and papers in this subject area has rapidly increased (Barber et al., 2004; Wanner et al., 2008), knowledge in this field still has gaps. The patterns identified using large-scale palaeoclimate reconstructions, including modelling based on pollen data (e.g. Cheddadi et al., 1997; Davis et al., 2003) are rather coarse in respect to both temporal and spatial scales. The existing data on the mid-Holocene climate variability within Europe suggest important regional differentiation both in temperatures and precipitation as well as different amplitudes and some opposite trends in climate changes between different regions (Brewer et al., 2007; Davis et al., 2003; Magny et al., 2003). Some climate reconstructions obtained by different methods and using different proxies are not coherent, indicating the need for a better understanding of physical and biological processes (Huntley, 2012; Seppä et al., 2009) and for new multiproxy records from particular regions and time-slices that could be used for data–model comparisons to validate palaeoclimatic simulations.
The Kashubian Lake District (Figure 1), bordering the southern coastal zone of the Baltic Sea, is of special interest regarding climate history. It lies in the transitional climatic zone where the dominating westerly airflow from the Atlantic meets continental air masses. The thermic regime in this area is distinctly dependent on the impact of the North Atlantic Oscillation and the strength of its blocking by the Russian and Scandinavian anticyclonic centre (Filipiak, 2004); therefore, any palaeoclimatic data from this region would be important for reconstructing past regional atmospheric circulation. Unfortunately, this area has been neglected for a long time as concerns modern, high-resolution palaeoecological studies. Only during the last few years have new projects begun in this region, some of which are still in progress. The sole mid-Holocene palaeoclimatic reconstruction is based on a multiproxy record from a raised bog and concerns the later phase of this period (Lamentowicz et al., 2010).
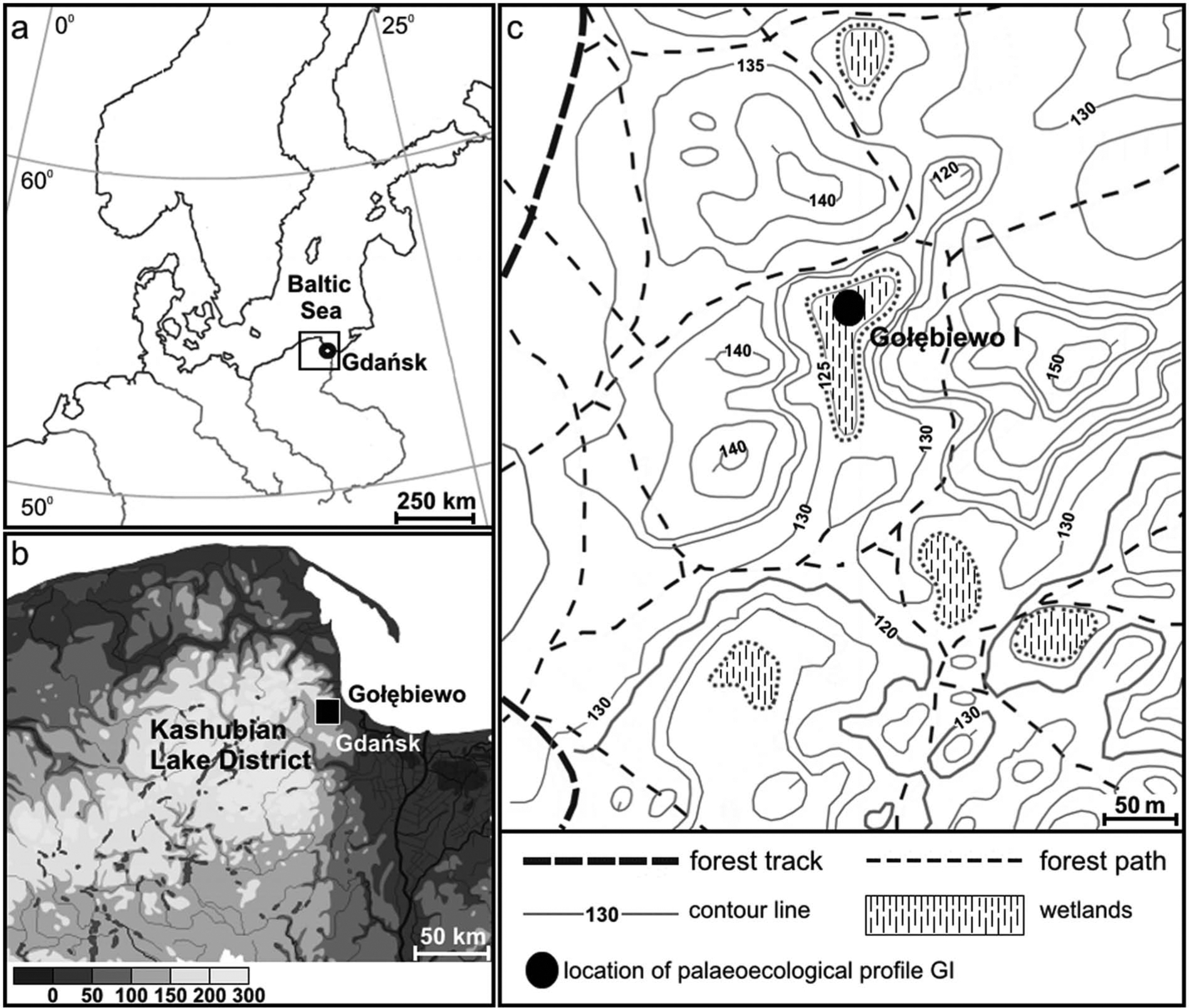
(a) (b) Geographic location of the study area. (c) Core location.
The use of indicator characters of plant taxa, as well as of other organisms, is a fundamental method in palaeoecology, enabling reconstruction of past environments, including climate parameters (Birks, 2003; Birks et al., 2010). Recently, the spectrum of organisms used as palaeo-bioindicators has grown because of developments in the analysis of non-pollen palynomorphs (NPPs), among them algal, faunal, and fungal microremains preserved in pollen samples, which are determined and counted along with pollen grains (van Geel, 2001). Fungal remains are of high potential in this context (van Geel and Aptroot, 2006), especially those of specific taxa that are well characterized in terms of their biology and ecological requirements. The use of NPPs essentially improves palaeoecological interpretation and helps with reconstruction of several specific ecological aspects of the palaeoenvironment, including local habitat conditions (van Geel, 1978; Yeloff et al., 2007), pathogen outbreaks (Innes et al., 2006), and frequency of herbivore visits at the site (Cugny et al., 2010; Feeser and O’Connell, 2010).
The aim of our paper is to present new palaeoecological data from N Poland, distinct by extremely high concentrations of Tilia spp. pollen and ascospores of Kretzschmaria (Ustulina) deusta in mid-Holocene deposits, illustrating specific long-term co-occurrence of a fungal parasite and a host-tree population. Results of pollen analysis supplemented with NPPs, analysis of plant macrofossils, and allogenic mineral matter content provide a basis for the reconstruction of climate conditions in the period between c. 9500 and 4600 cal. BP.
Study area
The Kashubian Lake District bordering the southern Baltic coastal zone (N Poland) is a distinctly elevated area reaching 328 m a.s.l., making it the highest point within the European Plain. The area has been shaped by the Vistulian (Weichselian) ice sheet, leaving morainic uplands and patches of outwashed plains, numerous channels, kettle holes, and valleys now filled with lakes and mires. The upland is dissected by depressions and dry gullies. The morainic formations of clay or sandy-clay are the most common substrates for soil cover; however, patches of sandy soils are also frequent. The area is located within the temperate nemoral zone where beech forests are the main potential vegetation type (Melico-Fagetum, Luzulo pilosae-Fagetum, and Fago-Quercetum) (Matuszkiewicz, 2008).
The proximity of the Baltic Sea directly influences the climate of the eastern part of the region, where our site is located. The air humidity is relatively high, with cold, cloudy, moderately severe winters and mild summers. The average annual temperature is 7.2°C. The seasons are clearly differentiated. Spring starts in March and is initially cold and windy, later becoming warm and often very sunny. Summer, which begins in June, is predominantly warm with plenty of sunshine interspersed with heavy rain. July is the hottest month (17.0°C on average). Autumn comes in September and is at first warm and usually sunny, turning cold, damp, and foggy in November. Winter lasts from December to March and includes periods of snow. The coldest month is January, with an average temperature of −1.5°C. Annual precipitation is rather moderate, c. 590 mm, but there is considerable diversity from year to year and with the season, with the highest values in July. Westerly winds dominate (Owczarek et al., 2007).
The site of Gołębiewo I (54°27′N; 18°30′E) is a small peat bog, c. 6 ha in surface, situated at the east-facing edge of the morainic upland. It fills a small, elongated depression about 150 m long and c. 40 m wide on average (Figure 1a, b). This form is surrounded by rather steep slopes covered by woodland; fertile Fagus sylvatica forest occurs in the north and northeast while an acid beech forest with an admixture of Pinus sylvestris and Quercus robur dominates in the remaining part of the catchment.
The present-day vegetation of the mire is strongly disturbed mainly due to a ditch dug out along the bog margin. It represents a transitional bog including elements typical of both minerogenic and ombrogenic mires. Sphagnum palustre, S. russowi, S. squarrosum, and S. fallax are common. Juncus effusus and Dryopteris carthusiana form large patches on wet, disturbed surfaces and along the bog margins while in other places Comarum palustre and Menyanthes trifoliata are among the most distinct species. In the central part of the mire, Vaccinium oxycoccos, Andromeda polifolia, and Ledum palustre are frequent. A large area is covered by Molinia caerulea and scattered low trees of Betula pubescens, Frangula alnus, and Pinus sylvestris.
Materials and methods
The profile was collected in the central part of the peat bog (Figure 1c) using Instorf samplers 10 or 5 cm in diameter. A preliminary description of lithology was made in the field and then supplemented according to examination of the sediments in the laboratory and the results of LOI. Dry weight was obtained after drying at 105°C for 24 h, and LOI was determined at 550°C for 8 h. A simplified Troels-Smith system (Aaby and Berglund, 1986) was applied for sediment descriptions. Our present study concerns only the lower part of the profile (316–492 cm).
Pollen samples 1 cm3 in volume were boiled in 10% KOH and then acetolyzed (Faegri and Iversen, 1989). If necessary, samples were first processed with HF to remove siliceous matter. Lycopodium tablets (University of Lund) were used to calculate pollen/spore concentrations (Stockmarr, 1971). Non-pollen microfossils (NPPs) (van Geel, 2001) were recorded along with the pollen and identified using nomenclature according to the rules described by Cugny et al. (2010). Percentages of dry-land taxa were calculated using the AP+NAP sums (excluding sporomorphs of aquatic and wetland plants, NPPs as well as corroded pollen). The proportions of K. deusta, corroded pollen, charcoal particles and wetland taxa are based on the sums as above plus the number of spores, pollen or particles of the respective taxa. The NPPs other than K. deusta are presented as concentrations of specimens/cm3 of sediment.
For macrofossil analysis, sediments were cut into slices 4–5 cm thick (100–150 cm3 in volume) considering lithological limits. The material was immersed in water with a small admixture of KOH for 24–28 h and then wet-sieved on a column of sieves of 0.2, 0.5, and 2.8 mm meshes. The residue was segregated under 16× magnification. Identifications were based on several specific keys and confirmed using the reference collection in the Laboratory of Palaeoecology and Archaeobotany at the University of Gdańsk. The results are expressed as number of specimens per 100 cm3 of sediment.
The chronology of the palaeoecological events is based on nine radiometric dating (AMS) performed at the Poznań Radiocarbon Laboratory, Poland. All dates were obtained from selected remains of terrestrial plants isolated from particular layers. To obtain calendar years, OxCal, ver. 4.1, based on the calibration curves IntCal 09 and Marine 09 (Reimer et al., 2009) was used.
Principal component analysis (PCA) (CANOCO for Windows 4.5) served for detection of interspecies relationships. The analysis was based on data from all pollen samples; sporadically occurring tree taxa were excluded.
Results
Litho-, chrono-, and biostratigraphy
The sediment is dark brown, strongly decomposed peat differentiated with respect to decomposition grade and with a mineral matter content ranging from about 50% to below 3% (Table 1). Radiometric dating (Table 2) indicates that the considered section of the profile covers the period of >9500–4600 cal. BP. The time/depth model is based on a linear approximation (Figure 2), which is considered the most appropriate choice in the case of expected abrupt changes in sedimentation rate. The model shows generally a very low accumulation rate with extreme slowing down at the levels around 440–430 cm (c. 9000–8000 cal. BP) and 355–340 cm (c. 6100–5300 cal. BP); we suggest potential hiatuses within both sections.
Litho- and biostratigraphic zonation; sediment designations according to the Troels-Smith system (Aaby and Berglund, 1986).
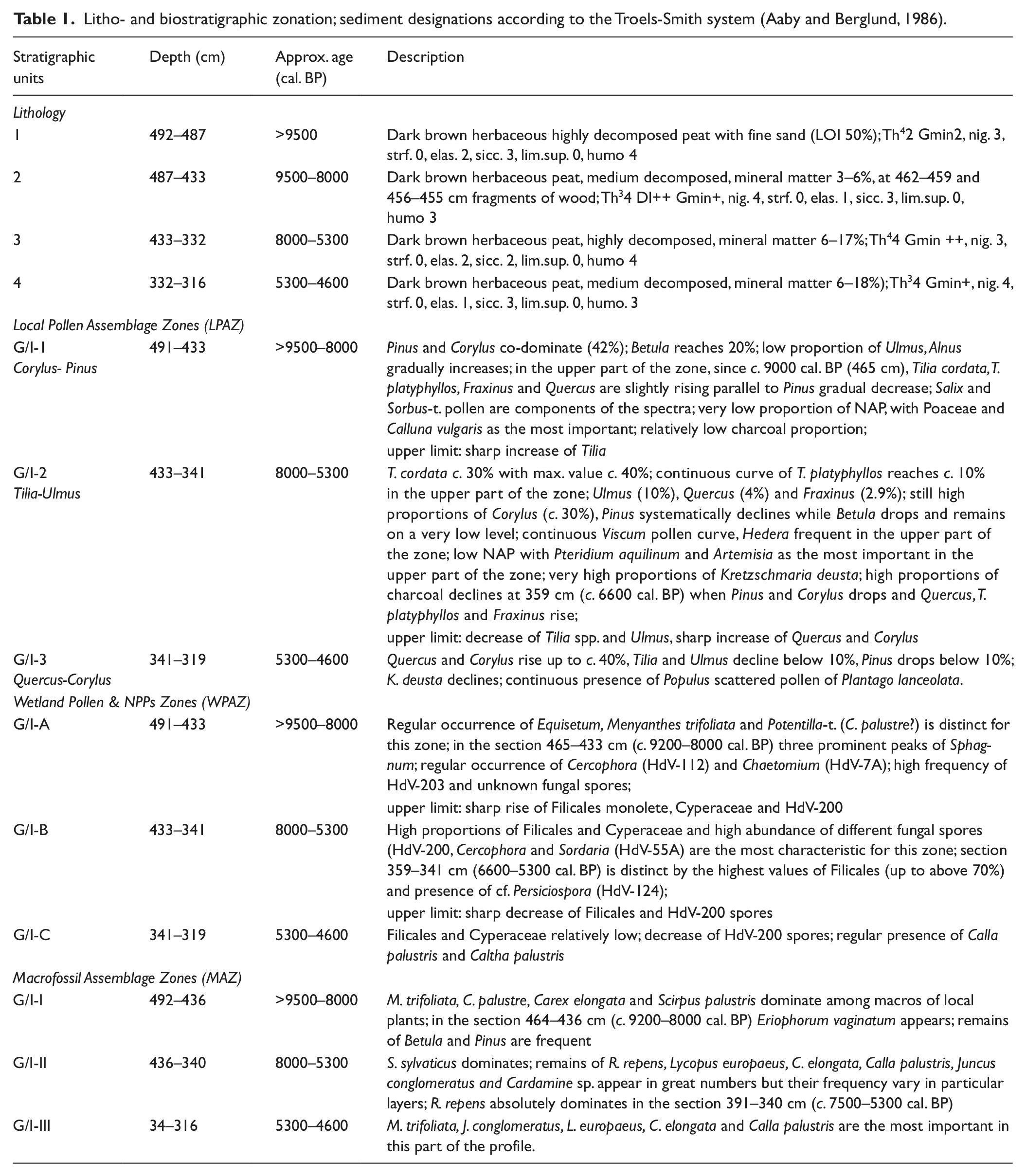
Radiometric dating.
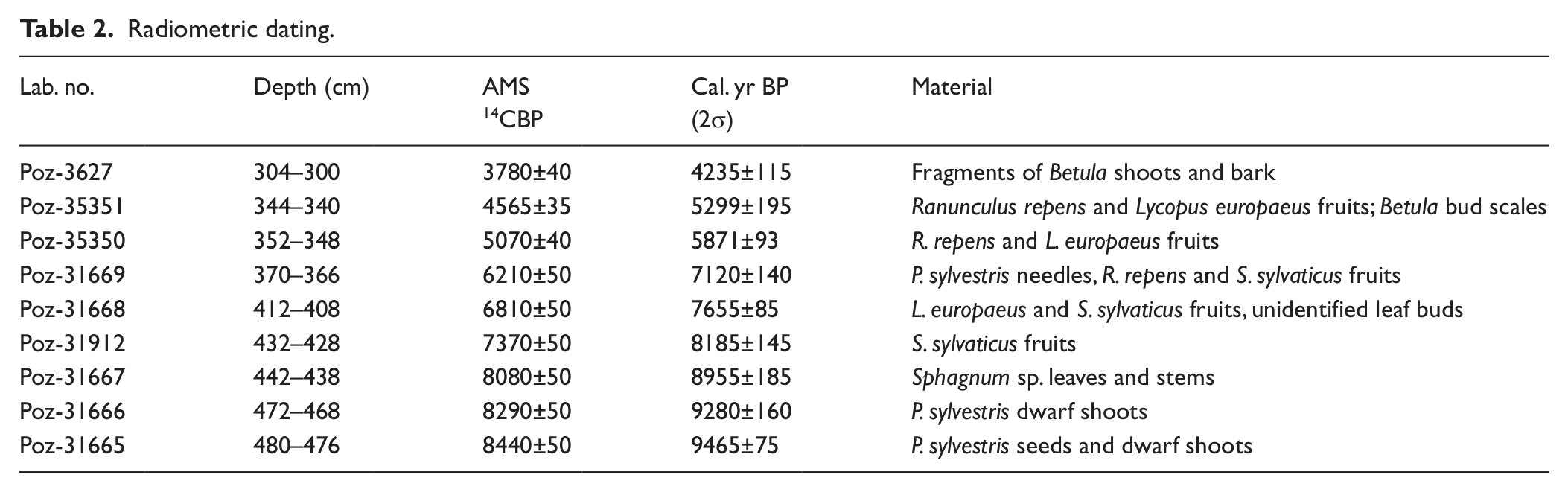
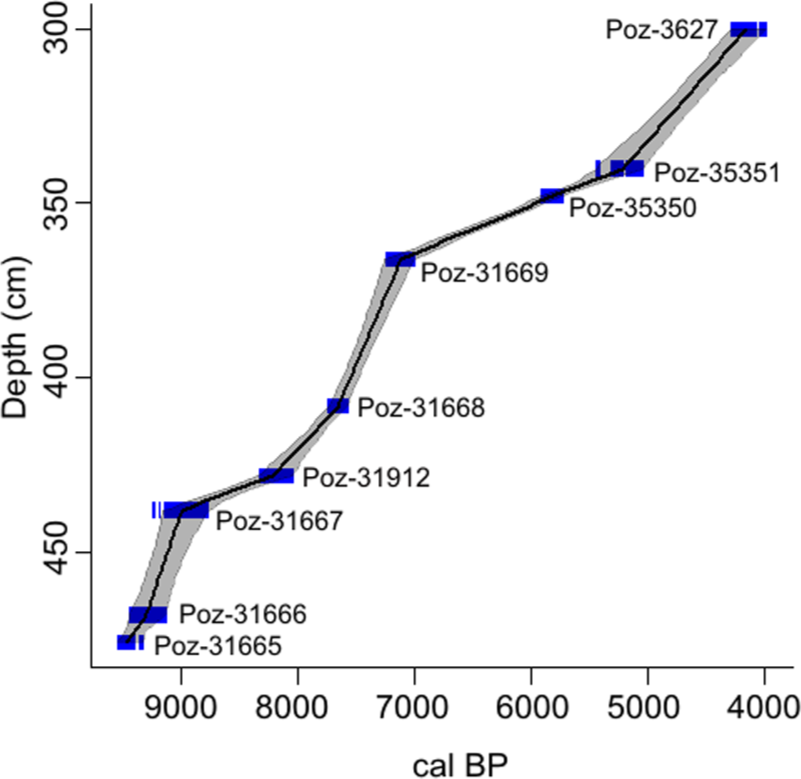
Age–depth model for the peat stratigraphy.
The pollen diagram of dry-land taxa (Table 1, Figure 3) demonstrates three main local pollen assemblage zones (GI/1-3 LPAZ). The most distinct features of this diagram are very high T. cordata pollen and K. deusta spore frequencies, the presence of T. platyphyllos, and very low values of Quercus, Alnus, and Betula in the GI/2 LPAZ dated to 8185±145–5299±195 cal. BP; highly corroded tree pollen among well-preserved sporomorphs and higher mineral matter content are other characteristic features of this section. The pollen concentration is generally higher in the lower part of the profile where Pinus, Betula, and Corylus are the main constituents of the pollen spectra, sharply decreases when Tilia spp. and Ulmus become important, and rises again in the top of the diagram where Corylus and Quercus dominate. The elevated pollen concentrations of all taxa occur at 435 cm, i.e. at the level of deceleration of the sediment accumulation rate (suggested hiatus).
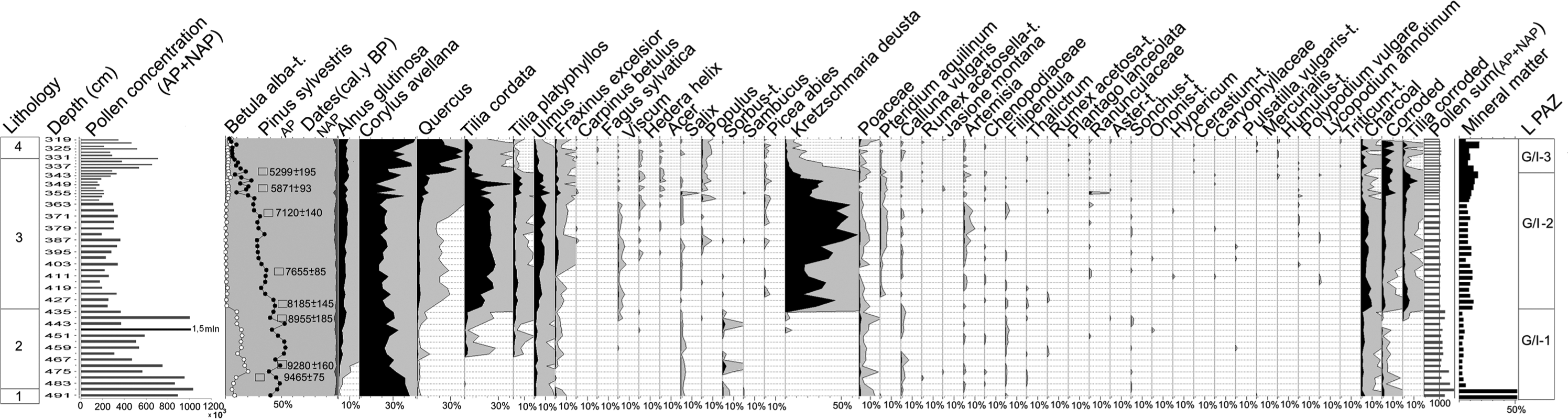
Pollen percentage diagram for dry-land taxa, total AP+NAP concentrations and content of mineral matter. For the lithological units see Table 1.
Three assemblage zones (GI/A–C) describing proportions of sporomorphs of wetland plants and NPPs have been distinguished (Table 1, Figure 4). They illustrate main phases in the bog development and the local ecological conditions during the period covered by the diagram. Also in this case, the middle part of the diagram is distinct because of specific features, mainly the abundance of Cyperaceae, Filicales, HdV-200, Cercophora (HdV-112), and Sordaria (HdV-55A) spores.
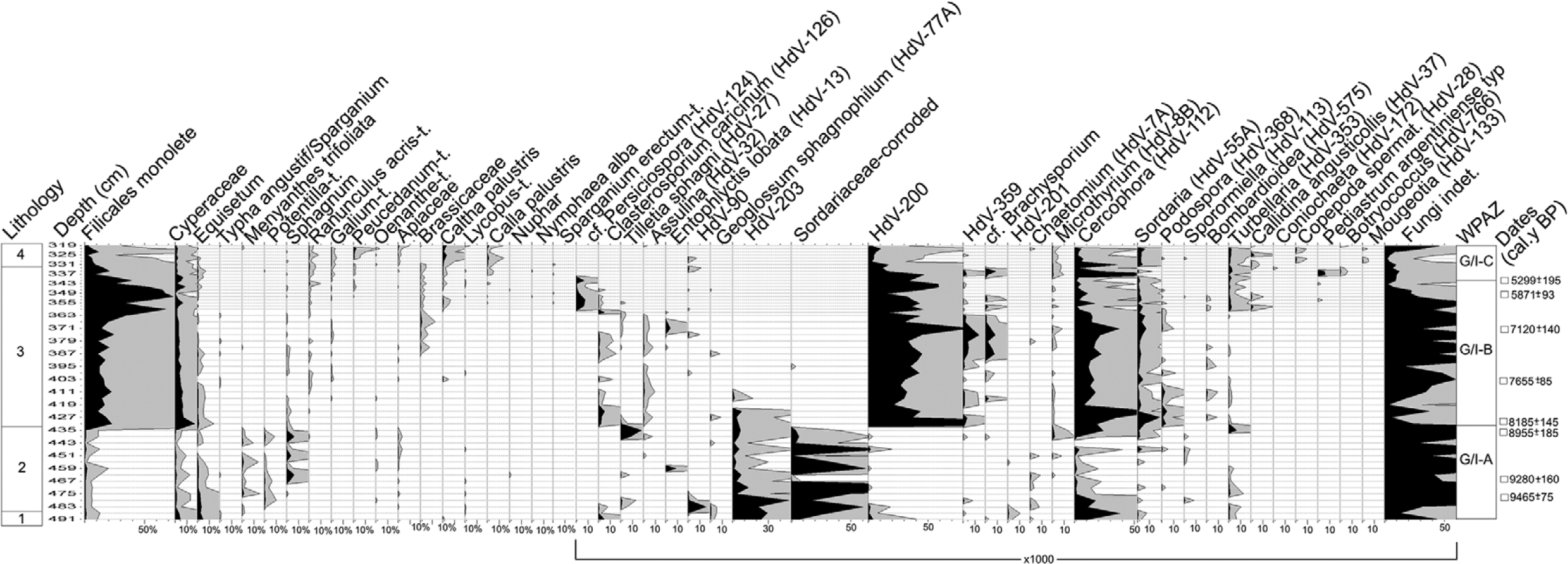
Pollen percentages for wetland taxa and NPPs concentration values. For the lithological units see Table 1.
The sediments are rich in macrofossils. Three macrofossil assemblage zones (GI/I–III) (Table 1, Figure 5) illustrate changes in mire vegetation. This general division follows the pollen assemblage zones delimited in the profile (Figure 3).
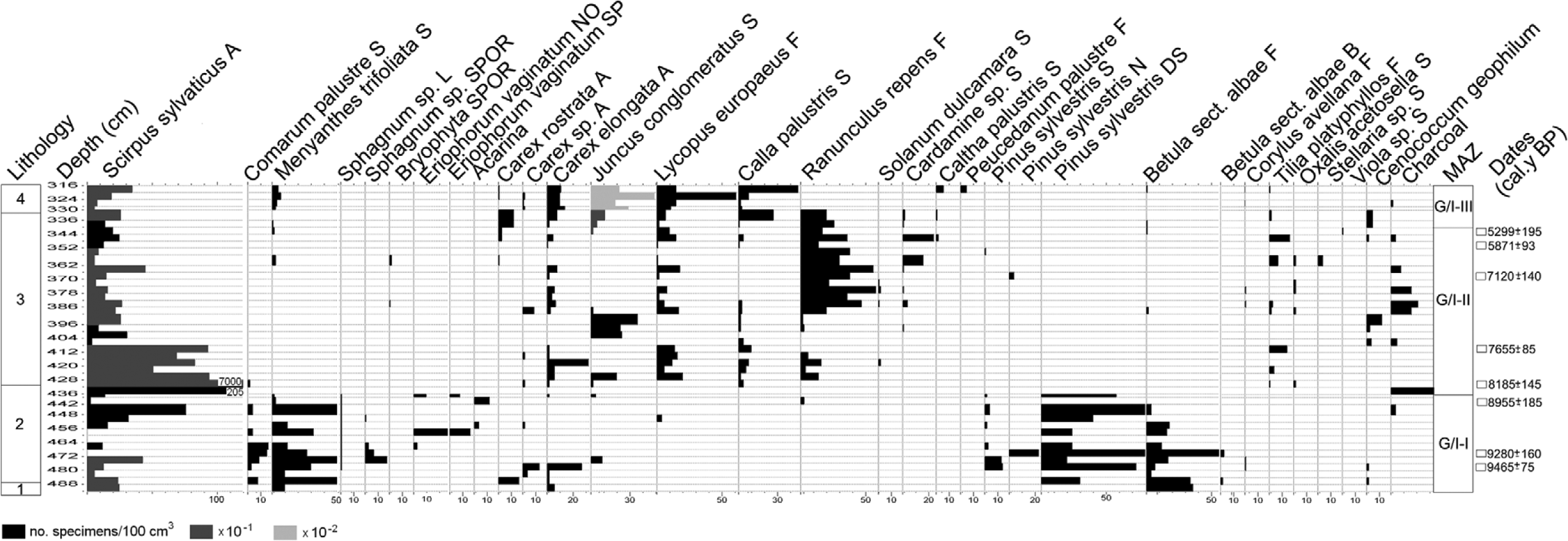
Plant macrofossil diagram. For the lithological units see Table 1. A: achenes; B: bracts; DS: dwarf shoots; F: fruits; L: leaves; N: needles; NO: nodes; S: seeds; SP: spindles; SPOR: sporangia.
Forest succession
In the surroundings of the site, pine and hazel dominated in a period preceding 9465±75 cal. BP; birch and elm were an important admixture. Alnus, Tilia cordata, T. platyphyllos, Fraxinus excelsior, and Quercus started to spread shortly after 9280±160 cal. BP. Higher proportions of Betula and appearances of Sorbus suggest temporary forest disturbances in the period c. 9500–8200 cal. BP, probably because of local fires as shown by the presence of microcharcoal.
The period between 8185±145 and 5299±195 cal. BP can be characterized as a time of Tilia forest dominance. Both T. cordata and T. platyphyllos (pollen and fruits in the subfossil material) were growing in the immediate surrounding of the site, together with C. avellana, Ulmus, and F. excelsior in an admixture. P. sylvestris was probably scarce around the site, and its contribution gradually declined in regional vegetation up to about 6600 cal. BP, when it almost disappeared from the habitats on nutrient-rich soil. Very low proportions of Quercus pollen, oscillating around 2%, indicate that oak stands were scarce in those forests; also, Alnus was insignificant in the local woodland. Viscum was flowering regularly, while Hedera helix blooming became frequent only after c. 7500 cal. BP. Very low proportions of Betula and herbs, including such common taxa as Poaceae, indicate that the forest was rather closed, without distinct openings. A badly developed herb layer favoured soil erosion, as demonstrated by higher mineral matter content in the sediments, input of corroded pollen, and the presence of macrofossils of forest plants (T. platyphyllos, C. avellana, Oxalis acetosella) and sclerotia of Cenococcum geophilum; we may also assume that at least a part of the great amount of Tilia pollen reached the site because of water transport from the local pollen stand. Small-scale forest disturbances accelerated at around 7500 cal. BP, as shown by a higher frequency of Pteridium aquilinum and Artemisia, a small peak of Populus, and an increased frequency of macrocharcoal particles.
It is impossible to reconstruct the history of this forest in the period c. 6600–5300 cal. BP in great detail because of a very low accumulation rate of the peat deposit in the corresponding section of the profile. In that period, population expansion of Quercus and Corylus at the cost of pine and Tilia initiated vegetation changes in neighbouring areas. The frequency of forest fires clearly decreased. At around 5299±195 cal. BP, a Quercus-Corylus community replaced the Tilia forest. The first pollen of Plantago lanceolata appears in the diagram at about 5300 cal. BP, suggesting the presence of the first farmers; however, anthropogenic indicators are scarce up to the top of the diagram.
Kretzschmaria deusta and its palaeoecological context
The whole Tilia forest phase is associated with extremely elevated frequencies of ascospores of Kretzschmaria deusta (Hoffm.) PMD Martin (c. 350,000/cm3 of sediment and up to 60%), suggesting that this parasitic fungus infected most of the potential host populations living in this area. The first ascospores appear together with the beginning of the Tilia spp. pollen curves (c. 9000 cal. BP), a time when pine and hazel were still the most important components of the forest surrounding the site. Their frequency rapidly increases in the same level as the Tilia pollen curve rises, around 8185±145 cal. BP, and then drops first at c. 6600 and then at 5299±195 cal. BP together with the prominent Ulmus and Tilia declines.
The PCA analysis of the interspecies relationships (Figure 6) indicates strong positive correlation between Kretzschmaria and both Tilia species, Ulmus, and Fraxinus. Other tree taxa, including Quercus, Corylus, Betula, and Alnus do not show such a correlation. It is evident from the topmost part of the diagram that Kretzschmaria started to decline when Corylus and Quercus began to replace Tilia (also Ulmus and Fraxinus) in the local forest.
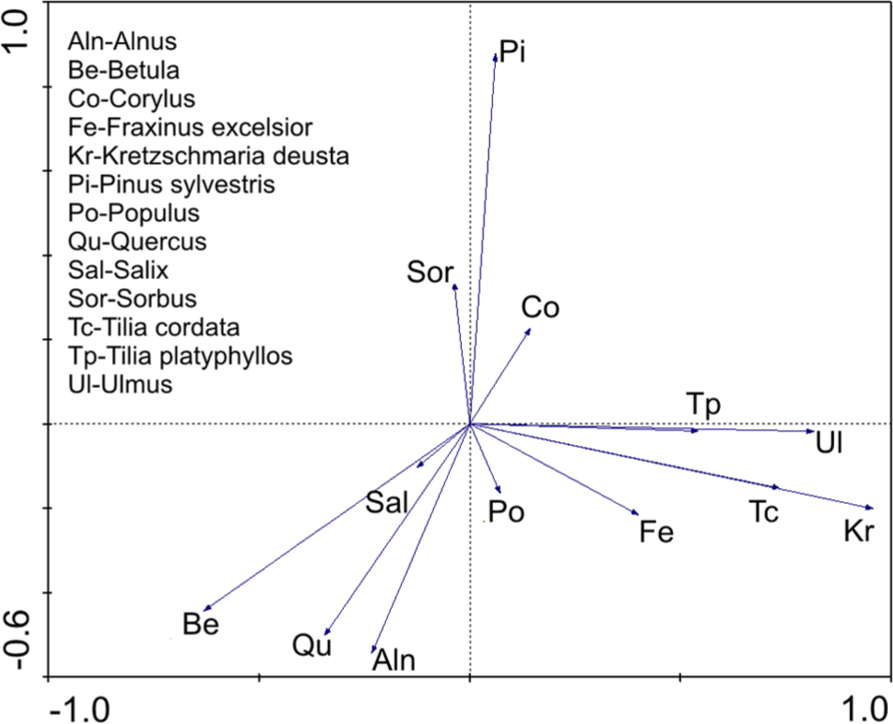
PCA plot illustrating the interspecies relationships in the pollen spectra from Gołębiewo I.
Mire development
In the earliest phase of the mire history, from >9465±75 to 8185±145 cal. BP, a plant community with Scirpus sylvaticus, Equisetum (palustre), Menyanthes trifoliata, Comarum palustre, and Carex elongata developed on this site as a result of paludification of the local depression; from 9280±160 cal. BP, trophic level on the mire was decreasing. Patches of Sphagnum sect. Palustria expanded or contracted according to the moisture of the mire surface; unstable hydrologic conditions are confirmed by the incidental appearances of Eriophorum vaginatum. In drier periods, plants were subject to fungal growth, as confirmed by the abundant presence of hyphae and spores. A hiatus between 8955±185 and 8185±145 cal. BP suggests dry conditions limiting accumulation of the peat deposit or even the partial disappearance of a sediment layer of unknown age.
During the phase 8185±145–c. 5300 cal. BP, the bog surface was covered by vegetation of a wet meadow type with S. sylvaticus, Lycopus europaeus, Ranunculus repens, C. elongata, and Filicales (Thelypteris palustris? Dryopteris carthusiana?) as the most common taxa, illustrating eutrophic and, for at least part of the growing season, relatively dry conditions. These are well characterized by NPPs and especially several types of fungal spores (cf. van Geel and Aptroot, 2006; van Geel et al., 1989). The most distinct feature is a high concentration of HdV-200 type but also Cercophora, Microthyrium (HdV-8B), Pleospora (HdV-3B), and other cellulose-decomposing fungi developing on drying-out stems of mire plants and all kinds of plant remains, all characteristic of dry conditions in mires. Some of these fungi, such as Cercophora, also occur on herbivore dung. Additionally in this phase, typical coprophilous fungi (Sordaria, Podospora (HdV-368), and Sporormiella (HdV-113) appear at much higher frequencies than in the earlier stage, suggesting a low water-table in summer allowing herbivores to visit the site for grazing. In this period, the hydrological situation on the mire varied only slightly. The regular occurrence of Calla palustris in the earlier stage (8185±145–7655±85 cal. BP) points to, at least periodically, a relatively higher water-table while the later expansion of R. repens (c. 7500–5000 cal. BP) arose from the general water-table drop. In this phase, however, several wetter episodes occurred, especially from c. 6600 cal. BP.
In the phase c. 5300–4600 cal. BP, the mire vegetation changed again because of the higher water-table as indicated by the re-appearance of C. palustris and then M. trifoliata, and occurrence of Botryococcus and Pediastrum argentiniense typ. The frequency of fungal spores typical of dry conditions declined.
Discussion
The mid-Holocene Tilia forest
Among the most interesting aspects of this study are the evidence of development of the Tilia forest at 8185±145 cal. BP, its persistence up to c. 5300 cal. BP, and abundant pollen production by lime trees. Similar records in NW and central lowland Europe are rare. Most pollen diagrams indicate relatively low maximum proportions of Tilia (5–10%), with values above 10% interpreted as lime dominance (Huntley and Birks, 1983) according to the poor dispersal of its pollen (Keatinge, 1982). The best data illustrating lime forest history have been obtained through analysis of sediments of small forest hollows and soil profiles in Denmark (Aaby, 1983; Andersen, 1984; Hannon et al., 2000; Iversen, 1969) and Britain (Bradshaw, 1981).
Also worth noting is the well-documented presence of T. platyphyllos on this site situated c. 400 km north of the present geographical range limit of this species (Boratyńska and Dolatowski, 1991). Most of the isolated localities north of this limit, both in N Poland and S Scandinavia, are not clear in respect to origin, and their anthropogenic background cannot be rejected (Dahl, 1998; Zając and Zając, 2001). On the other hand, similar to the data in this paper, T. platyphyllos pollen (Andersen, 1984; Latałowa, 1982) or pollen and macrofossils (Hannon et al., 2000) point to the presence of this species in the mid-Holocene woodlands along the Baltic coastal zone, suggesting that at least some of the recent populations might be relics of a more extended geographical range of this species in the past (Hannon et al., 2000).
In comparison with other tree taxa in NW and central Europe, both Tilia species display high thermal demands for generative reproduction. A daily temperature drop below 13°C destroys flowers of T. cordata, and the minimum temperature at which pollen of this species germinates is about 15°C, but the effective elongation of the pollen tubes, necessary for fertile seed production, occurs at above 21°C (Piggot, 1991). The much more restricted geographical range of T. platyphyllos, which is limited to central and southern Europe, points to its generally higher thermal demands and higher resistance against drought (Ellenberg et al., 1991). On the other hand, both species are rather resistant against low winter temperatures, but in this respect, T. platyphyllos is less so compared with T. cordata (Korotaev, 1994). Despite the high thermal requirements for generative reproduction, the simple presence of both Tilia species is not useful for more precise climate reconstructions because, as a result of vegetative reproduction, they have the capacity for long-term population survival in suboptimal climatic conditions (Huntley and Birks, 1983). However, at least the arrival and then the initial massive population expansion on the site must have resulted from vigorous generative reproduction, which had to take place in optimal climate conditions for both species.
In addition, the very high pollen production by the Tilia forest stand is notable. Apart from climate, light is among the most important factors for lime flower development. In forest stands, they start to bloom when reaching the canopy layer and only the topmost part of the canopy produces flowers (Barzdajn, 1991). Therefore, the massive pollen production by the Tilia forest palaeocommunity, the co-occurrence of Corylus, and the very low proportions of light-demanding Quercus, Betula, and herbs suggest a specific structure of this forest stand. We interpret these data as the presence of a dense, low-canopy forest enabling good light conditions for both limes and hazel but hampering development/regeneration of light-demanding species at the forest floor. Such a structure could have developed with the intensive production of lime offshoots.
Both Tilia species are distinct among other broadleaved trees by their strong ability to produce offshoots and reproduce vegetatively, especially after action of different destructive agents (Faliński and Pawlaczyk, 1991; Piggot, 1991). The offshoots appear immediately at the base of an old tree, replacing it after its destruction, or can develop up to 5 m distant from the maternal tree because of formation of horizontal xylorhizomes. The specimens sprouting from xylorhizomes have an ‘arboreal’ form, and after some years are not distinct from generative specimens. The offshoot growing enables limes not only to rebuild their stand but also to increase their share at other species’ expense in the phytocoenoses suffering from destruction (Faliński and Pawlaczyk, 1991). The most common destructive agents stimulating vigorous vegetative reproduction in limes are browsing and barking damage by cervids, especially by deer, and climatic factors such as catastrophic droughts, and parasitic infection (Faliński and Pawlaczyk, 1991). According to our study, all of the above factors could have played a role in formation of the Tilia forest community in Gołębiewo, with a parasite attack as the most important. In contrast to Innes et al. (2006), given the lack of evidence of human activity for the main period of Tilia dominance, we can exclude anthropogenic factors (coppicing and damage by domestic animals).
The explosive spread of K. deusta and its possible role in maintaining the Tilia dominance
The consistent phase of the elevated values of K. deusta ascospores in the sediments of Gołębiewo I, dated to c. 8185±145–5299±195 cal. BP, is among the exceptional palaeoecological records. The most similar data have been reported by van Geel and Andersen (1988) from a small forest hollow in Denmark illustrating Tilia forest succession in the mid-Eemian interglacial. In other sites of the Eemian (van Geel et al., 1986) or Holocene age (Innes et al., 2006; Kuhry, 1985; van der Wiel, 1982) in which Kretzschmaria appears in relatively high frequencies, these values are much lower and concern shorter periods. Van Geel and Andersen (1988) have emphasized that Kretzschmaria is usually under-represented in palaeoecological records because its fruiting bodies are located close to the ground, which is a limiting factor for more distant spread of ascospores. According to our hypothesis, the Kretzschmaria ascospores, which are produced from autumn to early spring (Wilkins, 1938), were accumulating on snow or on the ground surface and then were washed out into the basin with snowmelt. It is the only possible scenario for the massive spore dispersal onto the mire surface, which in the growing season was covered with a dense vegetation cover.
K. deusta is a parasitic fungus of Xylariaceae infecting a wide range of broadleaved tree taxa (Wilkins, 1934); however, different species are more or less resistant to the infections. Tilia, Fagus, Ulmus, Fraxinus, and Populus are particularly susceptible while, e.g. Quercus and Corylus are among the most resistant against this parasite (Baum and Schwarze, 2002; Baum et al., 2000; Schwarze and Baum, 2000; Wilkins, 1934). Our PCA analysis results for the interspecies relationships (Figure 6) strictly follow the groups separated by Wilkins (1934) according to the ability of K. deusta to infect particular tree species, but surprisingly all of the most susceptible taxa are positively correlated with the fungus and the resistant ones are in the opposition. A very high positive correlation between Kretzschmaria and Tilia is of special interest here because of the long period of c. 2700 years in which extremely high Kretzschmaria frequencies were concurrent with Tilia’s absolute dominance in the local forest. Such a phenomenon indicates that the Kretzschmaria infection did not negatively affect Tilia population survival. Van Geel and Andersen (1988) drew a similar conclusion from their work on K. deusta occurrence in Eemian deposits in Denmark. However, our hypothesis about the relationships between Tilia forest dynamics and the long-lasting high incidence of the fungus goes further. We suggest that Kretzschmaria infection could have positively affected Tilia dominance in the local forest. This hypothesis is based, first of all, on the ecological features of Tilia and specifically on its capacity for clonal reproduction of a wounded tree.
K. deusta is a mild, opportunistic parasite that mostly colonizes fresh sapwood at the base of old stumps and exposed roots after wounding (e.g. root damage) or by taking advantage of physiological stress (e.g. drought) (Deflorio et al., 2008; Rogers and Ju, 1998). In Gołębiewo, the abundance of coprophilous fungi remains during the Tilia-Kretzschmaria phase, indicates that the mire could attract game as a natural pasture. This open space was especially important because of the lack of herbs under the closed-canopy Tilia forest. Animals could damage trees around the mire through browsing and barking, enabling better spread of the fungus (Innes et al., 2006). Judging from the distinct drop of the water-table on the mire during the period of high K. deusta incidence, hydrological stress also should be considered as a factor accelerating disease spread. According to Baum and Schwarze (2002), lime has a very low ability to compartmentalize decay fungi, and fast growth of Kretzschmaria through the reaction zone is thus observed; however, Wilkins (1939) indicated that on young limes, the fungus may grow for a longer time without negative consequences for the tree. Because lime trees are not resistant against pathogens, their ability to continuously rejuvenate by offshoots prevents population decline (Faliński and Pawlaczyk, 1991). Our data from Gołębiewo seem to illustrate this process and suggest that infection by Kretzschmaria could be the main agent stimulating the vigorous vegetative development of Tilia at the site.
The fungal disease spreads through the root contacts by conidia or by ascospores. The season and climatic requirements for germination of both types of propagules are crucial for our interpretation. K. deusta is the only representative of the genus extending its geographical range beyond the tropics and subtropics, up to the temperate regions (Rogers and Ju, 1998). Its provenance from the warmer climates is underlined by specific, high thermal requirements. According to Wilkins (1938), production and germination of ascospores take place during October–March and require warm climatic conditions (10–35°C, with an optimum between 25°C and 30°C); thus, in central and northern Europe where autumns and winters are much colder than that, these thermal requirements are a limiting factor for disease transmission by ascospores. In contrast to ascospores, conidia are produced in warmer seasons, from the beginning of April until September or October (Wilkins, 1939), a period of potentially favourable conditions for their germination. They germinate in a range of temperatures from 10°C to 30°C, with an optimum between 20°C and 25°C. Wilkins (1939) points also to rainfall and relative humidity as the most important factors for effective production of conidia and their germination and infection of a host; in south Britain conditions, a well-distributed weekly average of not less than 0.5 inch (c. 12.5 mm) (c. 56 mm/month) of rain is essential for optimal conidia production and development. We may assume that the massive occurrence of K. deusta in Gołębiewo between 8185±145 and 5871±93 cal. BP took place in the optimal climatic conditions for its reproduction. Therefore, monthly precipitation in a warm period of a year was not less than 56 mm, i.e. within the recent limits for the period of May–September but above the monthly precipitation values for April (Owczarek et al., 2007). The monthly summer temperatures should be above 20°C, i.e. distinctly higher than the recent mean temperature of the warmest month, 17.0°C (Owczarek et al., 2007). The humidity conditions for the development of the fungus probably occurred close to the ground even in higher summer temperatures because of the dense structure of the forest stand and relatively low transpiration rate of Tilia trees (Popowa, 1985). It is worth noting that this phase had no analogue in the earlier or later periods of the history of the site. Even during the last millennium when Fagus, the ideal potential host for this fungus next to Tilia, dominates in the forest stand surrounding the site, only single ascospores irregularly appear in the pollen spectra (Pędziszewska, 2007).
The mid-Holocene climate
The combined record of the abundant occurrence of T. cordata and T. platyphyllos, high incidence of K. deusta, and hydrological changes on the mire give some important hints for the climate reconstruction in the period c. 9500–4600 cal. BP, illustrating three major and some minor climatic phases.
Phase 1 (c. 9500–8185±145 cal. BP) started with an increase in the groundwater-table, which initiated paludification in the local depression. Therefore, the development of a mesotrophic mire resulted from higher climate wetness compared with earlier in the period. This inference seems to be consistent with the mid-European phase of higher lake levels at 9550–9150 cal. BP (Magny and Bégeot, 2004), reflected also in the laminated sediments of Lake Gościąż, central Poland (Starkel et al., 1996). The unstable groundwater-table and decreasing trophy of the local habitats suggests a slightly drier climate after 9280±160 cal. BP, at least temporarily. Unfortunately, a drop in the peat accumulation rate or a hiatus between 8955±185 and 8185±145 cal. BP makes climate reconstruction for this time-slice impossible. The potential hiatus could have resulted from decomposition of a previously accumulated peat layer because of climate drought in the following period. We note, however, that the 8.2 ka cold event (Alley et al., 1997) is well expressed in other sites in the Kashubian region (Weisbrodt, 2011).
Phase 2 (8185±145–5299±195 cal. BP) is distinct by the T. cordata and T. platyphyllos population expansion and K. deusta infection taking place immediately after the 8.2 ka event, i.e. following the climate warming. The abundant occurrence of these species must have taken place in optimal climatic conditions for reproduction. As argued above, average summer temperatures were certainly higher than today by probably more than 3°C, exceeding 20°C. High temperatures resulted in summer drought, however, at least in a part of the spring–summer season, regular precipitation must have assured suitable relative humidity at the forest floor, stimulating K. deusta development. During this phase, the mire surface was distinctly drier than in the earlier period. Large proportions of the HdV-200 fungal spores, indicative for drying-out plant matter (van Geel et al., 1989), suggest that the water level was seasonally variable – higher in spring, enabling mire vegetation development, and lower in summer when high temperatures resulted in intensive evapotranspiration and plant drying and decay. Our data indicate a rather stable climate during the whole phase, with only minor fluctuations. As shown by the situation on the mire, probably between 8185±145 and 7655±85 cal. BP, the climate was generally slightly wetter, and then drier between c. 7500 and 5300 cal. BP, with some wet episodes. In an ombrogenic peat profile from Stążki (the same region), Lamentowicz et al. (2010) described six wet phases in this period. The intensive slowing down of the effective peat accumulation rates between c. 7500 and 5300 cal. BP is a common occurrence in NW Poland (Lamentowicz et al., 2010; Latałowa, 1982, 1992), indicating at least temporal water deficits. A strong decline of K. deusta and both Tilia species at 5299±195 cal. BP suggests a climate worsening for these taxa. We would like to refer here to the pollen data by Seppä et al. (2005) from south-central Sweden, which suggest relative cooling at 6100–5400 cal. BP, and to several other records indicating cold and wet events in the period 5600–5300 cal. BP (Magny, 2004; Magny and Haas, 2004).
The whole phase 8185±145 to 5299±195 cal. BP represents the Holocene Thermal Maximum (HTM). Both proxy data (Davis et al., 2003; Magny et al., 2003; Mayewski et al., 2004; Wanner et al., 2011) and model simulations (Brewer et al., 2007; Renssen et al., 2009) indicate a temporal and regional complexity in which – apart from orbital forcing and ‘behaviour’ of the Northern Hemisphere ice sheets affecting ocean and atmospheric circulation – latitudinal, continentality, and elevation gradients played an important role. Our results seem to be close to those from south Scandinavia and the eastern Baltic region; however, according to our data, the HTM came to an end earlier. In south-central Sweden, both oxygen isotope data (Hammarlund et al., 2003) and pollen-based climate reconstruction (Seppä et al., 2005) indicate warm (Tann. c. 2.5°C higher than today) and markedly dry conditions between c. 8000 and 4300 cal. BP. Similar results have been obtained through pollen-based reconstruction for Estonia (Seppä and Poska, 2004) while the HTM for Latvia (Heikkilä and Seppä, 2010) has been described as a period 8000–4000 cal. BP with Tsummer 2.5–3.5°C higher than modern values; the highest anomalies occurred between c. 7500 and 5000 cal. BP (+3–3.7°C), falling exactly into the driest period as recorded in Gołębiewo.
Phase 3 (5299±195–4600 cal. BP) represents unstable hydrological conditions starting with a drier phase up to c. 5000 cal. BP followed by increasing moisture of the local habitats that probably was caused by climate change. Although both Tilia species still are present in this phase, their rapid decrease and replacement by Quercus and Corylus might have resulted from climate cooling, limiting their flowering and generative reproduction. As has been argued, a very weak presence of human indicators cannot alone explain the scale of this event, as demonstrated by the pollen diagram. This phase fits roughly with cool and wet climate events described in many areas (Greisman and Gaillard, 2009; Magny, 2004; Starkel et al., 1996) and reflected in the regional data (Lamentowicz et al., 2010).
Conclusions
The palaeoecological data from Gołębiewo, and specifically changes in the local forest stand, long-lasting infection by the parasite fungus K. deusta, and changes in the mire ecosystem, afford several arguments for the reconstruction of mid-Holocene climatic conditions in the Kashubian Lake District. Our data are in accordance with the geographic pattern of the HTM temperature anomalies in northern Europe, including the Baltic region, which suggest gradients of positive anomalies from north to south and from west to east (Heikkilä and Seppä, 2010). According to these authors, such gradients might result from differential heating of the continental surface and the Atlantic Ocean, with its magnitude modified by atmospheric circulation. They suggest that the hot and dry summers of the HTM might be an effect of stable summer anticyclonic circulation in this area, possibly analogous with modern blocking anticyclonic conditions.
The data from Gołębiewo seem to illustrate the impact of climate warming on pathogen outbreak and suggest intensive vegetative reproduction as a Tilia strategy for population survival. Summer drought was probably important for the high success of K. deusta in infecting weakened Tilia trees. We may assume similar mechanisms and similar climatic conditions resulting in the abundant co-occurrence of Tilia and K. deusta in the mid-Eemian deposits in Egernsund/Denmark (van Geel and Andersen, 1988). Furthermore, our data may be a ‘lesson for the future’ concerning the increasing risk of K. deusta outbreaks in the scenario of present and forecasted climate warming and increasing dryness in many areas of deciduous woodland.
Footnotes
Acknowledgements
The authors are grateful to Bas van Geel for his comments on the manuscript.
Funding
The work was supported by the Rector of the University of Gdańsk through grant BW/L145-5-0096-9.