
Abstract
The steep, wet, forested, slopes of the mid-elevation Andes have often been dismissed as inimical to human occupation. A lack of ancient lakes in these habitats has limited the availability of paleoecologists to contribute to this discussion. Here, we present fossil pollen and charcoal data from Lake Pomacochas in northern Peru, which is a 70-m-deep lake lying at 2100 m a.s.l. We present a c. 3500-year history of both significant climate change and site occupancy by humans. Maize pollen is present in sediments throughout much of this record, though there are times when its cultivation is either abandoned, or moves back from the shoreline, for example, between c. 3500 and 2700 cal. yr BP and post 1200 cal. yr BP. By comparing our record with isotopic data derived from cave calcite we are able to show that dry times favored maize cultivation in this setting. In the last 1200 years, large-scale social changes in the Andes, such as the emergence of the Chachapoya culture and the invasion of Europeans, appear to be reflected in the fossil pollen and charcoal record.
Keywords
The role that climate change has played in Andean cultural history is actively debated. Some researchers view climate as playing a pivotal role in cultural transitions, for example, the end of monument building by the Tiwanaku (Ortloff and Kolata, 1993), Maya (Hodell et al., 2005), and Moche (Moseley and Deeds, 1982). Drought is also suggested to have driven the mass migrations of human populations that resulted in the mid-Holocene (c. 8000–4000 cal. yr BP; all ages are given in 14C calibrated years Before Present (cal. yr BP) taking present to be 1950) dearth of archaeological sites in the Chilean Andes (Núñez et al., 2001). Lake records show substantially lowered lake levels between c. 8000 and 4500 cal. yr BP stretching from Ecuador to Chile (Abbott et al., 1997; Ekdahl et al., 2008). Aridity on this scale, especially in the desert settings investigated by Núñez et al. (2001) and in dry intermontane valleys may have stymied human occupation. Shorter events, on the scale of decades that induced massive flooding or droughts have been identified in ice-core and sedimentary records (Bird et al., 2011a; Thompson et al., 1986, 2013). These rapid events have been invoked to explain the collapse of the Moche (c. 1900–1250 cal. yr BP; Moseley and Richardson, 1992) and Tiwanaku (c. 1600–950 cal. yr BP) cultures (Binford et al., 1997; Ortloff and Kolata, 1993). Some archaeologists cite evidence of drought to help explain both the rise and decline of the Wari empire (chapters in Bergh and Castillo, 2012).Evidence of emigration from the Andes into the coastal lowlands c. 750 cal. yr BP coincident with the collapse of the Tiwanaku and Wari cultures is suggested on the basis of DNA evidence (Hernández-Ugalde et al., 2011). Archaeologists who are unconvinced that climate underlay these cultural transitions refer to these arguments as ‘environmental determinism’ (Coombes and Barber, 2005; Erickson, 1999; Roosevelt, 2013).
Alternative viewpoints to that of ‘environmental determinism’ emphasize the capacity of people to respond to climatic and environmental crises through adaptive innovations (Erickson, 1999; Freilich, 1967). Abrupt cultural transitions may also be explained through internal social strife, or external conflict. The role of warfare is clearly evident in the military subjugation of the Chanka (c. 750–512 cal. yr BP) and the Chachapoya (c. 1200–470 cal. yr BP) cultures by the Inca (512–417 cal. yr BP), who in turn were conquered by the Spanish in AD 1533 (417 cal. yr BP; Arkush and Tung, 2013; Bauer et al., 2010; Church and Von Hagen, 2008). With some evidence appearing to support both sides of the discussion, the importance of climate change to past Andean cultures remains uncertain (Gosling and Williams, 2013).
Prehistoric agriculture in the central Andes
Agriculture has been practiced in the Andes for at least the last 10,000 years. The earliest crops included squash (Cucurbitaceae), cotton (Malvaceae), peanut (Fabaceae), and a quinoa-like pseudo-cereal (Amaranthaceae) and are known from the western Andes (Dillehay et al., 2007). Maize was introduced into Amazonia by 6000 cal. yr BP (Bush et al., 1989), and camelids were domesticated between 6000 and 4000 cal. yr BP (Stahl, 2008). Human modification of Andean landscapes reveals engineering ingenuity. Irrigation using small-scale gravity canals appears in northern Peru by 6000 cal. yr BP (Dillehay et al., 2005). In the Bolivian Andes, canal irrigation has been practiced for the last 3500 years, and the Tiwanaku used flooded raised fields to increase the growing zone of crops around Lake Titicaca (Dillehay and Kolata, 2004; Kolata and Ortloff, 1989), and terracing of steep slopes was practiced by the Wari c. 1450–950 cal. yr BP (Kendall, 2013). The effort invested in landscape manipulation reveals an impetus to overcome adversity.
Contributing to the reconstruction of the Andean past, paleoecologists have documented the addition of maize to the highland crop repertoire between c. 3200 and 2800 cal. yr BP coinciding with a decline in quinoa abundance in the Peruvian Andes at sites lying above 3050-m elevation. This transition was interpreted as response to wetter climates in the dry intermontane valleys of the Andes (Mosblech et al., 2012). A further cultural adaptation, in response to a need for wood, may have been the adoption of silviculture (Chepstow-Lusty and Winfield, 2000). Records from Lakes Marcacocha (3400-m elevation), Pacucha (3050-m elevation), and Huaypo (3500-m elevation) show a marked increase in the abundance of Alnus pollen between 1300 and 1000 cal. yr BP, clearly preceding the rise of the Inca c. 600–500 cal. yr BP (Mosblech et al., 2012).
In northern Peru and southern Ecuador, lakes within the range of modern cloud forests, for example, Lake Chochos (c. 3200-m elevation; Bush et al., 2005), and Lake Llaviucu (3180-m elevation; Colinvaux et al., 1997), did not reveal any conclusive evidence of agriculture. In the adjacent lowlands, however, at Lake Ayauchi (350-m elevation) in Ecuador, maize agriculture was evident for more than 6000 years (Bush et al., 1989).
Archaeologists had once considered the Chachapoyas region, between the Huallaga and Marañon Rivers, too wet and densely forested to sustain independent human occupation and cultural development (Bonavia, 2000). Investigations and discoveries over the last several decades, however, have revealed semi-permanent occupation since c. 3950 cal. yr BP in the upper montane forests (Church, 1996), with settlement and possibly early maize cultivation deep within the cloud forests by c. 2300 cal. yr BP (Church, 1994; Church and Von Hagen, 2008). Such early settlements have not been assigned to any larger regional culture, but between c. 1150 and 850 cal. yr BP a common Chachapoya cultural identity formed, although the extent of sociopolitical unity remains uncertain prior to Incan conquest c. 500 cal. yr BP (Church, 1994; Church and Von Hagen, 2008; Nystrom, 2006).
Climate change in the last 4000 years
High-resolution sedimentary, ice-core, and cave calcite isotopic records provide proxy records of precipitation for the late Holocene (Apaéstegui et al., 2014; Bird et al., 2011a; Ekdahl et al., 2008; Kanner et al., 2013; Moy et al., 2002; Thompson et al., 1992, 2013). These records showed an overall wetting trend as lake levels rose from their lowstands at 5500 cal. yr BP and were mostly at or close to modern levels by c. 4000 cal. yr BP (Baker et al., 2005). Local variability in climate tended to dominate the signal thereafter. Sites that responded strongly to El Niño Southern Oscillation (ENSO) forcing generally showed an uptick in activity around 2800 cal. yr BP with a peak in activity around 2000 cal. yr BP (Conroy et al., 2008; Moy et al., 2002). The manifestation of ENSO was spatially heterogeneous, that is, El Niño (La Niña) events were wet (dry) in coastal settings, and dry (wet) in the Central Andes (Vuille et al., 2000). Some sites were simply more sensitive to this Pacific forcing than others according to whether their precipitation source was primarily from the Pacific Ocean or the Atlantic Ocean (Bird et al., 2011b; Moy et al., 2002).
Within the last 1500 years, The Medieval Climate Anomaly and the ‘Little Ice Age’ were both represented by precipitation anomalies in the Andes. Most Central Andean proxy records indicated that the period between c. 1200 and 1500 cal. yr BP was wet, with some drying evident in the last 150 years (e.g. Apaéstegui et al., 2014). The peak of the Medieval Climate Anomaly c. 1000 cal. yr BP was a time of decadal scale droughts in parts of the high Andes, producing the strongest isotopic signatures of the past several thousand years (Thompson et al., 1986, 2013).
Here, we present a detailed record of fossil pollen and charcoal to reconstruct vegetation and agricultural history from 2000-m elevation. We compare our findings with independent proxies for regional climate change and seek to contribute new data to the ongoing discussion relating to impacts climate may have exerted on the development of pre-Columbian montane forest societies in northern Peru.
Methods
Study site
Laguna Pomacochas (5°49.63′S; 77°57.44′W; 2150-m elevation) is a large lake spanning c. 2.5 km from east to west and 2.3 km from north to south (Figure 1). Pomacochas has a mean annual temperature of 15.1°C, and the precipitation is 881 mm (mean from 1969 to 1973) with a 4-month dry season between May and August (Vargas, 2010). The surrounding geology consists of limestone, suggesting the lake basin may be an old sinkhole created by dissolution of underlying bedrock. A Tortora marsh surrounds the lake and ranges in width from 10 to 150 m. The Tortora gives way to a shallow shelf (c. 3- to 4-m water depth in July 2010) that quickly drops to maximum lake depths of 70–80 m (Figure 1). A bathymetric map was created using a Garmin GPS/sonar. The watershed consists of slopes that extend c. 1.5–3 km beyond the lake shores, with ridges reaching altitudes of 2700 m a.s.l. An outlet drains the lake to the north, feeding into the Chiriaco River. Pomacochas residents report recent changes in lake levels, influencing the width of the surrounding marsh.
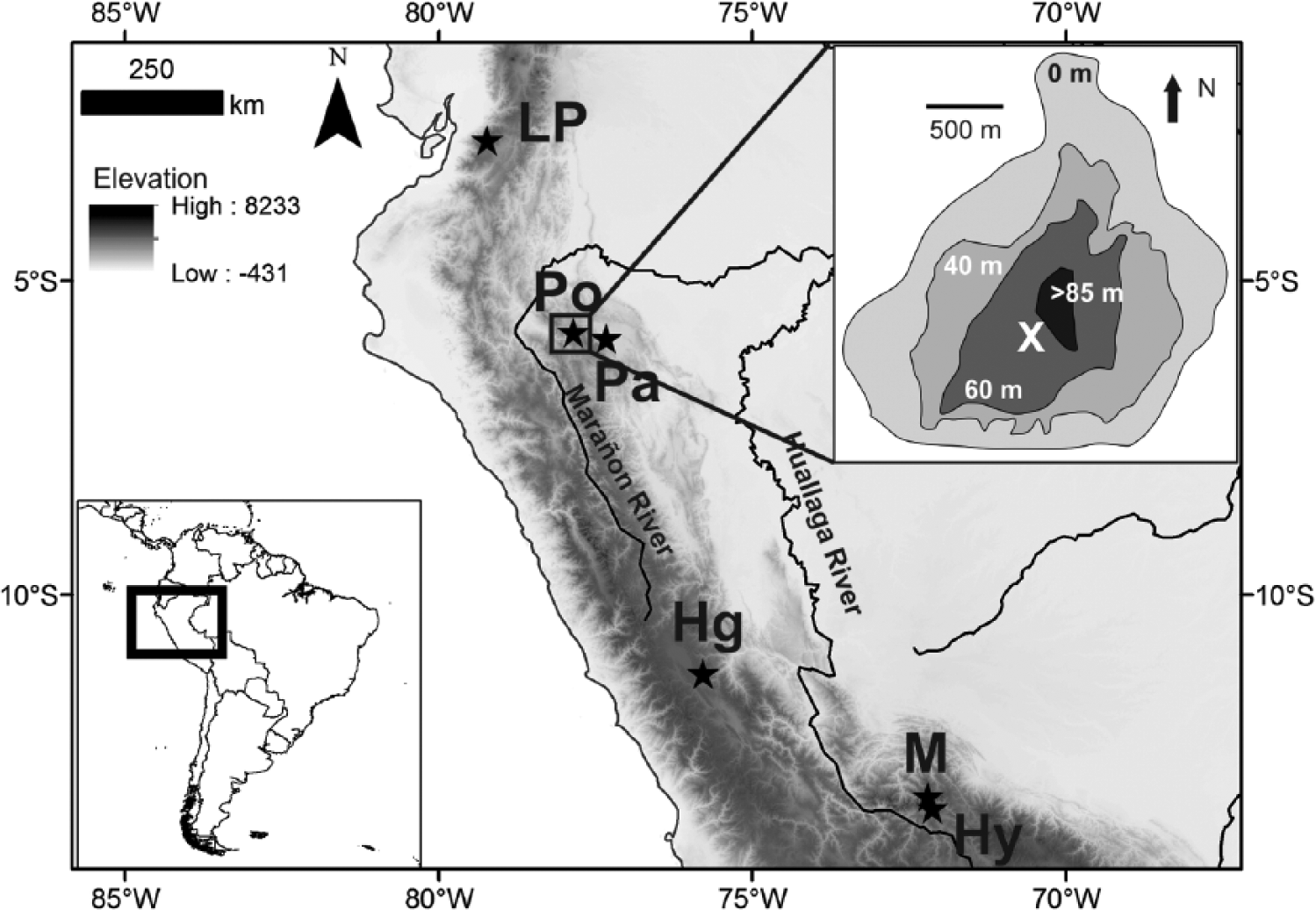
Topographic basemap showing the Peruvian and Ecuadorian Andes and adjacent Amazonia. Inset map top right shows the bathymetry of Lake Pomacochas. Stars denote the locations of lake and cave records mentioned in text. Pomacochas is marked by a circled star. Ch = Lake Chochos, Hg = Huagapo Cave, Pa = Palestina Cave, Pc = Lake Pacucha, Po = Lake Pomacochas, LP = Lake Pallcacocha, M = Lake Marcacocha, Hy = Lake Huaypo.
Around Pomacochas, the modern landscape has been cleared for agropastoral purposes. Local archaeological sites have been identified on the ridges to the southwest and southeast of Pomacochas, but are not yet dated or extensively studied. On the eve of the Spanish conquest of 1532, Inca imperial forces fought and defeated Chachapoya inhabitants of the lake basin identified as the ‘Indians of Pumacocha’ (De Marúa, 2008 [1613]).Despite the present lack of additional archaeological data, it is not unreasonable to expect human occupation and landuse spanning well-prior to the c. 3500 cal. yr BP basal radiocarbon estimate from the lake sediment core.
Introduced grasses and exotic trees, for example, Eucalyptus spp., dominate the vegetation of the modern basin and surrounding hillsides. Modern crops include a variety of potatoes, but maize is not grown around Pomacochas. Surrounding ridges contain patches of montane cloud forest, particularly on inaccessible slopes. Forests at this elevation would be expected to contain a diversity of species within Rubiaceae, Lauraceae, Melastomataceae, Podocarpus, Myrtaceae, Urticaceae, Hedyosmum, and Ericaceae. Tree branches would support a broad range of epiphytic vascular plants and mosses. Alnus and Cecropia are very abundant pollen types in forested settings at this elevation (Weng et al., 2004a, 2004b).
Immediately downslope of the Pomacochas Basin is a large protected area of forest that still retains a high diversity of species. The prevailing easterly winds are likely to transport anemophilous pollen (e.g. Urticaceae/Moraceae, Alnus, Cecropia) from these slopes to the lake.
A 2.9-m sediment core was obtained from Laguna Pomacochas in July 2010. The lake basin had a broad flat bottom suitable for coring, with minimal risk of obtaining slumped sediments. Due to the depth of the lake, 78 m at the coring site, a 69-mm diameter universal corer (Aquatic Research Industries) was used with a 4-m-long core barrel. The third core (PomaC3) yielded the most complete record and was used for the current study. The surface mud–water interface and the top 4 cm of unconsolidated sediment were collected and stored in whirlpaks. The top and bottom of the core were capped and sealed for transport to the Florida Institute of Technology. The cores were stored in a cold room (+4°C) when not in use.
Chronology
The Pomacochas chronology was established using Accelerator Mass Spectrometry (AMS) radiocarbon dates. No terrestrially derived organic macrofossils were found, and so five bulk sediment samples were dated. A chronology was constructed using the bacon package (Blaauw and Christen, 2011), with the 14C ages calibrated using IntCal13 (Reimer et al., 2013). IntCal13 was chosen rather than SHCal14 as the moisture arriving to Pomacochas originated largely from the subtropical Atlantic and the Amazon Basin (Vuille et al., 2003) and did not need a southern hemispheric correction. Ages for the record Pallcacocha (Moy et al., 2002) were similarly recalibrated.
Sediment analyses
Sediment colors were identified using Munsell color charts and imaged using a Geotek core logger (University of Florida). Subsamples of 0.5 cm3 were obtained for pollen analysis at 1- to 5-cm intervals from the Pomacochas C3 core. A total of 81 sediment subsamples provided c. 50- to 80-year temporal resolution. Samples were spiked with exotic Lycopodium tablets to calculate pollen concentration (Stockmarr, 1972) and treated with 10% HCl, KOH, NaP2O7, and HF, according to standard procedures (Faegri and Iversen, 1989). Processed samples were mounted in glycerin. Pollen counts of 300 grains (excluding aquatic taxa) were performed under 400× magnification using a Zeiss photomicroscope. If pollen concentration was low, counts were terminated after reaching 2000 Lycopodium spores. Pollen identification was aided by comparisons with a pollen reference collection of >3500 types, the Neotropical Pollen Database (Bush and Weng, 2007) and published references (Heusser, 1971; Hooghiemstra, 1984; Markgraf, 1978).
Charcoal samples were taken at continuous 1-cm intervals from the Pomacochas C3 core. After being heated in KOH and NaP2O7, the samples were filtered with 180-µm mesh (Clark and Royall, 1996). The filtered particles were examined using an Olympus stereoscope at 20× magnification. Charcoal fragments were photographed, and their surface area was calculated using ImageJ software (Rasband, 2005).
Loss on ignition was performed on 0.5-cm3 sediment samples at 1- to 10-cm intervals to provide a c. 70-year temporal resolution for organic matter and carbonate concentration. The sediment samples were placed in dry crucibles and weighed after 12 h at 105°C (mass of water loss), 4 h at 550°C (organic matter mass), and 2 h at 950°C (carbonate mass; Heiri et al., 2001). All results were plotted using C2 (Juggins, 2003).
Statistical analyses
Palynological results were analyzed using detrended correspondence analysis (DCA) to identify patterns in ecological change, using the vegan and paleoMAS packages within the program R (Correa-Metrio et al., 2010; R Development-Core-Team, 2011). To reduce noise in the dataset, the DCA was run on taxa that occurred in at least five samples and that attained at least 2% abundance (Birks and Gordon, 1985); this filter reduced the matrix to 34 taxa.
Results
Age model and sediment stratigraphy
The 2.96-m-long core raised for the center of the lake represented the limits of our coring capacity, and did not reach bedrock. Five bulk sediment radiocarbon dates showed that the 2.96-m sediment core from Laguna Pomacochas spanned c. 3530 cal. yr BP (Table 1; Figure 2). The resulting age model contained no reversals and indicated a decrease in the sedimentation rate at c. 1130 cal. yr BP.
Radiocarbon dates and calibrated ages using IntCal13 (Reimer et al., 2013) for bulk sediment samples from Lake Pomacochas, Peru.
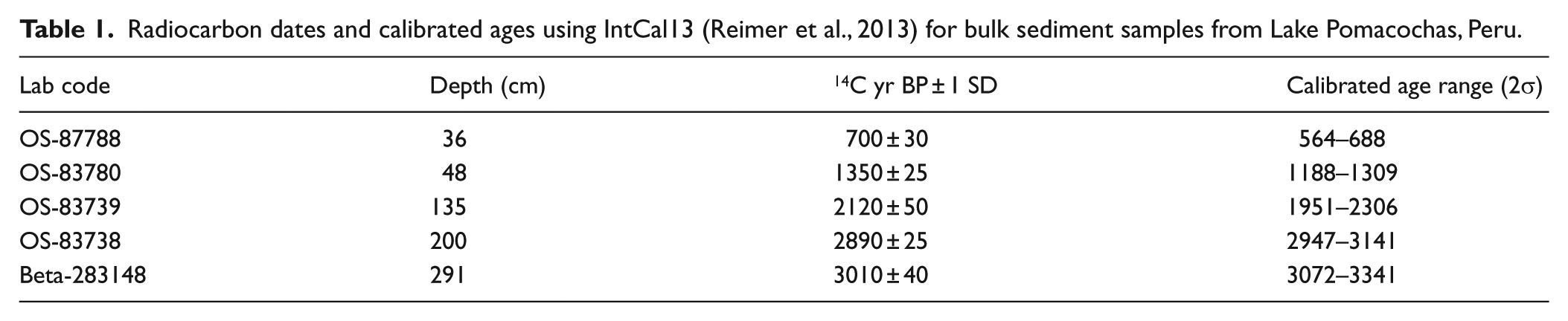
The stratigraphy, loss on ignition, and age–depth plot from bacon (Blaauw and Christen, 2011) for sediments from Lake Pomacochas, Peru.
The core was divided into two primary sedimentary units at a depth of 47 cm. The older unit (46–296 cm; 1140–3530 cal. yr BP) consisted primarily of laminated gyttja with one 1-cm layer of sand from 144- to 145-cm depth (c. 2250 cal. yr BP), which were all in the color range of very dark gray to olive gray. The upper unit (0–46 cm; modern to c. 1140 cal. yr BP) was much darker in color (black to olive gray) and was mostly solid gyttja, except for a section from 22 to 14 cm (520–300 cal. yr BP) of laminated gyttja.
Ordination
A total of 34 pollen types were included in the DCA ordination. The ordination of the 81 samples provided an output that showed considerable clustering. The samples were divided into six temporal groups that minimized overlap with neighboring groups. The resulting groups formed the pollen zones. A further three subzones were identified (Figure 3) based on their spatial and temporal separation.
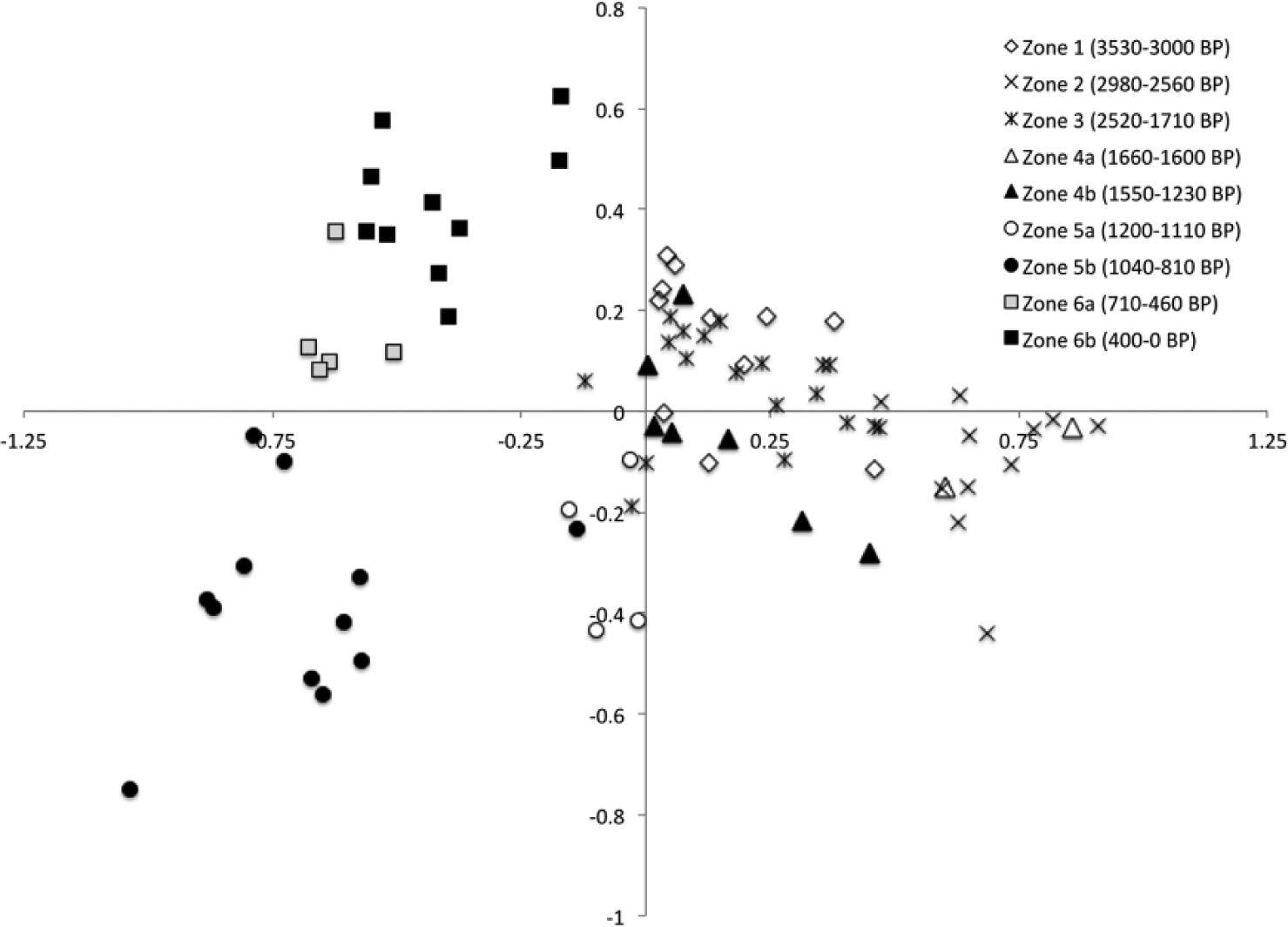
Plot of sample scores from a detrended correspondence analysis of percentage data for 34 pollen taxa found in the sediments of Lake Pomacochas, Peru. Samples were subdivided temporally to minimize spatial overlap. Groupings form the basis for the zonation of the pollen diagram.
The species ordination resulted in the positive extreme of Axis 1 being characterized by weedy trees, herbs, and maize, for example, Sapium, Zea, Virola, and Hypericum, plus a small psilate tricolporate grain (SOM Figure 1, available online). The negative extreme of Axis 1 was characterized by mesic forest types, for example, Psychotria, Schefflera (ex. Didymopanax), Struthanthus, and Malanea. Axis 2 subdivided forest types with Meliaceae, Gunnera, Clethra, and Lamiaceae at the negative extreme, and Trema, Clusia, Struthanthus, and Dodonea at the positive extreme.
Local pollen zones POMA-1 and POMA-3 were forest-rich samples that occupied similar ordination space with slightly positive scores on Axis 1 (Figure 3), while local pollen zones POMA-2 and POMA-4 were both grassland dominated, lying at the positive extreme of the Axis. POMA-4 was subdivided into samples with very high values of Poaceae and Zea pollen (zone 4a), from those that were still rich in grass, but had more moderate values and hence scores that lay closer to the origin of Axis 1.
Fossil pollen and charcoal zone descriptions
POMA-1. 296–220 cm (c. 3530–3000 cal. yr BP)
Laminated gyttja olive gray to dark gray with fine very dark gray bands (Figure 2, SOM Figure 2, available online) was present in this zone. Moderate abundances of organic material (14–20%) occurred during this period, while carbonate abundance remained low (<3%). Pollen concentrations were relatively low ranging from 3700 to 23,000 grains/cm3 (Figure 4). The fossil pollen within these basal samples contained high abundances of Andean forest taxa, c. 80–70% of the pollen sum and included Urticaceae-Moraceae (20–30%), Acalypha (10–20%), Elaeocarpaceae (most likely Vallea; 5%), Hedosymum (10–20%), and Melastomataceae (5–10%). Poaceae pollen was typically 20–25% of the sum. The largest peaks of charcoal within the core (1.88–2.45 mm2/cm3) were found within this zone, though the majority of samples contained no charcoal. Zea mays pollen was first detected in this record at c. 3290 cal. yr BP.
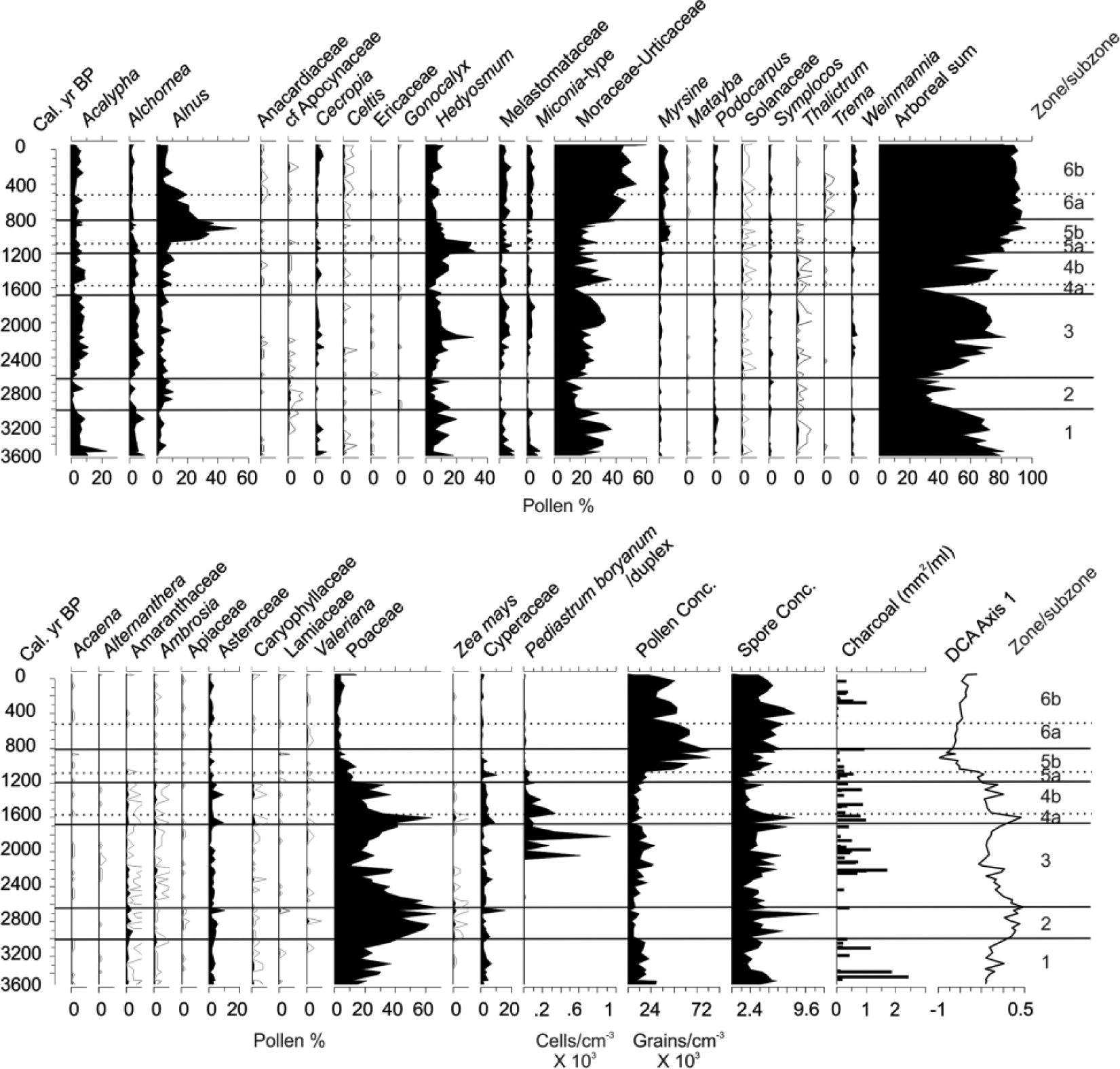
Percentage plot of the most abundant palynomorphs and total pollen concentration for samples from Lake Pomacochas, Peru. Also shown is the first Axis scores of the detrended correspondence analysis plotted against time and the charcoal concentrations for the 60- to 180-µm and >180-µm size fractions.
POMA-2. 210–170 cm (c. 2980–2560 cal. yr BP)
Laminated gyttja with relatively low pollen concentrations ranging between 2000 and 5000 grains/cm3 was present in this zone. This zone is marked by a sudden increase in Poaceae abundance rising to c. 70%. Zea mays pollen occurs consistently throughout the zone. Arboreal pollen occurrence falls to c. 30–40% and charcoal abundance falls to zero in most samples.
POMA-3. 167–95 cm (c. 2520–1710 cal. yr BP)
Laminated sediments were interrupted by a 1-cm-thick sandy layer at 144- to 145-cm depth (c. 2250 cal. yr BP). A reduction in Poaceae and increase in montane forest pollen types characterized this zone. Montane forest pollen increased throughout this zone reaching a peak of 80%. At 2110 cal. yr BP, Poaceae pollen reached its nadir (11%) and there was a peak of charcoal. Poaceae pollen gradually increased in abundance reaching c. 40% by the end of the zone. Zea was present in the lower part of the zone but was not recorded between 2160 and 1820 cal. yr BP. Charcoal was rare in the bottom of this zone but increased in abundance post-2300 cal. yr BP. Substantial blooms occurred of the green algal species, Pediastrum boryanus, twice exceeding 200% of the pollen sum.
POMA-4. 90–50 cm (c. 1610–1555 (subzone a), c. 1500–1180 (subzone b) cal. yr BP)
The laminated sediments were generally similar to those of preceding zones. Pollen concentrations were generally low with c. 9000–15,000 grains/cm3 in most samples. A sharp increase in Poaceae and Zea abundance marked subzone POMA-4a with a brief spike to >60% and 2.3%, respectively, corresponding to a decrease in forest pollen abundance to 26%. The last record of Zea pollen prior to European contact was at 1200 cal. yr BP. Charcoal was consistently present in these samples, but at lower concentrations than in the previous zone.
POMA-5. 48–32.5 (c. 1200–1110 (subzone a), c. 1040–810 (subzone b) cal. yr BP)
This zone is evident in a change in stratigraphy within the core beginning with an increase in organic content from 15% in previous zones to 20% at 48 cm and then 28% at 46 cm. Carbonate concentrations also rise through this zone reaching almost 10% of dry mass. At 46 cm, the change to a black gyttja is readily observable, and the organic content continued to increase until reaching c. 60% in zone 5b. Pollen concentrations increased as organic content rose increasing to >80,000 grains/cm3. Poaceae pollen declines to c. 10–13%, as forest pollen increases to >80%. Zea pollen was absent from this zone. The dominant pollen type in the spectra is the small tree Hedyosmum, which between 1130 and 950 cal. yr BP attained >30% of the pollen sum. Subzone POMA-5b was marked by a transition from Hedyosmum to Alnus. Alnus reaches 51% of the pollen sum at c. 840 cal. yr BP, before beginning a steady decline. Other forest types also increase including: Myrsine, Psychotria, Vallea, and forest pollen reaches its maximum within the record of 95%. Charcoal was present at low abundances throughout the zone, but exhibited a peak in the uppermost sample dated to c. 810 cal. yr BP.
POMA-6. 30–0 cm (c. 710–460 (subzone a), c. 400–0 (subzone b) cal. yr BP)
Organic content was generally 30–40% and carbonate content fell to c. 2% at 600 cal. yr BP before rising to c. 10% in subzone 6b. Sediments comprised dark gray gyttja, except for a section from 22 to 14 cm (c. 500–310 cal. yr BP) of laminated gyttja. Urticaceae/Moraceae, Myrsine, Weinmannia, Podocarpus, and Elaeocarpaceae increase in importance as Alnus declined to about 5%. Total forest pollen percentage stabilized at c. 80%. Charcoal was either minimal or absent apart from an event at 500 cal. yr BP. Zea was largely absent during this zone, only identified at c. 460 and 200 cal. yr BP. Pediastrum was present at very low levels.
Discussion
Very few natural lakes lie on the eastern flank of the Central Andes between elevations of 300 and 3100 m. The steep slopes and the lack of lake-forming processes, that is, below the limit of glaciation and above the elevation of river meanders, cause this rarity. Lying at 2150-m elevation, Lake Pomacochas is a large, deep, solution basin, with small inflowing and outflowing streams that offers rare insights into the ecological history of Andean mid-elevation forests. The finely laminated sediment, from beneath 78 m of water is consistent with a relatively productive lake with anoxic bottom waters.
Pomacochas in a regional climate context
The Pomacochas record comes from a habitat that is ostensibly stable. At 2150-m elevation, the lake lies in the heart of the sub-Andean forest far from ecotonal boundaries with grassland. Yet, this 3530-year record encompasses large shifts in the pollen record and apparent fire frequency. We compare the paleoecological record from Pomacochas with isotopic data located c. 67 km from Pomacochas from Palestina Cave (Apaéstegui et al., 2014). Lying at 870-m elevation, in a relatively wet environment, this low elevation cave record offers a valuable proxy for precipitation variability at Pomacochas. As the Palestina Cave record spans just the last 1600 years, a second cave record from Huagapo Cave (Kanner et al., 2013) is also presented. Huagapo Cave offers a highly resolved speleothem record that spans the last 3500 years, but lies above 3700-m elevation in a much drier Andean ecosystem about 800 km to the south of Pomacochas. From these data sources, we can infer an approximate precipitational history, although strong local variation is evident between the proxy records.
The millennial scale trajectory of climate change of the Holocene in the Andes was set by changes in precession of Earth’s orbit around the Sun (Bush et al., 2011a). The nadir of Dec–Feb (wet season) insolation at c. 10,000 cal. yr BP resulted in falling lake levels in the southern tropical Andes (Abbott et al., 2003; Fornace et al., 2014; Hillyer et al., 2009). Lakes were often below modern levels until c. 4000 cal. yr BP in much of the region (Baker et al., 2005), but there was an overall trend of increasing moisture availability from c. 6000 until c. 2500 cal. yr BP (Kanner et al., 2013). The precessional pattern of climate change is reflected in our record by increases and decreases in the proportion of pollen from forest taxa (Figure 5).
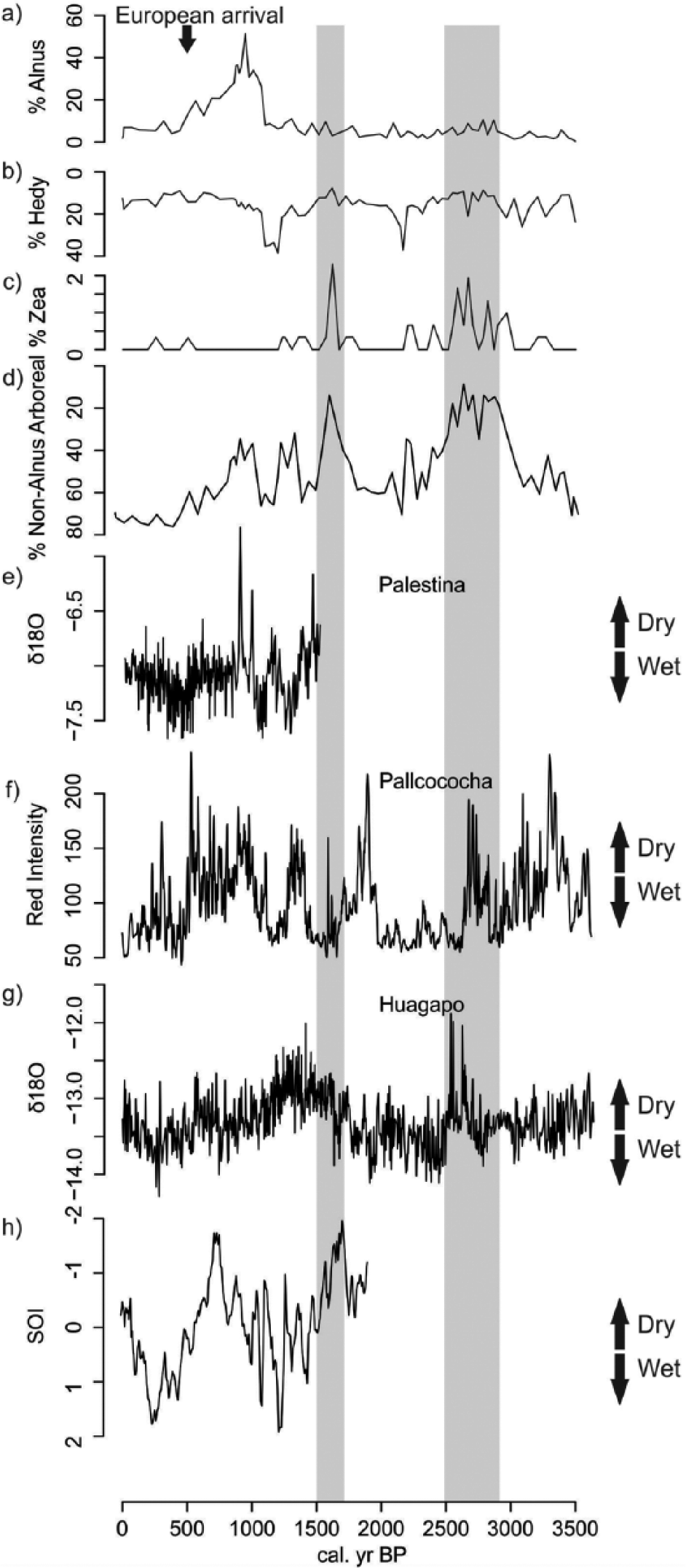
Changes in Pomacochas percentage pollen abundance relative to independent climate proxy records. (a) % Alnus pollen, (b) % Hedyosmum pollen, (c) % Zea pollen, (d) % Arboreal pollen excluding Alnus, (e) δO18 Palestina Cave calcite data (Apaéstegui et al., 2014), (f) Pallcacocha sediment red-coloration intensity data (Moy et al., 2002), (g) δO18 Huagapo Cave calcite data, (h) Reconstructed Southern Oscillation Index (Yan et al., 2011). Note that the scales for (b) and (d) are inverted to provide directional consistency with the independent climate records of wetter and drier conditions.
Between c. 2500 and c. 1450 cal. yr BP (Kanner et al., 2013), the Huagapo Cave record suggested a long-term transition to drier conditions. During this time, forest cover was reduced as grasses and herbaceous elements increased in importance (Figures 4 and 5). Times of elevated Poaceae pollen abundance coincided with dry events inferred from the Huagapo Cave record. This lake setting is so wet today that it is very unlikely that the drying would have caused forest to be replaced by grassland. A more parsimonious explanation may be that there was a change in pollen source area. The dry signature was interpreted to indicate a weakening of the South American Summer Monsoon (Kanner et al., 2013), which, in turn, lessened upslope winds emanating from the Amazon. The local effect of reduced convective rains and a reduction in easterly winds would have been to transport less forest pollen into the Pomacochas Basin. During these grass pollen–rich events, pollen concentrations fell, suggesting that the source area of the pollen was considerably reduced. Thus the increase in Poaceae abundance does not translate solely into a much more open landscape, but was also a change in the source area of the pollen.
Palestina Cave recorded a marked oscillation of wet and dry events, each lasting about 200 years between c. 1600 and 900 cal. yr BP (Figure 5), culminating in a very strong drought signal at c. 910 cal. yr BP. These events were captured in the Pomacochas pollen record with wet events resulting in an expansion of forest cover, while dry events resulted in higher Poaceae values. After c. 1200 cal. yr BP, a wet oscillation resulted in the expansion of a small tree, Hedyosmum, while a dry oscillation at c. 1000 cal. yr BP did not result in grassland expansion, but in the expansion of Alnus, a pioneer tree capable of colonizing disturbed settings. Between c. 545 and 360 cal. yr BP, the Palestina isotopic record suggested a wet setting, with a subsequent drying trend (Apaéstegui et al., 2014). During this time, pollen from a diversity of forest types reached c. 80% of the pollen sum, the highest forest representation of the record (Figure 5).
Even though the tropical Atlantic was probably the immediate driver of most western Amazonian climate change (Kanner et al., 2013) then as now, extreme ENSO events also induced major floods and droughts (Marengo et al., 2011). The Palestina data appeared to track the inferred ENSO signature of the Pallcacocha record (Moy et al., 2002) and the Southern Oscillation Index (Yan et al., 2011). Strong negative Southern Oscillation Index events (El Niño) corresponded to dry events suggesting that the flow of moisture coming across this section of the Amazon was modified by sea-surface temperature anomalies in the Pacific Ocean (Figure 5). Both the Pallcacocha and the Southern Oscillation Index revealed a strong El Niño influence between c. 1050 and 590 cal. yr BP, which corresponded to the relatively dry event seen in the Palestina Cave record, but contrastingly a wetter period at Huagapo, emphasizing that this more remote record may not reflect local conditions at Pomacochas.
The signature of fire and maize
As Pomacochas lies within the Andean volcanic gap (Räsänen et al., 1992), the source of ignition in the forests is either lightning or human activity. Lightning-initiated fires are liable to burn themselves out within a few meters (Alencar et al., 2004). That these wet forests on the flank of the Andes do not burn naturally makes sedimentary charcoal a sure sign of the presence of people (Bush et al., 2011b; Urrego et al., 2010). Burning such wet forests, however, is itself challenging, and breaking the initial forest cover is most likely to be successful during exceptionally dry years. At Lake Pomacochas, the largest charcoal peak of the entire record is at c. 3360 cal. yr BP, and occurs within a period of heightened El Niño activity (Moy et al., 2002). At the onset of the record, the proportion of Poaceae (15–20%) is higher than the c. 5% that would be expected in a closed canopy lowland forest (Bush, 2002). Grassy marshes have the capacity to increase Poaceae pollen counts, even when the adjacent forest is intact (Bush and Colinvaux, 1990). The values seen in the base of the Pomacochas record lie with the range of values (2–70%, mean 15%) found in moderately disturbed montane forests in the Andes (Bush et al., 2015). We infer that the elevated percentages of Poaceae represent either small openings in the forest or a grassy marsh adjacent to the lake.
The large charcoal peak at c. 3360 cal. yr BP was followed by an increase in Poaceae abundance, a decline in all forest taxa and the appearance of pollen of Zea mays at c. 3200 cal. yr BP. Cumulatively, these results indicated the expansion, perhaps onset, of maize cultivation close to the shores of the lake. Several points need to be made regarding this finding. The first is that humans were clearly occupying these mid-elevation cloud forests at a time when overall population densities were low. This observation contradicts assertions that these were uninhabited or uninhabitable locations. The second is that while maize at this time may have been of culinary importance, it may also have been grown for brewing Chicha, a Pre-Columbian beer, or ceremonial purposes (Nicholson, 1960). Regardless of its use, maize is an unequivocal indicator of human presence. Another large fire event was marked at c. 3000 cal. yr BP, and this presaged an even larger increase in Poaceae pollen at the expense of forest taxa; an event that led to the statistical separation of POMA-1 from POMA-2. For the next 600 years, there was near-constant evidence of maize agriculture, but no charcoal was recorded in the sediments.
This pattern of an initial opening of the landscape, with a peak of charcoal, followed by evidence of agriculture but no correlation with charcoal presence has been described repeatedly (Bush et al., 1992; Piperno et al., 1990). If cultivated land was being used repeatedly, such that when fires were set they burned grasses and herbaceous regrowth, little charcoal would have been produced. An alternative hypothesis is that people were deliberately suppressing fires near to their homes fringing the lake.
The cultivation of maize became more erratic after c. 2560 cal. yr BP. Relatively long gaps, lasting several centuries, occurred between samples that revealed Zea pollen. It is possible that Zea cultivation was abandoned during these phases or that it was located further from the lake. The reduction in Zea corresponded to increased abundances of Pediastrum, an alga that generally thrives in settings with moderately high values of pH, dissolved organic carbon, and nutrient availability (Weckström et al., 2010). The peaks of Pediastrum would be consistent with continuing human landuse in the catchment, even though Zea was not detected.
Although lakes may appear to be ideal sites for settlement and are thought to have attracted colonists in both the lowlands and the high Andes, the middle elevations around Pomacochas would have been especially problematic for agriculturalists. Wet, cool, and shrouded in cloud for much of the time, the Lake Pomacochas basin may have been only marginal farmland. One benefit of growing crops is the ability to store food, but in these wet conditions drying and storing grain may have been challenging, with substantial or catastrophic loss to mold during wet periods. The period when maize agriculture appears to have been at its peak, between c. 3300 and 2560 cal. yr BP, was one of the consistently driest times of the last 3400 years.
Between 2640 and 2280 cal. yr BP, Poaceae pollen abundance declined by a factor of three and forest recovery was evident. Hedyosmum attained a brief peak of 20%, suggesting the recovery of a wet forest (Figures 4 and 5). This pattern was interrupted by a fire event at c. 2180 cal. yr BP that was quickly followed by the influx of a 1-cm sandy lens of sediment and renewed expansion of Poaceae abundance. The timing of this event coincided with the onset of a period enhanced ENSO activity in the eastern equatorial Pacific (Conroy et al., 2008). The sandy layer at Pomacochas probably represented a significant episode of forest clearance and ensuing erosion, and this may have marked the time when the most extensive deforestation took place. The peaks of Pediastrum between c. 2030 and 1560 cal. yr BP suggest increasing productivity within the lake and would be consistent with erosion increasing nutrient availability.
Despite the evidence of human activity based on charcoal abundance and loss of forest cover, maize pollen was not found between 2150 and 1760 cal. yr BP. This absence of maize suggested that an alternative crop was being grown. We hypothesize that from c. 2150 until c. 1760 cal. yr BP and post-1200 cal. yr BP were times when this setting was so wet that people switched to other crops or in the post-1200 cal. yr BP period partially abandoned the site. The site was too low for quinoa, but potatoes, other solanaceous domesticates, squash, and beans may have been cultivated.
After the peak of Poaceae at 1600 cal. yr BP, which coincided with the highest abundance of maize pollen in the record, the forest cover expanded at the expense of grassland. The last maize of pre-Columbian times was recorded at c. 1200 cal. yr BP.
Reforestation and the emergence of Chachapoya culture
A transition in the record took place at c. 1200 cal. yr BP that was marked by the deposition of increasingly organic sediments and the recovery of forest taxa as climates became wetter (Figure 5). Hedyosmum again was the first tree to regain dominance in this wet period, reaching a local peak of abundance between c. 1170 and 1070 cal. yr BP. The increased carbon and carbonate contents in the last 1200 years suggest a substantial change in basin limnology. The reason for this change is not completely understood, but it apparently coincided with changes in landuse, and would have been consistent with co-precipitation of dissolved organic material and carbonate as a product of algal blooms (Reddy, 1995). Unfortunately, diatoms, which could have shed light on past lake conditions, were not preserved in this record. When dry events occurred post-1200 cal. yr BP, woody taxa, for example Alnus, rather than grasses, filled disturbed areas.
The change in landuse aligned to larger societal changes, and we observe that it was broadly coincident with the establishment of the Chachapoya culture c. 1150–1050 cal. yr BP (i.e. 800–900 CE; Church and Von Hagen, 2008). At approximately the same time, Wari imperial presence was spreading northward into the Cajamarca region, west of the Marañon River canyon, where it lasted until c. 950 cal. yr BP (Isbell, 2008). Several Wari artifacts have been recovered in the Chachapoyas region, and Wari influence is detected in some technological and style attributes observed in Chachapoyas textiles. However, no evidence of direct interaction between Wari and Chachapoya populations has been suggested. At Lake Pomacochas, the period of Chachapoya fluorescence was during the Hedyosmum and Alnus peaks of zone 5. The presence of charcoal throughout this zone indicates that people still occupied this site. About 1040 cal. yr BP, Alnus begins a steady decline in abundance as Moraceae and other forest elements increase. Had Alnus been promoted, directly or indirectly, its decline at this site marks a change in landuse. Clearly, antecedent cultures that used maize existed within this landscape for several thousand years before the time of the Chachapoya. While we still can say little about how the Chachapoya lived, it is apparent from this record that, at Pomacochas, they were not a maize-based society. Indeed, the timing of the regional inception of Chachapoya culture is coincident with the abandonment of maize agriculture at this site. A speculative explanation might be that the growing connectivity of communities allowed more local specialization, and that other crops better suited to this elevation than maize may have been grown and traded.
Alnus remains above 20% of the pollen sum from c. 1040 until c. 650 cal. yr BP. Both Hedyosmum and Alnus are capable of growing as early successional taxa, but it is somewhat surprising to find them not being replaced by other taxa for c. 400 years. Alnus is often massively over-represented in pollen records, but even so, this pattern is strikingly similar to that seen in the high Andes where Alnus is suggested to have been planted for silviculture as early as c. 1300 cal. yr BP at Lake Huaypo, and by 900 cal. yr BP at Lakes Pacucha and Marcacocha (Mosblech et al., 2012). The explanations for the elevated presence of Alnus might be quite different. In the high Andes, it has been suggested that Alnus was needed as a supply of fast-growing timber that could be coppiced (Chepstow-Lusty and Winfield, 2000). Whether timber was ever in such short supply at the elevation of Pomacochas as in the high Andes is doubtful, but Alnus may have been planted as a windbreak, for erosional control, or for firewood in much the same way that Eucalyptus is planted today. What is noticeable, however, is that the isotopic records indicate that Alnus thrived during a dry event. Alnus is a good pioneer species in dry settings (Weng et al., 2004a). Hence, while the Alnus abundance could reflect direct or indirect effects of human landscape modification, it probably also reflected a climatic shift to drier conditions. As that dry event faded so, Alnus was replaced by other pioneer taxa that favored wetter conditions, for example, Cecropia (Bush, 2000), and a more diverse set of woody taxa, for example, Myrsine, Weinmannia, Podocarpus, and Elaeocarpaceae. The return of charcoal to the record, after very low concentrations or a total absence lasting 500 years, at c. 310 cal. yr BP could coincide with the arrival of Europeans some time after c. 400 cal. yr BP (1550 CE).
Conclusion
At the onset of this record, the landscape at Pomacochas was already being used by people. This datum reinforces archaeological evidence that the forests of the Andean flank were used by people even when human populations were still relatively sparse. The Pomacochas record includes phases of grassland expansion and maize agriculture, alternating with forest regrowth. It is apparent that a simple trajectory of increasing human domination of the landscape does not fit the observed data. Neither does a simple analysis of wet versus dry times match the observed pattern of fire or agriculture. It appears that human activity induced more open landscapes, hence higher Poaceae and lower forest pollen representation during dry times, and that maize agriculture was practiced during these dry events. After c. 1200 cal. yr BP, however, maize agriculture was decoupled from climate change, as, presumably, a cultural change led to altered landuse. We find evidence of long-term but changing landuse patterns that appear to respond to both climate change and large-scale cultural changes in the Andes, that is, centennial scale changes in moisture abundance, the development of Chachapoya culture, and the invasion by the Europeans. We conclude that at Pomacochas, landuse and crop choice of Pre-Columbian societies were influenced by contemporary climate conditions, but concurrent changes in the geopolitical landscape and regional demography, or other social factors likewise may have governed such choices.
Footnotes
Acknowledgements
We thank the government and people of Peru for their hospitality. Bryan Valencia and Angela Rozas-Davila are thanked for their hard work in the field. Crystal McMichael prepared the figures and assisted with data analyses. William Gosling and Jo Williams are thanked for their insightful comments on the manuscript.
Funding
MBB thanks the National Geographic Society for funding fieldwork. NAM thanks NSF for support via the Graduate Research Fellowship Program.