
Abstract
Most rich-fen bryophyte species are currently declining and red listed in many European countries. Little is known about their past and current occurrence. We investigated past and recent bryophyte assemblages in mires in the land uplift coast of Finland. The study areas included primary succession areas where mire development started c. 6000 BP. In total, seven mires were studied in two successional transects along the land uplift coast. The past occurrences of rich-fen and other bryophytes were studied in 45 mire stratigraphic peat profiles, and recent occurrences were collected from 158 vegetation plots of 10 m × 10 m. Bryophyte subfossils indicated that the central parts of the basins developed first to wet rich fens with groundwater influence, which developed towards intermediate fens or poorer mire types. Most mire bryophytes occurred in about the same proportion in peat profiles as in recent vegetation. Five rich-fen species (Hamatocaulis lapponicus, H. vernicosus, Meesia longiseta, M. triquetra and Pseudocalliergon trifarium) were clearly more common in peat profiles than in the present mire vegetation, where these species were very rare and appeared only in small patches without sporophytes. These rich-fen species have had markedly wider distributions in the past, probably linked to climate fluctuations, whereas current occurrences are becoming scarce because of multiple anthropogenic factors such mire drainage and climate change.
Keywords
Introduction
The survival of rich-fen bryophyte species has been a long standing research problem in biological conservation. Most investigations attribute scarcity in rich-fen species to recent environmental changes such as atmospheric acidification (Kooijman, 2012) or drainage (Mälson et al., 2008). The continuous decline of the rich-fen mosses in the remaining rich fens has also been attributed to habitat fragmentation, eutrophication, occasional draughts and competition of Sphagnum mosses (Kooijman et al., 1994). However, only few studies have examined long-term temporal dynamics of rich-fen species in the entire course of mire succession (but see Hájková et al., 2012, 2015). This is a drawback since mire bryophyte species along with other organisms are known to react complex interactions between climate, peat accumulation and overall vegetation changes (Bragazza, 2006; Lamentowicz et al., 2013). We find that successful conservation and habitat restoration of mire sites to maintain rich-fen species requires better understanding of long-term dynamics of species: this is also crucial to forecast the future of rich-fen species under on-going climatic and land-use changes.
Moss subfossils in peat can be used accurately to determine what kind of vegetation there has been at different stages. In autochtonic peat layers, moss subfossils are very abundant, but they represent only the vegetation on that single spot. Therefore, a large number of survey points is needed to reflect the vegetation of larger peatland areas. The difficulty here is, above all, that part or sometimes all of the mosses decay beyond recognition, and different moss species decompose at different rates. The overall preservation can be used to validate the reliability in assessing species composition of past communities (Janssens, 1983a). Generally, the decay rate of peat moss layers depends on the speed at which they reach the constantly anoxic layers in peat (Clymo, 1983). Sphagnum species are known to be more resistant than many other mosses (Clymo and Hayward, 1984), while liverworts degrade very quickly (Janssens, 1983a) and have very few remnants in peat layers.
The analysis of moss residues in studying mire stratigraphic history is widely used to determine the development history of specific mire basins (Aartolahti, 1965; Huttunen, 1990; Ruuhijärvi, 1963; Tolonen, 1967a). A typical sequence starts from lush, thin-peated swamps with strong influence of mineral soil and flowing water and develops to thick-peated bogs or aapa mires. In these cases, the change in moss species composition reflects the process of mire development: typically loss of swamp or rich-fen species and increase of barren bog species, yet there is evidence that in some cases, increasing water flow may favour development of fen vegetation (Hughes and Dumayne-Peaty, 2002; Tolonen and Seppä, 1994). Major climate changes are reflected in the vegetation changes of raised bogs in particular, but it is also known that the changes are sensitive to local variation in hydrological and topographical factors (Aartolahti, 1965; Huttunen and Tolonen, 2006; Tolonen, 1967a).
Stratigraphical analysis is a useful method to assess the previous distribution and abundance of the species (Turetsky et al., 2012). Studies in Central Europe and the British Islands have showed that the contemporary occurrences of many moss species are small and scattered compared with the earlier more extensive and more uniform areas (Dickson, 1973). The current populations of rich-fen mosses have been interpreted to be remnants of the former, large and uniform growth area (Dickson, 1973). Later on, their occurrences have become scarcer which has been often associated with the development of Sphagnum-dominated bogs. According to Dickson (1973), the share of the northern-continental rich-fen moss-flora is inversely proportional to the share of Sphagnum. Obviously, a strong invasion of Sphagnum mosses took place about 5000 BP because of both autogenic succession and climate change. More recently, drainage for agriculture and forestry have destroyed most of the remaining populations. The situations of the species of spring-fed rich fens on the British Islands are typical examples, for example, Calliergon richardsonii, Helodium blandowii, Meesia longiseta (nowadays extinct from the British Islands, but there are subfossil or historical occurrences; Hall, 1978), Meesia triquetra (one recent occurrence in Ireland, now probably extinct; Smith, 2004), Hamatocaulis vernicosus, Paludella squarrosa, and Pseudocalliergon trifarium (only few isolated occurrences left; Smith, 2004).
Mires along the land uplift coast of northern Baltic Sea with continuous primary mire formation (Huttunen and Tolonen, 2006) create a successive series of mires. These mires provide a unique opportunity to explore the long-term dynamics of peatland moss species. By examining the whole chronosequence, it is possible to obtain same-time occurrence data from mires which differ in their age and successional stage. In the land uplift area, there is a continuous gradient from primary mire formations to central parts of aapa mires and from paludified forests to Sphagnum-dominated peripheral parts (Rehell et al., 2012b). These mires have been extensively drained for forestry and agriculture, which has led to fragmentation and areal shrinkage (Rehell and Heikkilä, 2009). Particularly, rich fens and their typical species have become threatened even in this area.
The aim of this study is to investigate the long-term trends of bryophyte species composition in mire vegetation by comparing their occurrences in peat profiles and present vegetation. The shifts in rich-fen bryophytes are likely linked to natural climate variations and recently also to direct anthropogenic land-use changes such as drainage.
Material and methods
The study was conducted on the land uplift coast of Bothnian Bay where uniform succession series from the seashore to the inland area occurs. In this area, aapa mires dominate (Ruuhijärvi, 1960). The age of the primary paludification of the basins was estimated using the regression diagrams for the region (Taipale and Saarnisto, 1991). The ages refer to C-14 years calibrated to calendar years (Table 1). The studied basins constitute two succession series on two different types of topography: one in an area with moraine topography and the other in an area with sandy topography (Figure 1).
Transects with elevations referring to main sites of primary mire formation. Areas 1–5 are on moraine topography, areas 6–7 on sandy topography. The ages of localities from regression curves (Taipale and Saarnisto, 1991) are expressed as calibrated years before present (AD 1950, Weninger et al., 2005).

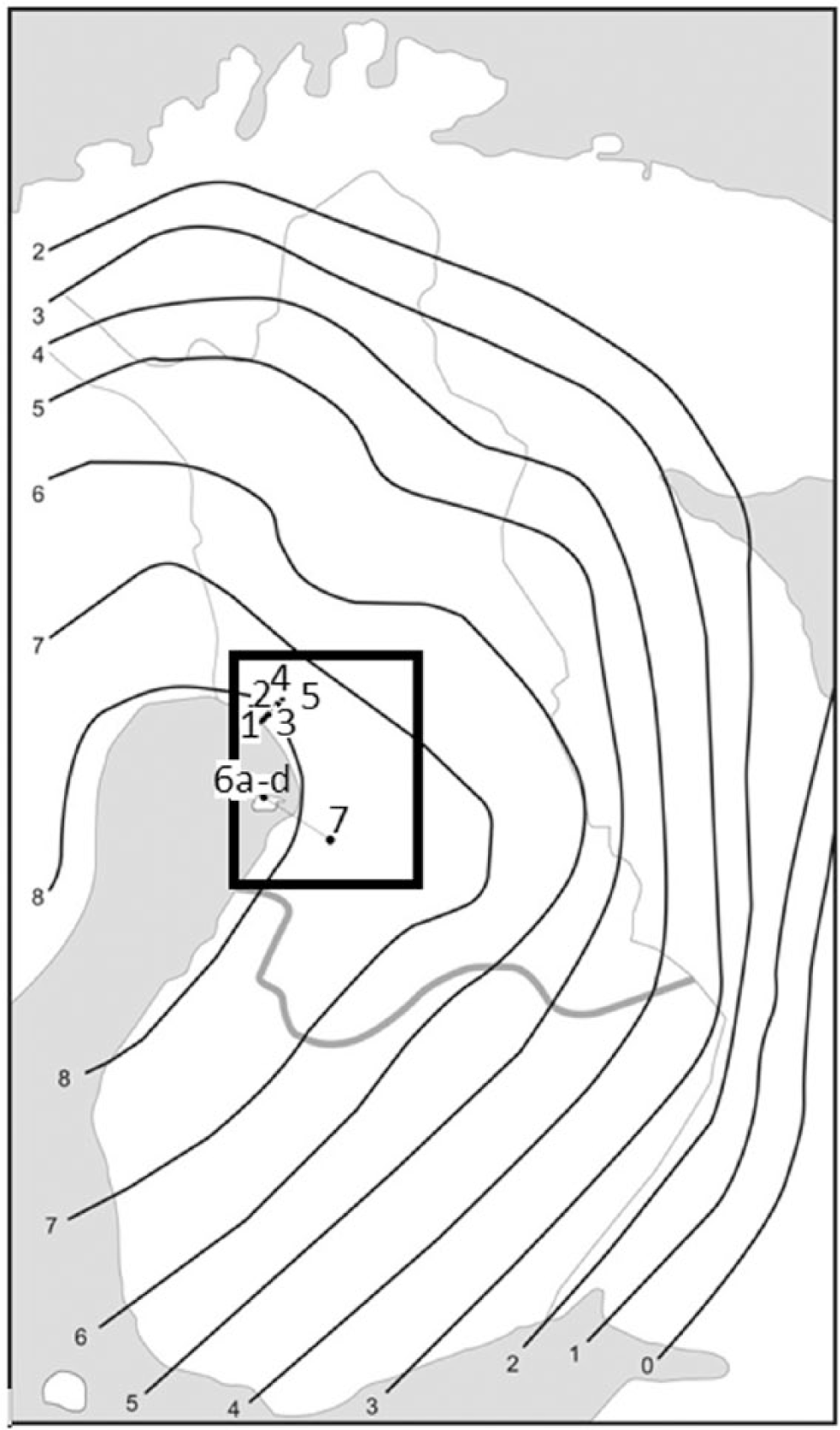
Studied mire basins along the succession series on the land uplift coast of the Bothnian Bay. The black contours show the isobases of land uplift and the grey line the border between the southern ‘bog-area’ and the northern ‘aapa-mire area’ in Finland. The rectangle shows the larger reference area used in Table 2 (western part of Northern Ostrobothnia).
The studied succession series with moraine topography is situated on the border between communes Simo and Kuivaniemi. Here, there is a uniform series of mire basins from the present seashore up to large aapa mires, which have started their development more than 6000 years ago (Table 1). The bedrock is composed mainly of acidic granitoid gneiss and migmatite with diabase joints and other more basic veins on the whole area (Geological Survey of Finland, 1997). It is overlain by layer of bottom moraine, which forms small drumlins in places. Along the succession series, study sites 1–5 in Figure 1 represent paludified basins bordered by mineral soil on different elevations above sea level. Each study site includes an entire catchment area with mire on the central part. The studied transects are placed across the whole basin so that they represent the whole paludified area with both central and peripheral parts (Laitinen et al., 2007).
The studied succession series in the area with sandy topography is situated on a large esker formation, which stretches from the island of Hailuoto towards southeast. Here, young mire basins near the seashore can be compared with aapa mires having started their development more than 4000 years ago (Table 1). This esker chain is composed mainly of fine sand with only little gravel and boulders. The coastal waves and wind have accumulated beach ridges and dunes stretching kilometres on both sides of the original core of the esker. The mires are situated on narrow basins between these ridges. The bedrock consists mainly of sedimentary siltstone (Geological Survey of Finland, 1997), which is covered by thick layers of sand, till and other loose sediments (Gibbard, 1979). The study sites along transects were selected following the same criteria as in the northern transect on moraine topography.
A thorough vegetation mapping and survey of rich-fen bryophytes was done for the study sites in 1998–2002 (Rehell and Heikkilä, 2009). Vegetation sample plots were studied on 10 transects drawn across the basins (Table 1). Vegetation studies were done on mires and paludified heaths of the large sample plots. On the littoral zone also seashore meadows and thickets were studied. Neither stable heath vegetation nor permanent water bodies were investigated. A total of 158 large vegetation sample plots were placed along longitudinal transects across successional mire basins so that both the parts of the mire basin near the water divides and the parts with a voluminous water flow became represented. As a rule, their area was 100 m2. For 17 sample plots distributed in mire basins all over the successional series, a larger sample plot size (200–600 m2) was used, because the set was originally made for a study of the initiation and formation of microtopographic levels along the aapa mire succession (Rehell et al., 2012a). The corners of sample plots were placed at intervals of 100 m within transects, the sides of the plots directing from the east to the west and from the south to the north. In the central part of the aapa mire complex in Nikkilänaapa-area (no. 4 in Figure 1), only every other plot was studied, because of the wetness of the large flark fen. When summing up the mean cover values of species, this was taken into account by multiplying the values of these plots by 2. Where transects crossed mineral soil islands or a small water body, one sample plot on both sides of the island or a pond was placed. In each large sample plot, one small sample plot of 1 m2 for each of uniform-looking plant community (microtopographic level) was studied. A total of 468 1-m2 plots were investigated. The percentage cover of species was estimated for 1-m2 plots. The field work on transects with morainic topography was carried out in July–September 2001 and 2002 and on sandy topography in July–September 2003.
The cover values for the large plots were calculated as averages from the values of 1-m2 plots, weighted in relation on the proportions of the microtopographic levels on the large plots. A list of species for all 100-m2 plots was made. For those species which did not occur on the 1-m2 plots but were found elsewhere on the studied mire basins (Hamatocaulis lapponicus, Meesia longiseta, Pseudocalliergon trifarium), the mean coverages had to be approximated by the extent of the patches found in the mapping of the basins.
Peat profiles were taken from some of the studied vegetation sample plots on different altitudes along the two succession series (Rehell et al., 2012a). The sites represented both the central and peripheral parts (Laitinen et al., 2007) of the mire basins. On some sites, two profiles were taken close to each other to compare the development of microtopographical patterns (e.g. one profile from the string and the other from the flark next to it; Rehell et al., 2012a). The samples from the cover 50 cm were taken with a metal cylinder with the diameter of 11 cm. From the lower peat horizons, samples were taken with a Russian borer (Tolonen, 1967b) with length of 50 cm and diameter of 4 cm with two replicate horizons close to each other. The peat profiles were cut into 10-cm pieces, which were studied (5-cm pieces were used on the layer of living mosses and sometimes also on the bottom layer).
Along succession series with moraine topography, 29 peat profiles were taken and 20 of them represented primary mire formation on seashore. Altogether 374 peat samples were studied from the study area on moraine topography. From the succession series with sandy topography, 16 peat profiles were taken and 12 of them represented primary mire formation. From the sandy areas, altogether 144 peat samples were studied.
The moss remnants were studied from a piece of peat profile put on 1- and 0.1-mm sieves. The peat was washed with water to remove amorphic humus material. The often used pre-treatment with KOH was not necessary because of the very low humification rate. Leaf and stem fragments of mosses were identified with the help of binoculars. The cell characters of moss remnants were checked using optical microscope. At the same time, the residues of vascular plants were studied. The percentages of different species of the whole volume of washed peat sample (taking into account also the amorphic mass remaining after the washing) were visually assessed using a version of the quadrat and leaf count (Mauquoy et al., 2010). The studied peat samples were mainly very weakly humified (H2-3 in von Post scale). More degraded peats (H5 or above) were situated only in thin layers, mainly in the bottom of profiles representing paludification of forest. The identification of the moss remnants to the species level was usually feasible. Some taxa close to each other were treated collectively in the comparisons of the peat profiles and recent vegetation. When counting the frequencies of mosses in peat samples, only findings with concentration more than 0.5% were taken into account, because smaller occurrences typically consisted of only single, often broken leaves and were thought as more uncertain.
The characteristics and type of the original vegetation were assessed based on the moss species composition in peat layers (for ecological characteristics of species, see Eurola et al., 1994). The profiles were divided into those representing primary mire formation and those representing paludification of forests. The age of profiles was estimated based on their elevation above sea level (see above). Those profiles that lacked typical seashore layers or which had clear evidence of forests on their bottom (coal fragments, Sphagnum girgensohnii, seeds of Carex globularis) could not be dated. The coarse dating of individual peat layers in profiles representing primary mire formation was made based on an assumption that the growth rate of peat has been stable along the development of the mire. The ages of the positions in the profiles were estimated as follows: the age of the bottom layer is the same as the age of the seashore stage on that elevation (Table 1) and the age of the living mosses is 0 years. For a group of rich-fen mosses, the occurrences at different periods of time were approximated by calculating the number of samples with occurrences of those mosses in each successive period of 200 years in the timeline.
Results
The type of vegetation which constituted peat layer was determined from the composition of moss remnants found in the peat. Layers indicating wet, swampy rich fens (with species such as Calliergon richardsonii and Meesia spp.) were found just over the layers of coastal swamps (where e.g. Drepanocladus aduncus, Phragmites australis dominated). All profiles with rich fens in some layer represented primary mire formation (Figure 2). The peat layers above the rich-fen horizons included minerotrophic peats obviously indicating the development of acid aapa mires. Mostly, they were composed of Carex-peats with mosses indicating wet, flark-level conditions (e.g. Sphagnum subsecundum, Warnstorfia exannulata). Carex-peats without identifiable mosses were also found in some profiles, but only occasionally. Sphagnum-dominated peat layers indicating lawn to hummock-level vegetation (with species like Sphagnum papillosum or S. fuscum) were encountered on the surface of some profiles.
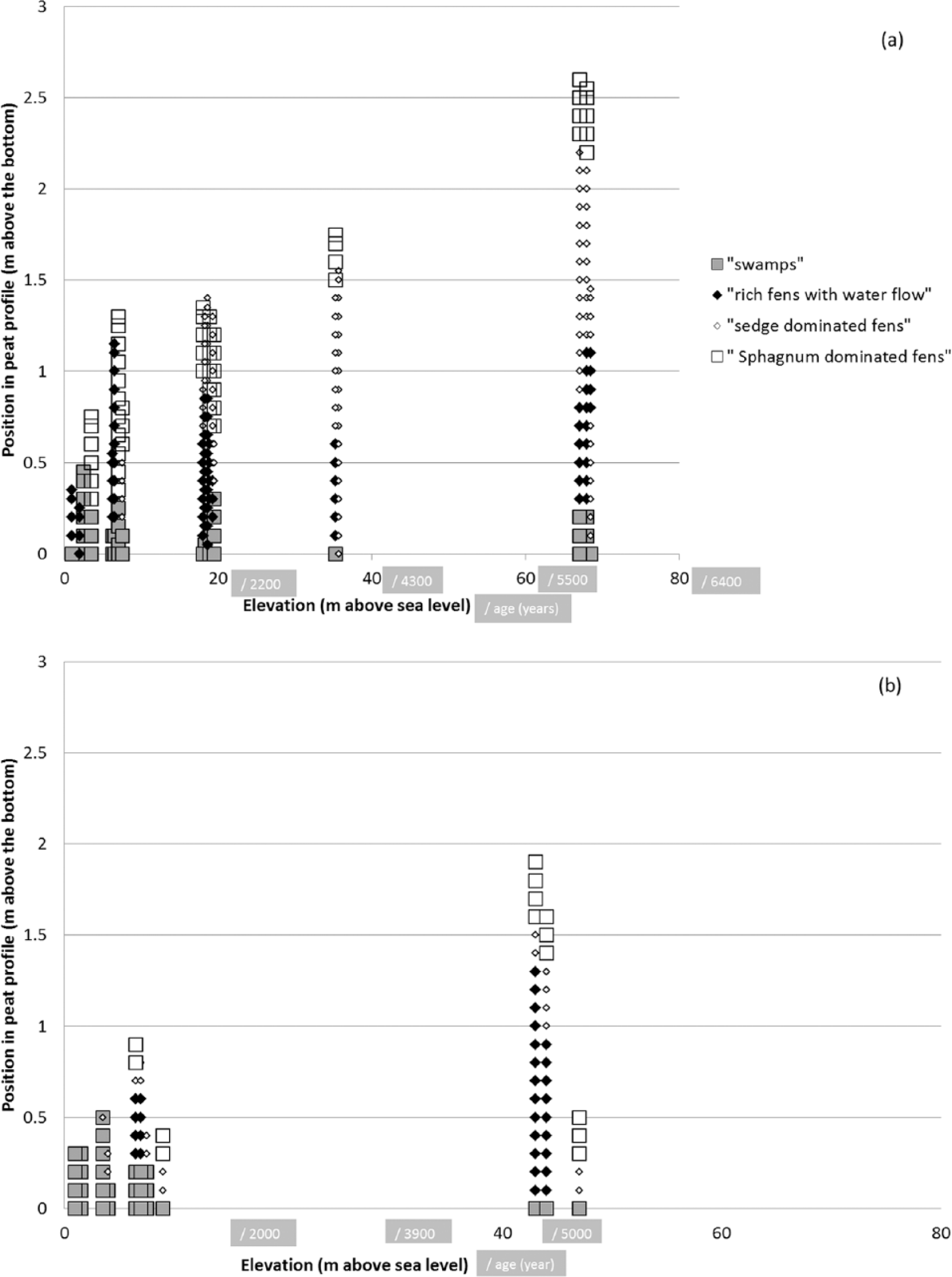
The occurrence of main mire vegetation categories in the studied peat profiles. The horizontal axis: the elevation of the studied profile from the current sea shoreline (m a.s.l.). The vertical axis: the distance of the peat sample from the bottom of the profile. (a) Moraine topography; (b) sandy topography. The young swamps and rich fens with strong water flow taken separately. The other types, even the poor, Sphagnum-dominated, include other, mainly minerotrophic types.
A comparison of bryophyte occurrences in present mire vegetation and in studied peat profiles revealed a pattern of groups of moss species (Figure 3). Most common mire mosses had mean coverage in present vegetation about the same magnitude as their mean concentration in studied peat samples (the dots near the cross-section line in Figure 3). These included many species common in either wet fens (Calliergon giganteum, Scorpidium spp., Sphagnum cuspidatum coll., S. squarrosum, S. subsecundum, S. teres, Straminergon stramineum and Warnstorfia exannulata) or bogs (Sphagnum magellanicum). Some Sphagnum mosses typical for lawn-level vegetation, especially Sphagnum recurvum coll., S. balticum and S. papillosum were situated on the upper part of the graph in Figure 3, well to the right of the cross-section line. They seemed to have large present mean cover in proportion to the concentration in peat, although their concentration in peat was not small. Some hummock-level Sphagna (S. russowii, S. fuscum) were situated close to them.
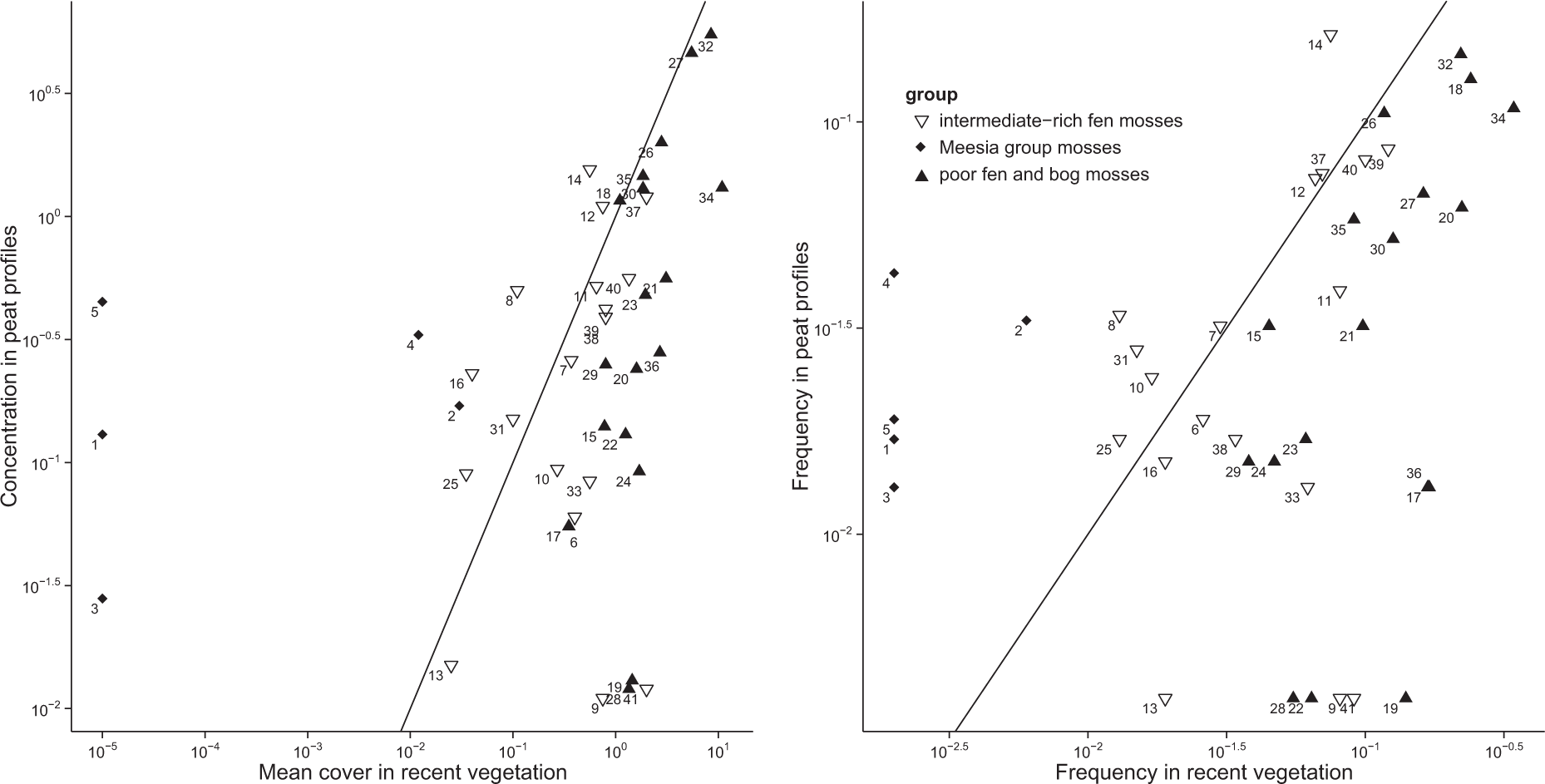
A comparison of bryophyte occurrences in present mire vegetation and in peat layers. The approach based on mean cover and concentration (left) and on frequencies (right). The graphs are based on all studied sample plots and peat profiles from both moraine and sandy topography. Diamonds (Meesia rich-fen mosses): 1 – Hamatocaulis lapponicus, 2 – Hamatocaulis vernicosus, 3 – Meesia longiseta, 4 – Meesia triquetra, 5 – Pseudocalliergon (Drepanocladus) trifarium. Open triangles (other intermediate rich-fen mosses): 6 – Calliergon cordifolium, 7 – Calliergon giganteum, 8 – Calliergon richardsonii, 9 – Campylium stellatum, 10 – Drepanocladus aduncus, 11 – Scorpidium cossonii/revolvens, 12 – Scorpidium scorpioides, 13 – Tomentypnum nitens, 14 – Warnstorfia exannulata/procera, 16 – Warnstorfia tundra, 25 – Sphagnum contortum, 31 – Sphagnum obtusum, 33 – Sphagnum platyphyllum, 37 – Sphagnum squarrosum, 39 – Sphagnum subsecundum/inundatum, 40 – Sphagnum teres, 41 – Sphagnum warnstorfii. Filled triangles (poor fen and bog mosses): 15 – Warnstorfia fluitans/pseudostraminea, 17 – Aulacomnium palustre, 18 – Straminergon stramineum, 19 – Pleurozium schreberi, 20 – Polytrichum sp. (P. commune, P. jensenii, P. schwarzii, P. strictum), 21 – Sphagnum balticum, 22 – Sphagnum capillifolium/rubellum, 23 – Sphagnum centrale/palustre, 24 – Sphagnum compactum, 26 – Sphagnum cuspidatum coll. (S. annulatum, S. cuspidatum, S. jensenii, S. majus), 27 – Sphagnum fuscum, 28 – Sphagnum girgensohnii, 29 – Sphagnum lindbergii, 30 – Sphagnum magellanicum, 32 – Sphagnum papillosum, 34 – Sphagnum recurvum coll. (S. angustifolium, S. fallax, S. flexuosum incl. S. pulchrum), 35 – Sphagnum riparium, 36 – Sphagnum russowii, 38 – Sphagnum subfulvum. The cross-section line is drawn to show approximately where the values are equal.
The species with dots on the right-hand side of the cross-section line near the bottom of the graphs included taxa whose present mean cover was relatively high while the occurrences in peat samples were infrequent (Figure 3). These species included mosses typical for very different environments: swamps and lawn-level rich fens (Calliergon cordifolium, Campylium stellatum, Sphagnum warnstorfii and Tomentypnum nitens) unstable mud-bottoms (Sphagnum compactum, S. platyphyllum), spruce mires (Polytrichum spp. and Sphagnum girgensohnii) and dry hummocks (Aulacomnium palustre, Pleurozium schreberi and Sphagnum capillifolium).
Species situated farthest to the left of the cross-section line occurred frequently in peat in relation to their scarcity in present vegetation (Figure 3). These species seemed to constitute an ecologically coherent group including five species typical for wet, groundwater-fed rich fens (Hamatocaulis lapponicus, H. vernicosus, Meesia longiseta, M. triquetra and Pseudocalliergon trifarium). Onwards, we call this group of five mosses the Meesia rich-fen moss assemblage or Meesia rich-fen assemblage, for brevity. Four additional, ecologically similar species, Calliergon richardsonii, Sphagnum contortum, S. obtusum and Warnstorfia tundrae, showed the same tendencies, but the further analyses were restricted to the five species mentioned above.
The analysis of occurrences of the Meesia rich-fen moss assemblage in peat profiles showed their relatively frequent occurrence in early successional mires (Figure 4). This contrasts with their virtual absence from present mire vegetation. Only Hamatocaulis vernicosus and Meesia triquetra occurred in small amounts in the sample plots of present mire vegetation. The analysis of relative occurrences of rich-fen species showed that they have been present in studied mires at least from c. 5500 BP to recent times (Figures 5 and 6). There is some variability, but they seem to occur especially frequently in peat profiles aged 1000–1600 and 200–600 BP (Figures 5 and 6).
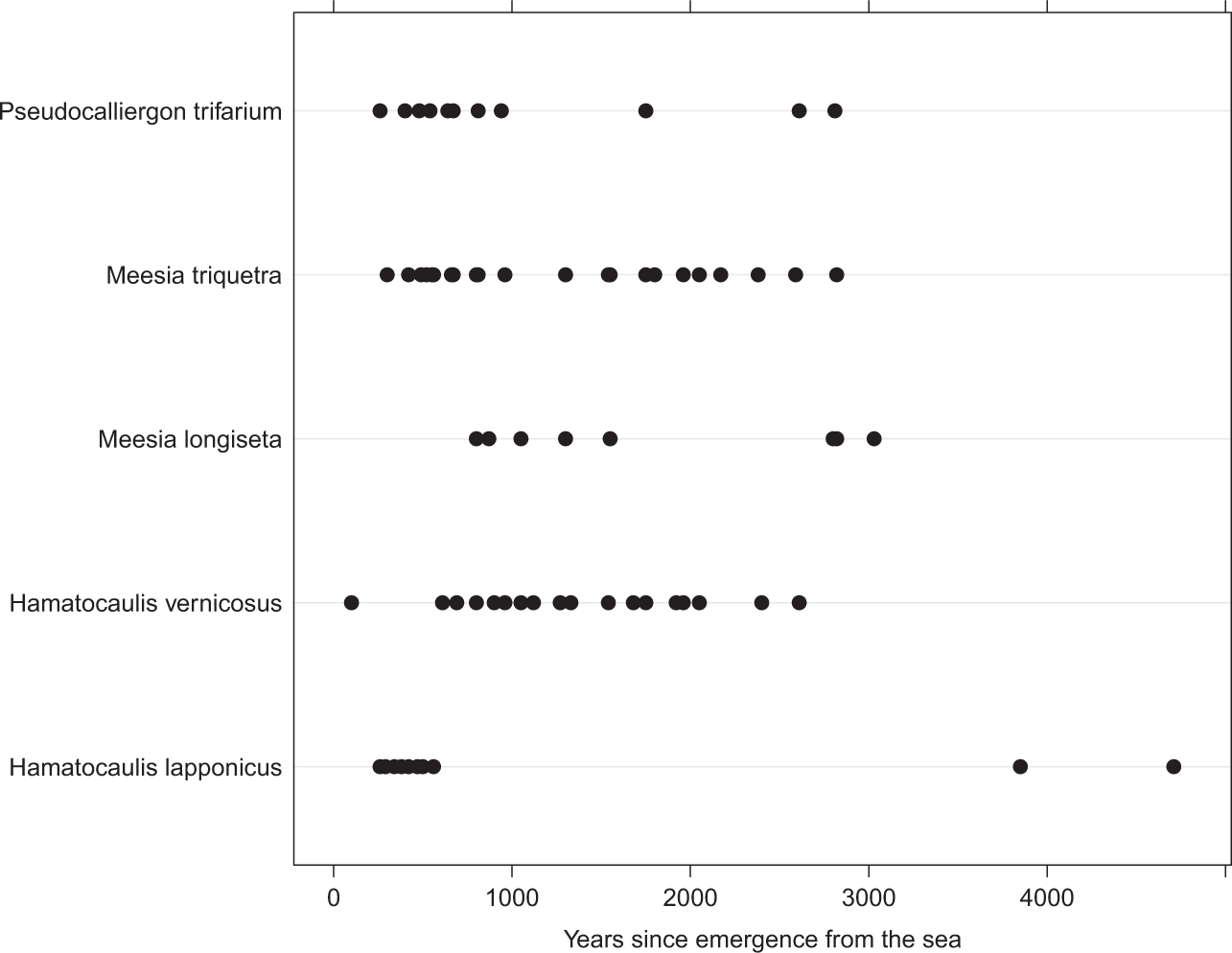
The occurrences of Meesia rich-fen assemblage mosses in peat horizons in the chronosequence of developing aapa mires. Each dot represents one sample, where some species of the group has been met. The horizontal time-scale shows at which time after the start of mire succession (years after the emergence of the place from the sea) the mosses have been growing.
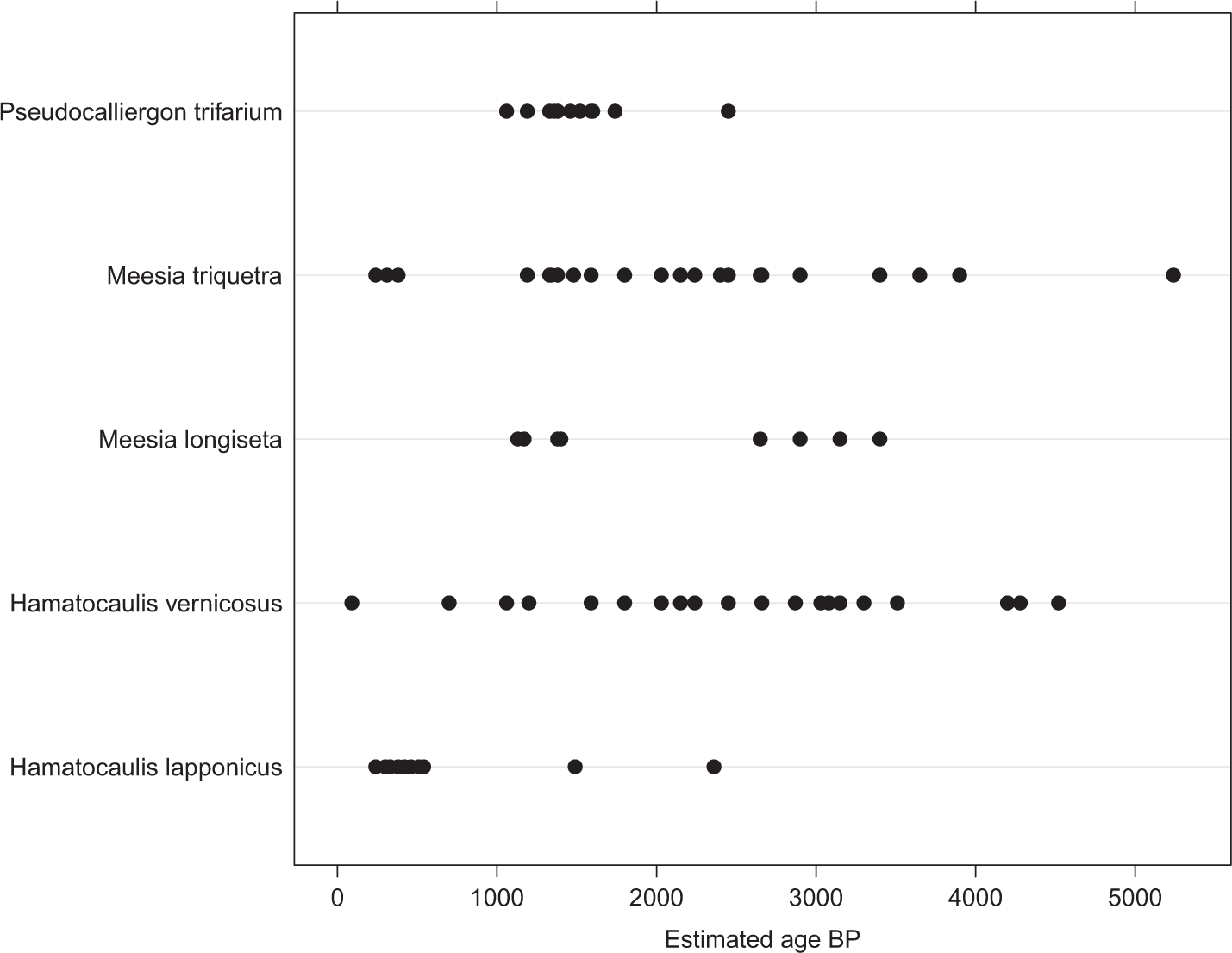
The estimated ages of the occurrences of Meesia rich-fen assemblage mosses found from the peat. Each dot represents species occurrences in one sample. The horizontal axis shows time-scale (years before present).
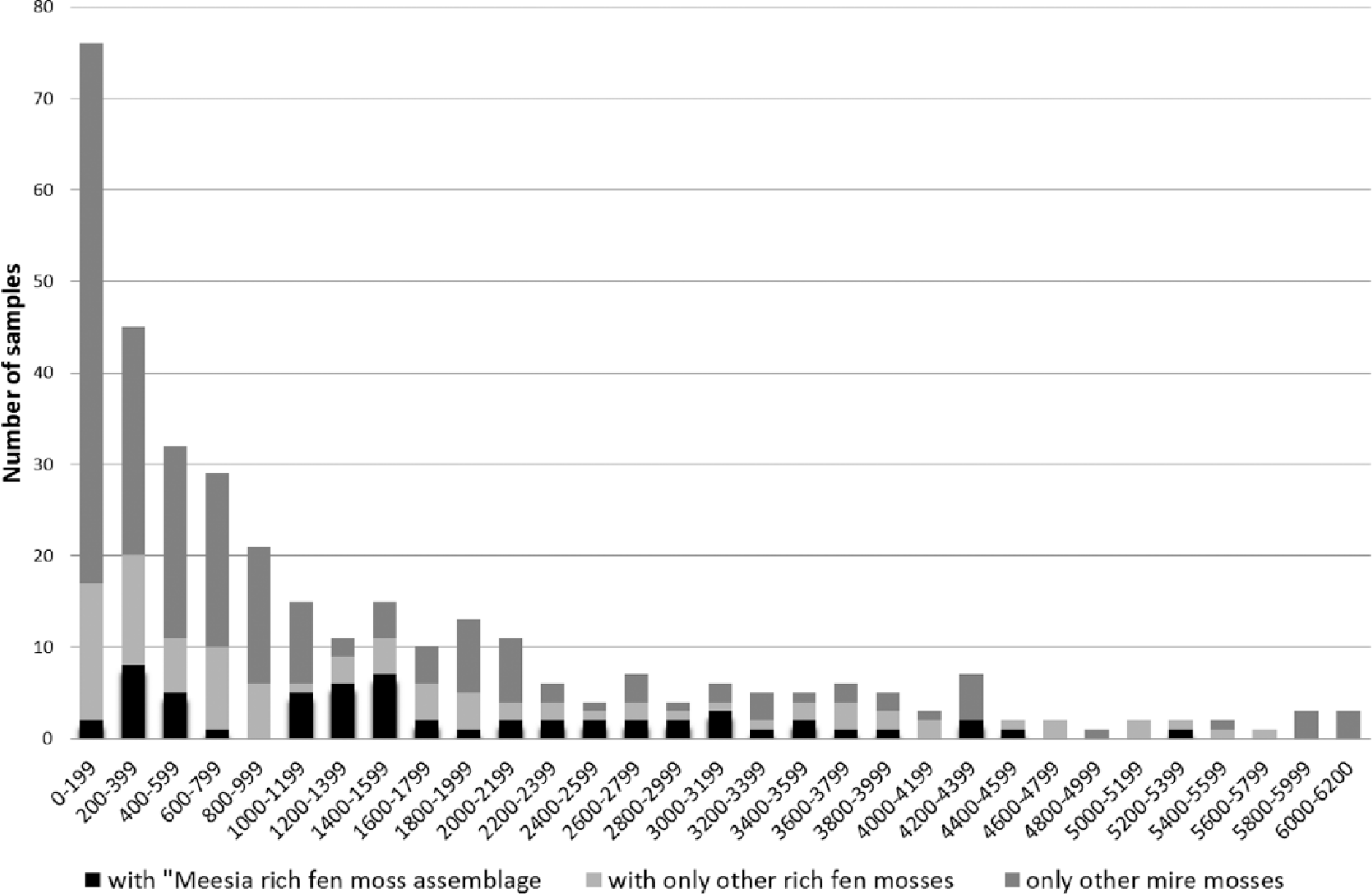
The number of peat samples with moss groups in different periods of time. The peat profiles have been divided into 200-year periods.
Discussion
The reliability of comparison between moss occurrences in peat and present vegetation
The studied peat profiles reveal that the initial succession of mires has been similar across sites during the last 5000 years. The central parts of the mire basins with groundwater influence developed from seashore meadows to wet rich fens and then to intermediate and moderately poor flark fens in a similar way in different times. The comparison of moss subfossils and current vegetation, however, shows that rich-fen mosses have decreased. The analyses of species occurrences in peat profiles are instrumental in revealing past species distributions, yet there are methodological considerations to be made. One has to consider the extent to which different mosses remain identifiable in peat. Sphagnum mosses are usually regarded as more resistant than other leafy mosses (Clymo and Hayward, 1984), but in suitable sites even very delicate and easily degradable leaf-mosses can stay unbroken and identifiable (Huttunen and Tolonen, 2006). The peat layers in the studied profiles were mostly composed of weakly humified peats, where the mosses were very well preserved. Therefore, the identification of the rich-fen mosses of the Meesia rich-fen moss assemblage seems to be reliable, because all the species were found from layers where the mosses had remained unbroken.
The robustness of used dating method
The datings are based on the presumption of uniform growth rate of peat layers on the whole profile. The actual growth rate of a peat layer can vary (e.g. Damman, 1996). However, we consider the estimated datings reasonably accurate because all the profiles represent young fens where local hydrologic conditions have very likely remained stable in their whole development history. Moreover, all profiles are composed of weakly composed peats with very little variation in humification rate; vast majority of the profiles are composed of uniform Carex- and Bryales-peats, and only the surface layers have Sphagnum-peats. Therefore, all the profiles, except some surface layers, can be considered to be produced of litter, which has accumulated quickly under anoxic conditions below water table. In these conditions, the peat accumulation rate depends mostly on organic matter production (Damman, 1996), which in this kind of wet fen ecosystem can be considered as relatively stable. Moreover, the actual targets of the datings, the occurrences of the rich-fen mosses, are mainly from either young profiles or from layers very close to the bottom. The dating of these samples probably is much less inaccurate than the dating of the other samples. The inaccuracy can be considered greatest in the upper and central parts of the longer and older peat profiles, where the changes in local hydrological conditions have probably led to changes in aerobic decay in peat, and the longer time has enabled more anaerobic decay in deeper peat horizons (Damman, 1996; Tolonen and Turunen, 1996). However, of all the findings of the Meesia rich-fen assemblage mosses, only two have been made in the upper parts of longer profiles, where the inaccuracy of even several hundreds of years could be possible (Hamatocaulis lapponicus findings given the ages 1400 and 2300 BP). It seems likely that the variation in the occurrences of the Meesia rich-fen assemblage mosses in peat layers reflects a real variation in the commonness of these mosses. The radiocarbon-dates from nearby peat profiles all concern ages of basal peats and give contribution to beginning of peat accumulation. Datings from the middle parts of the profiles could help to assess the reliability of the used supposition of linear peat growth, but unfortunately, datings of that kind seem to be lacking so far (Jukka Turunen, 2015, personal communication). The radiocarbon dating of fen peats is also problematic, because of deep-rooted sedges (Tolonen and Turunen, 1996).
The occurrence of mosses in peat layers and in current vegetation
The analysis of current and past occurrences of bryophytes (Figure 3) revealed species groups that are ecologically distinct but also may reflect habitat specific differences in the deposition in peat layers. The latter has to be considered in the interpretation of species relative occurrences in current and past mire assemblages. The species situated near the cross-section lines in Figure 3 are typical wide-spread bog and fen mosses for which suitable habitat complexes (flarks and hummocks) have been present from the early successional stages. Their occurrence in peat and present vegetation is about the same magnitude, and these analyses do not suggest any changes in their commonness. The species on the right-hand side of line, on the upper part of the graphs include mainly Sphagnum mosses which are quite resistant, and if present in peat, their remains could have been preserved. When comparing species growing typically on different levels (e.g. species of small patches of lawn- and hummock-level vegetation to species of larger wet levels), the graph based on mean coverages and concentrations (Figure 3, left) can, in our opinion, be considered as more reliable than that based on frequencies (Figure 3, right). Apparently, at least some lawn-level Sphagna having large present coverage in respect to their concentration in peat samples have become generally more common in current vegetation than they were in past. The taxa near the bottom of the graphs were found commonly in present vegetation, but only very little in peat profiles. They include species that occur in habitats, where the humification can be expected to be intense (spruce mires, dry heath-like hummocks, lawn-level rich fens, mud-bottom flarks). Typically, the leaves of these species were broken and difficult to identify. So, the scarcity of these species in peat likely reflects their relatively high humification rates. Undoubtedly, this holds true also for some species commonly found on recent vegetation, but not encountered in peat (e.g. Sphagnum fimbriatum and Loeskypnum badium).
The Meesia rich-fen moss assemblage seems to differ from other mosses by the fact that they are very scarce or lacking in present vegetation but found commonly in peat layers. The vast difference between the past and present occurrences strongly suggests that the species have genuinely decreased in mire vegetation.
Possible reasons for rich-fen bryophyte decrease
All species of the Meesia rich-fen moss assemblage are considered regionally threatened in Western Finland and three (Hamatocaulis lapponicus, H. vernicosus and Meesia longiseta) are considered threatened both in Finland and in whole Europe (The European Committee for Conservation of Bryophytes, 1995; Ulvinen et al., 2002). In Finland, all species of the group have had a large amount of occurrences all over the country, but today most of these have disappeared. Some larger occurrences still persist in central Lapland, but in the more southern parts the few occurrences are very small patches. In the vegetation of the studied mire basins, small patches of these species without sporophytes have been detected (Table 2). In the studied peat profiles, the remnants of these mosses were found in deeper layers (Table 2), but not in the living surface layer of the peat column, although some profiles were taken near living patches.
The occurrence of the Meesia rich-fen assemblage mosses on the study areas and on their surroundings. The columns: 1 – The number of discovered patches on the large reference area shown in Figure 1; 2 – The total area of the known occurrences on that area (m2) (Finnish Environment Institute database, last 25 years); 3 – An estimate of the occurrence of sporophytes on that area. Scale: − = not observed, (+) = only scattered sporophytes on some largest patches, + = some sporophytes even on small patches, but not regularly; 4 – The number of patches on the studied mire basins (m = only on moraine topography, s = only on sandy topography); 5 – The total area of patches on the studied mire basins (m2); 6 – The observations of the sporophytes on the studied mire basins; 7 – The number of studied peat profiles with occurrences of Meesia rich-fen assemblage moss subfossils (m = only on moraine topography, m,s = both on the moraine and sandy topography); 8 – The longest observed presence without breaks in the profiles (years).

The species of the Meesia rich-fen moss assemblage are typically boreal mosses with rather stringent ecological requirements. They can be considered mainly boreal species with a few occurrences in the arctic zone and with also disjunct occurrences at lower latitudes, mainly on mountains. Hamatocaulis lapponicus, however, is more restricted to north, partly also in the arctic zone. They have, however, numerous subfossil occurrences not more than a few thousand years old on areas where they are very scarce or totally absent today. These include discoveries of Meesia triquetra and M. longiseta from England (Dickson, 1973), Hamatocaulis vernicosus and Meesia longiseta from Poland (Galka et al., 2013), M. triquetra and Pseudocalliergon trifarium from Indiana (Swinehart, 1995), Hamatocaulis lapponicus from Minnesota (Griffin, 1977; Janssens, 1983b) and Meesia triquetra from Iran (Kurscher and Djamali, 2008). In Finland, occurrences of especially Hamatocaulis lapponicus but also other species of the group have been noticed in the peat layers of the land uplift coastland also outside the sites of the present study (Rehell, 1985). They all are fen species of meso-eutrophic conditions (Eurola et al., 1994). They grow in wet flark-level conditions where there is flowing ground- and surface-water (Eurola et al., 1994) which counteract the acidification of mire (Heikkilä et al., 2001; Sallantaus, 2006; Tahvanainen, 2004). One member of this group (Hamatocaulis vernicosus) grows in conditions with large variation in water quality (Stechova et al., 2008). In Finland, the typical habitats are rich fens not especially rich in calcium (Ulvinen, 2009), and this probably holds true for the other members of the group as well (see Hedenäs and Bisang, 2012 for the wide ecological niche of Pseudocalliergon trifarium and Ruuhijärvi, 1962 for the ecology of Hamatocaulis lapponicus). We thus find it unlikely that strict environmental requirements could explain their present scarcity (Söderström and Herben, 1997).
The typical occurrence of the Meesia rich-fen moss assemblage on separate, small patches suggests that the species are weak competitors (e.g. Ulvinen and Sallantaus, 2009). Considering the chemical requirements Hamatocaulis vernicosus is thought to have features typical for ruderal plants (Hájek et al., 2014). Seemingly, this species does not stand the competition of Sphagnum mosses, but grows in places where there are gaps in the Sphagnum cover. Factors reducing the growth of Sphagnum in this kind of habitat can be, for example, high pH and strong water flow (Clymo and Hayward, 1984), ferruginous precipitates and the effect of iron on the pH and organic acid concentration (Sallantaus, 2006), the erosion caused by ice, the traditional pasturing and hay-making of fens (Lindholm and Heikkilä, 2006). The dispersal ability of the Meesia rich-fen assemblage mosses seems to be restricted. They have relatively large spore sizes for which reason their ability to spread over longer distances can be weak and so the local abundance may be less directly related to wide-spread distribution (Hutsemekers et al., 2008; Virtanen, 2014). Except for Meesia longiseta, the species of this group are dioicous. That means that they can produce spores only in sites, where male and female patches meet. Small and separate patches remain sterile. The only autoicous species (M. longiseta) has sporophytes also in small occurrences, but their production frequency in extant populations is poorly known. Even for M. longiseta, the dispersal by spores is considered inefficient (Syrjänen, 2009) and a strong decline of this species has been observed in Europe (Hofmann et al., 2007).
The main reason why these rich-fen species have become threatened is undoubtedly the decline of the suitable habitats because of drainage operations. The typical growing sites, groundwater-fed fens, have been among the most heavily ditched mire site types (Lindholm and Heikkilä, 2006). They are, on the other hand, sensitive to hydrological changes happening even far from catchment areas. The decrease of groundwater flowing through the fen can cause acidification of the mire water and expansion of Sphagnum mosses (Heikkilä et al., 2001; Sallantaus, 2006). The species, which are weaker competitors, should be able to find new suitable sites to compensate the lost area. For this they need a very efficient ability to propagate. Many flark-level mosses can spread small distances by floating pieces of shoots (Cronberg, 1991; Ulvinen and Sallantaus, 2009). For longer distances, an efficient propagation by spores should be needed. In case of dioicous mosses, the decrease of populations leads quickly to the situation where the male and female gametophytes cannot meet and the production of spores cannot take place. The small, fragmentary populations are in a danger of disappearing (Longton, 1992; Longton and Hedderson, 2000). The autoicous mosses should be able to produce spores even in very small and scattered patches, but even for them inbreeding can decrease genetic diversity (Longton, 1992).
It is important to notice that the observed decrease of the Meesia rich-fen assemblage mosses has taken place in the studied mire basins where the shoreline displacement has brought new rich-fen sites. The loss of rich-fen habitats because of the natural development of aapa mires cannot explain the decline observed in this study. Additionally, there has been no ditching or other drainage on these studied basins, although the drainage operations in their surroundings have been very large. On the land uplift coast of Finland, about 95% of mires have been drained (Rehell and Heikkilä, 2009). So, these studied mires have become isolated fragments, a few hundreds of hectares in size. When the overall area of young rich fens on the whole coastland has collapsed, the overall amount of spores has diminished and the germination of them has become far more improbable than before.
Besides the drainages, one possible explanation for the decline of many rich-fen mosses is the cessation of traditional use of mires (Simola, 2006; Syrjänen, 2009; Ulvinen, 2009). The use of sedge growing fens for hay-making has been very common in Northern Finland. It started about 500–300 years ago and stopped abruptly about 50 years ago. At the same time, cattle also were grazing on mires in many places. This was especially common in the fairly densely populated coastal area. For example, on Hailuoto Island, where a group of young mire basins on sandy topography was studied, it has affected the development of vegetation very profoundly (Markkola and Merilä, 1982). However, it is clear that the rich-fen vegetation has had a long history in Finland before the start of the traditional cattle husbandry. So, although the Meesia rich-fen assemblage mosses as weak competitors may have benefited from the traditional use of mires, they have by no means been dependent on it. In large part of Northern Finland, the fens are still actively used as summer pastures for domestic reindeer. That can be one reason for the better situation of the Meesia rich-fen assemblage mosses in Lapland. The study areas 4 and 5 of this study are situated on the area of reindeer husbandry, but there these mosses were found only as subfossils from deep peat horizons. Present occurrences are situated on younger mire basins, where reindeer are met only occasionally.
The occurrences of the Meesia rich-fen assemblage species in peat profiles concentrate clearly to the young stages of mires (Figure 4), especially common they are in peat layers formed 300–2000 years after the emergence of the place from sea. Also on the present vegetation on moraine topography outside calcareous areas (Rehell and Heikkilä, 2009; Rehell and Laitinen, 2014), the rich-fen species demanding flowing groundwater have most of their occurrences on mires 300–2000 years old. On sandy topography, the species have many occurrences in peat profiles, but virtually no occurrences on present vegetation. This may be due to the fact that on sandy terrain, groundwater discharge can take place far away from the recharge areas and the studied, quite restricted mire areas do not contain true discharge areas. In samples representing older stages of mire development (over 3000 years after the emergence), there are only very few occurrences and of only two of the species, Hamatocaulis lapponicus and Meesia longiseta. These two species are known to occur on intermediate mires besides the rich fens (Ruuhijärvi, 1962). In single profiles, species of the Meesia rich-fen moss assemblage have been met typically in peat layers 10–50 cm thick. This indicates that the life-time of one patch has typically varied from a few tens of years to a few hundreds of years (Table 2).
The changes in moss assemblages could be linked to the colder and warmer epochs during the late Holocene. Such climatic changes may affect the developmental trends of mire vegetation; the warming and prolongation of the growth season is supposed to benefit the Sphagnum mosses (Alm and Saarnio, 2006; Lindholm, 1990; Tahvanainen, 2011). The occurrences of the Meesia rich-fen assemblage mosses in the peat samples seem to have great variation during different ages. The occurrences of other rich-fen mosses, on the contrary, do not show similar periodical variation, and their frequency seems to have been fairly constant. The samples with the Meesia rich-fen assemblage mosses seem to concentrate to horizons dated to periods known to have been cold (Eronen, 1991, 1997; Gunnarsson et al., 2011; Linderholm et al., 2010; Lunkka, 2008). The first maximum of occurrences (about 200–600 BP) could be interpreted to correlate to the ‘Little Ice Age’ (discontinuous cold period most intensive in the 17th century). In the period just before that (600–1000 BP), the Meesia rich-fen assemblage mosses nearly lack from the samples. This could be correlated to the warmer period of the Middle Age, when at times the temperatures may have been as high as today (Gunnarsson et al., 2011). The period before that is supposed to have been colder, and there is quite clear maximum at about the period 1200–1600 BP. The time before that is represented by so few samples that it is impossible to see any trends in occurrences, but the Meesia rich-fen assemblage mosses have been detected quite frequently also from the older layers.
Concluding remarks concerning the changes of rich-fen bryophytes
Our study presents new perspectives on how rare and endangered Meesia rich-fen assemblage mosses are driven by various ecological changes in different time scales. The results revealed their relatively high frequency in the past and a very dramatic disappearance in the last centuries. These rich-fen mosses show trends and fluctuations in occurrences that seem to be associated with known climatic oscillations in the later part of the Holocene. The influence of climatic trends on these mosses is probably due to the competition of lawn-level Sphagnum mosses. The latter mosses are known to drive mire succession (Van Breemen, 1995). They can benefit from the longer growing seasons with long, moist autumn season (Alm and Saarnio, 2006; Lindholm, 1990) and diminishing runoff because of increased evaporation (Tahvanainen, 2011). Recent expansion of lawn-level Sphagnum mosses on the mires of land uplift coast has been observed in peat profiles by Tuittila et al. (2012) and thought to coincide with the warm temperatures during the last 100 years. Thermal conditions can also affect the amount of dissolved organic acids in mire waters, which is crucial for the pH (Sallantaus, 2006). The present-day decline of Meesia rich-fen mosses is undoubtedly linked to the large drainage operations of rich fens, which have fragmented the populations too small and scattered. Warming of the climate and ceasing of traditional land use in mires have then enhanced their rate of disappearance.
Footnotes
Acknowledgements
Antti Huttunen and Raimo Heikkilä gave valuable comments on the issues of mire stratigraphy and ecology. Tauno Ulvinen helped in identifying moss species. Jukka Turunen gave information of the C-14 dates in GTK. Two anonymous referees made useful comments on the article.
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.