
Abstract
Two high-altitude lake-sediment sequences (Lake Lia, 1910 m a.s.l. and Lake Bucura, 2040 m a.s.l.) from the Retezat Mountains (South Carpathians, Romania) were analysed using multi-proxy methods to study responses of treeline, timberline and alpine/subalpine vegetation to climate change and human impact during the past 16,000 years. Woody species (Pinus mugo, Pinus cembra, Picea abies and Juniperus communis) reached Lake Lia between 12,000 and 11,800 cal. yr BP, whereas P. mugo colonised the shores of Lake Bucura at 9600 cal. yr BP. Lake Lia was in the timberline ecotone between 8000 and 3200 cal. yr BP, in semi-open P. cembra and Picea abies woodland, probably mixed with P. mugo on the steeper slopes. Lake Bucura was surrounded by the upper part of the krummholz zone during the mid-Holocene. The increase in P. cembra after c. 6000 cal. yr BP around Lake Lia suggests that the composition of the timberline forest changed. The disappearance of P. cembra and Picea abies around Lake Lia at ~3000 cal. yr BP reflects descent of the timberline. A large mean July temperature decline between 3300 and 2800 cal. yr BP may have driven or at least contributed to the descent of the Picea abies–P. cembra forests. An increase in human indicator pollen types in Lake Bucura around 4200 cal. yr BP may reflect human impact in the naturally open alpine zone in the Late Bronze Age. In contrast, human impact likely appeared considerably later, around 2650 cal. yr BP (Early Iron Age) around Lake Lia in the upper subalpine zone. Human impact likely intensified after 2200 cal. yr BP at both sites that resulted in the lowering of the krummholz zone. We conclude that climate change and human impact both played an important role in the lowering of the treeline and timberline in the late-Holocene.
Introduction
The treeline ecotone is a transitional belt between the upper limit of closed forest (altitudinal timberline) and treeless alpine vegetation in high mountains (Däniker, 1923; Lotter et al., 2006; Wieser and Tausz, 2007). In the treeline ecotone, the density of trees decreases upwards, and the highest elevation reached by single tree individuals taller than 2–3 m defines the treeline (Arno and Hammerly, 1984; Schwörer et al., 2013; Tinner and Theurillat, 2003). Dwarfed trees and shrubs growing above the treeline form the krummholz zone (Holtmeier, 1981; Körner, 2012a; Tinner, 2007).
Since the elevation of the potential timberline and treeline is primarily determined by climate (Holtmeier, 1981; Körner, 2003), researchers generally agree that the treeline ecotone will be strongly affected by present-day warming (Grace et al., 2002; Solomon et al., 2007). Signs of upward treeline and timberline shifts have already been detected in several decadal-scale studies (Harsch et al., 2009; Heiri et al., 2014; Körner, 2012b; Theurillat and Guisan, 2001; Tinner and Ammann, 2005), supporting predictions that ongoing global warming leads to rises in treeline and timberline with corresponding reductions in alpine grassland (Schwörer et al., 2013).
Treeline and timberline dynamics can be understood using a combination of short-term ecological observations with longer-term reconstructions based on palaeoecological proxies such as fossil pollen, plant macrofossils, stomata and charcoal, and in some cases megafossils and tree rings (Birks and Birks, 2000; Reasoner and Tinner, 2009; Tinner, 2007). Several long-term multi-proxy records from the Alps illustrate the effects of past climate and land-use changes on treeline ecology (Berthel et al., 2012; Finsinger and Tinner, 2007; Gobet et al., 2003, 2005; Lang and Tobolski, 1985; Rey et al., 2013; Schwörer et al., 2013, 2014; Tinner et al., 1996; Tinner and Kaltenrieder, 2005; Tinner and Theurillat, 2003; Wick et al., 2003; Wick and Tinner, 1997). The use of plant macrofossil and stomata analysis in many of these studies provides a detailed picture of local vegetation dynamics around the treeline ecotone (Tinner, 2007), because of the lower dispersal distance of plant remains compared with pollen (Ammann et al., 2014; Birks and Bjune, 2010). In contrast, plant macrofossil analysis has only recently been adopted to investigate long-term treeline dynamics in the Carpathians (Feurdean et al., 2016; Geanta et al., 2014), and previous reconstructions were mainly based on pollen records (Fărcaş et al., 1999, 2013; Feurdean and Willis, 2008). Because of the low number of long-term records and the inherent limitations of pollen records as a tool to reconstruct treeline dynamics, knowledge of the long-term dynamics of upper treeline and timberline in the Carpathians is scarce and fragmentary.
Currently, evidence for past human activity and their effects on mountain vegetation in the South Carpathians and transhumance is based on documentary archives, modern vegetation studies and archaeological data. According to historical evidences, the first shepherds moved up to the high mountain pastures from the plains only 800 years ago and started clearing forests to create pastures and meadows in the Romanian section of the Carpathians, later spreading northwards along the Carpathian chain (Mertens and Huband, 2004). Written sources document that grazing contributed to the lowering of the timberline in the Iezer Mountains (Mihai et al., 2007), while the enlargement of grazing fields by slash and burn activities is documented in the Fagaras Mountains (Nedelea et al., 2009). Prehistoric settlements are found in the lowlands surrounding the Retezat Mountains (Supplementary Figure 6, available online), and evidence for prehistoric pastoral farming is also available in the nearby areas (e.g. Foeni-Salaș, W-Banat, Romania; Arnold and Greenfield, 2006). A recent study by Juler (2014) focused on Medieval long-distance transhumance in Romania and its fast disappearance after communist times. In that study, an important transhumance route runs through the Retezat Mountains. Some vegetation studies conclude that intensive summer alpine pasturing had a direct influence on the treeline ecotone vegetation (Coldea and Cristea, 1998) and suggested that land use led to the appearance of new plant communities (e.g. Potentillo–Nardion and Calamagrostion) and facilitated the spread of heathlands (Loiseleurio–Vaccinion, Rhododendro–Vaccinion) in the low-alpine zone following clearance of the native woody vegetation (Rhododendro–Pinetum mugi and Bruckenthalio–Piceetum; Coldea and Cristea, 1998). Geanana (1991) concluded that logging and grazing widely affected the landscape during the 20th century lowering the elevation of the treeline about 100 m below the potential tree limit. These inferences contrast to records from the Eastern Carpathians and the Alps. The elevation of the treeline in the Central Alps was probably 150–200 m higher than today’s tree limit between 11,000 and 4500 cal. yr BP (Tinner and Theurillat, 2003); and the onset of the exploitation of the alpine grassland zone started much earlier (late Neolithic; Oeggl, 1994; Tinner et al., 2007). In the Alps, timberline descended around 6500 cal. yr (Tinner, 2007) and in the Eastern Carpathians around 6000–5000 cal. yr BP (Feurdean et al., 2016; Geanta et al., 2014).
The aims of this study are to investigate the vegetation changes at the current treeline ecotone over time and to understand whether the driving factors of these changes were climate change or human impact. To address these aims, we use plant macrofossil, pollen, stomata and charcoal records from two lakes situated at different altitudes in the same mountain valley. The study sites are located in the Retezat Mountains (Figure 1), about 300 km south of the sites in the Eastern Carpathians investigated by Feurdean et al. (2016) and Geanta et al. (2014). Neolithic and Bronze Age archaeological sites have been reported from the lowlands of the nearby Haţeg Basin and from south-east of our study area (Institutul National al Patrimoniului, 2013; Luca et al., 2005), indicating human presence in the region. Our key questions are as follows: (1) how did Holocene treeline ecotone vegetation in the Retezat Mountains respond to climate change? (2) did human activities influence high mountain ecosystems? and (3) how do Holocene treeline fluctuations in the Retezat Mountains compare with the treeline variations in the Eastern Carpathians and other European mountains?
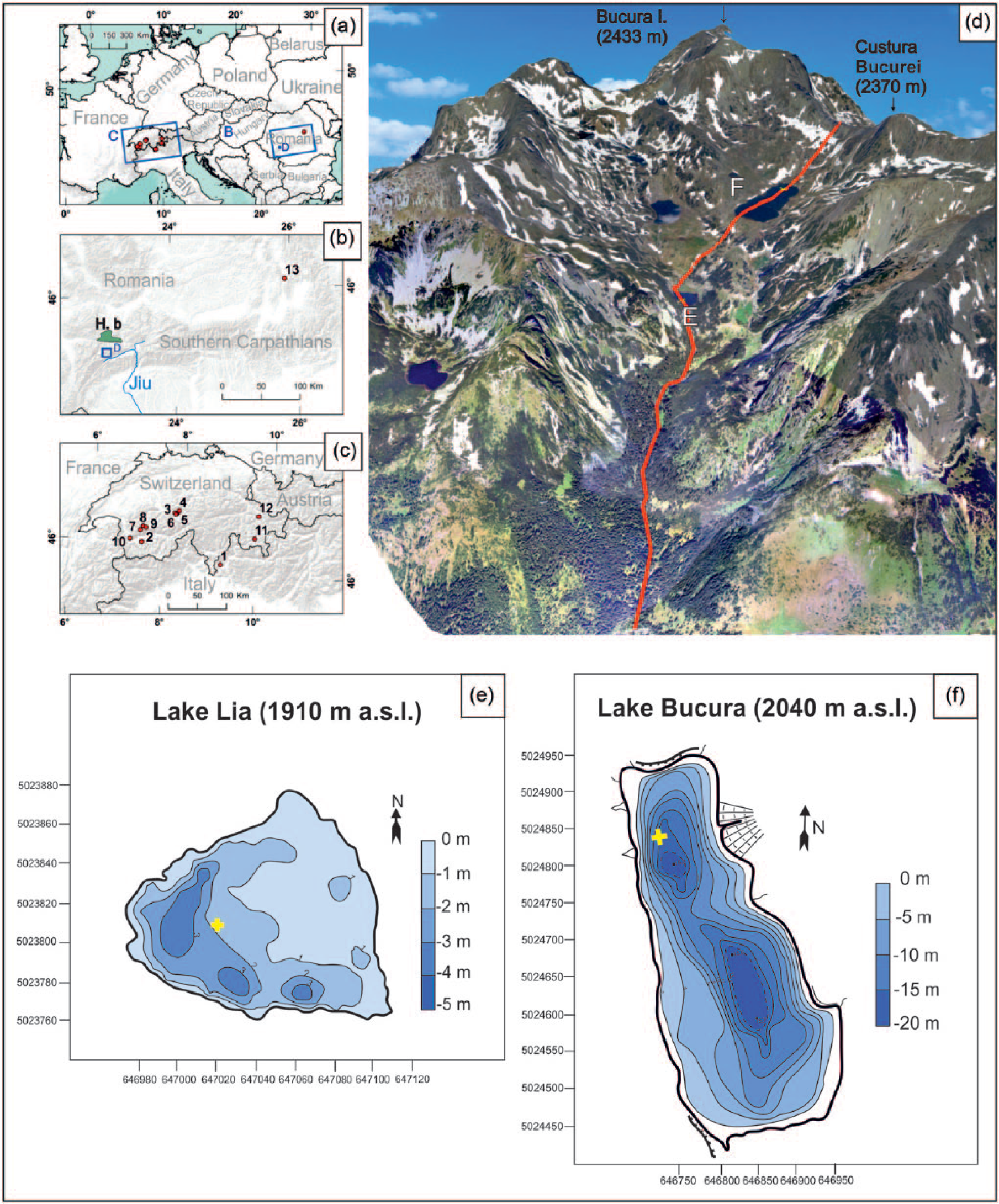
(a) The location of the Retezat Mountains (RM) in Europe and in the Southern Carpathians (H.b. = Haţeg basin). (b) Selected sites with important palaeorecords mentioned in the text (b and c): 1. Lago Basso (2250 m a.s.l.; Wick and Tinner, 1997), 2. Gouille Rion (2343 m a.s.l.; Wick and Tinner, 1997), 3. Sägistalsee (1935 m a.s.l.; Wick et al., 2003), 4. Hinterburgsee (1515 m a.s.l.; Heiri et al., 2003), 5. Bachalpsee (2265 m a.s.l.; Lotter et al., 2006), 6. Feld mire (2130 m a.s.l.; Lotter et al., 2006), 7. Emines (2288 m a.s.l.; Berthel et al., 2012), 8. Lauenensee (1381 m a.s.l.; Rey et al., 2013), 9. Iffigsee (2065 m a.s.l.; Schwörer et al., 2013), 10. Lac de Fully (2135 m a.s.l.; Finsinger and Tinner, 2007), 11. Lej da Champfèr (1791 m a.s.l.; Gobet et al., 2003), 12. Lej da San Murezzan (1768 m a.s.l.; Gobet et al., 2003), 13. Lake St Anne (Eastern Carpathians; 950 m a.s.l., Magyari et al., 2014). (d) Location of Lake Lia (e) (1910 m a.s.l.) and Lake Bucura (f) (2040 m a.s.l.) in the Bucura valley on the southern slope of Retezat Mountains. (e) Bathymetry of Lake Lia: 45°21′7.3″N, 22°52′39.3″E; coring point marked by a cross. (f) Bathymetry of Lake Bucura: 45°21′42.9″N, 22°52′27″E coring point marked by a cross. The bathymetric maps were modified after Vespremeanu-Stroe et al. (2008).
Study area
The Retezat Mountains are situated on the western side of the South Carpathians in Romania (Figure 1). The mountain range stretches from east to west between the Jiu river valley (about 45 km north from the valley) and the Haţeg Basin (about 25 km from the basin; Figure 1b), and most of it belongs to the Retezat National Park, which hosts a rich flora with 90 endemic and 130 rare and protected species (Borza, 1934; Csűrös et al., 1956; Nyárády, 1958). The bedrock is predominantly granite and granodiorite with crystalline schist intercalated between the northern Retezat and the southern Buta massifs (Jancsik, 2001). There are more than 50 mountain peaks reaching altitudes >2000 m a.s.l., and the highest is the Peleaga peak (2509 m a.s.l.).
The climate is temperate continental in the lowland area (Farcas and Sorocovschi, 1992), and alpine climate characterises the high mountains. Because of the combined oceanic and Mediterranean climatic influence, it is one of the wettest regions in the Romanian part of the Carpathians (Magyari et al., 2013). The present treeline is located at 45°35′N, 22°87′E at 1900 m a.s.l., while timberline runs at 45°34′N, 22°88′E at 1850 m a.s.l. on the southern slope. According to the study of Farcas and Sorocovschi (1992), January mean temperature is −8°C at the treeline (1900 m a.s.l.) and between −7°C and −8°C at the timberline on the southern slope of the Retezat Mountains. July mean temperatures are 10°C at the treeline and between 10°C and 11°C at the timberline. The annual mean precipitation is c. 1200 mm at the treeline, 1300 mm at the timberline and <1200 mm above 2000 m a.s.l. These environmental conditions determine the altitudinal zonation of the vegetation (Borza, 1934; Csűrös et al., 1956; Nyárády, 1958) that is summarised in the Supplementary Material (available online).
Study sites
We analysed two lake-sediment sequences from the Bucura valley on the southern flank of the Retezat Range (Figure 1d). Lake Lia at 1910 m a.s.l. is a small shallow lake (1.26 ha, average water depth: 0.8 m; Figure 1e) located at the present treeline. It has two inflow streams and one outflow. Dense Pinus mugo thickets occupy its steep western slope and only one dwarfed Picea abies individual is well established on the lakeshore. Picea abies and Pinus cembra are present near the southern shore c. 150 m away, and a few specimens grow on the ridge above the lake. The lake and its inflowing streams are fringed by herbs of mesic or wet soils (e.g. Caltha palustris, Rumex alpinus, Aconitum tauricum, Adenostyles alliariae, Epilobium angustifolium, Heracleum palmatum).
Lake Bucura at 2040 m a.s.l. lies above the treeline and is the largest lake in the Retezat (8.92 ha, maximum water depth: 17.5 m (Vespremeanu-Stroe et al., 2008) and Figure 1f). It receives water from two inflow streams and has an outflow on its southern side towards Lake Lia. It is surrounded by open alpine meadow vegetation and screes. At present, P. mugo is scarce on the lakeshore (10 individuals growing <20 m from its shores), but it also grows above the lake. On rocky terrain Juniperus communis is also present. Cattle grazing is frequent during the summer around both lakes. The treeline is formed by Picea abies and P. cembra individuals around 1900 m a.sl., while the timberline is around 1850 m a.sl. on the southern slope of Bucura valley.
Materials and methods
Sediment sampling
Sediment cores from Lake Lia (LIA-1) and Lake Bucura (BUK-5) were obtained in 2008 using Livingstone and Kullenberg piston corers. The 880-cm-long sediment core from Lake Lia was taken close to the western shore (45°21′7.3″N, 22°52′39.3″E) under 1.1 m water (Figure 1e). The 550-cm-long sediment core from Lake Bucura was taken in the northern sub-basin (45°21′42.9″N, 22°52′27.0″E) under 13.9 m water (Figure 1f). The Bucura sediment was contaminated below 495 cm depth because of technical problems, so only the top 495 cm was analysed. Because of the unavailability of a specialised surface sediment sample, the core tops do not cover the most recent times. In the laboratory, the sediment cores were photographed and lithostratigraphically described following Troels-Smith (1955; Figure 2), sliced into plastic bags and refrigerated.
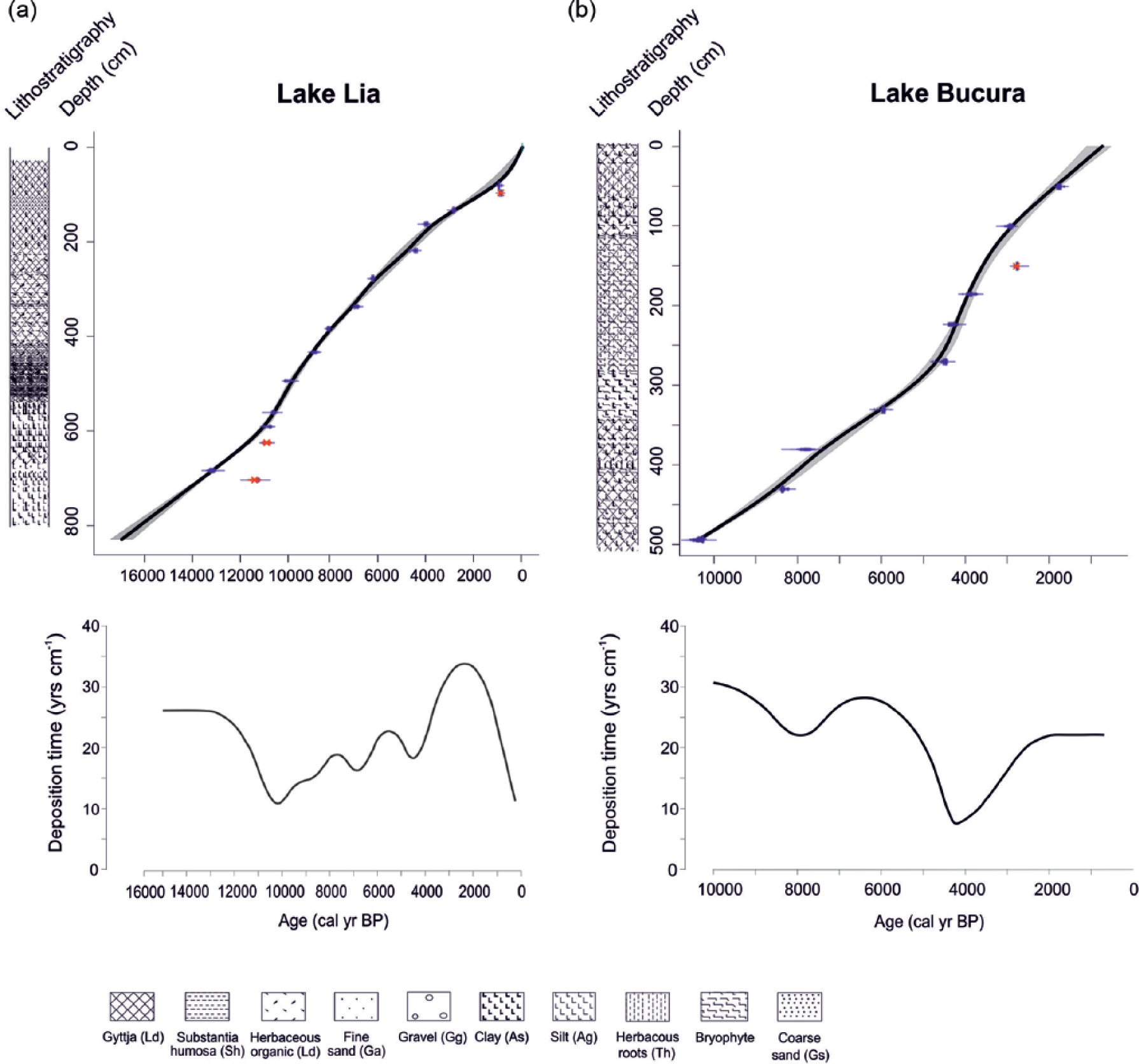
Age–depth models and deposition times for the (a) Lake Lia and (b) Lake Bucura sediment profiles. The age–depth models are based on 16 14C dates from Lake Lia and 12 14C dates from Lake Bucura sequence calibrated using CALIB Rev. 7.2.0. and the IntCal13 data-set (Table 1). The age–depth models were created with the smooth-spline curve fitting function in CLAM v2.1 (Blaauw, 2010). The lithostratigraphy is represented using the symbols of Troels-Smith (1955) on the left of each model and detailed in Supplementary Table 1 (available online). Troels-Smith’s symbols: Gyttja (Ld), substantia humosa (Sh), herbaceous organic mud (Ld), fine sand (Ga), gravel (Gg), clay (As), silt (Ag), herbaceous remains of bryophytes (Th), coarse sand (Gs).
AMS 14C dates from Lake Bucura (BUK-5) and Lake Lia (LIA-1). 14C measurements were done in the Hertelendi Laboratory of Environmental Studies, Institute of Nuclear Research of the Hungarian Academy of Sciences, Debrecen. Outliers were detected with the BACON R package (Supplementary Figure 1, available online). For the LIA-1 core the age of the lake-sediment surface was estimated by extrapolation.
Depths (cm) are the bottom depths of samples; all dated sample width is 1 cm long.
Low temperature (LT): 400°C; high temperature (HT): 800°C.
Radiocarbon dating and chronology
Table 1 lists the 28 accelerator-mass spectrometry (AMS) 14C measurements obtained. Plant macrofossils were 14C dated plus three dates on the <250 µm organic-rich sediment fraction abundant in fine organic debris, Cladocera and Chironomidae remains. Previous attempts to compare the 14C-dating results of various sediment components of the lake sediments in the Retezat Mountains suggested that the lakes show no reservoir effect (Hubay et al., 2016; Magyari et al., 2009a, 2012); the bedrock is poor in calcium, and generally terrestrial plant macrofossil components provide similar 14C ages to the aquatic animal components (Magyari et al., 2009a, 2012). No macrofossils were found in the highly inorganic late-glacial sediment of Lake Lia, so we dated bulk sediment. 14C dates were calibrated into calendar years before present (AD 1950) using the IntCal13 calibration curve (Reimer, 2013). Bayesian age–depth modelling was used to detect outlier radiocarbon dates (Blaauw and Christen, 2013) with the R-BACON package (Supplementary Figure 1, available online). The programming algorithm of BACON uses the deposition time of the previous depth interval to predict age–depth relations in the subsequent interval. Even though this so-called ‘memory time’ can be modified, when there are rapid shifts in deposition time the model still provides biased age–depth curves. Therefore, we experimented with more traditional curve fitting methods in CLAM v2.1 using the outlier information from BACON (Figure 2). Finally, we decided to use the smooth-spline curve fitting function to model the age–depth relationship in the two lake-sediment sequences (Blaauw, 2010), as shown in Figure 2 together with the sediment deposition time. Further details on the radiocarbon dating and age–depth modelling of the Retezat lake sediments are provided in Hubay et al. (2016).
Plant macrofossil analysis
Two-cm-thick subsamples (average volumes 21 and 5.4 cm3 in Lake Lia and Lake Bucura, respectively) were taken at 8- to 16-cm intervals from the LIA-1 core and at 8-cm intervals from the BUK-5 core. Bucura subsamples were briefly soaked in 10% NaOH and wet-sieved through 250 and 180 µm meshes. Clay-rich samples from Lake Lia were treated with Na4P2O7 and all were sieved through a 125-µm mesh. Plant remains were picked out systematically under a stereomicroscope (Olympus SZ 51 and LEITZ, ×10 magnification), identified using keys (Bojnanský and Fargašová, 2007; Katz et al., 1965; Schermann, 1967; Tomlinson, 1985; Velichkevich and Zastawniak, 2008) and determined to the smallest taxon level by comparison with the reference collections of the MTA-MTM-ELTE Research Group for Paleontology, the Hungarian Natural History Museum and the Palaeoecology Laboratory of Bergen University. For conifer needles, the numbers of complete needles (WN), needle fragments (NF), needle tips (NT) and needle bases (NB) and the highest value among the needle tips and needle bases (NH) were noted. The minimum number of needles (MN) in a sample was estimated using the following formula: MN = (WN + (NF/2) + NH). Sediment components (e.g. Cladocera, chironomids, bryophyte leaves and Cenococcum sclerotia) of Lake Bucura were counted and calculated according to a modified version of the QLCMA method of Barber et al. (1994), as detailed in Jakab et al. (2004). Plant macrofossil concentrations were standardised to a reference sediment volume of 20 cm3 for Lake Lia (average samples volume was 21 cm3) and 5 cm3 for Lake Bucura (average sample volume was 5.4 cm3). Macrofossil concentration diagrams were drawn using TILIA software (Grimm, 1992) and divided into macrofossil assemblage zones (LM 1–5, BM 1–5) visually, taking into account major compositional and concentration changes in the macrofossil assemblages.
Pollen and stomata analyses
Subsamples of 1 cm3 were taken at 8-cm (Lake Lia) and at 4- to 8-cm (Lake Bucura) intervals and processed for pollen analysis using standard methods (Bennett and Willis, 2001), but excluding the acetolysis step. Pollen, spores and stomata were identified and counted under an Olympus CX41 light microscope at ×400 and ×1000 magnification. At least 500 terrestrial pollen grains were counted at each level. For identification, the pollen atlases of Reille (1992, 1995, 1998) and the pollen identification keys of Moore et al. (1991) and Beug (2004) were used. For the identification of the Sordariales-type spores, descriptions in Van Geel et al. (1989) were used. Pollen diagrams were drawn using Psimpoll v.27 (Bennett, 2005). Local pollen-assemblage zones were determined using the method of optimal splitting by information content on the terrestrial pollen taxa that reached at least 5% in one sample as implemented in Psimpoll. Seven pollen zones were identified in each sequence. Pollen percentages are based on the terrestrial pollen sum (M). Relative frequencies (F) of aquatic pollen types + fungal spores + algae (A) were calculated according to the formula F = ΣA/(M + A).
Stomata of Picea abies, Abies alba, J. communis, Larix decidua, P. cembra and species of the subgenus Pinus (diploxylon pines) were identified and counted using the keys of Sweeney (2004) and Magyari et al. (2012). Stomata of the subgenus Pinus (diploxylon pines) are probably produced by P. mugo, because it is the only diploxylon pine at mid and high altitudes in the Retezat Mountains today.
Pollen types indicative of human impact are presented in Figures 5 and 7 and Supplementary Figure 5 (available online). They include herbs associated with pastures, meadows and trampled ground at both high and low altitudes: Plantago lanceolata, Plantago major/media, Artemisia and Poaceae. Because of their wide distribution, good pollen productivity and dispersal, we interpreted the relative frequency increases in these pollen types as indicative of either local or regional (lower elevation) human impact. Alpine grasslands and mats respond to pasturing and mowing by the expansion of nutrient-demanding herbs (Oeggl, 1994), many of which are insect-pollinated, producing low amounts of pollen that do not travel far from their origin. They are therefore underrepresented in the pollen records. Supplementary Table 2 (available online) lists some of these taxa in the Retezat Mountains frequently encountered on our pollen slides. We interpreted the relative frequency increase in these herb pollen types as indicative of local and extra-local human impact, mainly pasturing in the vicinity of the studied lakes. These taxa include Vaccinium-type, Adenostyles, Achillea, Sedum and Phyteuma-type. Pollen studies from high altitudes in the Alps show that the pollen of local pasturing indicator herbs can be used successfully to identify high-altitude pasturing (Bortenschlager, 1991; Court-Picon et al., 2006; Vorren et al., 1993).
Loss-on-ignition
Subsamples of 1 cm3 sediment were taken at 1-cm intervals from core LIA-1 and at 2-cm intervals from core BUK-5. Samples were dried at 105°C overnight and then burned at 550°C for 4 h. Organic matter content was estimated as the percentage loss in dry weight after ignition (Heiri et al., 2001).
Charcoal analyses
For macrocharcoal analysis, subsamples were taken at every centimetre from the top 100 cm of the BUK-5 core, followed by 2 cm contiguous (merged) sub-sampling below 100 cm. For core LIA-1, samples were taken at 1-cm contiguous intervals. Methods were as described in Finsinger et al. (2016), and they resulted in counts of macrocharcoal particles and their surface area (macrocharcoal accumulation rates (macroCHAR)).
Microcharcoal particles (>10 µm) were counted on the pollen slides following Tinner and Hu (2003) and Finsinger and Tinner (2005) and presented as microcharcoal accumulation rates (microCHAR). These records are used to reconstruct regional (20–50 km extent) fire activity around the study sites following Tinner et al. (1998). Experimental and empirical evidence indicates that most macrocharcoal particles (>160 µm in diameter) originate from fires nearer to their source area (max. 10–20 km) because of the more limited dispersal of larger charcoal particles (Lynch et al., 2004; Oris et al., 2014; Peters and Higuera, 2007). Long-term variations in total macroCHAR may reflect biomass burning, that is, an integrated signal of charcoal production and fire frequency. To determine long-term variations in regional fire activity and local biomass burning, microCHAR and macroCHAR records were smoothed with a 900-year-bandwidth lowess (Cleveland and Devlin, 1988). Because the sample deposition times (years sample−1) for Lake Bucura were too high below 100 cm depth, the record was not analysed to infer local fire episodes using the decomposition approach (Finsinger et al., 2014; Higuera et al., 2009; Kelly et al., 2011).
Results and interpretation
Chronology and sediment stratigraphy
For a detailed lithostratigraphic description of the profiles, see Supplementary Table 1 (available online). The lithostratigraphy is shown in Figure 2 using the Troels-Smith’s (1955) notation.
The BACON age–depth model of Lake Lia suggested three outlier 14C dates (Table 1, Supplementary Figure 1, available online) that were excluded from the smooth-spline age–depth model. The smooth-spline age–depth model (Figure 2a) was constructed between the top and bottom of the core (between 31 and 830 cm). The estimated ages of the top and bottom samples (~250 and ~17,100 cal. yr BP, respectively) were obtained by extrapolation of the age–depth model and therefore have high uncertainty. The average sediment deposition time is 26 yr cm−1 before 12,000 cal. yr BP, 16 yr cm−1 between 12,000 and 6900 cal. yr BP, increasing to 22 yr cm−1 between 6900 and 2000 cal. yr BP, before decreasing towards the present.
One outlier 14C date was detected at Lake Bucura (Table 1, Supplementary Figure 1, available online) and was excluded from the smooth-spline age–depth model (Figure 2b). The estimated age of the top of the core is ~650 cal. yr BP, obtained by extrapolation of the age–depth model and therefore it has a high uncertainty. The average sediment deposition time is 26 yr cm−1 between 10,300 and 6300 cal. yr BP, 18 yr cm−1 between 6300 and 4200 cal. yr BP and 17 yr cm−1 between 4200 and 700 cal. yr BP.
Vegetation histories
Lake Lia
The macrofossil record from Lake Lia is presented in Figure 3 and the pollen record in Figure 5a. The basal plant macrofossil zone, LM-1 (between 15,900 and 12,000 cal. yr BP; 782–636 cm), contained one charred Picea abies needle fragment at 14,600 cal. yr BP and some seeds of Silene sp., R. alpinus-type and Carex sp. indicating that the treeline was located below Lake Lia. The single charred Picea abies needle fragment could have been transported for hundreds of metres in the convective columns of fires at lower altitudes (Pisaric, 2002).
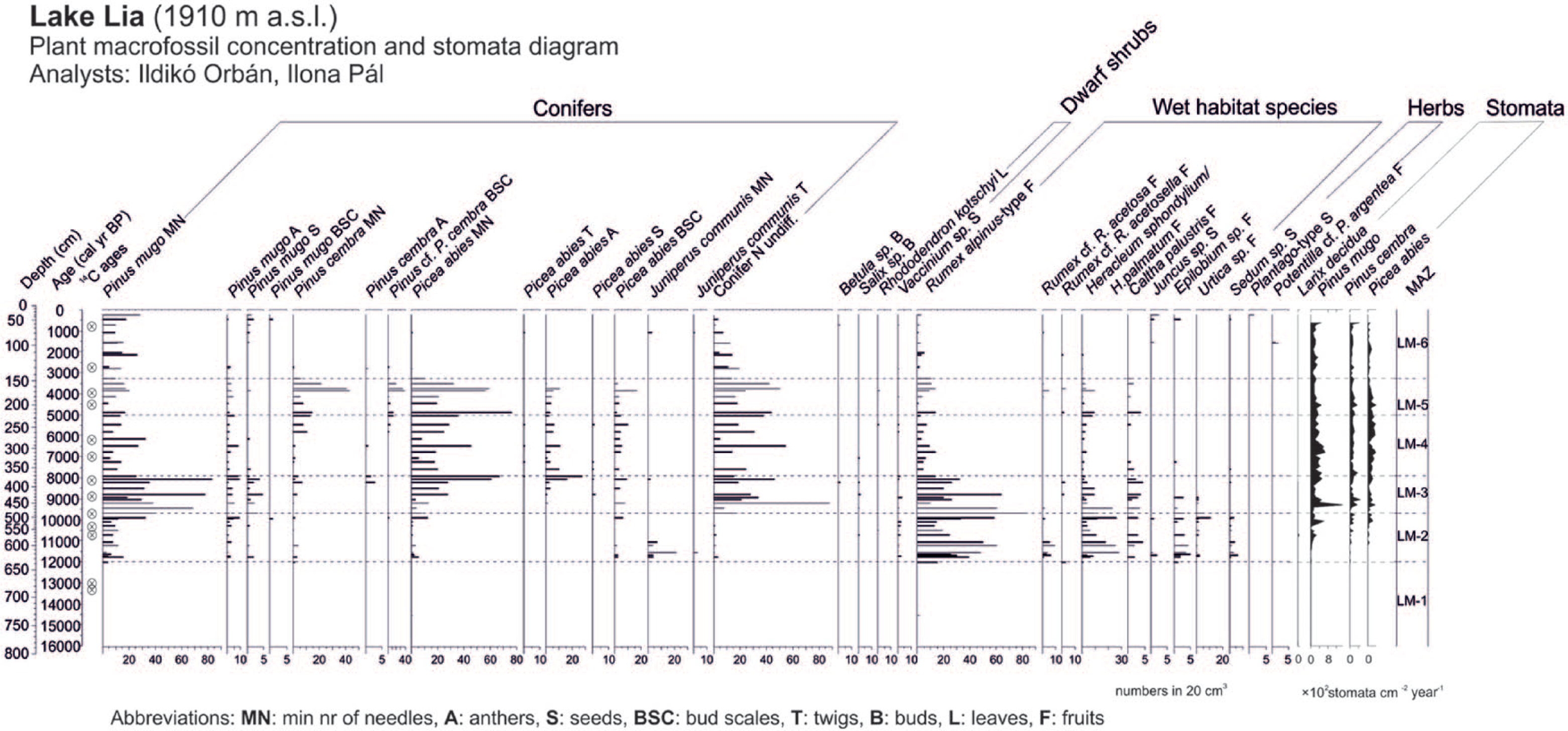
Plant macrofossil concentration diagram of Lake Lia plotted on a calibrated age scale showing selected taxa together with the stomata accumulation rates. All macrofossil samples are standardised to numbers in 20 cm3 sediment. Stomata were counted along with pollen. LM zones are local macrofossil assemblage zones in Lake Lia, delimited visually.
The treeline reached the catchment around 12,010 cal. yr BP (start of zone LM-2; 12,010–9700 cal BP; 636–483 cm) as shown by the first appearance of P. mugo and low concentrations of P. cembra needles that were closely followed by the first occurrences of macrofossil remains of Picea abies and J. communis. Stomata of P. mugo–type were first detected at 11,400 cal. yr BP, closely followed by the appearance of P. cembra and Picea abies stomata at 11,000 cal. yr BP. One L. decidua stoma was found at 10,700 cal. yr BP (568 cm). During the equivalent pollen zone, L-4 (Figure 5) percentages of Picea abies, Corylus and tree pollen of Quercetum mixtum (Quercus, Ulmus, Tilia and Fraxinus) increased and herb pollen of Artemisia, Chenopodiaceae and Poaceae distinctly decreased reflecting the vegetation response to the Younger Dryas/Holocene transition that is also evident in other records from the region (Magyari et al., 2012). Macrofossils of dwarf shrubs, such as Rhododendron kotchii and Vaccinium sp., were also present suggesting their rapid local establishment at 1910 m. R. alpinus–type and Heracleum sphondylium/H. palmatum became abundant around the lakeshore together with Rumex cf. acetosa, C. palustris, Epilobium sp. and Sedum sp. (Figure 3). This herb assemblage indicates damp ground and open dry rocks and soil. Rumex species may have favoured base-rich disturbed surfaces after ice retreat, while the presence of Sedum sp. remains points to dry habitats. The most common Sedum species is Sedum atratum in the mountain today that usually occupies scree surfaces. The high number of herbaceous remains may indicate strong discharge of the inflowing streams or a great extent of permanently saturated soils near the lakeshore because of high precipitation and water levels resulting from melting of large amounts of snow and of the discontinuous permafrost in rock glaciers next to the lake (Kern et al., 2004; Urdea, 2004; Vespremeanu-Stroe et al., 2012).
Around 9700 cal. yr BP (onset of zone LM-3, 483–371 cm), the timberline was probably very close to the elevation of Lake Lia as shown by increased macrofossil concentrations of Picea abies and by the presence of P. cembra remains. Increasing abundance of Picea abies macrofossils from 9700 to 8000 cal. yr BP indicates that the timberline was rising throughout this period. However, P. mugo, which competes well in conditions of thick snow cover and can also colonise avalanche tracks because of its high flexibility and resistance to breakage (Holtmeier, 2009), became more abundant in the local vegetation as shown by high needle concentrations and continuous presence of stomata. This suggests that the woodland was relatively open. Betula cf. pubescens fruits occurred around 8300 cal. yr BP. On the basis of its modern ecology, B. pubescens is a good coloniser because of its resistance to breakage by snow pack (Holtmeier, 2009), but it was rare in the Bucura valley.
Between 7915 and 5030 cal. yr BP (zone LM-4, 371–225 cm), Picea abies macrofossils were dominant. Conspicuously lower concentrations of P. mugo and herbs indicate that the timberline was located above the lake, which was now surrounded by closed Picea-dominated forest. Around 6000 cal. yr BP, the composition of the timberline forest changed with the increase in P. cembra.
Between 5030 and 3300 cal. yr BP (zone LM-5; 225–147 cm), Picea abies still dominated the woodland around Lake Lia, with a low abundance of P. mugo. Pinus cembra needle concentrations were higher than in the previous zone, suggesting that P. cembra became part of the Picea abies–dominated forest. The increased abundance of P. cembra suggests that the timberline was shifting downwards, or that the P. cembra population expanded at the timberline. It is possible that P. mugo was not only part of the Picea abies–P. cembra forests, but may have dominated the steep scree slope to west, which is covered by P. mugo thickets today (Figure 1d).
The uppermost zone between 3300 and 245 cal. yr BP (zone LM-6, 147 to 31 cm) was characterised by a slight increase in P. mugo needles and the near absence of Picea abies and P. cembra: one P. cembra needle at 2700 cal. yr BP and one Picea twig at 79 cm (1090 cal. yr BP) and one Picea anther at 95 cm (1550 cal. yr BP). Overall, this suggests that the timberline may have shifted below the elevation of lake at 3300 cal. yr BP. However, the continuous presence of Picea abies, P. cembra and P. mugo stomata and of P. mugo needles shows that the treeline was still located above the lake, as is the case today.
The first nearly continuous appearance of pollen of Plantago lanceolata and increased Plantago major/media and Artemisia pollen in Lake Lia were recorded from c. 2200 cal BP (Figure 5a), while Adenostyles, Achillea and Phyteuma-type pollen frequencies started to increase from 3000 cal. yr BP (Supplementary Figure 5, available online). In combination with higher Poaceae pollen percentages, these probably reflect the expansion of human activities in the mountain valleys and also at higher altitudes. However, the marked increase in Poaceae pollen (at c. 2700 cal BP; LPAZ L-7a; Figure 6a) preceded the first appearance of Plantago lanceolata and coincided with a decrease in loss-on-ignition (LOI) values suggesting that soil erosion increased after the timberline descent. The sediment accumulation rate also increased at this time (inverse of deposition time, Figure 2). A single Plantago sp. seed was found at 31 cm (~245 cal. yr BP), which could reflect grazing and/or trampling near the lake. Intensified human activity around the lake was also confirmed by the re-appearance of J. communis and Potentilla cf. P. argentea macroremains during the late-Holocene (from 1100 and 300 cal. yr BP; Figure 3).
Lake Bucura
Lake Bucura is located 130 m above Lake Lia. The macrofossil record is presented in Figure 4 and the pollen record in Figure 5b.
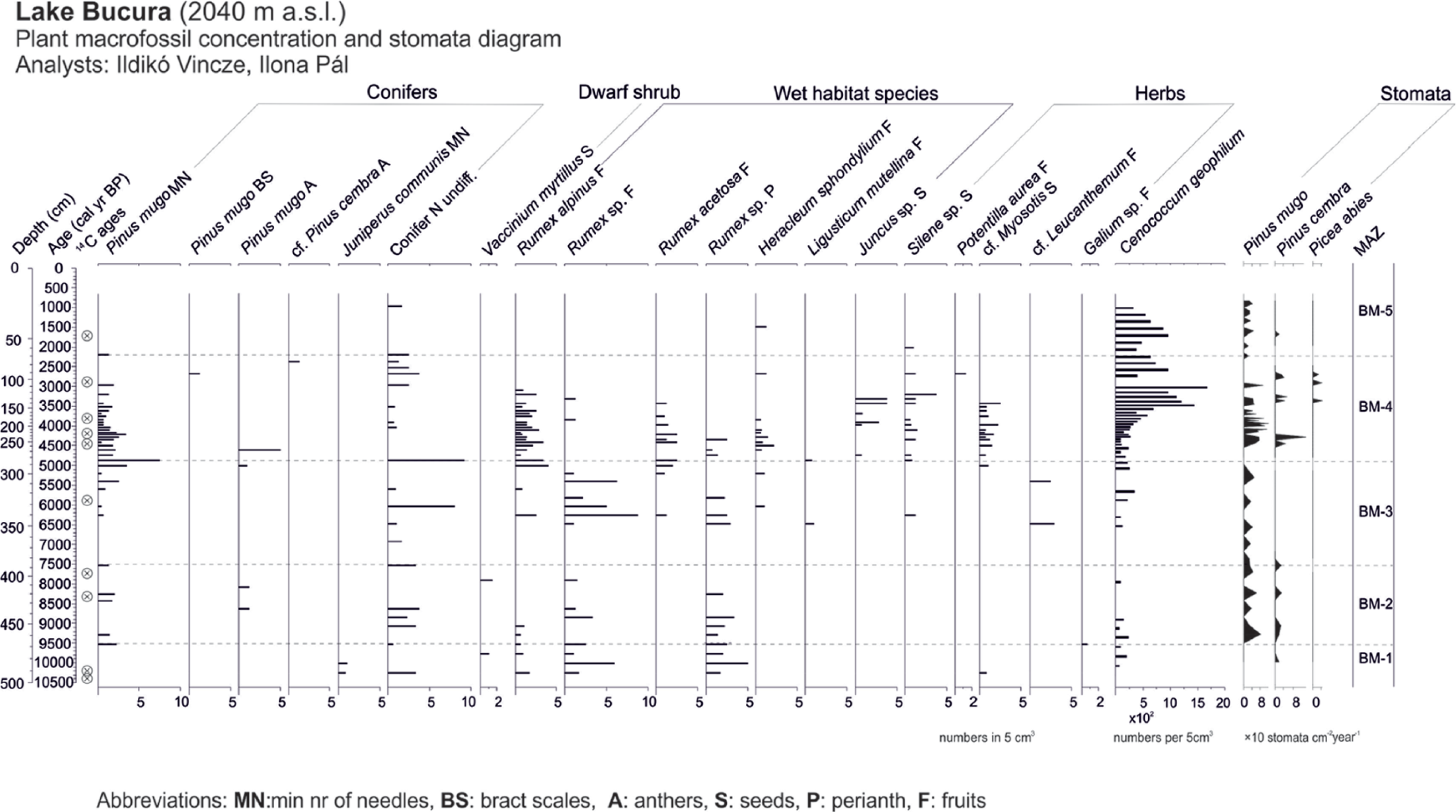
Plant macrofossil concentration diagram of Lake Bucura plotted on a calibrated age scale, together with the stomata accumulation rates. All macrofossil samples are standardised to numbers in 5 cm3 sediment. Stomata were counted along with pollen. BM zones are local macrofossil assemblage zones in Lake Bucura delimited visually.
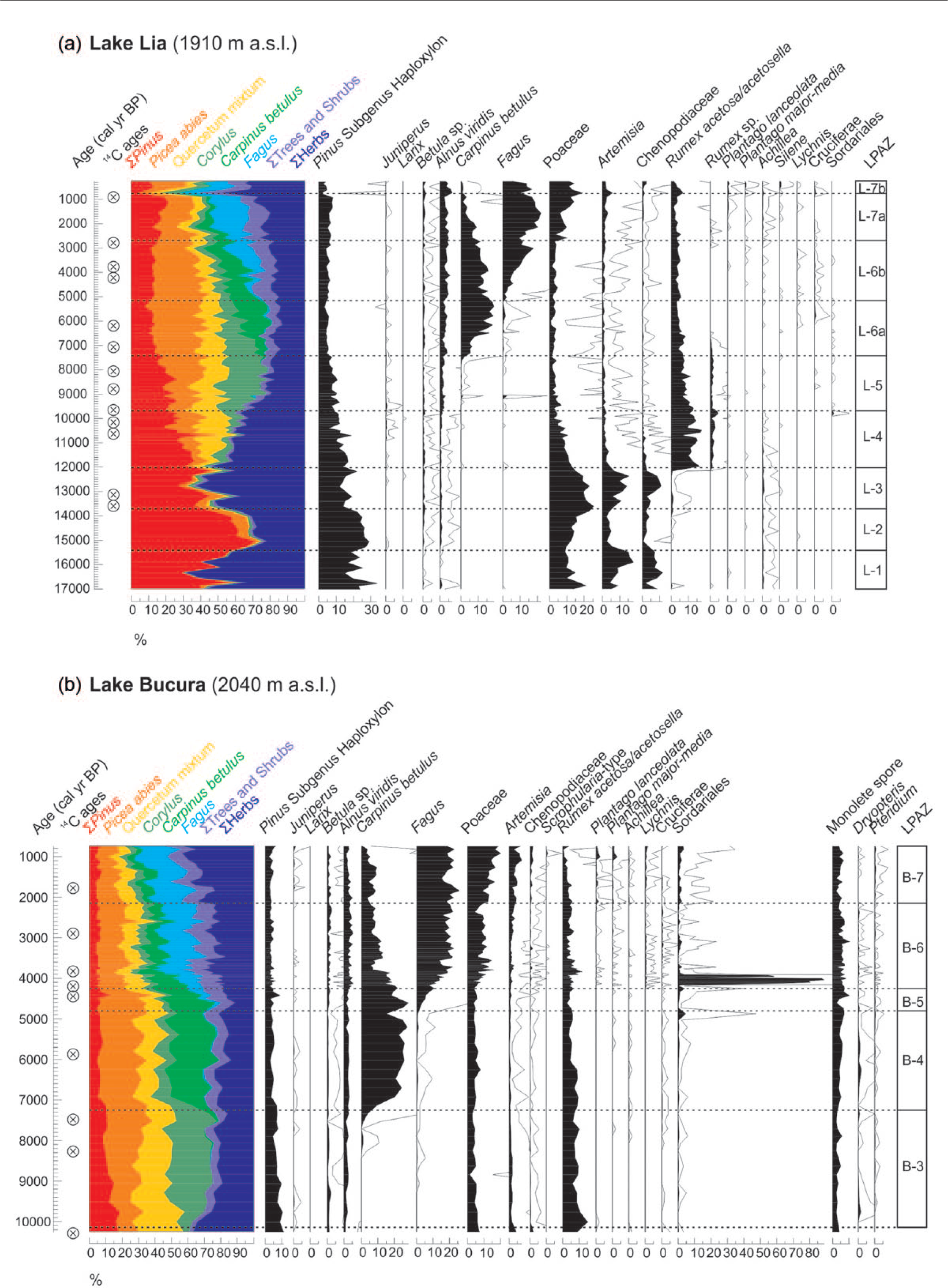
Pollen percentages of selected arboreal and herbaceous pollen types and fungal spores in (a) Lake Lia and (b) Lake Bucura. The pollen sum for calculation was at least 500 terrestrial pollens from each sample. Quercetum mixtum includes Quercus, Ulmus, Tilia and Fraxinus pollen percentages. Empty curves show ×10 exaggerations. LPAZ zones are local pollen-assemblage zones in Lake Lia and Lake Bucura. Pollen analyst: Ilona Pál.
J. communis, R. alpinus, Vaccinium myrtillus and Galium macrofossils are present in the oldest zone (BM-1: 10,300–9530 cal. yr BP, 492–468 cm). The absence of P. mugo and other tree macrofossils clearly indicates that this lake was located above the treeline at a time when Lake Lia was between the treeline and the timberline. In zone BM-2, from 9800 to 9500 cal. yr BP, P. mugo needles and stomata together with P. cembra stomata appeared in Lake Bucura regularly, indicating that the treeline was ascending and that small numbers of P. cembra trees reached Lake Bucura by 9500 cal. yr BP. Therefore, Lake Bucura was now in the krummholz zone and the treeline was not far below it. Hence, the krummholz zone reached Lake Bucura about 2000 years later than Lake Lia, which was at this time probably located below the timberline.
In zone BM-3 (7500 and 4900 cal. yr BP, 388–284 cm,) no P. mugo macrofossils were found between c. 7500 and 6300 cal. yr BP and P. cembra stomata were absent. However, unidentifiable conifer needles were present throughout the zone and P. mugo reappeared after 6300 cal. yr BP suggesting that P. mugo persisted on the lakeshore and therefore the lake was still in the krummholz zone. Rumex alpinus and R. acetosa also disappeared from the record at the beginning of the zone, but later they reappeared in high concentrations.
Zone BM-4 (4900 and 2200 cal. yr BP; 284–68 cm) was characterised by high but decreasing concentrations of P. mugo needles and stomata, the presence of P. cembra and some Picea abies stomata, probably reflecting the highest elevation of the treeline at that time. Concentrations of Cladocera ephippia and Chironomidae head capsules increased during the past 4000 years (Supplementary Figure 3, available online). The increase in Cladocera may be because of increasing nutrient availability and hence increasing phytoplankton production stimulated by disturbance in the catchment. Soil inwash is also indicated by increasing amounts of the soil mycorrhizal fungus Cenococcum geophilum (Figure 4) and gradually decreasing LOI values (Figure 7) after 4200 cal. yr BP. Pinus mugo needle concentrations gradually decreased although unidentified needle fragments were still detected until 2200 cal. yr BP. At 4200 cal. yr BP, percentages of Sordariales fungal spores (Supplementary Figure 2, available online) peaked at Lake Bucura (Figure 5b). The start of increasing percentages of Poaceae and herb pollen types, including Plantago lanceolata, Plantago major/media, Artemisia, Adenostyles, Phyteuma-type and Vaccinium-type, and the decreasing percentages of Picea pollen likely indicate the start of human impact both in the mountain valleys and at higher altitudes.
Rumex alpinus macrofossils disappeared at 3000 cal. yr BP when Lake Bucura was located above the treeline, while they persisted at Lake Lia (Figures 3 and 4) coinciding with decreasing Rumex pollen percentages at both sites (Figure 5a and b). Rumex species (mainly R. alpinus and Rumex thyrsifolius) are still abundant in the area today, and the absence of flowering and the increasingly vegetative reproduction at increasing altitude (Šťastná et al., 2012) may explain the absence of Rumex seeds in Lake Bucura. At Lake Lia, R. alpinus probably found ideal growing conditions associated with soil nitrification near animal enclosures (Šťastná et al., 2012).
After 2200 cal. yr BP (BM-5; 68 cm), the absence of P. mugo needles, of unidentified conifer needles, and the lack of tree stomata (apart from a single P. cembra stoma at 1700 cal. yr BP) are taken to indicate that the treeline continued to descend. Only Heracleum and Silene sp. seeds, fruits and a few stomata of P. mugo were still found in Bucura until 1000 cal. yr BP. The corresponding increases in Juniperus, Poaceae, Plantago lanceolata, Plantago major/media and Sedum pollen suggest more intense land use during the past 2000 years.
From 2200 cal. yr BP, recurring human impacts are detected, which likely affected the montane and subalpine forests and the alpine grassland zone. These are indicated by the relative pollen frequency changes in trees and herbs (Figure 5 and Supplementary Figure 5, available online, red arrows), especially by the gradual decrease in Picea abies pollen. The fluctuation of Carpinus betulus, Fagus sylvatica and Corylus percentages at both sites probably reflects human impact at lower elevations (Figure 5 and Supplementary Figure 5, available online, green arrows) together with increases in Poaceae, Alnus viridis, Artemisia, R. acetosa/acetosella and other herb percentages.
Fire histories
MicroCHAR (Figure 6) in both sequences indicates higher fire activities between about 10,000 and c. 8000–7000 cal. yr BP in the early-Holocene. Fire activity increased again at Lake Lia around 500 cal. yr BP. However, Finsinger et al. (2016) have shown that the increase in CHAR values around 500 cal. yr BP is probably not significant. Similarly, the high microCHAR values in Lake Bucura between 5000 and 3500 cal. yr BP may be because of a sharp change in the sediment accumulation rates (Figure 2) and sediment focusing.
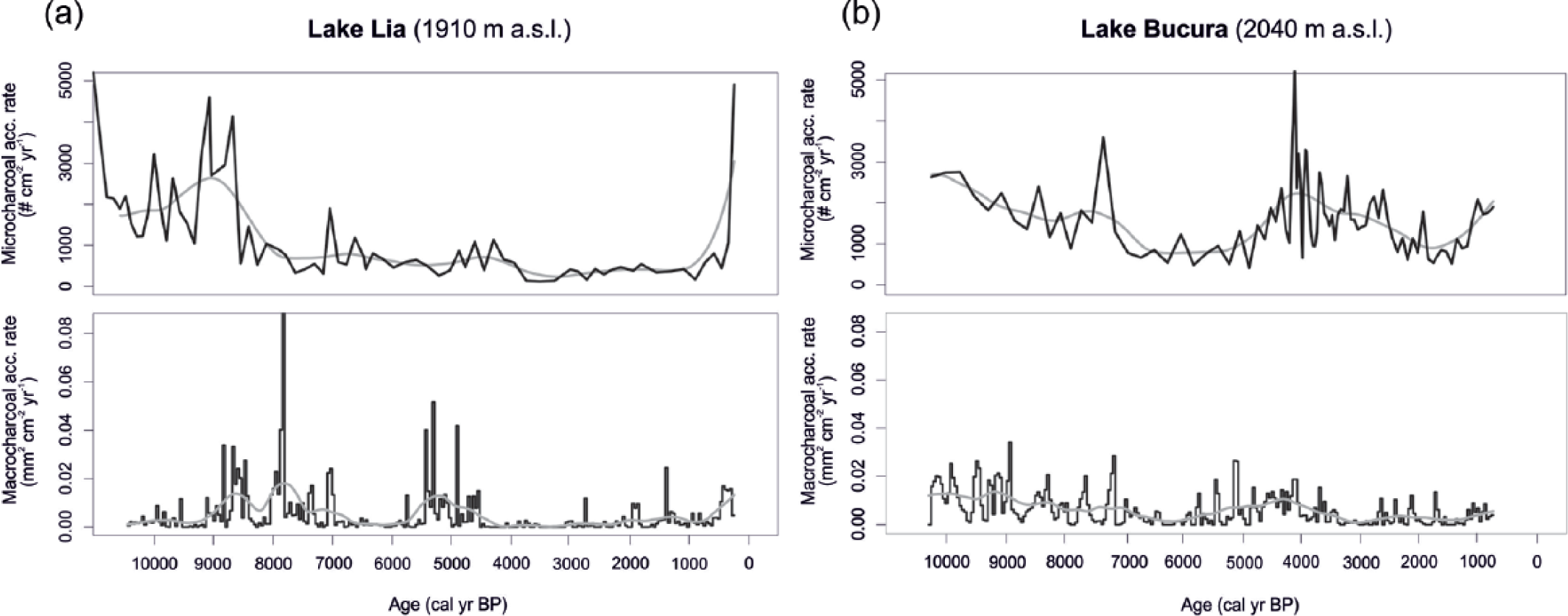
Microcharcoal (microCHAR) and macrocharcoal (macroCHAR) accumulation rates in (a) Lake Lia and (b) Lake Bucura. The black lines are raw data. Grey lines are the data smoothed by lowess (see text).
MacroCHAR (Figure 6) show higher peaks at Lake Lia than at Lake Bucura, reflecting high early-Holocene burning of the forest before c. 7000 cal. yr BP and another high biomass burning phase centred on 5000 cal. yr BP (Finsinger et al., 2016). Local fires do not seem to have been intense at Lake Bucura, probably because of the lower biomass availability or the greater distance to burnt areas. Periods of increased biomass burning as inferred from the smoothed macroCHAR record from Lake Lia occurred between 9000 and 7000 cal. yr BP, 5500 and 4500 cal. yr BP and around 500 cal. yr BP. A slight increase in macroCHAR starting from 2000 cal. yr BP could reflect a moderate increase in biomass burning. At Lake Bucura, highest macroCHAR values were recorded between 10,000 and 7000 cal. yr BP and between 6000 and 3500 cal. yr BP. However, given the likely influence of sediment focusing on microCHAR values, the latter high biomass burning period is considered an artefact. A slight increase in smoothed macroCHAR between 2700 and 1700 cal. yr BP may reflect moderately increased biomass burning at that time.
MacroCHAR records partially reflect variations in microCHAR (notably at Lake Bucura between 10,000 and 7000 cal. yr BP, at Lake Lia around 9000–8000 and 500 cal. yr BP) reflecting either a synchronicity between local and regional fire occurrences or the deposition of microcharcoal particles because of local fires.
Discussion
The position of the timberline is sensitive to climate changes (Tinner and Theurillat, 2003), because it is mainly influenced by the accumulated heat during the growing season that is closely connected to the mean temperature of the warmest month (Körner, 2012a) and land-use change (Holtmeier, 2009). Our results suggest that between 8000 and 3200 cal. yr BP, the timberline reached its highest elevation and was located above Lake Lia (1910 m a.s.l.), but below Lake Bucura (2040 m a.s.l.; Figures 7 and 8). They also suggest that Picea abies reached its highest elevation between 3500 and 3000 cal. yr BP, probably because of more favourable soil forming conditions (Henne et al., 2011). To investigate the role of summer temperature changes and human activities on the timberline and treeline, we compare the reconstructed treeline and timberline changes (Figure 8) with the charcoal and human indicator pollen proxies (Figure 7) and with the chironomid-inferred July temperature changes in Lake Brazi on the northern slope of the Retezat Mountains (Tóth et al., 2015). We ask the following questions: Was the elevated timberline position a response to higher growing-season temperatures in the mid-Holocene? Was the timberline descent at ~3000 cal. yr BP because of Holocene cooling, or did it reflect human impact?
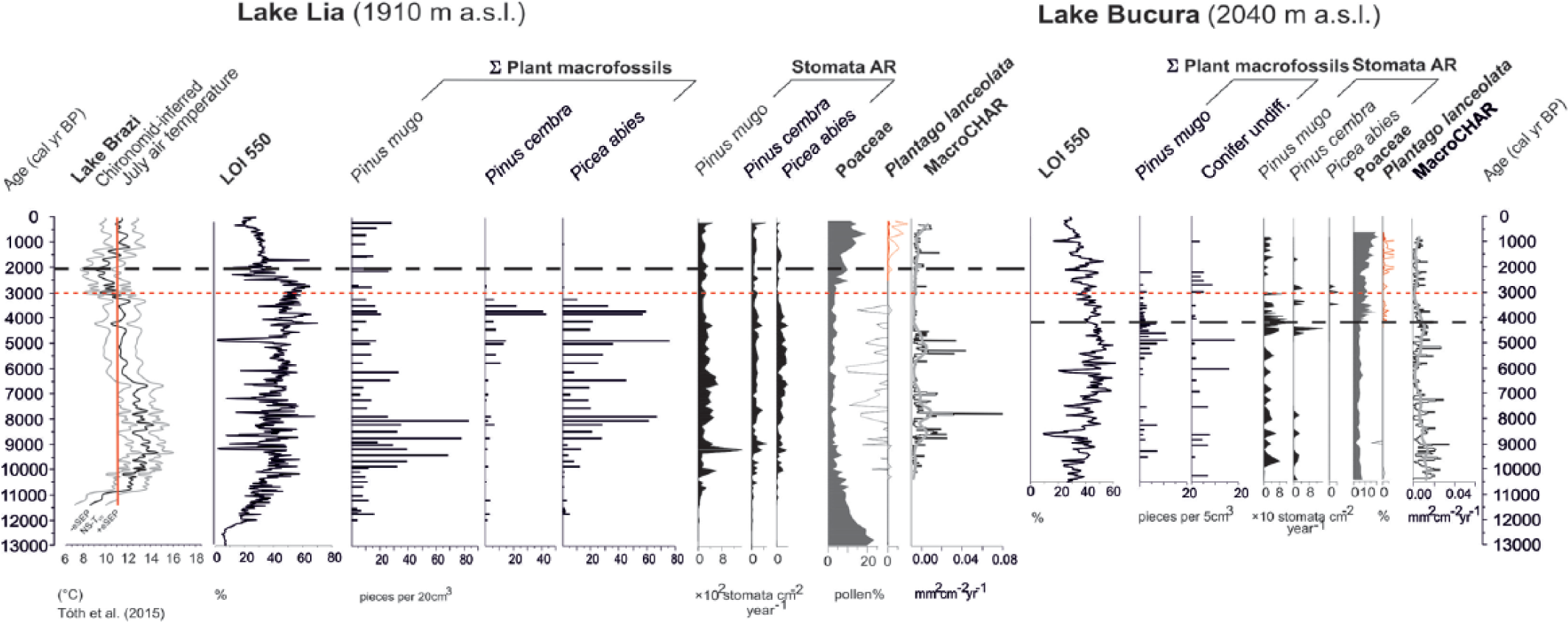
Summary diagram showing selected proxy records from Lake Lia and Lake Bucura and the chironomid-inferred mean July air temperatures ± 1 standard errors from Lake Brazi (north slope of Retezat Mountains, 1740 m a.s.l.; Tóth et al., 2015). The vertical red line indicates the present-day mean July air temperature (~11.2°C; Magyari et al., 2013). LOI 550 = loss-on-ignition at 550°C. Macrofossil records are summed fragments for the selected taxa. MacroCHAR = macrocharcoal accumulation rate. Stomata AR = stomata accumulation rate. The pollen of Plantago lanceolata is shown in red as a human indicator pollen type. Macrocharcoal analysis of Lake Lia focused on the Holocene part of the sediment sequence.
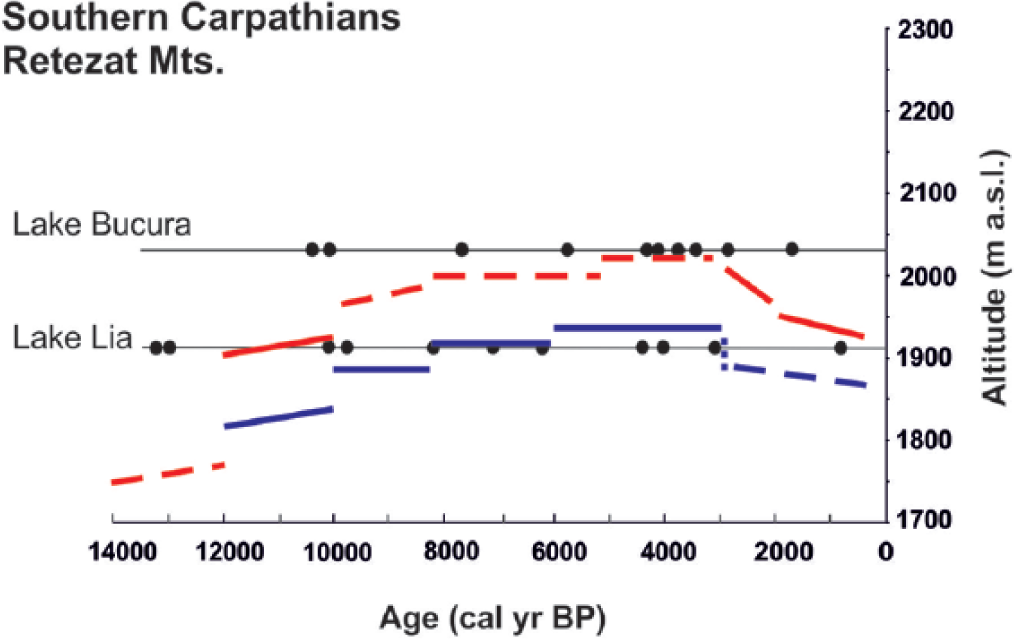
Schematic illustration of the altitudinal changes in the treeline (red line) and timberline (blue line) on the southern slope of the Retezat Mountains during the past 14,000 years. The first woody macrofossils appeared at Lake Lia around 12,000 cal. yr BP. The limits of treeline and timberline are placed between the sites recording the presence of the respective vegetation type as inferred by macrofossil and stomata analyses, following Tinner and Theurillat (2003). Dashed lines denote stronger uncertainties in the past altitudinal positions. The chronology relies on AMS dating of sediment components from Lake Lia and Lake Bucura; calibrated radiocarbon dates are shown with black dots at the elevations of the lakes.
Did summer temperature change at the time of treeline ecotone changes in the Bucura Valley?
Summer temperatures increased rapidly in the early-Holocene (Figure 7) between 11,500 and 10,200 cal. yr BP (Tóth et al., 2015). Concomitantly, treeline ascended and conifers first appeared around Lake Lia. Warmer-than-present conditions lasted until around 6300 cal. yr BP (Tóth et al., 2015), when July temperatures decreased towards present-day levels. On the basis of the Lake Brazi diatom record, this summer cooling was accompanied by rising lake levels after 6000 cal. yr BP (Buczkó et al., 2013). These temperature and hydrology changes coincided with the start of an increase in P. mugo at Lake Bucura. At Lake Lia, P. cembra became more abundant in the Picea forest after 6000 cal. yr BP. The increase in P. cembra and the continuing abundance of Picea abies suggest closed forest conditions. Pinus cembra is more frost resistant than Picea abies (Holtmeier, 2009) and is most abundant near the altitudinal timberline and in the treeline ecotone. This may suggest that the timberline was moving downwards.
The chironomid-inferred temperatures show a marked decrease to 1.8–1.9°C below modern values between 3000 and 1000 cal. yr BP (Tóth et al., 2015). The onset of this decrease broadly coincides with the timberline descent below Lake Lia and with the treeline descent further below Lake Bucura. In addition, the diatom record between 3300 and 2800 cal. yr BP characterises the highest lake level in Lake Brazi during the Holocene (Buczkó et al., 2013). Overall, summer temperatures played an important role in the macrofossil-inferred timberline upward and downward shifts during the early and mid-Holocene.
The low plant macrofossil concentrations in Lake Bucura may imply more open vegetation cover than at Lake Lia. However, taphonomic factors may have been responsible. Lake Lia is small and shallow, whereas Lake Bucura is large and deep. In addition, the bottom of Lake Bucura is covered by aquatic mosses (Supplementary Table 1, available online), which might have hindered the dispersal of terrestrial macrofossils to the coring site (see, for example, Birks et al. (2012)), which was located in the centre of the northern sub-basin. In addition, lower sediment-sample volumes in Lake Bucura (mean 5 cm3) reduce the likelihood of finding plant macrofossils in the sediments compared with Lake Lia (mean volume 21 cm3). However, the reconstructions of the timberline and treeline changes are not based on absolute differences in plant macrofossil concentrations among the sites, but on the variations in plant macrofossil concentrations within each record and the composition of the macrofossil and pollen assemblages.
What was the role of humans in the Holocene treeline and timberline changes?
Pollens from cultivated plants (e.g. Cerealia-type) were not found in the sediment of the two lakes. At Lake Bucura, Plantago lanceolata pollen first appeared at 4200 cal. yr BP and percentages increased together with increases in Plantago major/media, Poaceae, Artemisia, Adenostyles, Vaccinium-type and A. viridis pollen. The coincident increase in C. geophilum sclerotia (Figure 4), the decrease in LOI values (Figure 7, Table 2) and decreasing P. mugo needle concentrations all indicate the opening of the P. mugo thickets. This could reflect a natural, macroclimate-induced expansion of alpine meadows with the gradual recession of the treeline and timberline already by 4200 cal. yr BP. The coincident appearance and increase in secondary anthropogenic indicator pollen types, on the other hand, suggest that human impact in the form of grazing likely appeared around Lake Bucura coincidentally or shortly after the expansion of the alpine meadow areas. Soil erosion may have resulted from both natural and anthropogenic processes at this time. Regarding the geographical position of the human impact (whether lowland piedmont or high mountain origin), it is important that no comparable changes were found at Lake Lia around 4200 cal. yr BP. Here, the montane Picea forest persisted until at least 3200 cal. yr BP and increases Plantago major/media, Plantago lanceolata and Poaceae pollen percentages occurred later (Supplementary Figure 5, available online, Figures 5 and 7, Table 2). This difference in timing is probably real, because the increase in F. sylvatica percentages occurred at about the same time in both records (around 5000 cal. yr BP: the onset of LPAZ L-6b and B-5; Figure 5) implying that the age–depth models are correct.
Indications of human impact (age, cal. yr BP) in Lake Lia and Lake Bucura. The different proxy records are considered to be indicators of human impact, when accompanied by major changes in the pollen record. Secondary anthropogenic indicator pollen types (Plantago lanceolata, Plantago major/media, Poaceae, Achillea, Artemisia), tree species from lower altitudes (Fagus, Carpinus betulus), Picea abies and Pinus pollen percentage changes were considered as indicators of human impact (see Supplementary Figure 5, available online). Note: +: the proxy likely supports human impact; −: the proxy does not support human impact; 0: no data. The synthetised consequence of human impact is based on any combination of the previously mentioned comparable proxies from both lakes.
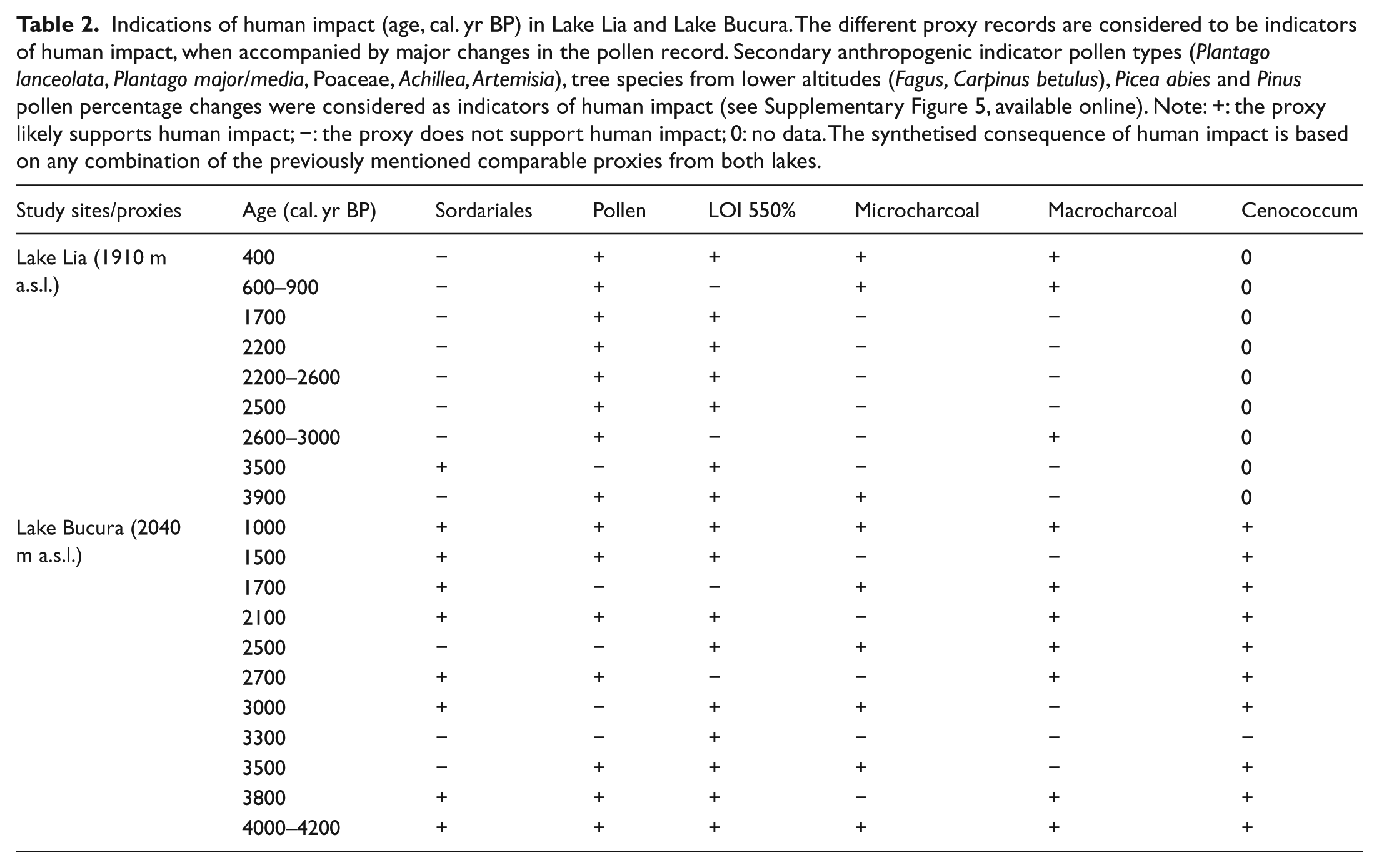
One possible explanation for the earlier detection of human impact indicators at the higher lake might be the smaller representation of regional pollen rain in the lower lake, which was surrounded by denser tree cover than the upper lake (Pop et al., 1965). Surface-pollen studies in the Retezat suggest that the increase in Poaceae pollen and associated herbaceous pollen types is a characteristic feature of Lake Bucura that is surrounded by extensive alpine meadows even today. The pollen frequencies of Poaceae far exceed the piedmont values (Pop et al., 1965). The simultaneous increase in Poaceae, Plantago lanceolata, Plantago major/media, Phyteuma-type, Adenostyles and Vaccinium-type pollen thus more likely indicate that the naturally open habitats in the alpine zone and treeline ecotone were the first to be utilised for seasonal pastural farming during the Bronze Age.
Our systematic archaeological data survey suggests that Bronze Age settlements were present in the lowland and piedmont zones (up to 750 m a.s.l.) of the Hunedoara and Gorj counties, located north and south of the Retezat Mountains (Supplementary Figure 6, available online), but there is no evidence for occupancy of strictly alpine situations. Grazing may have contributed to a decrease in the P. mugo population around Lake Bucura at 4200 cal BP, but similarly low macrofossil concentrations were found in the mid-Holocene when human impact was probably negligible. To disentangle whether the percentage peak of Sordariales-type spores in Lake Bucura between 4200 and 3800 cal. yr BP was derived from decaying wood or dung, we examined samples of both substrates from the Retezat Mountains (goat, sheep, cow, bear faeces and decaying Picea abies and P. mugo wood). However, none of these samples contained Sordariales-type spores (Supplementary Figure 2, available online). Sordariales-type spores showed a prominent increase in the sediments of Lake St Anne (Eastern Carpathians, Ciomadul Mountains) at 3300 cal. yr BP (Magyari et al., 2009b). In that case the Sordariales increase was not related to grazing but to rottening waterlogged wood because of a lake-level rise (Magyari et al., 2009b). In addition, dung fungal spores (e.g. Sporormiella; Baker et al., 2013), were not found at Lake Bucura and Lake Lia. Hence, we conclude that Sordariales-type spores likely indicate seasonal water-level fluctuation (shore inundation followed by low water level) and related fungal growth on waterlogged wood between 4200 and 3800 cal. yr BP at Lake Bucura. On the other hand, other lines of evidence, such as the first increase and first appearances of Plantago lanceolata and Plantago major/media, Adenostyles and Phyteuma-type pollen, the prominent increase in Poaceae pollen and the start of the increase in Cenococcum together with the decrease in LOI values suggest the start of human impact at this time.
Our pollen data suggest that human impact likely intensified from ~2200 yr cal BP at both sites. Increasing Poaceae and anthropogenic indicator herb pollen percentages coincide at Lake Bucura with the disappearance of P. mugo macrofossils, suggesting that the local extirpation of P. mugo was likely human-induced. This inference is also supported by the macrocharcoal results that suggest that episodic local fires continued after 2600 cal. yr BP around Lake Bucura (Figures 6 and 7). The late-Holocene fires, possibly man-made, could have played an important role in the opening up of the alpine krummholz zone in the past 2600 years (Finsinger et al., 2016). The intensification of grazing around Lake Bucura, suggested by increases in Poaceae, Plantago lanceolata, Plantago major-media, Sedum and Juniperus pollen frequencies after 2000 cal. yr BP (Figure 5b), can be attributed to Iron Age cultures (Luca et al., 2005).
At Lake Lia, the first human impact may have started at 2600 cal. yr BP, about the time of intensified human impact around Lake Bucura. The expansion of pastures around Lake Lia is indicated by increases in Poaceae pollen and anthropogenic indicator herbs (Table 2, Figures 5a and 7, Supplementary Figure 5, available online) and lower LOI values reflecting increased soil erosion. The plant macrofossil–inferred timberline decrease preceded these changes in pollen percentages by ~400 years. This implies that the timberline descent was likely not directly human-induced, but occurred in response to decreasing summer temperatures (Tóth et al., 2015). MacroCHAR showed a significant fire episode ~50 years earlier, at 2650 cal. yr BP (Figure 7; Finsinger et al., 2016) suggesting that the forest burnt at least once in this period. We also infer that without continuous grazing and burning, scattered trees would probably still grow around Lake Lia. The survival of Picea abies and P. cembra trees not far from the lake is attested by the stomata record that shows decreases at 2600 cal. yr BP, but increases afterwards (Figures 3, 4 and 7). The intensified human impact since 2200 cal. yr BP did not affect the overall abundance of P. mugo as the macrofossil record indicates its persistence around the lake, probably on the steep, rocky western slope that is unsuitable for grazing.
At about 1000 cal. yr BP, the peaks in pollen percentages of Artemisia, R. acetosa/acetosella, Plantago lanceolata, A. viridis and other herbs decrease in LOI in both sites, and the decreases in Pinus diploxylon–type and Picea abies pollen at Lake Lia suggest a short-term intensification of clearance of alpine habitats (Table 2, Figure 5a).
Comparison of the Holocene timberline and treeline history in the Alps, the Carpathians and the Balkans
The early-Holocene species composition of the treeline on the southern slopes of the Retezat Mountains was characterised by Picea abies together with P. cembra (Figures 3 and 4). These tree species also expanded in the early-Holocene in the Rodna Mountains (Eastern Romanian Carpathians), but their expansion started later, around 9800 cal. yr BP (Geanta et al., 2014). In the Central Alps, Picea expanded much later than in the Southern and Eastern Carpathians, at ~5000 cal. yr BP (Tinner et al., 1996). The timberline forests in the Central Alps were dominated by L. decidua and P. cembra at that time, while L. decidua was replaced by P. mugo at high altitudes in the Retezat in the early-Holocene (Magyari et al., 2012). These differences may reflect different climatic characteristics in the Southern Carpathians that favoured the early expansion and niche acquisition of Picea abies (Holtmeier, 2009).
Plant macrofossil records from the two lakes show that the maximum timberline elevation between 8000 and 3000 cal. yr BP was about 100 m higher than today in the Retezat Mountains. The maximum treeline position was also about 100 m higher than today (i.e. 2000–2050 m). By comparison, plant macrofossil studies in the Central Swiss Alps showed that the maximum elevation of the timberline occurred between 10,000 and 6000 cal. yr BP at Gouillé Rion (2343 m a.s.l., Central Swiss Alps, Figure 1c) and Lago Basso (2250 m a.s.l., North Italian Alps, Figure 1c; Wick and Tinner, 1997) and between 8500 and 4000 cal. yr BP at Lac de Fully (2135 m a.s.l., Figure 1c; Finsinger and Tinner, 2007). The Lac de Fully record is most similar to the Retezat Mountains. The later advance of the timberline in the early-Holocene could be because of local factors such as local dryness, topography and local fire events, interacting with the regional climate (Orbán et al., 2017).
In the Central Alps, the timberline progressively descended by about 180 m from 6000 to 5000 cal. yr BP (Tinner, 2007; Tinner and Theurillat, 2003). By comparison, on the southern slope of the Retezat, an increase in P. cembra at the timberline occurred suggesting timberline descent (Figure 3), but the major descent of the timberline commenced only around 3000–3500 cal. yr BP (Figure 8), later than in the Central Alps. This descent was clearly connected to July temperature decrease in the Retezat Mountains (Figure 7), while in the Alps the regression of the timberline was primarily caused by human impact (Tinner, 2007).
The intensity of human activities was less in the Retezat Mountains than in the Alps and in the Eastern Carpathians (Feurdean et al., 2016; Tinner, 2007; Tinner and Theurillat, 2003). Our data show no evidence for the influence of Neolithic cereal cultivation, and Plantago lanceolata–type pollen was detected only after 4200 cal. yr BP. This is despite the clear presence of Neolithic settlements in the surrounding lowland and piedmont zones (Supplementary Figure 6, available online). It is possible that Neolithic people did not use the high mountain pastures for grazing in the Retezat Mountains. In contrast, human indicator pollen types in pollen diagrams of the Alps from c. 8650 cal. yr BP show that Mesolithic and Neolithic cultures started to exploit these mountains (Lotter, 1999; Tinner et al., 2007). Tinner (2007) points out that the regression of the timberline during the past 5000 years was primarily caused by human activities. Similarly, late-Neolithic livestock grazing was demonstrated in the subalpine belt of the Rodna Mountains in the eastern Romanian Carpathians (Feurdean et al., 2016).
In the Bronze Age, grazing activity does not seem to have resulted in the lowering of the timberline in the Retezat Mountains, in contrast to the Alps. Our data suggest that grazing first focused on the alpine meadows and the naturally semi-open krummholz zone in the upper Bucura valley. The first shepherds contributed to the thinning of the P. mugo thickets and some tree cutting in the treeline ecotone. The timberline forests were not affected until later, when intensified human impact in the Iron Age and Middle Ages led to a considerable lowering of the timberline, as also attested partly by historical documents (Maderspach, 1868; Téglás, 1888a, 1888b). Thus, the treeline ecotone was extended, and the timberline is lower today than its potential elevation in the Retezat Mountains.
The treeline reached higher altitudes in the Balkan Mountains than in the Retezat during the early-Holocene probably because of higher growing-season temperatures. Macrofossil records suggest that treeline reached 2250 m in the Pirin Mountains during the late-glacial and was formed by Betula, Juniperus and Pinus peuce in the Southern Balkans (Atanassova and Stefanova, 2003; Stefanova et al., 2006; Tonkov et al., 2006, 2011). This species composition persisted during a time of high summer insolation, relatively warm temperatures and increased aridity until 6700 cal. yr BP when Pinus sylvestris, P. peuce and A. alba expanded in the treeline ecotone suggesting increasing moisture availability in the second half of the Holocene (Tonkov and Marinova, 2005). The altitudinal extent of the treeline ecotone appears to have been much larger in the Southern Balkans. The little available information suggest that the closed forest limit ascended around 6700 cal. yr BP (Tonkov and Marinova, 2005). This mid-Holocene upward shift is not detected in the Retezat Mountains. The P. cembra increase around Lake Lia (Figure 3) in response to cooling postdates this event by c. 700 years suggesting regional climatic differentiation caused vegetation responses at different times in these mountains.
Conclusion
The plant macrofossil, pollen, charcoal and LOI analyses from the two study sites on the southern slope of the Retezat Mountains provide the first records of treeline and timberline variability in this region and contribute to an improved understanding of the long-term impact of climate and land-use changes on these high-altitude ecosystems. The study confirms that the timberline and treeline were higher in the past, as hypothesised from documentary evidence (Mertens and Huband, 2004). Both the treeline and the timberline reached maximum elevations between 8000 and 3200 cal. yr BP and their altitudinal fluctuations were in the order of 100–150 m. The vegetation around 2040 m a.s.l was dominated by P. mugo during the mid-Holocene, whereas at 1910 m a.s.l. closed mixed P. cembra and Picea abies forest developed. The pollen and macrofossil evidence that human impact likely became important after 4200 cal. yr BP above 2000 m a.s.l. in the Bronze Age, further back in the past than documentary evidence is able to show. It probably intensified in the treeline ecotone (c. 1900 m a.s.l. at that time) during the Iron Age and Medieval Period, around 2600 cal. yr BP. The alpine meadows dominated by Nardus stricta and Festuca spp. today developed under continuous grazing on the southern slope over the past 2200–2600 years.
On the basis of our results, we conclude that future changes in alpine and treeline vegetation in the Retezat Mountains will be a function of the amplitude and rapidity of climate change combined with forest and land-use management practices. Global warming with decreasing land use of the high-elevation ecosystems would certainly lead to the upslope movement of both Picea abies and P. cembra. However, compared with the Alps, the altitudinal increase in the treeline would be expected to be less, in the order of 100–150 m. Changing land use with reduced grazing intensity and increased recreational use may also affect the vegetation of the Bucura valley at lower elevations, but further research is needed to provide an overview for the whole region.
Footnotes
Acknowledgements
The authors are grateful to Jordan Fevre for analysing the macrocharcoal from Lake Lia. They appreciate the archaeological information from Hunedoara county (Romania) from Dr IV Ferencz (Muzeul Civilizatiei Dacice si Romane Deva, Department of Archaeology, Deva). They are grateful to HJB Birks for helpful comments. They also thank the Retezat National Park support and the people who helped them in the field (Gergely Boros, Krisztina Buczkó, Péter Eszenyi, Edit Hathi, Tamás János Juhász, István Konyhás, Barnabás Körmöndi, Máté Mile, Judit Nédli, István Papp, Gellért Puskás, Csaba Schnitchen, László Somay, Szabolcs Struba, Andor Umman, Gábor Umman, Hajnalka Veres). IV, EKM and GJ did the plant macrofossil and macrocharcoal analysis of Lake Bucura; IO and HHB did the macrofossil analysis of Lake Lia and IO calculated the age–depth models of both sediment sequences. IP did the pollen, stomata and microcharcoal analyses of Lake Lia and Lake Bucura. LOI measurement was done by KH on both sediment cores. MB and EKM carried out the sediment coring. WF contributed to the macrocharcoal data analyses. TB drew the 3D model of the study site in ![]() . CD provided valuable information about the study site. IV, EKM, IP, WF and HHB wrote the manuscript. The project that financed this research was led by EKM. The authors thank the two anonymous reviewers for their constructive comments. All authors read and approved the final manuscript.
. CD provided valuable information about the study site. IV, EKM, IP, WF and HHB wrote the manuscript. The project that financed this research was led by EKM. The authors thank the two anonymous reviewers for their constructive comments. All authors read and approved the final manuscript.
Funding
This study was supported by the Hungarian National Scientific Fund (OTKA NF 101362), the Humboldt fellowship to EKM and was financially supported by GINOP-2.3.2-15-2016-00019. This study was also supported by a short-term scientific mission provided by the INTIMATE COST Action ES0907 awarded to IV and the University of Bergen, Norway, to IO. This is MTA-MTM-ELTE Paleo contribution no. 231.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.