
Abstract
This paper presents the first pedoanthracological study carried out on two mountains of the Northern Apennines, Monte Cimone, and Corno alle Scale, where the results provided new palaeoenvironmental data. The pedoantracological sampling followed an elevation gradient from the current timberline to the highest possible elevation, also adapted to the geomorphological characteristics of the study areas. Based on radiocarbon datings (16 at Monte Cimone and 9 at Corno alle Scale), the soil charcoal fragments provided data about the vegetation dynamics and plant landscape reconstruction, primarily indicating the Late Holocene. The landscape over the last 3000 years appears very similar to the current one with very small altitudinal variations of the timberline and treeline. The present study shows a lack of radiocarbon dates for the Middle Holocene and two hypotheses are discussed to explain this finding. Data suggest that past fires were locally linked to periods of climate optimum and possibly with the management of natural resources (especially animal grazing) by human societies. Some questions concerning the absence of Abies alba, today only present as plantation and normally associated with Fagus sylvatica at these latitudes, are discussed.
Introduction
The current plant landscape results from the interaction of ecological processes and social practices under the influence of the various climatic phases that occurred during the Holocene (e.g. Butzer, 2012; Mercuri et al., 2011). This definition is established for the mountain landscape thanks to numerous multidisciplinary studies (e.g. Bal et al., 2015; Bonanomi et al., 2018; Cevasco, 2004; Cevasco and Poggi, 2000; Davasse et al., 1997, 2006; 2016; Davasse and Galop, 1990a, 1990b; Métailié, 1986; Moreno and Montanari, 2008). This article aims to improve the reconstruction of the plant landscape of the Northern Apennines and to study the relationships between human societies and their environment during the Late Holocene based on pedoanthracological data.
The plant landscape of the Northern Apennines has been investigated by palaeoenvironmental and palaeoecological studies, mainly pollen analyses (e.g. Bertolani Marchetti et al., 1994; Branch, 2004, 2013; Cruise, 1990a, 1990b; Guido et al., 2013; Lowe, 1992; Molinari and Montanari, 2018; Vescovi et al., 2010a, 2010b; Watson, 1996), and by some studies on the archaeology of environmental resources located mainly in the Ligurian Northern Apennines (e.g. Cevasco and Parola, 2013; Cevasco and Poggi, 2000; Molinari and Montanari, 2018; Moreno and Montanari, 2008; Stagno, 2016). However, the Northern Apennines still lack high-spatial resolution palaeoecological research, in particular for the Tuscan-Emilian area. In the high mountains of the Northern Apennines, palaeobotanical data are rarely associated with the human action, because the archeological studies and findings are less numerous than those of the plain (Cardarelli and Malnati, 2006). Because of the limited number of human settlements in the mountain area, archeological research has mainly involved surface findings while there are rare excavated archeological sites (Cardarelli and Malnati, 2006; Ferrari et al., 2006; Fontana, 1997). The lack of a traditional archeology approach, largely studying artifacts, can be overcome thanks to environmental archeology, widely applied in Europe but little in Italy – pedoanthracology falls into this area and is suitable for studying contexts apparently devoid of ‘archeological traces’ as they are traditionally considered (e.g. Branch, 2004; Maggi, 1997; Maggi and De Pascale, 2011).
Within the Northern Apennines, we decided to study Monte Cimone and Corno alle Scale that are key mountains for several reasons. Thanks to historical sources we know that both are characterized by strong sylvo-pastoral traditions, which were maintained until the second half of the 20th century, when tourism began to prevail over agro-sylvo-pastoral activities. Human action, both for agro-sylvo-pastoral and tourism needs, has strongly modified these mountain environments by reducing woodlands (Albani, 1964; Alessandrini et al., 2010, 2003; Benatti et al., 2018; Fontana, 1997; Rombaldi and Cenci, 2013; Tomaselli et al., 1996). Moreover, Monte Cimone and Corno alle Scale are two of the highest mountains of the Northern Apennines and their treeline is lower than the potential ecological limit of beech trees at this latitude (Pignatti, 1982) and in other Northern Apennine sites (1825 m a.s.l. – Pezzi et al., 2008). This led us to investigate the causes and timing of this limit lowering. In particular, we decided to apply pedoanthracology, which is the study of charcoal fragments contained in soils and produced by natural and/or anthropogenic fires, introduced by Thinon (1992), to obtain palaeoenvironmental data at high spatial resolution (Carcaillet and Thinon, 1996; Talon et al., 1998). The use of this method is expected to provide new information about the known history of these mountains and the Northern Apennines in general, improving knowledge on the relationships between sylvo-pastoral societies and their territory. The focus of the paper is on the following dynamics observed at local scale: the evolution of timberline and treeline limits over time; the past forest composition and the vegetation dynamics (at slope scale); the analysis of fire events over time, and the use of plant resources of the mountains by the sylvo-pastoral societies of the past, showing the connections between fire history and vegetation development (e.g. Branch, 2004; Maggi, 1997; Maggi and De Pascale, 2011).
Pastoralism in the Tuscan-Emilian Apennines from historical sources
Pastoralism is one of the most practiced activities in the high mountains and has had a great influence on the development of the plant landscape of the mountains. We know after historical sources that the pastoral activity in the Tuscan-Emilian Apennines (where the study areas are located), as well as that in the Alpine regions (Carrer, 2012), had a lower occurrence compared to the Central and South Italian regions, such as Abruzzo, Apulia, Sardinia (Cazzola, 1993), and Basilicata (Florenzano, 2019). Despite this, it was nevertheless very important for the economy of the region, particularly for the mountain area (Borri, 2014; Cazzola, 1993). The research on historical and ethnographic sources has shown the importance of these two mountains, Monte Cimone and Corno alle Scale, for pastoral activities during the last centuries. The apogee of sheep and goat breeding in our study area coincides with the mid-19th century, when the number of sheep and goats was the highest (Roncaglia, 1850). In the late 19th/early 20th century, there was the first serious crisis in pastoral activity (Antilopi, 1989; Mordini and Mordini, 1994; Roversi, 1983). After the mid-20th century, there was a second intense crisis of pastoral activity that continues until today (Albani, 1964).
Ethnographic research demonstrates that the ancient tradition of transhumance has had a very important role in the study areas (Borri, 2014; Cazzola, 1993; Franceschi, 2004; Marcaccini and Calzolai, 2003; Zagnoni, 2004). As shepherds descended from the Apennine mountains with their flocks, they used to burn grasslands and mountain heath (including Vaccinium shrubs) in order to regenerate the pastures (Alessandrini et al., 2003). Today it is no longer possible to carry out a transhumance exclusively on foot due to the profound territorial and social changes that have occurred in the area over the last 50 years. For this reason, it is important to preserve the historical memory and so in March 2018, Italy, together with Greece and Austria, applied to UNESCO for transhumance to be considered an ‘Intangible cultural heritage of humanity’ (La Repubblica, 2018).
Materials and methods
Study area
Monte Cimone (44°11’36”N, 10°42’00”E) and Monte Corno alle Scale (44°07’23”N, 10°49’50”E) (Figure 1) are two of the highest peaks of the Northern Apennines (2165 m a.s.l. and 1945 m a.s.l., respectively) and are c.13 km apart. Mean annual temperatures are about 2°C and mean annual precipitation ranges from about 690 mm (registered at the Monte Cimone meteorological station) to 2500 mm (registered in other mountain stations at lower elevation) (Alessandrini et al., 2010; Şerban et al., 2007; Tositti et al., 2014). Geologically, this area is mainly characterized by sandstone and clay outcrops (Bettelli et al., 1989; Tellini et al., 1988). There is also evidence of glacial and periglacial landforms (Tomaselli et al., 1996).
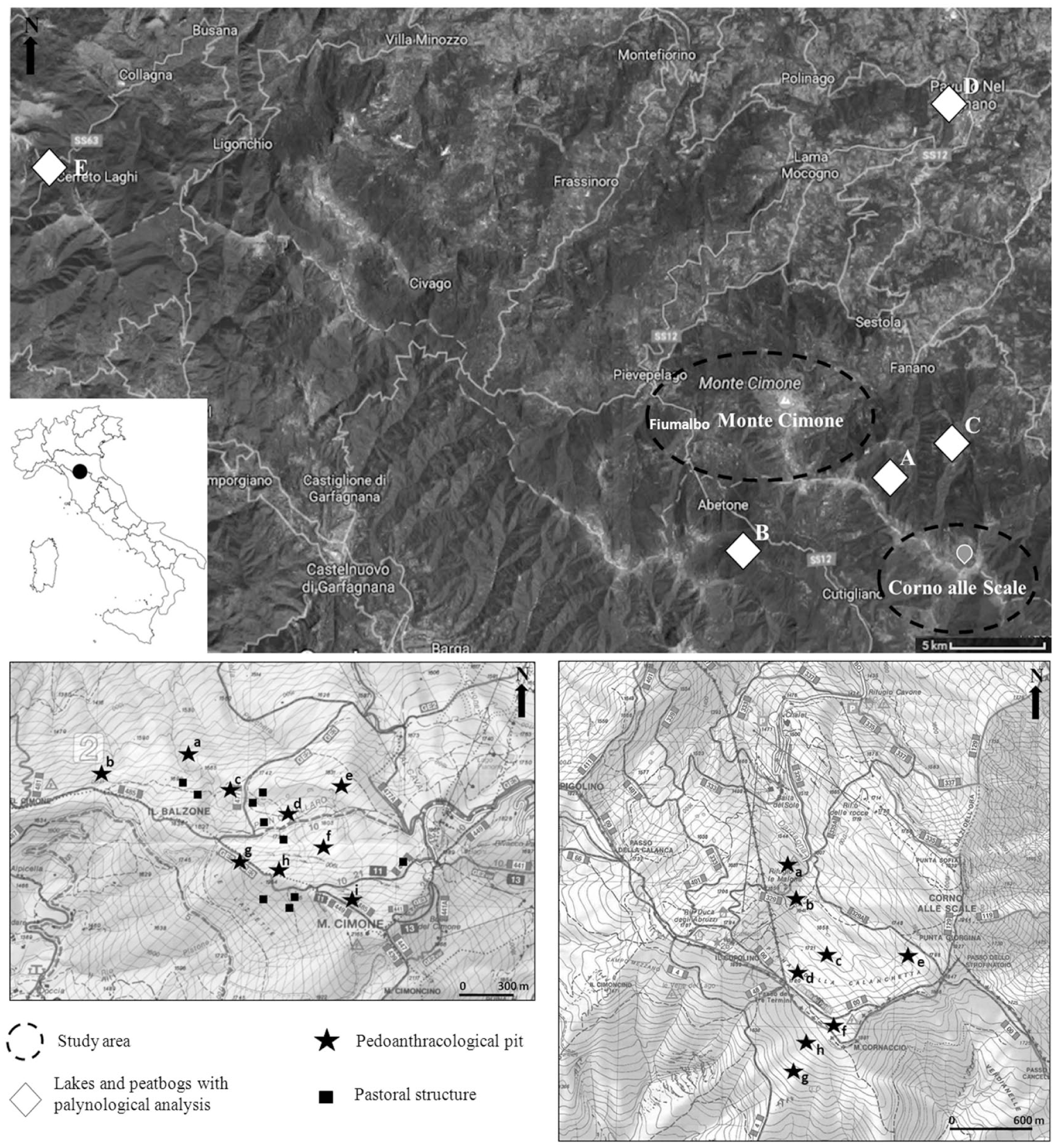
Location of Monte Cimone and Corno alle Scale study areas with the localization of the pedoanthracological sampling (pits), charcoal platforms, probable pastoral structures, and the closest and most recent pollen analysis of the area carried out in lakes or peatbogs (Vescovi et al., 2010a, 2010b; Watson, 1996)
In the study areas, the vegetation is composed of forest systems of Fagus sylvatica L., once widely coppiced and now left to natural wood development, with some occurrence of Acer pseudoplatanus L., Laburnum alpinum (Mill.) Bertcht. & J. Presl, and Abies alba Mill. (Tomaselli et al., 1996), from about 900 m a.s.l. to the timberline. A. alba is currently known only as cultivated and naturalized by plantations (Alessandrini et al., 2010) and it has not been detected near our sampling points. The current timberline reaches 1700 m a.s.l. at Monte Cimone and 1600 m a.s.l. at Corno alle Scale. Within the forest, close to the timberline, there are shrub communities with predominance of Ericaceae (dominated by Vaccinium myrtillus L.) or Fabaceae (e.g. Cytisus scoparius (L.)Link. and Genista tinctoria L.) (Alessandrini et al., 2010). In the forest system, there are also meadow-pastures dominated by Nardus stricta L. and Brachypodium genuense (DC.) Roem&Schult. (Alessandrini et al., 2010). In the mid-19th century, the beech climate zone underwent to a reforestation with conifers with the aim of safeguarding the soil in areas degraded by erosion. Reforestation was carried out with native species of the study area like Picea abies (L.)H.Kast. and Abies alba, and other species like Pinus nigra Arnold, P. mugo Turra and Larix decidua Mill. Below the beech belt, there are mesophilic woods dominated by Ostrya carpinifolia Scop., Quercus cerris L. and Castanea sativa Mill. Above the timberline, there are plant communities typical of cliff or debris areas, grasslands (Trifolium thalii Vill., Nardus stricta, Brachypodium genuense), and mountain heathland (Vaccinium myrtillus and V. gaultherioides Bijelow) (Alessandrini et al., 2003; Tomaselli et al., 1996).
According to historical and ethnographical sources, these mountains were very important for pasture and charcoal production (Albani, 1964; Rombaldi and Cenci, 2013). The presence of hypothetical pastoral structures (georeferenced but not dated – Figure 1) and charcoal platforms, still in activity in the last century, highlights the long history of using the natural resources of these areas (Benatti et al., 2018). These two mountains are located in two regional parks, Parco del Frignano and Parco del Corno alle Scale, where there are sites important for their biodiversity and protected within the Natura 2000 network. In fact, this territory has a rich and diversified floristic heritage with many endemisms due to its geographical position between two great phytogeographic regions: the Euro-Siberian and the Mediterranean (Tomaselli et al., 1996). Today, on these mountains, there are many recent human structures and buildings, such as ski slopes for winter sports.
Sampling strategy
In order to carry out the pedoanthracological study in the area of Monte Cimone we selected the north-west slope and in the area of Corno alle Scale we selected two zones: the north side (west slope of Monte Corno alle Scale and north slope of Monte Cornaccio) and the south side (south slope of Monte Cornaccio) (Figure 1). These slopes, in fact, are suitable for pedoanthracological sampling: they have large plateaus that were used as pastures in the past and are still used today; their good herbaceous cover reduces erosion and their timberline stops sharply lower than the natural ecological limit. The other slopes are very steep and rocky with numerous ski lifts and ski slopes.
Pedoanthracological soil pits were excavated following an altitudinal transect from the beech forest to the open landscape, suitable to identify timberline and treeline variations in the past. Meanwhile, on the large pastoral plateaus the sampling was carried out to verify whether the cause of vegetation dynamics might be identified as pastoral pressure. The pits were dug far from archeological elements (e.g. charcoal platforms and pastoral structures) to obtain samples from wild (or not directly human-influenced) contexts.
Pits were labeled as ‘MtC’ (Monte Cimone) and ‘CaS’ (Corno alle Scale) followed by a letter; the letters correspond to the altitude of the sampling points (a indicates the lowest pit for each study area). A total of nine pits were excavated on the north-west slope of Monte Cimone from 1650 to 2078 m a.s.l. Seven pits were located in the top belt area currently covered by open landscapes, while two pits (MtC-a and MtC-b) were just below the present timberline (Figure 1). A total of eight pits were dug in the Corno alle Scale area: six on the north side from 1602 to 1860 m a.s.l. and two others in the south side at 1700 and 1750 m a.s.l. All pits were located in the current open landscape; CaS-a was located just above the current timberline (Figure 1).
Soil charcoal analysis
Sampling
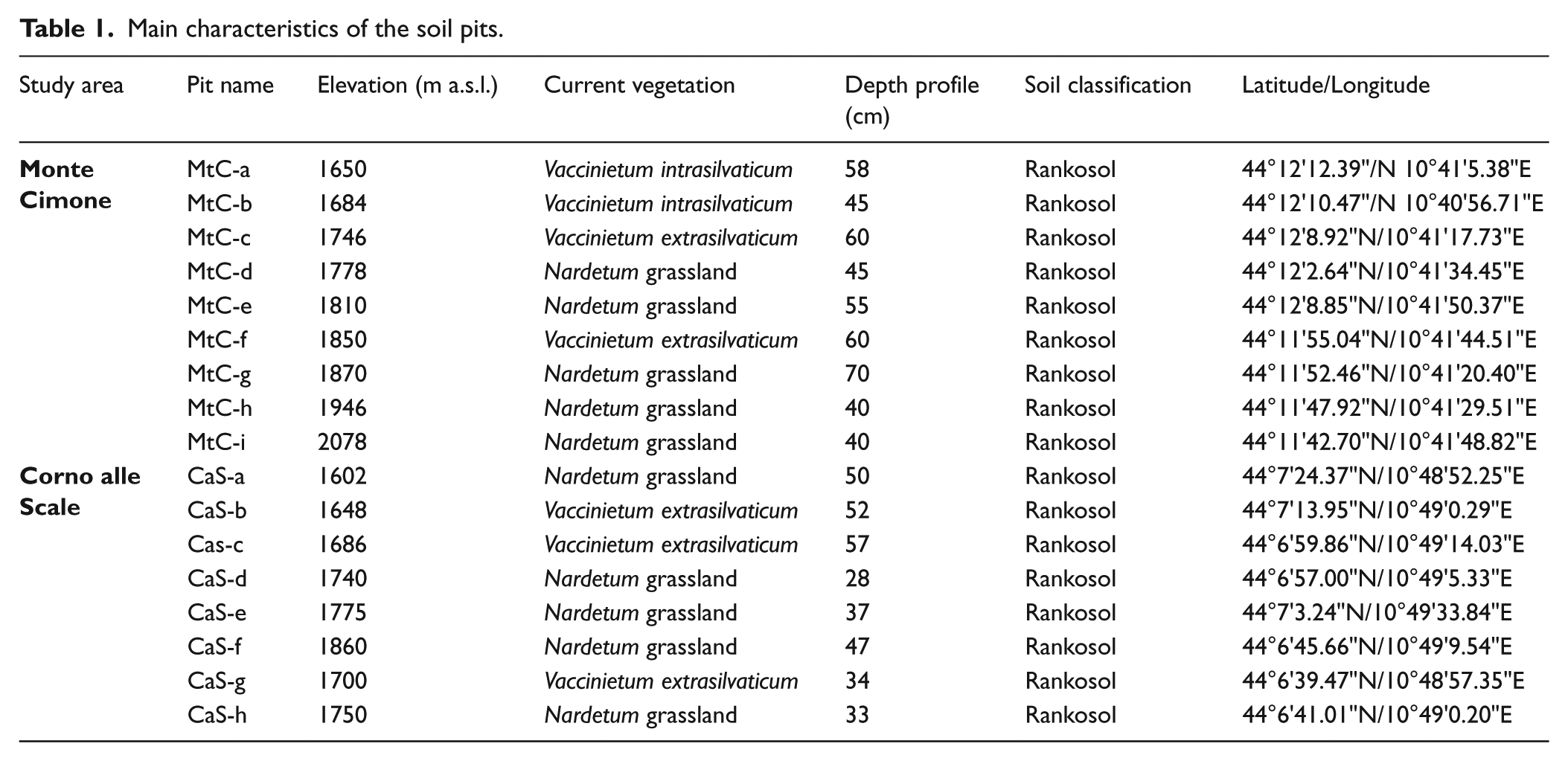
For each pit, after turf removing, soil samples were taken every 10 cm approximately, collecting about 5 kg of wet soil for each sample, from the top part of the sampling point to the bedrock (inverse direction from what has been done in other similar studies: e.g. Carcaillet, 2001; Carcaillet and Thinon, 1996; Robin et al., 2013; Saulnier et al., 2015; Talon, 2010). This method, that has already been tested in previous pedoanthracological studies, offers the advantage of simplifying the sampling without excavating a large pit (see Bal et al., 2015; Garcia Alvarez et al., 2017). A total of 43 soil samples from Monte Cimone and 37 from Corno alle Scale were collected. After sampling, a field description of soil profiles was done. The soils sampled in our study areas can be classified as ‘ranker’ soils, whose depth varies from 40 to 70 cm at Monte Cimone and from 28 to 57 cm at Corno alle Scale. The main characteristics of soil pits are indicated in Table 1.
Main characteristics of the soil pits.
Extraction and identification
Soil samples were dried to allow the potential charcoal fragments present in the samples to dry up to their inner part, thus becoming more resistant and less breakable for the analysis to follow (Carcaillet and Thinon, 1996) and were sieved with water jet using a column of four sieves with increasingly finer meshes (5, 2, 0.8 and 0.5 mm). Charcoal fragments retained in these sieves were large enough to not be transported far by wind and by the hot air flows generated by fires, providing indications about the vegetation that was burned near the sampling point (Talon et al., 1998). Moreover, their size still allows for taxonomic identification (Carcaillet and Thinon, 1996). For each soil sample, about 3 kg of dried soil were sieved. Charcoal sorting was carried out with a binocular microscope (at 10×–15×).
Taxonomic identification of charcoal fragments was performed by observing their transversal, longitudinal tangential, and longitudinal radial sections under a reflected-light microscope (100×, 200×, and 500×; Nikon Eclipse LV100) with the help of wood anatomy atlases (Jacquiot et al., 1973; Schweingruber, 1990; Vernet et al., 2001) and a reference collection of charred wood at GEOLAB in Limoges. All charcoal fragments from sieves with meshes of 5, 2, and 0.8 mm were analyzed; soil remains from 0.5 mm meshes were kept aside because the charcoal fragments contained in the other sieves provided sufficient amount of charcoal and sufficient botanical information, as has often been done in similar studies (e.g. Carcaillet and Thinon, 1996; Cunill et al., 2012; Moser et al., 2017).
Quantification and counts
The charcoal fragments of each sampling level were grouped on the basis of their taxon and weighed in order to obtain the anthracomass, which is a charcoal concentration value expressed in mg / kg. This is the milligrams of charcoal per 1 kg of dry soil, excluding the inorganic soil fraction greater than 5 mm (Carcaillet and Thinon, 1996). This value, called Specific Anthracomass (SA), was calculated for each pit (Figure 2) (GSA – General Specific Anthracomass) and for each taxon of each level of each pit (Figures 3–5) (SAT – Specific Anthracomass Taxonomic) (Talon et al., 1998).
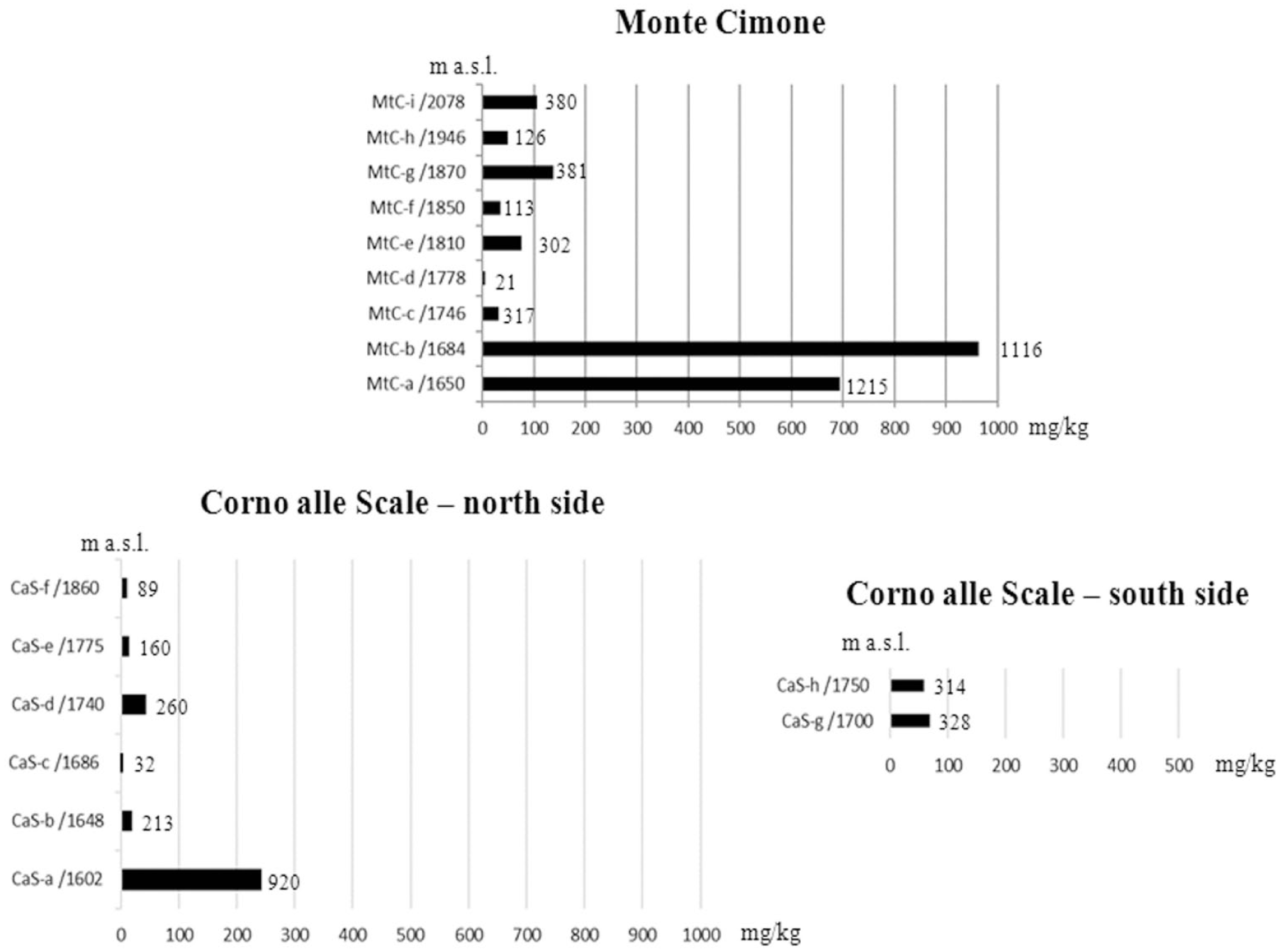
GSA – General Specific Anthracomass, that is the Anthracomass calculated for each pit. The number of charcoal fragments for each pit is indicated.
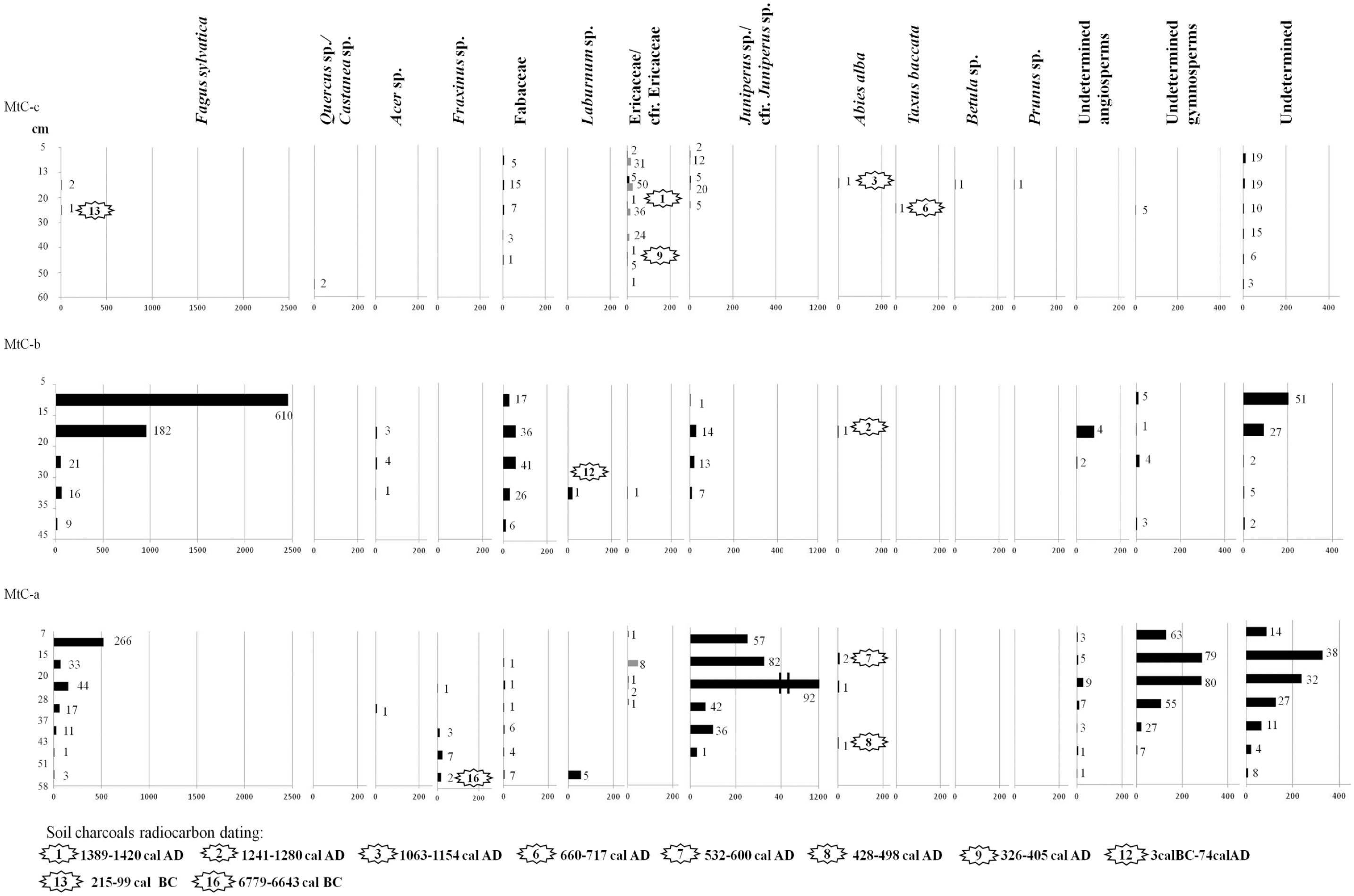
SAT – Specific Anthracomass Taxonomic, referring to MtC-a, MtC-b, and MtC-c of Monte Cimone (expressed in mg/kg; x-axis below) per identified taxon (x-axis above) in soil levels of the pits. Soil levels of sampling are indicated on y-axis. The number of charcoal fragments by taxon and by level is indicated. Gray color in the diagram refers to charcoal fragments identified as cfr.
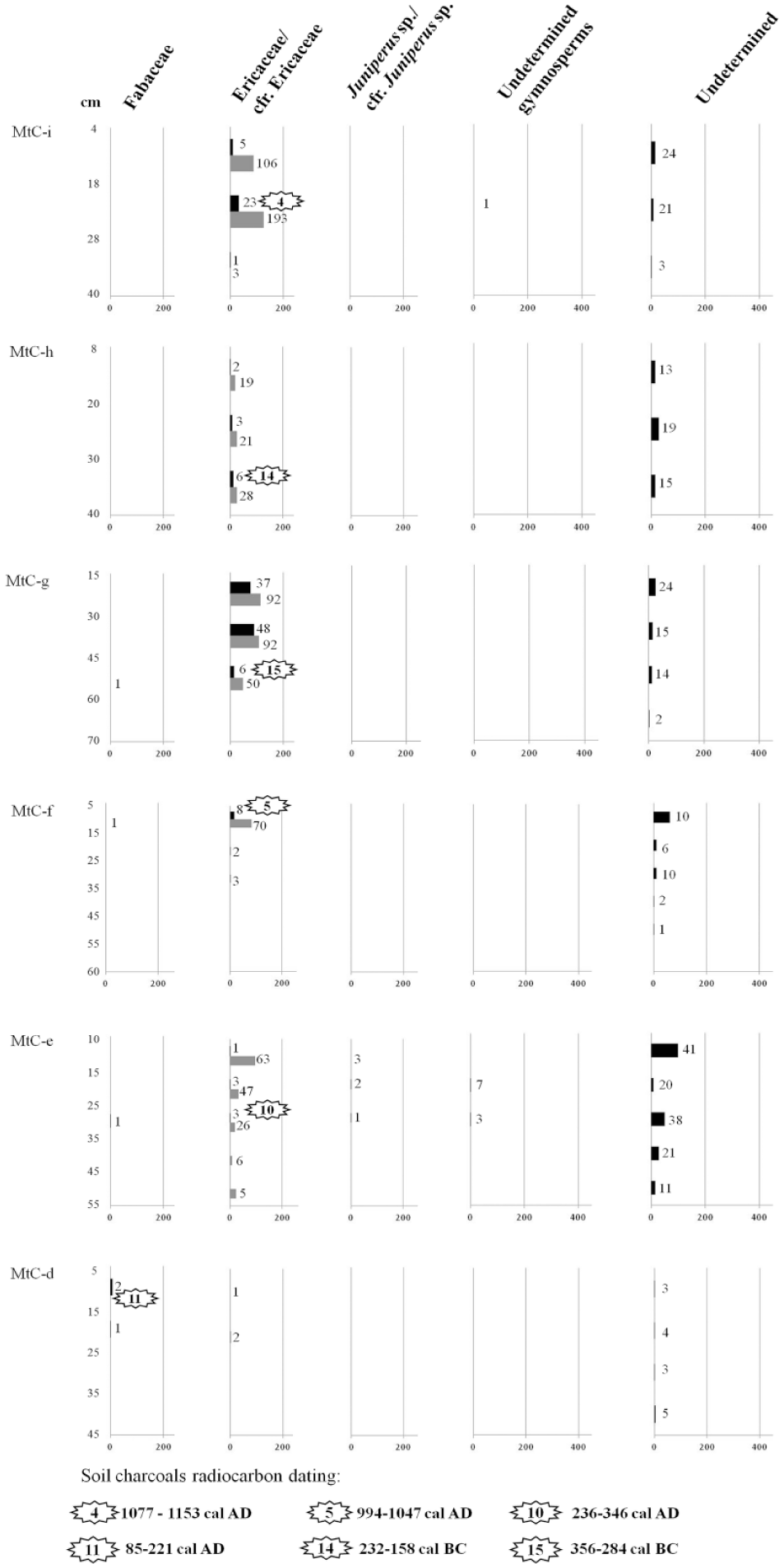
SAT – Specific Anthracomass Taxonomic, referring to MtC-d, MtC-e, MtC-f, MtC-g, MtC-h, and MtC-i of Monte Cimone (expressed in mg/kg; x-axis below) per identified taxon (x-axis above) in soil levels of the pits. Soil levels of sampling are indicated on y-axis. The number of charcoal fragments by taxon and by level is indicated. Gray color in the diagram refers to charcoal fragments identified as cfr.

SAT – Specific Anthracomass Taxonomic, referring to the pedoanthracological pits of Corno alle Scale (expressed in mg/kg; x-axis below) per identified taxon (x-axis above) in soil levels of the pits. Soil levels of sampling are indicated on y-axis. The number of charcoal fragments by taxon and by level is indicated. Gray color in the diagram refers to charcoal fragments identified as cfr. VS indicates the south slope of Monte Cornaccio..
Next to the calculated anthracomass for pit and for taxon from each level of each pit, the corresponding number of charcoal fragments was noted (Bal et al., 2015) in order to compare if the two types of values (number or anthracomass) were similar as quantitative values and their representativeness.
Charcoal fragments datings
A total of 25 charcoal fragments were dated with 14 C Accelerator Mass Spectrometry (AMS) at the Center for Applied Isotope Studies (University of Georgia, USA): 16 from Monte Cimone and 9 from Corno alle Scale area. The dated charcoal fragments were chosen on the basis of the best compromise between their taxonomic identification, the weight (sufficient to be dated), the location of the pits from which they come, and their position along the soil profile. The dates were calibrated as BP and BC/AD by the CALIB program (Stuiver et al., 2005), version 5.0, based on the data set Intcal04 (Reimer et al., 2004) and reported as intercept with 2 sigma.
Results
Anthracomass
All the pedoantracological excavated pits contained charcoal fragments (Figure 2).
A total of 3971 charcoal fragments were analyzed from Monte Cimone and 2316 from Corno alle Scale. The General Specific Anthracomass calculated for each pit of Monte Cimone ranged from a minimum of 4.9 mg/kg (pit MtC-d located at 1778 m a.s.l.) to a maximum of 962.8 mg/kg (MtC-b, 1684 m a.s.l.). The GSA of Corno alle Scale ranged from a minimum of 1.6 mg/kg (CaS-c, 1686 m a.s.l.) to a maximum of 243.4 mg/kg (CaS-a, 1602 m a.s.l.).
The GSA matches the trend of the relative total number of charcoal fragments; in other words, the number of charcoal fragments reflects the general tendency of the results expressed in anthracomass. This relationship has also been found in other studies where the number of charcoal fragments is reported (e.g. Carnelli et al., 2004; Cunill et al., 2012; Jansen et al., 2013; Bal et al., 2015).
Taxonomic identification
In the Monte Cimone area, 14 arboreal/shrubby taxa were identified: Abies alba, Acer sp., Betula sp., Ericaceae, Fabaceae, Fagus sylvatica, Fraxinus sp., Juniperus sp., Laburnum sp., Prunus sp., Quercus/Castanea, Taxus baccata, undetermined angiosperms and undetermined gymnosperms (Figures 3 and 4). In the Corno alle Scale area, the nine arboreal/shrubby taxa were: Acer sp., Ericaceae, Fabaceae, Fagus sylvatica, Fraxinus sp., Juniperus sp., Laburnum sp., undetermined angiosperms, and undetermined gymnosperms (Figure 5).
Many Ericaceae (cfr.) were affected by a certain degree of vitrification, a phenomenon that often affects small twigs under high temperatures (Marguerie and Hunot, 2007). About 77% of the undetermined angiosperms of Monte Cimone and about 68% of those of Corno alle Scale had a certain degree of vitrification; the same phenomenon was present in about 10% of the undetermined gymnosperms of Monte Cimone and in about 14% of those of Corno alle Scale.
The percentage of charcoal fragments in a poor state of preservation, and therefore undetermined, was quite high in both study areas; at Monte Cimone it varied from a minimum of 7% in the MtC-b and MtC-i pits to a maximum of 50% in the MtC-e pit; at Corno alle Scale it ranged from a minimum of 1.5% in pit CaS-a to a maximum of 64.8% in CaS-c. The amount of the undetermined charcoal fragments is very variable from one sample to another and this may be due to taphonomical issues which however did not leave clear macroscopic traces in the sediment. Despite the results obtained from the pits are affected by high values of undetermined charcoals, which if taken individually should be interpreted with caution, the sampling strategy (many samples involving specific points over a large area) allows a valid spatial interpretation of the results obtained.
Distribution of the anthracomass
Observing the altitudinal distribution of the anthracomass of the taxa found at Monte Cimone, we distinguished four zones and the presence of a gradual altitudinal succession of vegetation: from a wooded to an open vegetation zone crossing an intermediate transition zone: (1) in the MtC-a and MtC-b (just below the current timberline – 1650 and 1684 m a.s.l.), tree taxa dominate, and Fagus sylvatica prevails, accompanied by traces of other tree taxa, such as Fraxinus sp., Acer sp., and Abies alba; (2) in MtC-c (1746 m a.s.l.) shrubby taxa dominate (Ericaceae and Fabaceae) with small quantities of arboreal taxa, Fagus sylvatica, Quercus sp./Castanea sp., Abies alba, Taxus baccata, Betula sp., and Prunus sp.; (3) in MtC-d (1778 m a.s.l.), there is clear prevalence of Fabaceae and total disappearance of tree taxa; and (4) in the MtC-e, MtC-f, MtC-g, MtC-h, and MtC-i (from 1810 to 2078 m a.s.l.) there is a dominance of Ericaceae while the arboreal taxa continue to be absent.
On the north side of Corno alle Scale, we distinguished four zones and the lack of a gradual altitudinal succession of vegetation: (1) in CaS-a (just above the current timberline – 1602 m a.s.l.) tree taxa, especially Fagus sylvatica, accompanied by Acer sp. are dominant; (2) in CaS-b and CaS-c (located in the central part of the pastoral space – 1648 and 1686 m a.s.l.) there are only shrubby taxa (Ericaceae and Fabaceae) and the trees are absent; (3) in CaS-d and CaS-e (1740 and 1775 m a.s.l.) again the shrubby taxa (Fabaceae and Ericaceae) dominated but Fagus sylvatica reappear; and (4) in CaS-f (1860 m a.s.l.) again there are only shrubby taxa (57% Ericaceae and 4% Fabaceae) without trees.
On the south side of Corno alle Scale (corresponding to the south versant of Monte Cornaccio), CaS-g and CaS-h (1700 and 1750 m a.s.l.) show the dominance of shrubby taxa (Ericaceae, Fabaceae, and Juniperus sp.) with small amounts of trees such as Fagus sylvatica and Fraxinus sp.
Radiocarbon dates
Radiocarbon dates are distributed from 9906-9658 cal BP to 10 ± 25 cal BP (Figure 6). Almost all the dates (15 out of 16 at Monte Cimone and 8 out of 9 at Corno alle Scale) were from the Late Holocene. One date for each study area fell in the Early Holocene (Fraxinus sp. from Monte Cimone and Ericaceae from Corno alle Scale). For the Middle Holocene, there is therefore a gap in the dates (Figure 6).
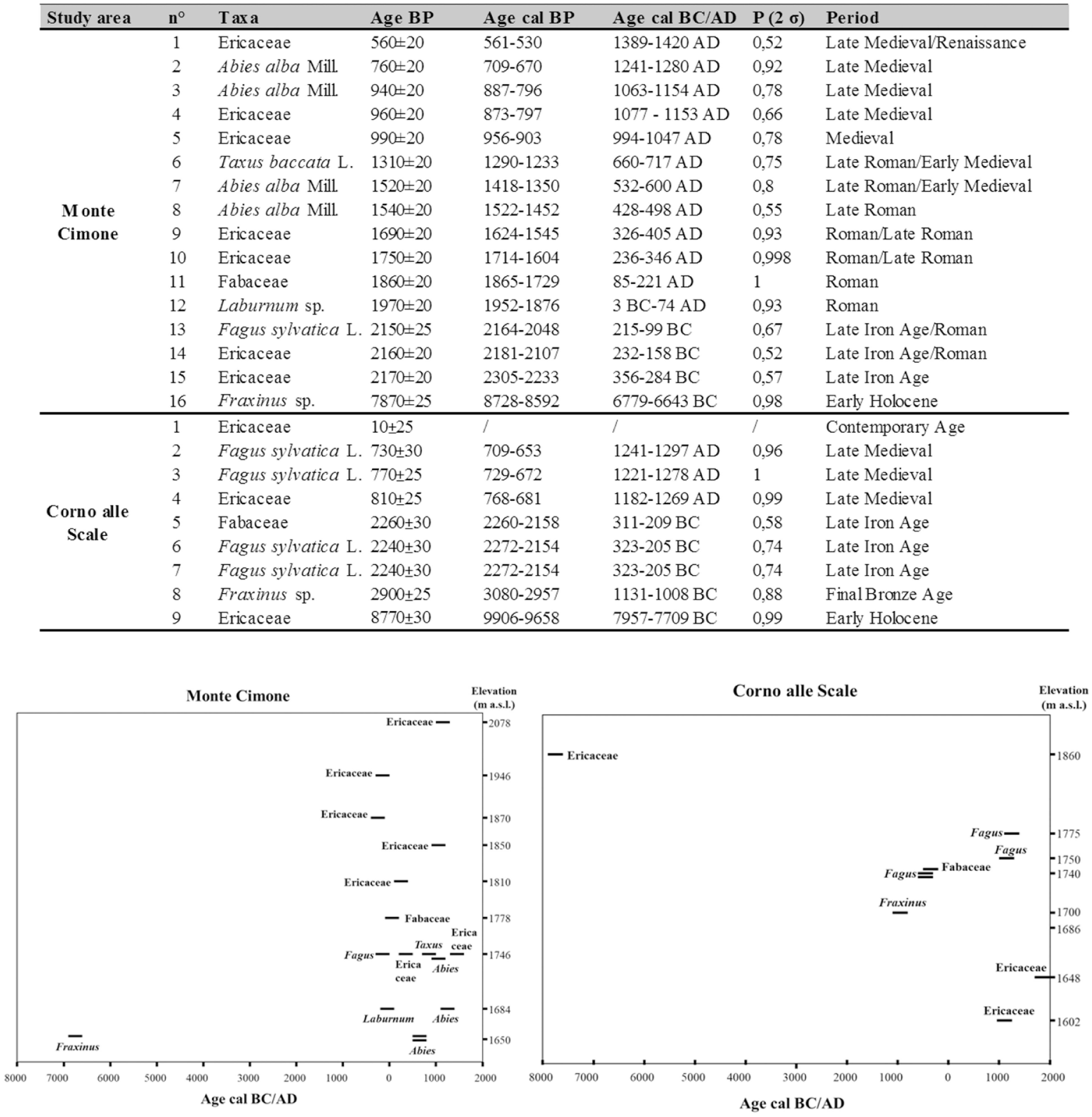
Radiocarbon dates of soil charcoal fragments and their temporal and altitudinal distribution (2 σ probability intervals).
Discussion
Based on floristic composition and radiocarbon dates, the pedoanthracological data give information on Early- and Late-Holocene vegetation dynamics in the studied area. The charcoal fragments contained in soils without a very steep surface come from the plant communities burned in the station within a radius of some tens of meters (Talon et al., 1998; Thinon, 1992).
Forest composition and high altitude prairies in the Early Holocene
Two radiocarbon dates of the Early Holocene provide some information on early floristic composition in the two areas. The fragment of ash (probably F. excelsior, on the basis of its anatomy combined with the ecology of this species) dated to 8728-8592 cal. BP (MtC-a) at 1650 m a.s.l., just below the current timberline of Monte Cimone, attests its past presence 150 m higher than today (Pignatti, 1982). This is consistent with pollen data that show thermophilous tree taxa, including ash, in the woods located at higher altitudes and dominated by fir during Preboreal and Boreal periods (Vescovi et al., 2010a; Watson, 1996). Moreover, high quantities of charcoal fragments of this tree were part of the anthracological spectra from fireplaces of Mesolithic sites discovered in the Tuscan-Emilian Apennines (Castelletti et al., 1976; Cremaschi and Castelletti, 1975; Cremaschi et al., 1984).
The charcoal fragment of Ericaceae dated to 7957–7709 cal. BC (CaS-f) and located in the highest pit of Corno alle Scale, at 1860 m a.s.l., in association with lack of tree charcoal in the same pit, suggests that, in that period, the Apennine ridge was already open and bare of arboreal vegetation.
The ancient forest close to the timberline during the Late Holocene
During the Late Holocene, the ancient forest composition just below the current timberline seems to have been characterized by Fagus sylvatica dominance with the presence of small quantities of Fraxinus sp., Acer sp. and Abies alba (Figures 3 and 5). Pedoanthracological data attest to the presence of fir at Monte Cimone for the Late Roman/Early Middle Ages and the Late Middle Ages. No fir charcoal fragments were found at Corno alle Scale. The small amounts of Abies alba found in our study areas (few mg/kg in a pit) seems to be in contrast with the high quantities (up to about 4000 mg/kg in a pit) of fir charcoal fragments found in another pedoanthracological study carried out at Monte Cusna (Northern Apennines – 25 km away from our study areas), at the same elevation and during the Late Holocene (Compostella et al., 2013). This high difference suggests that the current distribution of Abies alba was largely influenced by anthropic action. In fact, fir is very sensitive to fire and grazing (Bernetti, 1995; San-Miguel-Ayanz et al., 2016). The different distribution of the fir charcoal between Monte Cusna (Compostella et al., 2013) and our study is linked and supported by the current distribution of natural fir populations; in the Apennines of the province of Reggio Emilia (where the Monte Cusna is located) there are currently five spots of natural populations of A. alba (Alessandrini and Branchetti, 1997) whereas in the Apennines of the province of Modena (where the Monte Cimone is located) natural spots of fir are absent (Alessandrini et al., 2010).
The presence of forest opening attempts and intrasylvatic open spaces, consisting mainly of Fabaceae and Ericaceae close to the timberline, is suggested by two charcoal fragments: at Monte Cimone, Laburnum sp. dated to 3 BC–AD 74, the Roman Period, and at the Corno alle Scale, Ericaceae (probably Vaccinium since this is currently the most widespread representative of the family in the subalpine moors of the Northern Apennines) dated to 1182 AD–1269 AD, the Late Middle Ages. These plants suggest that some clearings may have been made by human societies using fire to open spaces, most probably for pastoral activities, both during the Roman period (Geri, 1992; Giordani, 2006) and during the Middle Ages (a key period for large-scale European deforestation by agro-pastoral activities – Bal et al., 2015), during which the importance of sheep breeding is attested in the highest belts of the Apennines (Cremaschi and Dall’Aglio, 1988).
The ancient timberline and treeline
On the northwest slope of Monte Cimone, the pedoanthracological findings show that during the Late Holocene, the timberline was located at about 1746 m a.s.l., 46 m higher than the current one. This timberline probably had phases of closed forest, as indicated by the fragments of Fagus sylvatica (dated between Late Iron Age and Roman Period – 215 BC–99 BC) and Abies alba (dated to the Late Middle Ages – 1063 AD–1154 AD) alternated to phases of open forest as suggested by the fragments of Prunus and Betula (not directly dated); this succession has been also identified in other European mountains (e.g. Bal et al., 2015; Cunill et al., 2012; Talon, 2010). Remarkably, the charcoal fragments of Taxus baccata (660 AD–717 AD) found at the ancient timberline is an important find because today yew is a relict species with a fragmented distribution; in the province of Modena it has only two spots of natural populations (15 km away from our study areas) and is protected by parks (Alessandrini et al., 2010). The use of fire to create and keep open pasturelands (Alessandrini et al., 2003), over time, may have led to the loss of the memory of the ancient forests above the current timberline. The ancient treeline is difficult to detect due to the lack of tree taxa above 1746 m (MtC-c pit).
On the north side of Corno alle Scale, the pedoanthracological study shows that small beech groves lived on the walls of the glacial cirque, at 1740 m a.s.l. at the end of the Iron Age and at higher elevation, 1775 m a.s.l., in the Late Middle Ages (140 and 175 m higher than the current timberline, respectively). Also on the south side, there was an ancient beech forest that lived at 1750 m a.s.l. during the Late Middle Ages (150 m higher than the current timberline). These ancient remains of Fagus sylvatica forest were probably spared from grazing. In fact, beech charcoal fragments are absent in intermediate pits between these pits and the current timberline (CaS-b, CaS-c and CaS-g) while shrubs are prevalent suggesting that the regions at about from 1600 to 1740 m a.s.l. have been used as pasture for a long time. As at Monte Cimone also at Corno alle Scale, the current timberline seems to have been stable during the last 3000 years.
The slight shift of the timberline identified in our study areas contrasts strongly with the large displacements (from 250 to 400 m) identified in other European mountains (e.g. Alps – Talon et al., 1998, 2010; Alps – Carcaillet and Brun, 2000; Alps – Carnelli et al., 2004; Alps – Ali et al., 2005; Pyrenees – Cunill et al., 2012; Sierra de Gredos – Garcia Alvarez et al., 2017; Alps – Badino et al., 2018). This can be regarded as a peculiar feature of this part of the Northern Apennines which seem to have been characterized by a similar plant landscape during the whole Late Holocene.
Open landscapes above the current timberline
The current open landscape of Monte Cimone, above the timberline (from 1870 to 1946 m a.s.l.) (Figure 4), seems to have been completely void of trees and dominated by Vaccinium already at the end of Iron Age/beginning of Roman Age. The evidence from our data is supported by the pollen profiles of six lakes in the Ligurian and Emilia-Romagna Apennines that show an increase in herbaceous taxa matching the opening of the landscape from 3000 cal BP (Branch and Marini, 2014).
At Corno alle Scale, radiocarbon datings did not allow us to infer the opening and first use of pastoral spaces located just above the current timberline. Yet the Ericaceae charcoal fragments dating back to the 20th century may be a testimony of an ancient practice that until recently had been maintained. One informant mountain dweller, Mr. Dario Brugioni, hailing from Fiumalbo (a small village near Monte Cimone) recalls that, until the 1960s, when he was young, in September before returning to the plains, the shepherds used to set fire to the dry grass to regenerate the high altitude pastures. So, from the timberline to the ridge, all the pastures were potentially subject to fire. Obviously, there was a rotation of the areas to be regenerated, that is, the same pastureland was not necessarily set on fire every year (Brugioni, in verbis).
Altitudinal migration of some taxa
Some taxa (in brackets the hypotheses of the most probable species given the elevation habitat) were found at higher altitudes compared to their current altitudinal limit; in particular: Quercus sp. (Q. cerris or Q. pubescens) – 250–550 m; Prunus sp. (P. spinosa or P. avium) – about 250 m; Taxus baccata – 150 m; Fraxinus sp. (F. excelsior) – 150 m (Pignatti, 1982). Charcoal fragments from these taxa are larger than 400 μm and this provides good evidence of a local fire; charcoal fragments ⩾400 μm do not enter into suspension in the convective column of the fire and they are hardly transported by rolling (Clark, 1988; Ohlson and Tryterud, 2000). The presence of these taxa at higher altitudes in the past may have been favored during periods of optimal climate and by the presence of open environments, a condition of more favorable growth compared to a dense beech forest.
Fire events correlated with human practices
The presence of charcoal fragments in the pits along the altitudinal transects (from 1650 to 2078 m at Monte Cimone; from 1602 to 1860 m at Corno alle Scale) demonstrates the importance of fire in shaping the current vegetation. As mentioned above, 23 out of 25 dates fall in the Late Holocene, and the fire events fall during both warm and cold periods (Figure 7). Most of the fires occurred during warm periods (17 dates), while only a few fell into cold periods (5 dates), and one was in a transition period (Figure 7). Figure 7 shows two key phases of high fire frequency (as inferred from datings) in the history of our study area: one is the transition between Late Iron Age and Roman Age (beginning of the Roman climatic optimum) and the other is the beginning of the Medieval climatic optimum. It seems therefore that climate optimum phases have favored changes of the landscape, probably triggered by the expansion of human activities (Poschlod, 2015).
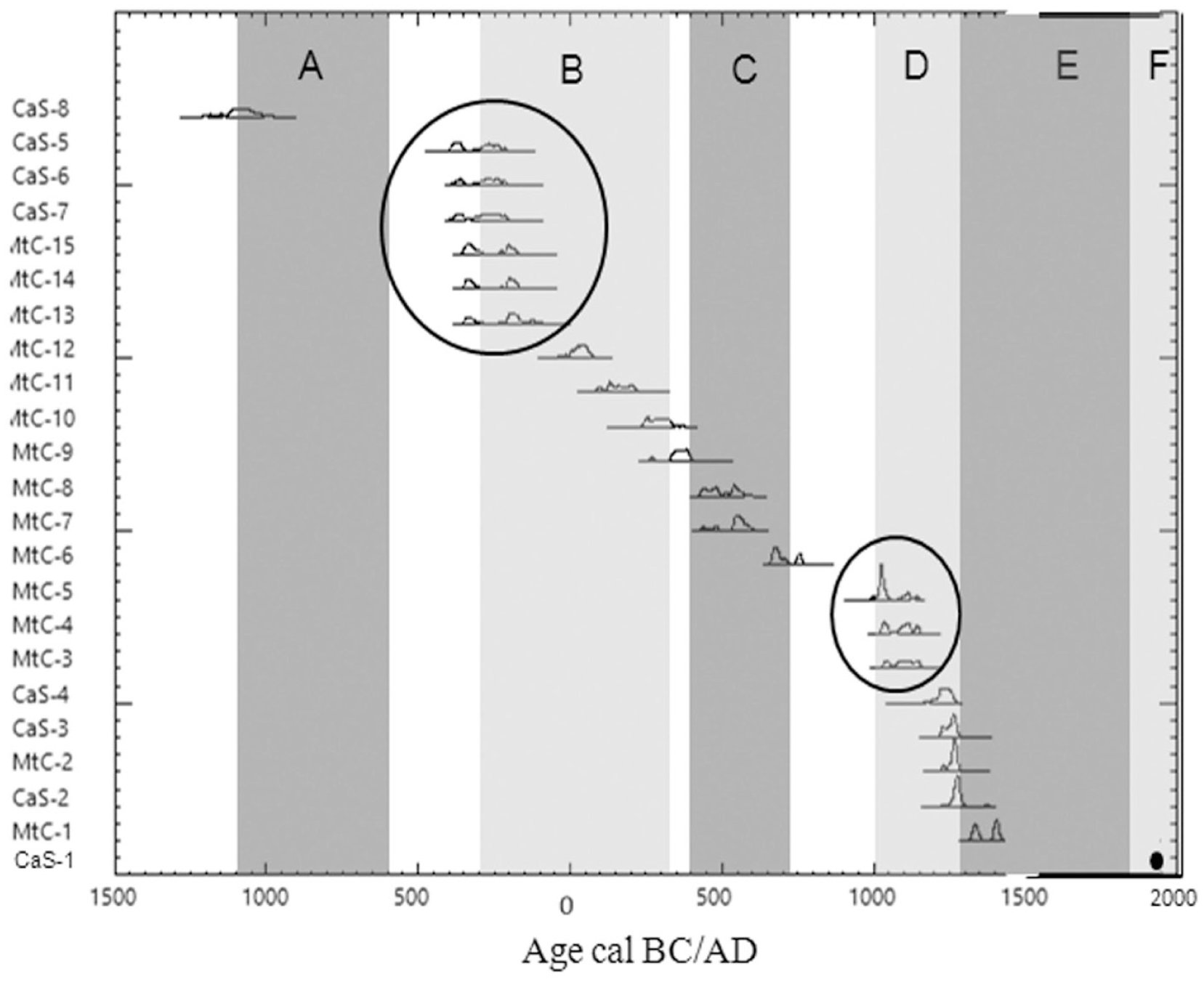
Temporal distribution of the 23 radiocarbon dates falling into the Late Holocene with the indication of the climatic optimum (light gray) and of the climatic deterioration (dark gray).
The pedoantracological results, combined with the spatial analysis of the sampling points and the archaeological evidence of human presence in the area during the Late Holocene (Cardarelli and Malnati, 2006; Guidanti, 1996, 1998), enabled us to propose a scenario for pastoral and forest areas management in these Apennines. This has already been observed previously in other European mountains (e.g. Davasse, 2006; Galop, 1998; Pescini et al., 2017).
For both study areas, there is a point of sharp decrease in the anthracomass, between MtC-b (1684 m a.s.l.) and MtC-c (1746 m a.s.l.) at Monte Cimone and between CaS-a (1602 m a.s.l.) and CaS-b (1648 m a.s.l.) at Corno alle Scale. This narrow belt coincides with a change in the geomorphological conformation, that is, the transition from a steep slope (where the forest is currently present) to completely open pastoral plateaus (where grazing is practiced). This observation suggests that the inhabitants of the two mountain areas exploited the natural physical conformation of the territory to use and manage natural resources through sylvo-pastoral activities, thus using the forest on the steep slopes for the production of charcoal (on charcoal platforms which were being used until the mid-20th century; Benatti et al., 2018) and the plateaus for livestock grazing. This management can also be inferred by pits distribution (Figure 2) and anthracomass amounts, which are counterintuitive with respect to their location. Pits in flat points in the central part of pastoral plateaus (MtC-c, MtC-d, MtC-e and MtC-f; CaS-b and CaS-c), and therefore in a position where erosion should be less intense, have lower anthracomass values than those of the pits placed at higher altitudes and with a certain slope (MtC-g, MtC-h and MtC-i; CaS-d, CaS-e and CaS-f), located in points where the erosion should be more intense. The preference for the flattest parts for grazing may have hindered the development of arboreal and shrubby plants and favored herbaceous ones, thus reducing the produced anthracomass. The fires set by the shepherds to extend the pastures at the expense of Vaccinium heath (Cevasco, 2012; Geri and D’Addario, 1997) and the tree cover may have, over time, caused a lower production of charcoal during fires. The openings created by fire were then maintained by the presence of the flocks and consequently the shepherds had less need to use the fire.
The Middle-Holocene gap in the radiocarbon datings
Although the radiocarbon dating did not provide dates for the Middle Holocene, it is unreasonable to hypothesize a total lack of fires for this chronological phase, especially because the presence of large amount of settlements is known in the region since prehistoric times (Cardarelli and Malnati, 2006; Mercuri et al., 2015). Many studies carried out in Europe show that during the Neolithic period, under the influence of the climate optimum, agro-pastoral societies were able to shape the plant landscape (Mercuri et al., 2019a) including woods (Bal et al., 2010; Cunill et al., 2012; Poschlod, 2015; Schwörer et al., 2015; Talon, 2010). In that period, a significant increase in fire activity has been recorded in sedimentary records (Ali et al., 2005; Carcaillet, 1998, 2001; Carcaillet et al., 2002; Mocci et al., 2005; Vannière et al., 2011).). Moreover, the palynological studies carried out on the Northern Apennines show the occurrence of the first openings in the forest cover during the Middle Holocene (Branch and Marini, 2014; Vescovi et al., 2010a, 2010b; Watson, 1996). These openings may have been caused by pastoral societies (Cardarelli and Malnati, 2006; Cevasco, 2012; Stagno, 2016; and see also Mercuri et al., 2012).
To explain the lack of records dated to the Middle Holocene, we have two hypotheses: (1) problems of charcoal preservation in soils during the Middle Holocene. Intense erosion phenomena (such as landslides) may have occurred in the period before the end of the Bronze Age that removed portions of soil and the relative charcoal fragments. Geoarchaeological, geomorphological, and micromorphological studies carried out in the nearby Apennines areas indicate that these soils have often suffered from very active slope instability during the climatic deterioration of the Subboreal (between the end of the Bronze Age and the beginning of the Iron Age), which led to frequent erosion and sediment removal events (Bertolini, 2007; Compostella et al., 2014, 2013; Cremaschi et al., 1984); and (2) high fire activity in the Late Holocene (Vescovi et al., 2010a, 2010b) may have masked that of previous periods. Therefore, the intense fire activity from the final Bronze Age to the recent times may have produced a greater number of charcoal fragments than in the previous period; as a consequence, the probability of dating charcoal fragments prior to the final Bronze is low. Considering the current and past physical and geomorphological situation of the study area with the presence of important erosive phenomena, the loss of charcoal seems to be the most probable and most important hypothesis whose effects may have been amplified by high fire activity in the Late Holocene.
Conclusion
The pedoanthracological analysis carried out in this study has provided new data with a large spatial resolution on the history of the plant landscape especially on the last 3000 years of history of the Northern Apennines. As elsewhere (e.g. Mercuri et al., 2019b), climate and human action have influenced the vegetation and fire dynamics even in the mountains, and pedoanthracology has provided information on a reasonable and consistent spatial distribution of woods and pastoral practices in these areas.
The research identified a Late-Holocene plant landscape that was quite similar to the current one. In both study areas, in fact, the analysis of soil charcoal fragments shows: (1) a forest belt, which almost coincides with the current one, dominated by beech and with the presence of intra-forest clearings; (2) an intermediate area, coinciding with current pastoral plateaus, characterized by an open landscape dominated by Ericaceae and Fabaceae from an anthracological point of view, but with small wooded portions at some points; this suggests that wooded pastures were widespread in the past, and may have been an important component of the cultural landscape of this study area; (3) a completely open area, close to the ridge, dominated by Ericaceae and Fabaceae as today. Results opened also interesting perspectives for the reconstruction of the history of some tree species like Taxus baccata and Abies alba, which are endangered or less spread today with respect to the past.
Soil charcoal fragments analysis suggests that the soils of the two study areas contain mainly Late-Holocene charcoal. We suggest that this may be due to two main causes: (1) frequent erosion and slope instability that occurred before the Late Holocene (and before the Bronze Age), which may have hindered the conservation of charcoal in the soils; (2) high and intense fire activity during the Late Holocene that may have masked that of the Early and Middle Holocene.
The spatial resolution of pedoanthracology and the archaeological and historical data have identified a logic of space use. In both study areas, it seems that the mountain societies first started to exploit the more flat portions of the mountain territory as pasture, and then moved on to other areas when the increased grazing pressure required additional surfaces.
Footnotes
Acknowledgements
We want to thank the regional parks Parco del Frignano and Parco del Corno alle Scale and the city of San Marcello Pistoiese for the sampling authorizations. Many thanks to all who helped in sampling: Daniele Bertoni, Paolo Gibertini, Salvia García álvarez, and Graziella Rassat. The authors are also grateful to Dario Brugioni, Donato Labate, and Marcello Crotti for their help with fieldwork and their archaeological and historical knowledge of the study area. Finally, we would like to thank the reviewers, for their constructive suggestions and comments.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was developed in the framework of the PhD thesis project entrusted to A.B., and carried out between the School 613 Sciences de la Société, Territoires, Sciences Économiques et de Gestion of the University of Limoges (France), and the School M3ES Models and Methods for Material and Environmental Sciences of the University of Modena and Reggio Emilia (Italy). Financial support was provided by the Project AGRESPE (headed by M.C.B.) and the Limousin Region of France with the PhD scholarship.