
Abstract
This study integrates diatom analysis with existing records of pollen, charcoal, elemental composition, and stable light isotopes to expand upon the 4200-year history of human activity and climate change from Laguna Los Mangos in southern Pacific Costa Rica. We counted diatoms in peroxide-treated samples and analyzed community composition using cluster analysis, revealing four distinct assemblage zones with diatom variability most closely correlated with phosphorus, titanium, and organic content. The earliest assemblage (Zone D, 4150–3430 cal yr BP) was dominated by Encyonema silesiacum and Nitzschia incognita and aligned with a period of deforestation, erosion, and abundant macrophytes. Gomphonema affine proliferated in Zone C (3430–2450 cal yr BP), reflecting increased pH and productivity likely caused by agriculture-induced nutrient loading. We attributed the preservation gap from 3290 to 2970 cal yr BP in Zone C to silica depletion and erosional deposition that induced decline in diatom abundance by diluting valve concentrations in lake sediments. Nitzschia incognita and G. affine became the dominant taxa in Zone B (2450–1400 cal yr BP), likely reflecting eutrophy, increasing conductivity, and drying climate. Dominance of Diadesmis confervacea indicated reduced lake level in Zone A (1400 cal yr BP–modern) at the onset of the Terminal Classic Drought (TCD). A hiatus in the record indicates lake desiccation from 950 to 450 cal yr BP. During the Little Ice Age (LIA), diatoms reflect conditions similar to Zone B indicating increased lake level, circumneutral pH, and eutrophy. Refilling of the lake indicates increased precipitation during the LIA despite evidence of severe regional drought reported at other sites. Variable precipitation during this period likely resulted from the combined effects of Spanish contact, agricultural collapse, forest recovery, and shifts in Atlantic and Pacific climate forcing mechanisms. Overall, the Los Mangos diatom record reflects shallow, slightly alkaline, eutrophic conditions influenced by nutrient enrichment, erosion, and deforestation associated with maize agriculture.
Introduction
Humans have modified landscapes for millennia with agriculture and the use of fire, but the history of these activities and their connection to changing climates would remain uncertain without paleolimnological records. Time-series analysis of fossil diatoms, pollen, charcoal, isotope ratios, elemental composition, and other proxies in lake-sediment records makes it possible to reconstruct timelines of human activity, climate, and environmental change in specific watersheds. Diatoms are an especially valuable proxy because they are abundant, morphologically distinct, ecologically diverse, and responsive to changes in lake conditions. Diatom records provide site-specific records revealing relative changes in lake depth (Battarbee, 1988), nutrient status (Battarbee, 1978), pH (Battarbee, 1984), and the degree of stratification (Battarbee, 1988).
Diatom composition reflects physical and ecological conditions within a waterbody, while other proxies like carbon isotope ratios, charcoal, pollen, and elemental composition are primarily influenced by conditions in the surrounding watershed. Multivariate analysis of diatom composition in conjunction with other proxies enables the reconstruction of physical conditions within a waterbody such as pH, salinity, lake stratification, nutrient status, and productivity that cannot be reconstructed from individual proxy records. This technique also reveals how variables are interrelated and how they respond to environmental changes driven by human activity and changes in climate. In this study, we use fossil diatom analysis and constrained cluster analysis to describe changes in lake ecology and limnological conditions in response to land clearance, agriculture, fire, and drought over the Late-Holocene, or Meghalayan, at Laguna Los Mangos in southern Pacific Costa Rica. Our objectives are to (1) Interpret past water conditions reflected by diatom community composition; (2) Compare the timing of shifts in diatom assemblages at Laguna Los Mangos to shifts in other proxies studied in the same core; and (3) Describe the responses of diatom communities to human disturbance from forest clearance and agriculture and to two Late-Holocene climate events: the Terminal Classic Drought (TCD) and Little Ice Age (LIA).
Holocene climate and cultural transition in Central America
During the Holocene global temperatures increased, glacial volume decreased, and sea levels rose as climatic conditions became more suitable for crop cultivation. Changes in climate during the Holocene facilitated the transition from hunter-gatherer to agricultural societies independently in at least eight regions across the globe (Bhattacharya et al., 2015; Diamond, 2002). Crop cultivation increased food availability, driving nomadic hunter gatherers to embrace more sedentary lifestyles. Horticulture, described as the small-scale cultivation of wild and domesticated plants, preceded the use of slash-and-burn agriculture, where large areas of forest were cleared with fire for crop cultivation (Cooke, 2005). In Central America, the introduction of maize agriculture led to population growth, establishment of sedentary villages, colonization of forests, and environmental degradation (Piperno et al., 1985).
Precipitation and atmospheric circulation in the neotropics are primarily driven by El Niño-Southern Oscillation (ENSO; Moy et al., 2002) and the annual migration of the Intertropical Convergence Zone (ITCZ; Lane et al., 2014). The interactions of these climate-forcing mechanisms have been invoked to explain two major climatic events in the Late-Holocene: the Terminal Classic Drought (TCD) ca. 1200–850 cal yr BP (Lane et al., 2014), and the Little Ice Age (LIA) ca. 550–100 cal yr BP (Lane et al., 2011). In the Caribbean, the TCD and the LIA were both expressed as periods of drought attributed to Atlantic and Pacific climate variability, specifically southward shifts in the ITCZ (Bhattacharya et al., 2017; Lane et al., 2011; Sachs et al., 2009) and increased ENSO frequency (Krishnaswamy et al., 2001; Yanuskiewicz et al., 2022). The TCD, which occurred in two distinct phases of severe drought from 1200 to 1075 cal yr BP and 950–875 cal yr BP (Hodell et al., 2005), has also been attributed to the effects of widespread deforestation and agriculture (Cook et al., 2012; Oglesby et al., 2010). The LIA is generally characterized by cooler, drier climate associated with global-scale glacial advance (Ledru et al., 2013), which some have attributed to forest regrowth following widespread agricultural collapse caused by European conquest and disease during the 16th century (Koch et al., 2019; Loughlin et al., 2018). Forest regrowth can also reduce soil erosion as new roots secure sediments, while the shade cast by new forest canopy can change local microclimates by reducing soil temperatures and water evaporation (Guariguata and Ostertag, 2001).
Droughts during these periods had pronounced but regionally distinct effects on pre-Columbian civilizations in the Americas; in the cloud forests of the Peruvian Andes, droughts were coincident with the expansion of agriculture into previously forested areas as conditions became more suitable for maize cultivation (Åkesson et al., 2020). Similar patterns were reported in the Caribbean Antilles, where evidence of drought during the TCD coincided with increased population size, increased reliance on drought-resistant maize and cassava crops, and proliferation of Ostionoid cultural traditions in Hispaniola (Aguilu et al., 1991; Curet, 2005; Keegan et al., 2013; Lane et al., 2014; Rouse, 1992). Contrarily, intense multiyear droughts associated with the TCD were attributed to crop failure, war, and the collapse of the Maya civilization in the Yucatan peninsula (Haug et al., 2003). Pre-Columbian societies’ responses to climate change vary depending on the timing and severity of drought, local environmental conditions, and the ability to adapt or relocate their agricultural practices.
Central American diatom records
Paleolimnological records from southern Pacific Costa Rica reveal a long history of deforestation, agriculture, and climate change during the Holocene (Anchukaitis and Horn, 2005; Clement and Horn, 2001; Horn, 1993, 2006; Johanson et al., 2019, 2020; Kerr et al. 2020; Lane et al., 2004; Taylor et al., 2013, 2015, 2020; Wu et al., 2017, 2019; Yanuskiewicz et al., 2022), but few studies from this region include diatom analysis (Haberyan and Horn, 1999, 2005, 2023). Diatom records reflect changes in the physical and ecological conditions within a waterbody, which also reflect site-specific environmental conditions, human disturbance, and changes in climate that may not be reflected by other proxy records. Supplementing previous reconstructions from Laguna Los Mangos with diatom analysis will provide a more detailed history of the watershed that can be compared to other Central American paleorecords.
A multiproxy analysis of diatoms, pollen, elemental composition, and carbon from lake Las Margaritas, a shallow, low elevation lake in Central Colombia, showed a transition from savanna to rainforest consistent with increased precipitation and seasonality during the early Holocene (Vélez et al., 2005). Detrended Correspondence Analysis (DCA) supported the finding that Eunotia abundance corresponded to deeper and more diluted water, while Aulacoseira abundance corresponded to shallow, open water. Diatom-inferred oscillations in lake level indicate increased seasonality and variable precipitation from 4180 to 2750 cal yr BP, followed by a dry period with increased nutrient levels and turbidity which may reflect agricultural activity until 1870 cal yr BP. This period is followed by a shift to shallow, well-mixed, open water with lower productivity until modern times.
The diatom record from Lake Wodehouse, a low-elevation grassland swamp located in the Darien of Panama, reflected similar patterns of climate variability and human disturbance (Bush and Colinvaux, 1994). Dry periods inferred from the decline of planktonic Aulacoseira and Cyclotella spp. were observed from 3800–3700 cal yr BP, 3400 to 2500 cal yr BP, and 1900 cal yr BP–modern times. Charcoal fragments, declining arboreal pollen, and maize pollen indicate agricultural activity beginning as early as 3910 cal yr BP, with diatoms reflecting a gradual shallowing and eutrophication of the lake since the start of the record (Bush and Colinvaux, 1994).
Multiple paleolimnological analyses were conducted on Laguna Zoncho, a shallow, slightly alkaline, mid-elevation lake located 65 km southeast of Laguna Los Mangos (Clement and Horn, 2001; Haberyan and Horn, 2005; Taylor et al., 2013, 2015). The Zoncho diatom record revealed a gradual replacement of Aulacoseira and Staurosira with Pinnularia, Eunotia, and Encyonema by 1020 cal yr BP (Haberyan and Horn, 2005). The new assemblage, which persisted until 460 cal yr BP, was indicative of reduced lake depth, acidification, and more stagnant conditions attributed to drought. Pollen and charcoal records reflected intensive agricultural activity predating the sediment record (Clement and Horn, 2001), but agriculture did not appear to increase lake productivity (Haberyan and Horn, 2005; Taylor et al., 2015). Taylor et al. (2015) attributed low sedimentation rates and productivity to agriculture-induced erosion predating the start of the record. Agricultural proxies reflect decline beginning around 1400 cal yr BP and intensifying by 1100 cal yr BP during a period of drought indicated by peak δ2H values (Taylor et al., 2015, 2020). The pollen record reflects forest recovery after 600 cal yr BP following largescale abandonment of cultivated land (Taylor et al., 2020). δ2H values steadily decrease after 600 cal yr BP, indicating increased precipitation during the LIA (Taylor et al., 2020). Increased Aulacoseira abundance reflects a shift to wetter conditions around 460 cal yr BP, which is consistent with forest recovery (Haberyan and Horn, 2005).
Study site and sediment core
Laguna Los Mangos (9.0895°N, 83.4665°W) lies within the Diquís subregion of the Gran Chiriquí archeological region of southern Pacific Costa Rica (Figure 1). Located at an elevation of 475 m in the Fila Costeña mountain range, Los Mangos is one of several lakes formed by landslides within the geological Curré formation comprised of Upper Miocene conglomerates, sandstones, and shale (Horn et al., 2018). The lake had a surface area of 0.3 ha and maximum depth of 0.5 m when it was surveyed in March of 2014 (Johanson et al., 2019). Horn et al. (2018) completed a limnological survey at Laguna Los Mangos and noted it as a polymictic lake that turns over frequently without observed stratification. The surrounding area is classified as tropical moist forest in the Holdridge (1967) system, with an average annual temperature of 24°C and annual precipitation of at least 2000 mm.
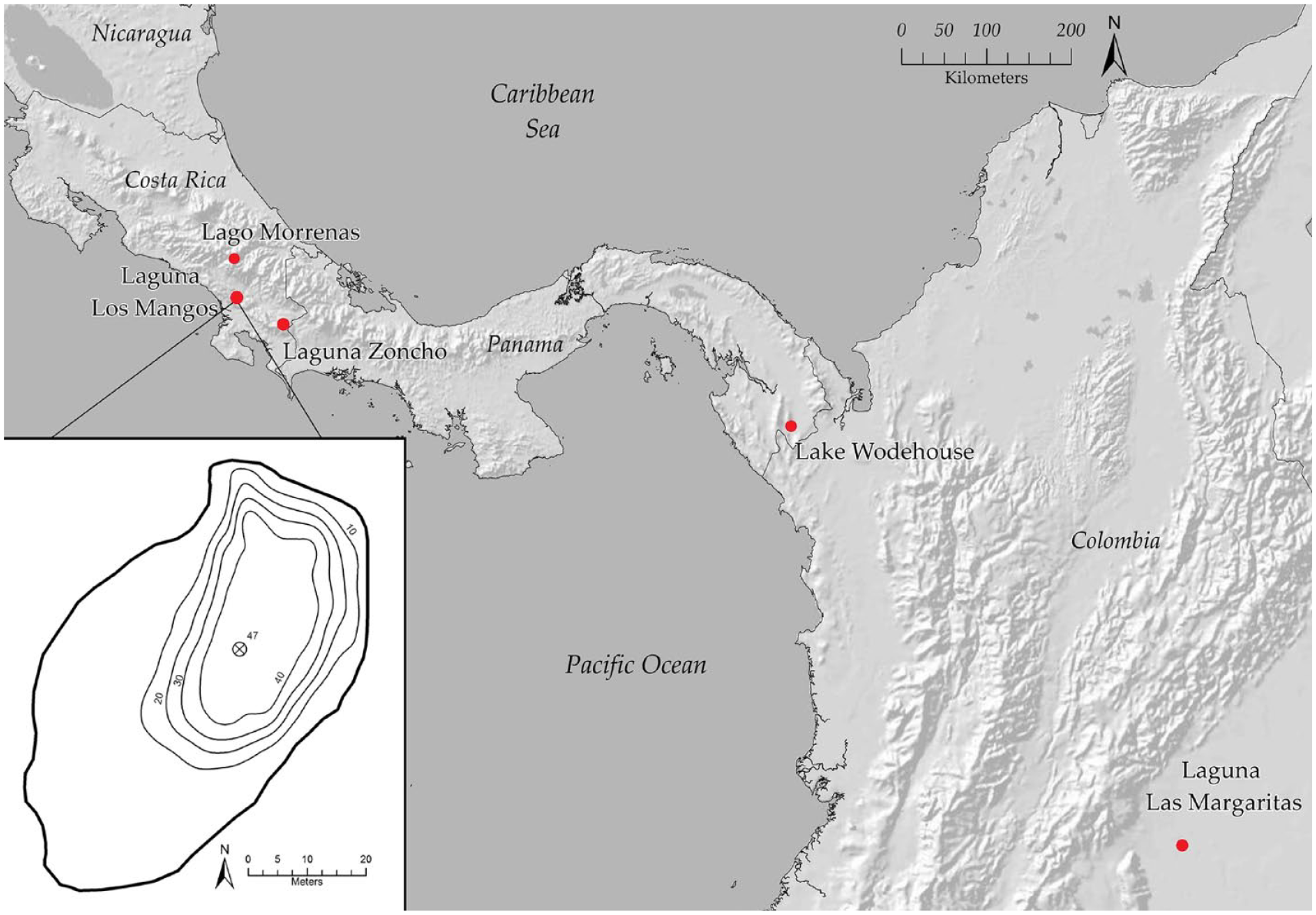
Map of Central America showing the positions of the study site at Laguna Los Mangos and other lakes in Costa Rica, Panama, and Colombia used for comparison. The inset bathymetric map depicts Laguna Los Mangos when it was surveyed in March of 2014 by Johanson et al. (2019). Each isobath line represents a 10 cm increase in depth moving toward the center of the lake, where an X denotes the coring location at 47 cm lake depth.
Johanson et al. (2019) extracted a 7.5 m long core from Laguna Los Mangos in 2014, using successive 1-m drives with a Colinvaux-Vohnout locking piston corer (Colinvaux et al., 1999), beginning 30 cm below the mud-water interface. Recovery was incomplete for some drives. An age model was developed from eight AMS radiocarbon dates on macroremains of wood and leaves (Table 1 and Figure 2) using the CLAM age-modeling package in R (Blaauw, 2010) and the IntCal 13 calibration dataset (Reimer et al., 2013). The model estimates the basal age of the core at ca. 4200 cal yr BP and reveals a hiatus in sedimentation at 192 cm corresponding to ca. 950–450 cal yr BP (Johanson et al., 2019).
Summary of radiocarbon date determinations from the Laguna Los Mangos 2014 core (adapted from Johanson et al., 2019).
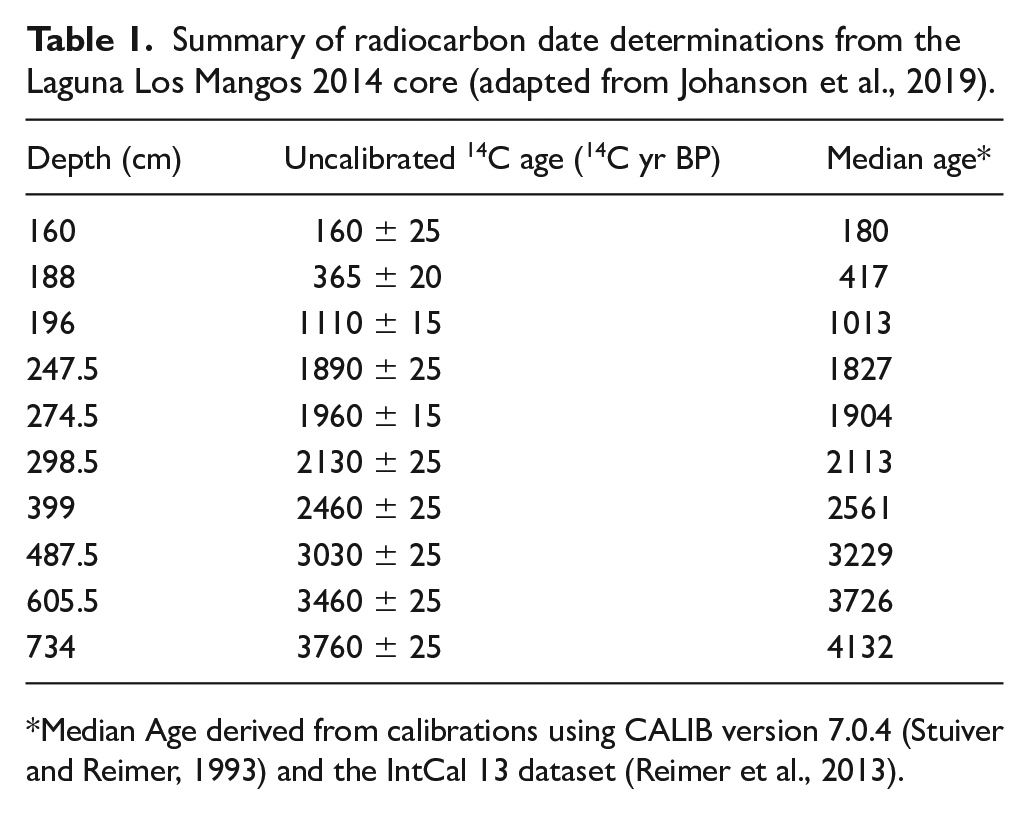
Median Age derived from calibrations using CALIB version 7.0.4 (Stuiver and Reimer, 1993) and the IntCal 13 dataset (Reimer et al., 2013).
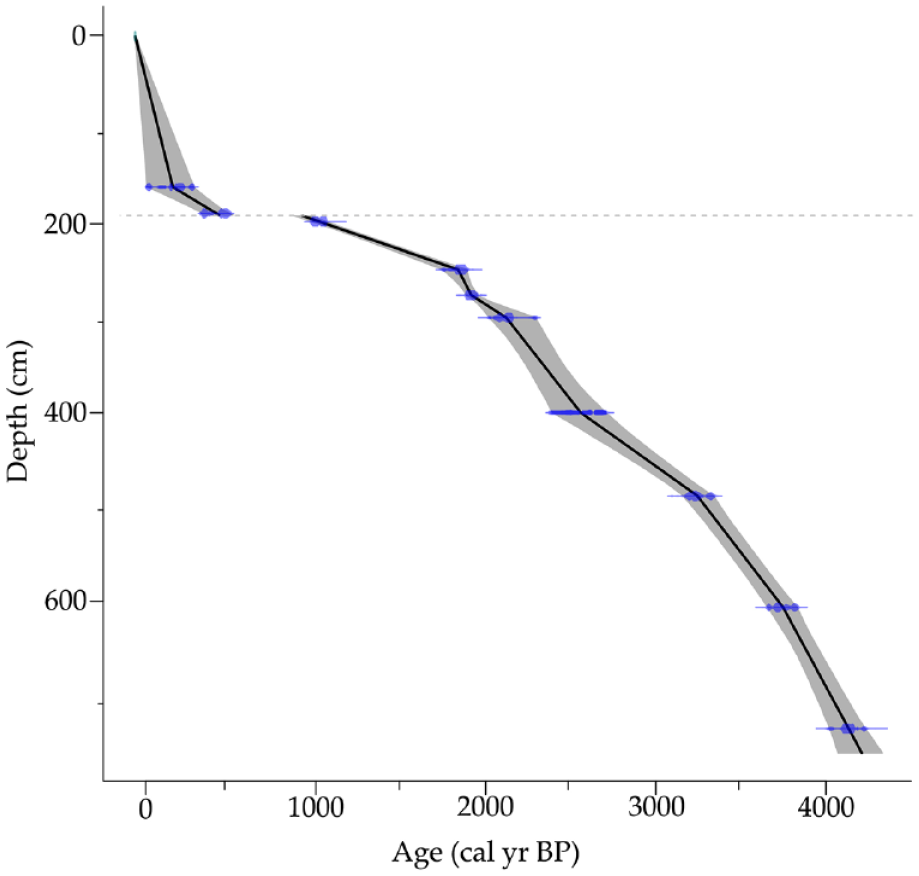
Laguna Los Mangos age-depth model from Johanson et al. (2019). Dashed line indicates sedimentation hiatus at 192 cm spanning ca. 950–450 cal yr BP. Blue dots and shading represent the estimated ages and confidence intervals of radiocarbon-dated macrofossils in the Laguna Los Mangos sediment core. The black line represents the estimated age (cal. yr BP) of core sediments by depth with the confidence interval shown in gray.
Methods
We extracted 38 sediment samples from the Los Mangos core at an average of 17 cm intervals corresponding to an average of 110 ± 127 years between samples. Variability in sampling resolution was due to limited sediment availability following prior analyses. Sampling procedures were adapted from Renberg (1990). We treated 0.1 cm3 of sediment from each depth with 10% HCl for 5 min before adding 30% H2O2 to each. The following day, we placed the samples in a hot water bath at 85°C with additional H2O2, where they remained for 8 weeks until organic material dissolved completely. During the hot water bath treatment, we added H2O2 as needed to prevent sample desiccation. After 8 weeks we rinsed each sample by centrifuging at 3000 rpm for 5 min, decanting, and resuspending the pellet in distilled water, repeating for a total of five rinses. Using plastic pipettes, we added three drops of each sample to coverslips, heated them over medium heat until dry, and mounted them onto glass slides with Naphrax resin on a hot plate at 130°C.
At each depth, we counted 300 valves at 630× magnification under a Zeiss AxioLab compound microscope. Only fragments representing at least one-half of an intact valve were counted. All taxonomic groups were identified to the species level except for Eunotia spp., which were grouped at the generic level because species-level designations were not feasible with the available equipment. Samples for which 300 valves could not be counted were omitted from our analysis to reduce error from selective dissolution. Unidentifiable valves were retained in the total, including valves that were too corroded for confident identification. Paleoenvironmental inferences were based on references from Patrick and Reimer (1966, 1975), Gasse (1986), Krammer and Lange-Bertalot (1986, 1991a, 1991b), Krammer and Lange-Bertlaot (1994), Krammer and Lange-Bertalot (1997), Van Dam et al. (1994), Haberyan and Horn (2023), and the taxonomic nomenclature was updated following Spaulding et al. (2022).
Constrained cluster analysis guided the defining of zones within the diatom record. Taxa with maximum abundance less than 2.5% were omitted to minimize the weight of rare taxa in zone determination. For simplicity, samples with Euclidean distance greater than 50 from adjacent samples were grouped with the adjacent zone whose assemblage was more similar.
Results
Diatom composition
Of the 31 taxa identified in the record, 12 abundant taxa made up >2.5% of the total species composition (Figure 3). Nitzschia incognita, Encyonema silesiacum, Aulacoseira granulata, and Gomphonema affine were the most abundant diatoms through the record, making up 18.9%, 18.2%, 8.7%, and 8.5% of the total species composition, respectively. Only one strictly planktonic species, Aulacoseira granulata, had total average abundance greater than 1%. Eunotia was at least partially represented by E. metamonodon, but it was uncertain if morphological differences between certain individuals were due to taxonomic distinction or morphological plasticity. Laguna Los Mangos diatoms range from oligosaprobes to α-mesosaprobe/polysabrobe intermediates (Krammer and Lange-Bertalot, 1986, 1991a, 1991b, 1997; Krammer and Lange-Bertlaot, 1994; Van Dam et al., 1994). All of the identified diatom taxa are obligate autotrophs except for Diadesmis confervacea and Gomphonema parvulum, which are facultative heterotrophs (Van Dam et al., 1994).

Laguna Los Mangos diatom taxa. Abundant diatoms (>2.5%) in the Laguna Los Mangos sediment profile. Scale bar = 20 μm. (a) Aulacoseira granulata. (b) Diadesmis confervacea. (c) Gogorevia exilis. (d) Lemnicola hungarica. (e) Cocconeis placentula. (f) Staurosira venter. (g) Eunotia spp. (h) Encyonema silesiacum. (i) Nitzschia incognita. (j) Gomphonema parvulum. (k) Gomphonema affine.
Seven depths exhibited poor diatom preservation for which 300 valves could not be identified (specifically 452.5, 468.5, 483.5, 500.5, 612.5, 624.5, and 756.5 cm), resulting in gaps in the record corresponding to ca. 4190, 3790–3760, and 3290–2970 cal yr BP. Valves at these depths were highly corroded, fragmented, and sparse such that species identifications were not feasible. These depths were omitted from the record to reduce potential bias from selective dissolution.
Diatom composition by zone
Four major zones, A–D, were recognized, and the assemblages in three samples (366.5, 516.5, and 724.5 cm) were inconsistent with adjacent samples compared using constrained cluster analysis (Figure 4) to evaluate the role of core depth in diatom composition (Grimm, 1987). Despite considerable differences in diatom composition, these depths were grouped with adjacent zones for simplicity (Table 2).
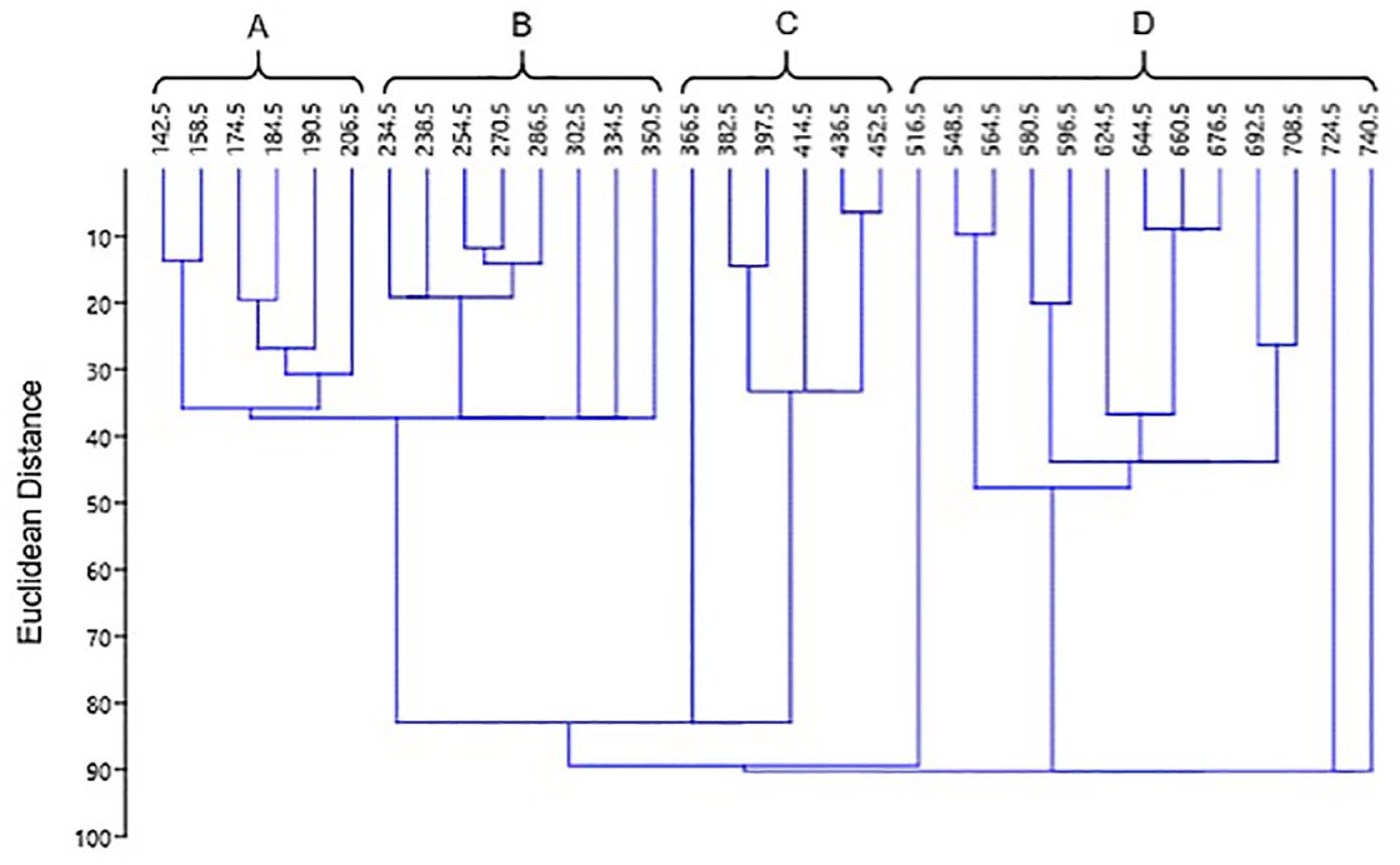
Euclidean similarity index of the Laguna Los Mangos diatom record calculated using the constrained cluster analysis paired groups (UPGMA) technique in PAST (Hammer, 2001). Zones are labeled (A–D).
Summary of diatom zones, most important species, and interpretation from Laguna Los Mangos.
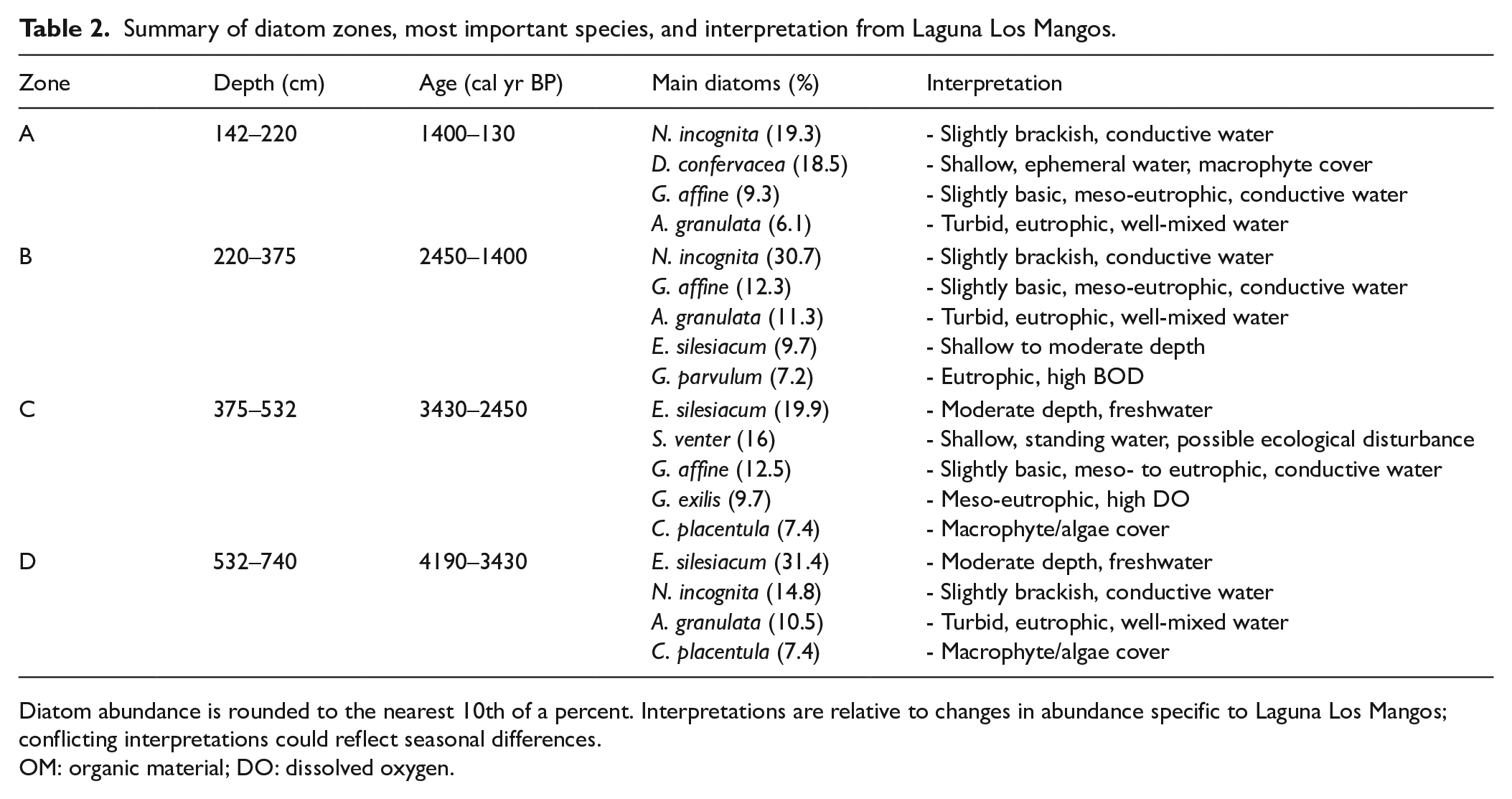
Diatom abundance is rounded to the nearest 10th of a percent. Interpretations are relative to changes in abundance specific to Laguna Los Mangos; conflicting interpretations could reflect seasonal differences.
OM: organic material; DO: dissolved oxygen.
Zone D: 4190–3430 cal yr BP
The record begins with a preservation gap from 4190 to 4140 cal yr BP followed by a sample dominated by A. granulata (76.9%) ca. 4100 cal yr BP (Figure 5), which averages 10.5% of the total diatom abundance in Zone D. Aulacoseira granulata is a planktonic species that prefers shallow, turbid, well-mixed, eutrophic waters where silica and phosphorus are nonlimiting (Kilham and Kilham, 1975; Kilham et al., 1986; Krammer and Lange-Bertalot, 1991a; Krammer and Lange-Bertlaot, 1994). Another smaller peak in A. granulata is observed from ca. 3950 to 3850 cal yr BP and is followed by another period of poor diatom preservation from 3790 to 3760 cal yr BP where diatoms could not be counted.
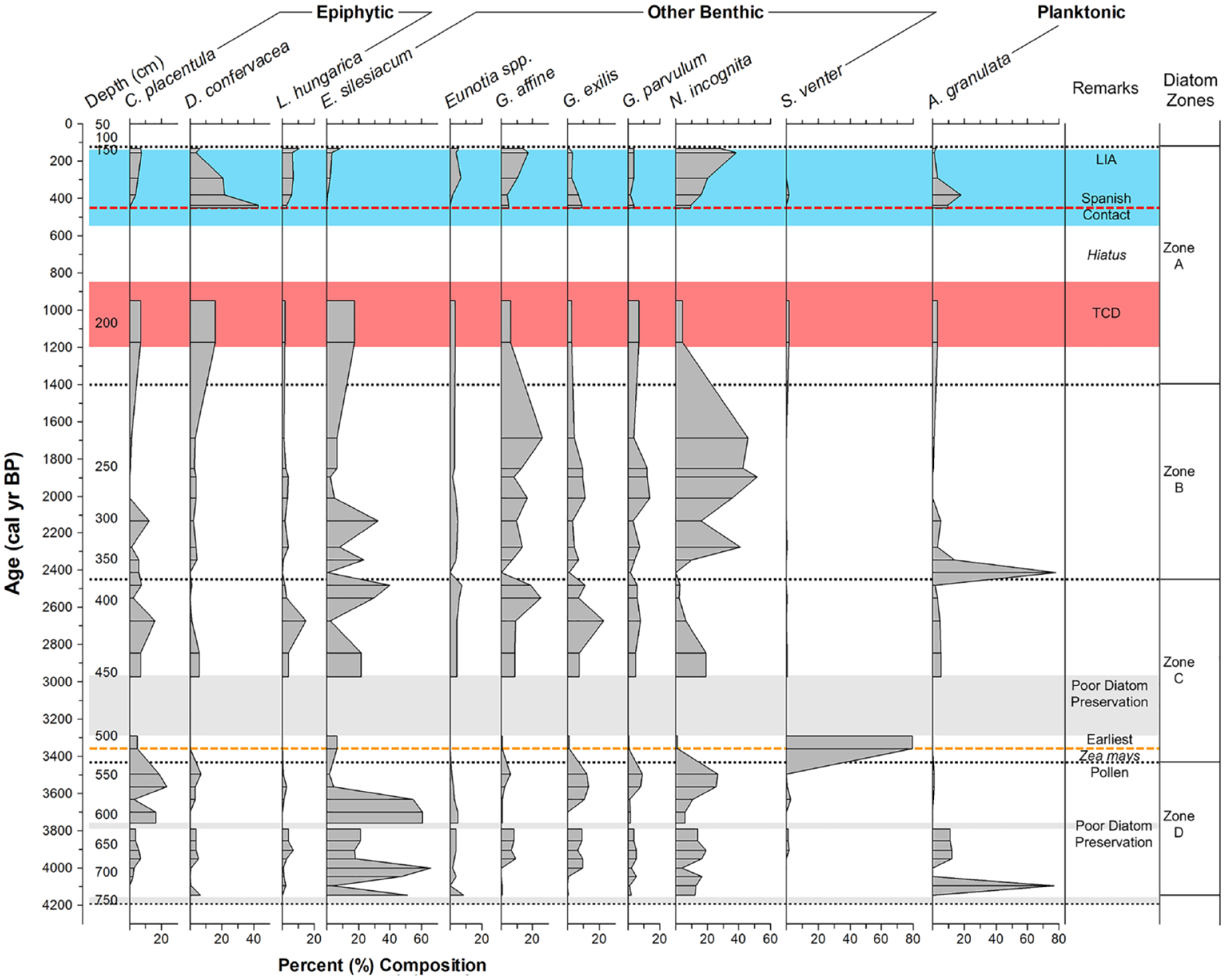
Laguna Los Mangos diatom record. Percent (%) composition of abundant (>2.5%) diatom taxa over time. Blue shading delineates the LIA (ca. 550–100 cal yr BP). Red shading delineates the TCD (ca. 1200–850 cal yr BP). Gray shading indicates periods of poor diatom preservation. Red dashed line indicates the timing of Spanish contact (ca. 450 cal yr BP). Orange dashed line indicates the first appearance of maize (Zea mays) pollen (ca. 3360 cal yr BP) identified by Johanson et al. (2019).
The upper section of Zone D maintains a more consistent community composition dominated by E. silesiacum (31.4%), a eurytopic, pH indifferent, and oligohalobous species (Patrick and Reimer, 1975; as Cymbella minuta var. silesiaca), and N. incognita (14.8%), an epipelic, motile, brackish-fresh water species (Van Dam et al., 1994) that thrives in urban lakes with elevated phosphorus, biological oxygen demand (BOD), and suspended solids (Winter and Duthie, 2000). The genus Eunotia reaches a maximum ca. 4140 cal yr BP. Members of this genus are common in acidic, oligohumic lakes (Kwandrans and Eloranta, 2005). Cocconeis placentula, a cosmopolitan epiphyte found in slow-moving waters (Gasse, 1986; Krammer and Lange-Bertalot, 1991a; Patrick and Reimer, 1966), maintains an average abundance of 7.4% in Zone D and Zone C. Major declines in E. silesiacum and A. granulata are also noted at the end of Zone D, with the latter all but disappearing from the record. In general, Zone D is characterized by periods of poor preservation, sporadic peaks in A. granulata, and dominance of E. silesiacum.
Zone C: 3430–2450 cal yr BP
Zone C begins with an assemblage characterized by peak abundance of Staurosira venter (79.5%) ca. 3360 cal yr BP, a cosmopolitan species that prefers shallow, standing, oligotrophic to mesotrophic waters of medium pH and low mineral content (Bradbury, 1971; Gasse, 1986; Patrick and Reimer, 1966). This assemblage coincides precisely with the first observation of Zea mays pollen in the record by Johanson et al. (2019). Staurosira venter averages 16% abundance in Zone C and is scarce (<1%) throughout the rest of the record. Poor preservation resulted in a large gap in this zone from ca. 3300 to 2970 cal yr BP where no reliable species data could be collected. The remainder of Zone C is dominated by E. silesiacum, averaging 19.9%, and Gomphonema affine, averaging 12.5% composition. Gomphonema affine prefers circumneutral, mesotrophic to eutrophic waters (Patrick and Reimer, 1975). Gogorevia exilis a benthic alkaliphil preferring mesotrophic to eutrophic waters (Lange-Bertalot, 2000; Moro and Fürstenberger, 1997; as Achnanthes exigua), and Eunotia spp. reach their highest average abundance of 9.7% and 4.3%, respectively, during this period. Lemnicola hungarica, an epiphyte that thrives in hypereutrophic conditions (Patrick and Reimer, 1966; Van Dam et al., 1994; as Achnanthes hungarica) peaked in abundance ca. 2670 cal yr BP. Cocconeis placentula maintains the same average abundance of 7.4% from the previous zone into Zone C. In general, Zone C is characterized by a large peak in S. venter, a prolonged preservation gap, maintained dominance of E. silesiacum, and peaks in G. affine, L. hungarica, and G. exilis.
Zone B: 2450–1400 cal yr BP
Zone B begins with an assemblage ca. 2410 cal yr BP characterized by a peak in A. granulata that makes up 78% of the total composition at this depth and averages 11.3% of the total diatom abundance in this zone. The rest of Zone B is dominated by N. incognita, which reaches its highest average abundance of 30.7%, and G. affine, averaging 12.3% composition. Gomphonema parvulum reaches its highest average abundance of 7.2% in this zone, while E. silesiacum decreases to 9.7% abundance. Gomphonema parvulum is a facultative heterotroph (Van Dam et al., 1994) and an α-mesosaprobe/polysaprobe intermediate that has been associated with elevated nutrient levels and BOD (Krammer and Lange-Bertalot, 1986, 1991a; Krammer and Lange-Bertlaot, 1994; Patrick and Reimer, 1975). N. incognita and G. affine reach their maxima in the upper section of Zone B before declining sharply into Zone A. Overall, Zone B is characterized by a large peak in A. granulata, dominance of N. incognita and G. affine, and high but declining abundance of E. silesiacum and G. parvulum.
Zone A: 1400–130 cal yr BP
Nitzschia incognita remains dominant in Zone A with average composition of 19.3% and is closely followed by Diadesmis confervacea, an epiphytic, aerophilic α-mesosaprobe known to thrive in shallow, marshy environments (Krammer and Lange-Bertalot, 1997; Ruwer and Rodrigues, 2018; Van Dam et al., 1994), which increases to its peak average abundance of 18.5%. A sedimentation hiatus suggestive of dry conditions occurred in the middle of Zone A from ca. 950 to 450 cal yr BP, for which period no proxy data could be collected. Another small spike in A. granulata (17.8%) is observed ca. 380 cal yr BP, with average composition of 6.1% in Zone A. In general, Zone A is characterized by peak abundance of D. confervacea, dominance of N. incognita, and a prolonged period of lake desiccation spanning the end of the TCD.
Discussion
Temporal variability in diatom composition and preservation are consistent with that of other records by Johanson et al. (2019) indicating extensive landscape disturbance from deforestation, agricultural activity, erosion, and fire throughout the record. The presence of maize pollen at ca. 3360 cal yr BP is a clear indicator of human occupation by this time (Johanson et al., 2019), but earlier occupation by hunter-gatherers or small-scale horticulturalists is also possible. δ13CTOC represents the ratio of carbon-13 to carbon-12 of total organic carbon (TOC), which increases with drought and the relative abundance of C4 plants including maize and agricultural weeds. Plants that use the C4 photosynthetic pathway make up only a small portion of the native flora in Costa Rican forests, and increased δ13CTOC values indicative of increased C4 plant abundance can be interpreted as evidence of either drought or the presence of maize and associated agricultural weeds (Johanson et al., 2019; Lane et al., 2004). When maize pollen is scarce, δ13CTOC can be a useful proxy for agricultural activity (Lane et al., 2009). Charcoal peaks indicating fire activity around 4000 cal yr BP, deforestation beginning around 3700 cal yr BP, and consistently increasing δ13CTOC beginning around 3800 cal yr BP all occurred during a period of higher-than-average precipitation prior to the earliest maize presence (Johanson et al., 2019; Yanuskiewicz et al., 2022) and may constitute evidence of earlier human presence at Los Mangos. Yanuskiewicz et al. (2022) used compound-specific stable carbon and hydrogen isotopes alkanes with specific chain lengths (δ13CC29, δ2HC29) as proxies for vegetation composition and precipitation, respectively, at Laguna Los Mangos. δ13CC29 aligned closely with δ2HC29 prior to 3360 cal yr BP, indicating that vegetation composition was primarily influenced by precipitation until maize agriculture was introduced (Figure 6). Paleolimnological studies based in the Americas show that human settlement increases lake input of nutrients and sediments and reduces water clarity as a result of deforestation and increased erosion (Clement and Horn, 2001; Dubois et al., 2018; Haberyan and Horn, 2005; Johanson et al., 2019; Vélez et al., 2005; Velez et al., 2011, 2021).
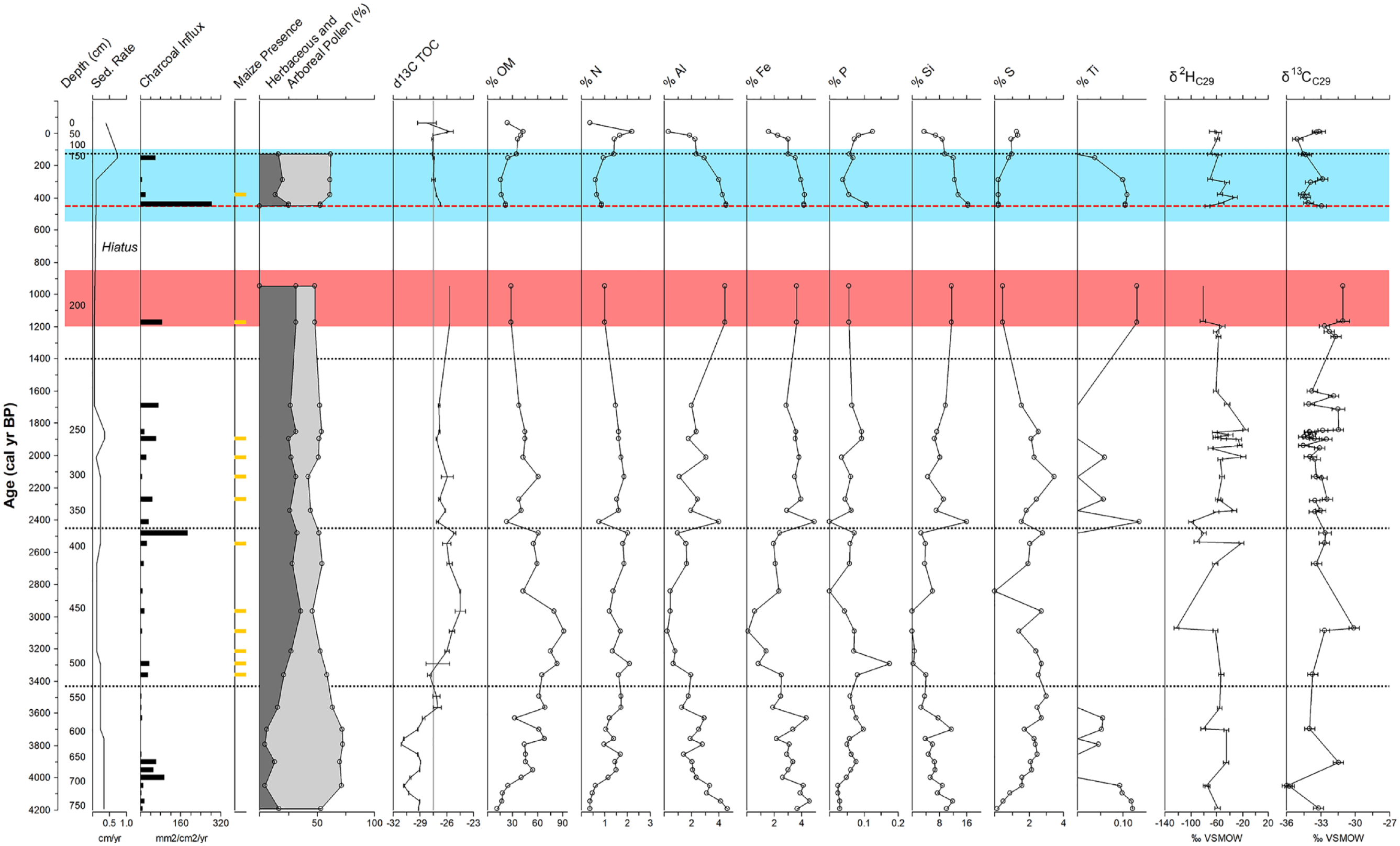
Previously published proxy records from Laguna Los Mangos. Sedimentation rate (cm/yr), charcoal influx (mm2/cm2/yr), pollen composition, organic material (%OM), nitrogen (%N), C:N ratio, elemental composition, and δ13CTOC records are from Johanson et al. (2019). δ2HC29 and δ13CC29 records are from Yanuskiewicz et al. (2022). Herbaceous pollen types included Poaceae, Cyperaceae, Asteraceae and Amaranthaceae, while arboreal/shrub types included Urticales diporate and triporate, Hedyosmum, Ficus, Burseraceae, Myrsine, Alchornea (Euphorbiaceae), Piper, Melastomataceae/Combretaceae, Cecropia, Sapium (Euphorbiaceae), Anacardiaceae, Ulmus, Myrica, Alnus, Podocarpus, Weinmannia, and Quercus. Blue shading delineates the LIA (ca. 550–100 cal yr BP) and red shading delineates the TCD (ca. 1200–850 cal yr BP). Red dashed line indicates the timing of Spanish contact. Orange dashed line indicates where maize (Zea mays) pollen was first observed in the record. LIA: Little Ice Age; TCD: Terminal Classic Drought.
Zone D: 4190–3430 cal yr BP
Variable but increasing values of δ13C of total organic carbon (δ13CTOC), carbon to nitrogen ratios (C:N), Poaceae pollen, nitrogen (N), organic material (OM), sedimentation rates, and peaks in charcoal influx indicate considerable landscape instability attributed to deforestation, erosion, and fire activity in the beginning of the record (Johanson et al., 2019; Figure 6). Alternating peaks in E. silesiacum and A. granulata are interspersed with periods of poor diatom preservation ca. 4190, 3790–3760, and 3290–2970 cal yr BP, reflecting ecological variability in response to unstable environmental conditions.
Deviations between δ13CC29 (Yanuskiewicz et al., 2022) and δ13CTOC (Johanson et al., 2019) ca. 4190, 4050, 3700, and 3360 cal yr BP may have resulted from variable algal productivity and its differential effects on δ13CTOC (Yanuskiewicz, 2019). Periods with low diatom abundance align with or closely follow deviations of δ13CTOC from δ13CC29, which may reflect a change in limnological conditions that reduced diatom productivity. Elevated aluminum, iron, and titanium concentrations aligning with peak sedimentation rates in the beginning of the record (Johanson et al., 2019) likely reflect landscape instability and heavy erosion following the landslide that caused lake formation. Landslide-induced erosion could explain gaps in diatom preservation from 4190 to 4140 cal yr BP, 3790 to 3760 cal yr BP, and 3290–2970 cal yr BP as a result of heavy sediment deposition “diluting” valve density in lake sediments, reduced light availability limiting diatom productivity, or both. Heavy input of N, P, and OM may have similarly hindered diatom preservation during the latter preservation gap when maize cultivation was first implemented.
The δ2HC29 record from Laguna Los Mangos reflects increased precipitation from 4050 to 3900 cal yr BP, which is followed by a 200-year period with drier conditions (Yanuskiewicz et al., 2022). This dry period coincides with increases in charcoal, titanium, iron, sulfur, and sedimentation rate as well as reduced arboreal pollen, indicating increased fire activity, deforestation, and heavy erosion, which may be interrelated. Peaks in A. granulata, which prefers turbid, well-mixed, eutrophic waters (Kilham et al., 1986; Krammer and Lange-Bertalot, 1991a; Krammer and Lange-Bertlaot, 1994), were observed ca. 4100 cal yr BP and 3950–3850 cal yr BP in alignment with increased charcoal influx, sedimentation, and metal(oid) deposition. An analysis of diatom composition and chemical and limnological characteristics of 86 Costa Rican lakes found that A. granulata abundance was related to shallow depth, low to moderate inflow/outflow, and elevated sulfur concentrations (Haberyan and Horn, 2023). Peak average abundance of E. silesiacum, which reportedly prefers moderate depth, low concentrations of magnesium and sodium, and high concentrations of sulfur (Haberyan and Horn, 2023), was observed in Zone D. Alternating peaks of S. venter and A. granulata likely reflect changes in lake mixing, depth, turbidity, and solute concentration, with A. granulata indicating more turbulent, shallow, and concentrated conditions ca. 4100 cal yr BP and 3950–3850 cal yr BP.
Zone C: 3360–2450 cal yr BP
Poaceae (grass) had replaced arboreal Melastomataceae/Combretaceae and Ficus as the dominant pollen type by 3360 cal yr BP when maize (Zea mays) pollen was first observed in the record (Johanson et al., 2019). Native vegetation was cleared with fire to cultivate maize, which altered the landscape and produced clear signals in records of δ13CTOC, pollen, and charcoal after 3360 cal yr BP (Johanson et al., 2019). Increased δ2HC29 indicates moderate drought at the start of this zone until ca. 3070 cal yr BP.
Zone C began with a sample dominated by S. venter, likely reflecting shallow, standing water at this time. Staurosira venter is known as a pioneer species early to colonize shallow, slightly alkaline lakes with heavy input of clastic material (Haworth, 1976). Wojtal and Sobczyk (2012) also reported that S. venter increases with conductivity and nitrate concentration. Proliferation of this species may have occurred in response to heavy erosional input, agriculture-induced eutrophication, reduced lake depth, or a combination of these factors.
This sample was followed by a core section with poor diatom preservation spanning 3290–2970 cal yr BP that showed peak values of nitrogen, phosphorus, organic material, and δ13CTOC, indicating heavy nutrient loading from agricultural runoff (Johanson et al., 2019; Yanuskiewicz et al., 2022). Phosphorus has a well-documented association with primary productivity (Conley et al., 2009; Ramstack Hobbs et al., 2016), but diatom productivity declines in favor of cyanobacteria when silica becomes a limiting nutrient (Battarbee, 1978). Silica concentrations declined to 0% as phosphorus reached peak concentration and diatom abundance plummeted, which may reflect a regime shift favoring cyanobacteria as agricultural activity intensified. Agriculture-induced nutrient runoff, organic input, and erosional deposition may have induced decline in diatom abundance by diluting valve concentrations in lake sediments, reducing light availability in the water column, or driving a regime shift favoring cyanobacteria. Drought can cause increases in nutrient and ion concentrations, turbidity, and even water temperature in affected lakes (Brasil et al., 2016; Howard and Noble, 2018; Li et al., 2018). Drought-induced decline in lake level has been reported to select for motile Nitzschia spp. (Klamt et al., 2020) and cyanobacteria when drought is more severe (Brasil et al., 2016; Howard and Noble, 2018). Dry climate likely intensified the effects of intensive maize cultivation observed during this period.
Zone C shows relatively similar diatom composition to Zone D with increases in G. affine, G. exilis, and L. hungarica following the preservation gap reflecting a meso- to eutrophic system with increased productivity, pH, and aquatic vegetation cover (Moro and Fürstenberger, 1997; Patrick and Reimer, 1966, 1975; Simkhada, 2006; Van Dam et al., 1994). Lemnicola hungarica is associated with duckweed, which is common in ponds affected by eutrophication and/or shading (Emson et al., 2018). In abundance, duckweed can cause anoxia by blocking light and oxygen from the water column below (Janes, 1998).
Increased diatom abundance following the period of poor preservation coincided with a shift to wetter conditions ca. 3070–2670 cal yr BP (Yanuskiewicz et al., 2022) likely due to increased inflow, lake volume, and dilution of agricultural runoff. Increased precipitation likely supported increased agricultural activity inferred from charcoal influx, reduced arboreal pollen, increased δ13CC29, and the presence of maize pollen during this period (Johanson et al., 2019; Yanuskiewicz et al., 2022).
Zone B: 2450–1400 cal yr BP
Increases in A. granulata, titanium, silica, aluminum, iron, and sedimentation rate around 2400 and 2100 cal yr BP follow a large peak in charcoal influx at ca. 2500 cal yr BP (Johanson et al., 2019). These trends reflect landscape instability caused by seismic disturbance, fire induced erosion, or both. Sharp declines in δ13CTOC, OM, and N ca. 2400 cal yr BP indicate the start of a gradual agricultural decline which intensified by 1840 cal yr BP, when increased δ2HC29 indicates a 250-year period of drought (Johanson et al., 2019; Yanuskiewicz et al., 2022). Nitzschia incognita, an epipelic diatom which prefers higher conductivity, turbidity, P, and salinity (Van Dam et al., 1994; Winter and Duthie, 2000), increases during this dry period, which may reflect increased solute concentration from lake drying and/or input of clastic material from a seismic disturbance. Gomphonema parvulum, which increases in the beginning of Zone B, is a facultative heterotroph (Van Dam et al., 1994) and an α-mesosaprobe/polysaprobe intermediate that prefers eutrophic waters with elevated BOD (Krammer and Lange-Bertalot, 1986, 1991b; Krammer and Lange-Bertlaot, 1994; Moro and Fürstenberger, 1997; Patrick and Reimer, 1975). This species reaches peak abundance in this zone before decreasing into Zone A, indicating productive, eutrophic water as the lake dried and became more concentrated. L. hungarica and C. placentula prefer higher nutrient levels and macrophyte cover, and they also decline in abundance in this zone.
Zone A: 1400–130 cal yr BP
In the beginning of Zone A ca. 1400 cal yr BP, the abundance of D. confervacea increased while N. incognita and G. affine suddenly declined. The desiccation of Los Mangos from ca. 950 to 450 cal yr BP is consistent with dry climate during the TCD, after which more negative δ2HC29 values indicate periods of increased moisture in the beginning (ca. 450–430 cal yr BP) and the end (ca. 300 cal yr BP–present) of the LIA. Diadesmis confervacea is an aerophilic epiphyte known to thrive in shallow, marshy environments (Krammer and Lange-Bertalot, 1997; Ruwer and Rodrigues, 2018; Van Dam et al., 1994) that is associated with macrophyte cover (Haberyan and Horn, 2023) and high levels of organic nitrogen (Kilroy et al., 2017). The increase in this species from ca. 1200 to 450 cal yr BP despite reduced lake levels throughout the TCD likely indicates that macrophyte cover increased as lake level declined. The ability of Diadesmis confervacea to survive outside of waterbodies may explain its competitive advantage over the other epiphytic diatoms, C. placentula and L. hungarica, which were low in abundance before and after the sedimentation hiatus during the TCD. Diadesmis confervacea later decreased in abundance while C. placentula and L. hungarica increased around 450 cal yr BP, coinciding with a small peak in A. granulata and likely wetter conditions at Laguna Zoncho (Haberyan and Horn, 2005; Wu et al., 2019). This may indicate increases in lake level and floating aquatic vegetation during the LIA. Charcoal, pollen, and δ2HC29 indicate a large fire event contemporaneous with the arrival of the Spanish ca. 450 cal yr BP. Spanish conquest was followed by a period of forest recovery and wetter conditions reflected by increased abundance of arboreal pollen and the resurgence of submerged diatom taxa. These signals of the TCD/LIA transition at Los Mangos align with a dry period indicated by reduced Aulacoseira spp. abundances in Laguna Zoncho from ca. 1020 to 460 cal yr BP and the wetter period that followed (Haberyan and Horn, 2005).
Constrained cluster analysis indicates that samples at 158 cm (ca. 150 cal yr BP) and 142 cm (ca. 130 cal yr BP) were more similar to each other than to samples corresponding to dates prior to ca. 290 cal yr BP in Zone A. Those samples from the earlier section of Zone A show higher similarity in diatom composition with the periods before and after the TCD than with the two most recent samples, implying a shift in lake conditions during the middle of the LIA. This may be a result of the shift to wetter conditions indicated by more negative δ2HC29 values ca. 300 cal yr BP (Yanuskiewicz et al., 2022) and/or modern recolonization following agricultural collapse attributed to Spanish contact.
General trends and comparison to other studies
The Laguna Los Mangos diatom record reflects a shallow, alkaline, eutrophic system affected by agriculture-induced nutrient loading, climate-induced changes in lake level and concentration, seismically-induced deposition events, and gradual sediment infilling and declining lake depth throughout the Late-Holocene. These factors contribute to a diatom record with highly variable species composition and preservation. Periods of poor diatom preservation and low sampling resolution limit the accuracy of zone divisions, the detection of brief shifts in species composition, and the interpretation of environmental conditions during periods where diatoms are scarce. Despite these limitations, we utilize other proxy records in conjunction with the diatom record to provide a more detailed interpretation of past environmental conditions at Laguna Los Mangos.
The Costa Rican landscape is highly unstable due to steep slopes and seismic activity, with earthquakes and landslides resulting in high levels of erosion and sediment flux especially during intense precipitation events (Alvarado et al., 2008; Krishnaswamy et al., 2001; Mora, 1989). Laguna Los Mangos was formed by a landslide (Horn et al., 2018), indicating landscape instability at the start of the record. Elevated metal(loid) concentrations and higher sedimentation rates reported around 4100, 2400, and 450 cal yr BP align with peaks in A. granulata and are indicative of erosion amplified by landscape instability, fire activity, and changes in precipitation. Aulacoseira granulata reportedly thrives in turbid, eutrophic, well-mixed waters where silica is nonlimiting (Kilham and Kilham, 1975; Kilham et al., 1986; as Melosira granulata). Bush et al. (2016) noted a similar pattern in A. granulata and charcoal influx at Lake Sauce in Peru, suggesting that lake inflow increased as the landscape became more open. Intensive burning causes increased erosion and lake deposition of iron and copper as charred vegetation and sediments previously secured by roots are carried downstream (Dunnette et al., 2014; Pelletier et al., 2020). These and other metal(loid)s were elevated in the beginning of the Los Mangos record, which may have resulted either from fire activity prior to the start of the record or from the landslide that caused lake formation (Johanson et al., 2019). Later peaks in iron, copper, aluminum, titanium, silica, and A. granulata all coincide with peaks in charcoal and shifts in δ2HC29, reflecting a dynamic interaction of fire, deforestation, and erosion during periods of drought. It is important to consider that most peaks in A. granulata were present in only a few isolated samples, so it is possible that other peaks in this species were missed during periods with low sampling resolution.
Severe environmental disturbances often decrease biodiversity by producing harsh conditions tolerable by a limited number of species (Chase, 2007). Drastic reduction in diatom diversity associated with peak abundance of S. venter and major decline in diatom abundance indicate severe environmental and ecological disturbance to the lake and surrounding watershed caused by intensive agricultural activity. Smaller disturbances reflected by A. granulata peaks, brief preservation gaps, and evidence of increased terrestrial input ca. 4100, 3950, and 2410 cal yr BP likely reflect deposition events associated with landslides and possibly seismic activity. Diatom preservation gaps associated with these deposition events were generally smaller, briefer, and more frequent than that associated with agriculture, which also appeared to drive more permanent shifts in diatom composition and lake chemistry at Laguna Los Mangos.
Eunotia and Encyonema silesiacum are the only abundant (>2.5%) taxa in the record that are oligohalobous (Patrick and Reimer, 1975; Van Dam et al., 1994), and they are absent in samples with peak A. granulata abundance. Haberyan and Horn (2005) noted a similar pattern in these taxa in Laguna Zoncho in which Aulacoseira, which indicated increased pH and water column mixing, alternated with Encyonema and Eunotia minor, which were suggested to indicate reduced pH, productivity, and turbulence. Nitzschia incognita, which gradually increases at the expense of E. silesiacum through the record, reportedly thrives in slightly brackish (Van Dam et al., 1994) urban lakes with elevated phosphorus, BOD, and suspended solids (Winter and Duthie, 2000). Throughout the record, E. silesiacum is gradually replaced by N. incognita, signaling an increase in conductivity and nutrient content and declining lake level since its formation. These trends are common in lakes heavily influenced by human activity because fire, deforestation, and agriculture increase the input of nutrients and sediments into the surrounding watershed (Anderson, 2014; Dunnette et al., 2014). Periodic drought likely amplified this trend by reducing lake depth and increasing conductivity and nutrient concentrations.
The diatom record reflects variability in lake depth, macrophyte cover, pH, nutrient levels, and solute concentrations in response to nutrient loading, declining forest cover, frequent burning, and heavy erosion indicated by other proxy records following lake formation. Periods of poor diatom preservation were attributed to heavy erosional input possibly caused by seismic activity and the landslide that caused lake formation. Poor diatom preservation attributed to unstable lake conditions was also reported at Las Margaritas during this period (Vélez et al., 2005), which may reflect regional seismic activity.
Evidence of early human settlement and agriculture at Laguna Los Mangos, Laguna Zoncho, and Lake Wodehouse aligned with inferred increases in fire activity, erosional input, and deforestation (Bush and Colinvaux, 1994; Haberyan and Horn, 2005; Johanson et al., 2019). At Lake Wodehouse, pollen and charcoal evidence indicate agricultural activity beginning as early as 3910 cal yr BP. Charcoal peaks ca. 4000 cal yr BP at Laguna Los Mangos occur during a period of increased precipitation, which could indicate an earlier human presence prior to the establishment of maize agriculture near the lake. Variability in pollen composition, nutrient levels, elemental composition, and elevated sedimentation rates during this period could be attributed to human presence, climate variability, landscape instability, or a combination of these factors.
Diatom assemblages from Los Mangos and Las Margaritas were more variable than those from Zoncho and Morrenas 1, which are deeper and located at higher elevation. This pattern could be a result of increased sensitivity of shallower lakes to shifts in climate and environmental conditions. A study comparing diatom assemblages in deep and shallow lakes throughout Guatemala, Panama, and Colombia by Velez et al. (2021) reported that assemblages from shallow lakes changed gradually over time and were more responsive to subtle environmental changes, while deeper lakes showed little change before exhibiting a quick shift into a novel assemblage. Velez et al. (2021) attributed the responsiveness of shallow lakes to higher rates of nutrient cycling and more complete water column mixing. Pronounced variability in the Los Mangos diatom record may be attributed to the combined effects of climatic variability, deforestation, agricultural activity, and heightened sensitivity of shallow lakes to climate and environmental changes.
Records from Los Mangos, Wodehouse, and Las Margaritas all reflect variable precipitation beginning around 4200 cal yr BP, possibly signaling a change in seasonality. Laguna Los Mangos was the only one of the previously mentioned lakes to dry out completely during the TCD. Intensive deforestation at Laguna Los Mangos may have contributed to an earlier onset and higher intensity of drought during the TCD, as suggested by Oglesby et al. (2010) and Cook et al. (2012). Drought and agricultural decline beginning ca. 1840 cal yr BP likely supported forest recovery around the lake as fields were abandoned. Persistent dry conditions led to the desiccation of Laguna Los Mangos from 950 to 450 cal yr BP, after which more negative δ2HC29 values indicate increased moisture at the beginning (ca. 450–430 cal yr BP) and the end (ca. 300 cal yr BP–present) of the LIA. This is inconsistent with other records that show longer droughts during the LIA with severity comparable to the TCD (Haug et al., 2001; Koch et al., 2019; Lane et al., 2011; Ledru et al., 2013). Forest cover increases and maize disappears from the record after 400 cal yr BP, likely due to severe decrease in Amerindian populations caused by diseases associated with Spanish conquest. A similar shift to wetter conditions during the LIA was noted at Laguna Zoncho by 460 cal yr BP, which was a suggested response to forest regrowth following Spanish conquest (Haberyan and Horn, 2005; Taylor et al., 2020; Wu et al., 2019). Forest recovery prior to and during the LIA likely contributed to wetter microclimates associated with increased canopy cover during this period (Johanson et al., 2019; Yanuskiewicz et al., 2022). Differences in the timing of drought onset during the TCD and LIA at these lakes may also be attributed to the variable, site-specific effects of elevation, ENSO events, shifts in the ITCZ, and other climate forcing mechanisms.
Footnotes
Acknowledgements
We thank E. Yanuskiewicz and C. Lane for sharing isotope data, and A. Oleinik, C. Shadik, and M. Bush for guidance on the analysis and interpretation of the Laguna Los Mangos diatom record.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We would like to thank the Florida Atlantic University Department of Geosciences and the Paleoenvironmental Change Specialty Group of the American Association of Geographers for supporting this research. Radiocarbon dating of the Los Mangos core was supported in part by the National Science Foundation under grant #1660185.