
Abstract
While few places on earth have been as deeply impacted by the human-horse relationship as the steppes of Mongolia and eastern Eurasia, gaps in the archaeological record have made it strikingly difficult to trace when and how the first domestic horses were integrated into ancient societies in this key region of the world. Recently, organic materials preserved in melting mountain ice have emerged as a key source of archaeological insight into the region’s deep past. Newly-identified artefacts recovered from melting snow and ice in the Altai Mountain range of western Mongolia (including metal artefacts, skeletal remains, and hoof fragments) provide archaeological evidence for the use of horses at high altitudes from the Bronze Age through the 20th century. Direct radiocarbon dating and genomic sequencing demonstrate the presence of Przewalski’s horse in the region during the early second millennium BCE, suggesting that this taxon may have once foraged at high altitudes frequented by human hunters. Importantly, directly-dated remains of horse hoof trimmings provide some of the oldest direct evidence of horse transport in the Eastern Steppe as early as the 14th century BCE, and suggest a role for high-mountain hunting in the innovation of reliable mounted riding.
Introduction – horse domestication and dispersals in East Asia
Despite the broad impact of later Mongolian horse cultures on global history and prehistory, archaeological insights into the earliest chapters of the human-horse story in Mongolia are scarce. Using interdisciplinary analysis of recent discoveries from high altitude archaeological survey, this article seeks to understand the role of wild and domestic horses in early human lifeways in the Altai, and contextualize their implications for the broader trajectory of horse domestication in East Asia. While debates continue over the possibility of earlier domestication events of Przewalski’s horse (Gaunitz et al., 2018; Taylor and Barrón-Ortiz, 2021), recent comprehensive genomic studies demonstrate that the origins of modern domestic horses can be traced to western Eurasia, when the first ancestors of what is known as the DOM2 lineage first appear in the archaeological record by the late third millennium BCE (~4200 years ago), before dispersing rapidly across much of Inner Asia (Librado et al., 2021). In Mongolia, early pastoralists appear to have migrated to the region as early as ca. 3000 BCE (Kovalev and Erdenebaatar, 2009; Wilkin et al., 2020), but the first domestic horses appear much later. The earliest directly dated domestic horses in the Mongolian steppes date to ca. 1200 BCE, when horse remains are appear at sites of a culture known as the Deer Stone-Khirigsuur (DSK) culture, named for anthropomorphic stone monuments (deer stones) and burial mounds (khirigsuurs) that emerged across a wide swath of eastern Eurasia centred on the Mongolian steppes. The DSK culture included ceremonial horse burials sometimes numbering in the hundreds (Allard and Erdenebaatar, 2005). Archaeozoological and biomolecular data show that these late second and early first millennium BCE horses were bridled and used for transport (Taylor et al., 2015), provided with veterinary care (Taylor et al., 2018), consumed for meat (Houle, 2010) and used for milk (Ventresca Miller et al., 2022; Wilkin et al., 2020). Some skeletal evidence on DSK horses suggests that they could have been used for early mounted horseback riding (Taylor and Tuvshinjargal, 2018), while other contextual data demonstrate a clear role for chariot use in DSK culture (Taylor et al., 2021a). Recent biomolecular study indicates that these animals were close relatives of early domestic horses, known as the DOM2 lineage from western Eurasia (Fages et al., 2019), and widespread appearance of domestic horses at DSK sites appears to accompany the initial dispersal of domestic horses into Shang-period China, where horses appear primarily in elite chariot burials during the final second millennium BCE (Rawson et al., 2021; Taylor et al., 2017).
The rapid appearance of domestic horses in DSK ritual features could be linked with their initial introduction into the eastern steppe (Taylor et al., 2017), but arguments have been made for the likelihood of an earlier introduction (Esin et al., 2021; Honeychurch, 2015). Isolated skeletal elements of equids predating the DSK period are sometimes recovered from archaeological sites (Houle et al., 2022; Janz et al., 2017; Kovalev, 2014), but the limited nature of the region’s early archaeofaunal record, absence of quality reference collections for morphological identifications, and the lack of reliable morphological criteria for distinguishing wild lineages from domestic, have prevented a conclusive assessment of the timing of early domestic horse use in Mongolia.
Glacial archaeology
Over recent years, mountain ice melting due to warming climate has emerged as an important source of archaeological insights into Mongolia’s earliest pastoral cultures. Around the globe, snow and ice features known as ‘ice patches’ – permanent non-glacial features often located on shaded slopes in high-latitude and high-altitude areas – have begun to melt at extraordinary rates, in the process exhuming ancient biological and cultural materials that provide a key record of human activity (Dixon et al., 2005). This record has proven to be particularly significant in Mongolia, where material culture is biased towards organic materials, but taphonomic conditions rarely favour good organic preservation except in dry or frozen contexts (Clark et al., 2021). In western Mongolia, initial surveys of glaciers and ice patches yielded some surprises about the dynamics of early pastoral subsistence, including the recognition that wild game hunting, particularly of large game such as argali sheep (Ovis ammon), was an important subsistence activity from the DSK period through the modern era (Taylor et al., 2021b).
In other cold environments across the ancient world, such as regions of southern Norway, survey of ice patches has provided a rare snapshot into the use of domestic horses, revealing skeletal remains and horse equipment such as special snowshoes that reveal the use of mountain passes for transport and commercial activity (Pilø et al., 2020). Poor survey coverage of melting cultural features, and continued melt of snow and ice features, raises the possibility that archaeological discoveries at ice patches and glaciers could provide snapshots into the use of domestic horses in Mongolia’s earliest prehistory.
Materials and methods
Archaeological survey
In the 2022 field season, we conducted full-coverage survey of melting snow and ice margins at five localities in the immediate vicinity of Tsengel Khairkhan, located in Sagsai sum of Bayan-Ulgii province, western Mongolia (Tsengel Khairkhan glacier, Tsengel Khairkhan Terrace 1, Tsengel Khairkhan Terrace 2, Khulzuut Ice Patch 3, and Tsengel Lower Patch 1). With a team of four archaeologists, we conducted pedestrian survey of snow and ice margins along the glacier’s western and northern margin, as well as two permanent ice patches (Terrace 1, first studied by Taylor et al., 2021a, 2021b, and Terrace 2, newly surveyed in 2022). Team members conducted survey in 5 m transects, beginning from the ice margin at the time of survey and radiating outward until no artefacts or ecofacts were recovered in three consecutive transects.
For each faunal specimen recovered from the ice, we conducted morphological identifications using limited comparative collections of domestic animal remains at the National Museum of Mongolia, alongside a large database of photographs and 3D models of comparative wild and domestic animals in collections housed in the Archaeozoology Laboratory at the University of Colorado Museum of Natural History. Specimens identified as horse were then sampled for radiocarbon dating using Accelerator Mass Spectrometry in the Department of Earth Sciences at the University of California-Irvine and genomic identification in the Centre for Anthropobiology and Genomics at the University of Toulouse in Toulouse, France. Prior to destructive sampling, we produced a high-resolution textured 3D scan of each specimen using a structured light scanner (EinsScanSE for smaller objects, and Artec LEO for larger cranial specimens). All artefacts recovered during the analysis are housed at the National Museum of Mongolia, Ulaanbaatar, Mongolia.
Radiocarbon dating
Preparation of bone and tooth samples followed Shammas et al. (2019) following mechanical surface cleaning if required, samples of 200 mg of bone or dentin were crushed to ~1 mm powder and decalcified overnight in a measured amount of 1 N HCl calculated as just sufficient to dissolve the entire sample if all of the material was hydroxyapatite. They were then washed with MQ water, gelatinized overnight at 60°C in 0.01 N HCl, ultrafiltered in precleaned Vivaspin 15 devices to select a high molecular weight fraction (>30 kDa), and freeze dried.
Samples of 1.5–2 mg of pretreated organics were combusted with CuO and Ag wire getter at 900°C in quartz tubes sealed under vacuum, graphitized by hydrogen reduction at 525°C with an iron powder catalyst (Santos et al., 2007), and the Fe-graphite mixture was pressed into Al sample holders. Radiocarbon was measured at the Keck Laboratory at University of California Irvine on a National Electrostatics 1.5SDH Compact AMS system (Southon et al., 2004). Radiocarbon ages are shown as conventional 14C ages corrected for isotopic fractionation, with 1 sigma uncertainties that reflect scatter in repeated runs and uncertainties in the measurement of blanks and normalizing standards (NIST Oxalic Acid 1, SRM4990B) as well as counting statistics. Aliquots of ~0.7 mg of ultrafiltered collagen for stable isotope ratio and elemental (%C and %N) analyses were placed in tin capsules and measured using a Fisons NA 1500NC elemental analyser and Finnigan Delta Plus isotope ratio mass spectrometer, at precisions of ±0.1% and ±0.2% for δ13C and δ15N, respectively. All dates produced quality control metrics, including C:N ratio and collagen yield that meet even strict suggestions for dating in Mongolian horse samples (Zazzo et al., 2019). Radiocarbon ages were calibrated using the INTCAL20 calibration curve (Reimer et al., 2020) in OxCal for ancient specimens, and using CALIBOMB for 20th century specimens (Reimer and Reimer, 2010).
Ancient DNA sequencing
The archaeological remains from sample TK101 consisted of tooth material, and was prepared in the ancient DNA facilities of the Centre for Anthropobiology and Genomics of Toulouse (CNRS/Université Paul Sabatier). DNA was extracted from 330 mg of osseous powder, following the procedure from Boessenkoel and colleagues (Boessenkool et al., 2017), including bleach pre-treatment and a short pre-digestion step. More specifically, the osseous powder was incubated with bleach (0.5%) for 10 min at room temperature under agitation, before being washed three times using 1 ml of sterile water. The bone powder was centrifuged for 30 s at 12,000 rpm in order to remove the bleach/water supernatant, and was dried out at room temperature after the third final washing step. The bleach-treated and cleaned powder was then incubated with agitation in 3.85 ml of extraction buffer at 37°C for 60 min, before being collected following a centrifugation step as described above, and resuspended in the same volume of a fresh extraction buffer. It was left for digestion at 42°C on a rotating rotor overnight until complete digestion. Following DNA purification, a total of 22.8 μL of DNA extract was treated with the USER enzyme (0.235 units/μL; New England Biolabs©) as described by Fages et al. (2019) and 14.9 μL of the USER-treated extract was used for library construction, using 25 μL as a final reaction volume. The procedure followed Librado et al. (2021), and included the ligation of adapters containing 7 bp-long indexes following the sequence of the sequencing PCR primer, as originally described by Rohland et al. (2015). Library amplification followed the methodology from Librado et al. (2021) using the Accuprime Pfx Taq polymerase, and resulted in the addition of a supplemental, 6 bp-long, index as part of the Illumina P7 adapter. Therefore, individual DNA libraries were characterized by a combination of three unique indexes, which annihilates possible index hopping, thus cross-contamination, following read demultiplexing (van der Valk et al., 2020). A total of 12 PCR cycles were carried out within 25 μL reactions, using 3 μL of the unamplified DNA libraries. Amplified DNA libraries were purified using Agencourt AMPure XP reagents (1.4:1 bead-to-library ratio) and eluted in 25 μL of EB buffer supplemented with 0.5% Tween-20. Aliquots consisting of 1/10th dilutions were quantified and size profiled on the Qubit fluorimeter (HS dsDNA assay, ThermoFischer) and TapeStation 4000 (Agilent) instruments, using the Qbit 1X dsDNA HS Assay and the High Sensitivity DNA ScreenTape analysis kits, respectively. DNA libraries were further pooled at equimolar concentration before shallow sequencing on the Illumina MiniSeq instrument (Paired-End, 101 × 2 cycles), providing sufficient sequence data to estimate endogenous DNA content and assessing DNA mis-incorporation and DNA fragmentation profiles with mapDamage2 (Jónsson et al., 2013), following index demultiplexing and read mapping against the horse EquCab3 reference genome sequence (Kalbfleisch et al., 2018), as described below. A total of 2 DNA libraries were prepared from the DNA extract obtained following one extraction session, before carrying out deep-sequencing on the Illumina NovaSeq 6000 instrument (S4 lane, 100 × 2 cycles) at Genoscope.
DNA sequence processing
Paired-end DNA sequencing reads were demultiplexed, and collapsed when showing sufficient overlap (–minadapteroverlap 3 – 5 mm), using AdapterRemoval2 (Schubert et al., 2016) tolerating one mismatch at best on each internal 7 bp-long indexes (–barcode-mm-r (Taylor et al., 2021a) 1), trimmed for low-quality ends (–trimns), and filtering out templates shorter than 25 bp. Both reads that could be collapsed in single templates, and those that could not be collapsed, were aligned against the horse nuclear (EquCab2 (Wade et al., 2009), and; EquCab3 (Kalbfleisch et al., 2018), supplemented by the Y-chromosome contigs from Felkel et al., 2019), and mitochondrial reference genomes (Accession nb: NC_0016440), using Paleomix v1.2.13 (Schubert et al., 2014), as described by Taylor et al. (2023). Sequence alignments corresponding to PCR duplicates and showing mapping quality scores strictly below 25 were disregarded. Average depth-of-coverage estimates were calculated using the Paleomix coverage module, ignoring read groups (Sample TK101, coverage = 0.923X). The EquCab2 nuclear genome alignments were further processed through Zonkey (Schubert et al., 2017) to confirm that the archaeological material derived from the horse species and to identify their molecular sex; according to this procedure, sample TK101 was identified as a horse male. The EquCab3 nuclear genome alignments were used in all other analyses described below, together with those obtained using the same methodology for a comparative panel of ancient horses selected to encompass the whole range of genetic lineages, both extant and extinct, hitherto characterized (Table S1 and Figure S6). The genome alignments of two domestic donkey individuals, one ancient specimen (AC14415, from Acemhoyük) and one modern specimen (ETH5B, from Ethiopia), were added as outgroups, using the sequence data from Todd et al. (2022). The list of 445 BAM genome alignments was processed through ANGSD (build December, 10th 2022, htslib 1.16 (Korneliussen et al., 2014)), as described by Taylor et al. (2023), to obtain a matrix including 10,395,964 transversion SNP variation across all 31 horse autosomes, in which each sample was pseudo-haploidized (i.e. the sequence of one random-read was considered at those sites covered more than once). We required a maximum missingness of 80% per site, and excluded sites corresponding to the 99.5% percentile and above of the cumulated coverage distribution across all individuals, in order to avoid mis-alignments resulting from possibly unidentified structural variants. The final matrix of SNP variation was produced in the tped format, and was further converted in eigenstrat format using Plink v1.90b6.24 (Chang et al., 2015) (www.cog-genomics.org/plink/1.9/) and convertf (v4510, from the Eigensoft package (Patterson et al., 2006)).
Molecular phylogenetic reconstructions
Horse mitochondrial genome alignments were generated following the methodology from Librado et al. (2021), disregarding sites showing coverage below 5 and base qualities below 30. Mitochondrial alignments were further masked for the repeat region located between the positions 16,124 and 16,359 of the NC_0016440 reference. The sequence of the complete mitochondrial genome of a modern donkey individual was downloaded from Genbank (Accession nb: CM027722.2), and aligned manually using Seaview (v5.0.5 (Gouy et al., 2010)) to provide an outgroup. Maximum Likelihood phylogeny was reconstructed using IQtree (v1.6.12 (Minh et al., 2020)), selecting the best substitution statistical model (GTR + F + R10) out of a total number of 286 possible models on the basis of the AICc information criterion. Node supports were assessed using ultrafast bootstrap (UFBoot) approximation (Hoang et al., 2018) and the SH-like approximate likelihood ratio (aLRT) test (Guindon et al., 2010) with 1000 pseudo-replicates (Figure S1A). Y-chromosome alignments for males (N = 259) were processed following the same procedure as that described above for the autosomes, conditioning on those regions identified as single copy by Felkel et al. (2019) and those sites showing at most 50% missingness. We then used the resulting 15,578 polymorphic sites to construct a Neighbour-Joining phylogeny with fastme (v2.1.6.2 (Lefort et al., 2015)), and assessed bootstrap support from a total of 100 bootstrap pseudoreplicates (Figure S1B). We followed the same methodology to reconstruct a Neighbour-Joining phylogeny of the autosomal genome-wide variation, using the matrix of 10,395,964 transversion sites (Figures 4a and S7). All molecular phylogenies were manually rooted using the donkey outgroup(s).
Population structure
Two Principal Component Analyses (PCA) of the autosomal variation were carried out in smartPCA (v16000 (Patterson et al., 2006)), after removing the donkey outgroups using Plink v1.9 (–remove option). In both analyses, the PCA was constructed from the genetic variation encapsulated by the comparative genome panel, projecting the ancient specimens characterized in this study (lsqproject: YES), and turning on the ‘inbreed’ flag while considering no outliers (numoutlieriter: 0). In the first analysis, the autoshrink option was turned off (data not shown), while it was turned on in the second analysis (Figures 4 and S5). Both analyses returned consistent projections of the ancient individuals considered.
We further explored the genomic makeup of all individuals present in our dataset, using unsupervised Admixture (v1.3.0 (Alexander et al., 2009)) and considering 2–10 admixture components (Figures 4d and S4). The optimal number of ancestry components (K = 6) was selected using a 10-fold cross-validation procedure. Additionally, all individual pairwise f3-Outgroup statistics in the form of (X,Y:Outgroup) were calculated using qp3Pop (v420, from the Admixtools package (Patterson et al., 2012)), first considering the two donkeys as outgroups (Figures 4b and S2). The analysis was repeated excluding donkeys, and considering the two divergent Late Pleistocene North American specimens as a closer outgroup (Figures 4 and S3).
Results
Archaeological survey results yielded both faunal remains and artefacts linked with use of domestic horses in high mountain environments. At Tsengel Khairkhan (Figure 1), we recovered iron horseshoes (Figure 2a and b) indicative of domestic horse use in two hillside contexts higher than 3300 m. The absence of dateable material associated with the finds make them difficult to date at finer resolution than the Iron Age or after (after ca. 500 BCE). Stylistic comparison of these finds shows that they are comparable in form and technology to modern and early historic horseshoes from western Mongolia, although without a reliable or dated typology, it is difficult to make further inferences regarding their age. At the nearby locality of Ice Patch 3 at Khulzuut, 2022 survey also recovered an iron tool melting directly from the ice margin that appears to be a hoof pick (Figure 2c), a specialized tool used to clear debris from the sensitive, V-shaped and concave sole area underneath the hoof itself known as the ‘frog’. No material was recovered with the hoof pick that could provide a direct date, although previous survey at this locality identified a camel-hair artefact interpreted as a possible horse bridle, which was radiocarbon-dated to the fifth or sixth century CE (Taylor et al., 2021b), and in 2022, a bone/antler projectile with arrow shaft was found nearby dating to ca. 758–520 cal. BCE (2 sigma calibrated range, 2475 ± 15 14C YBP, UCIAMS-268497). These radiocarbon data from nearby artefacts shows that human activity at the site may extend across most of the Iron Age, from at least the first millennium BCE through the first millennium CE and into the modern era.
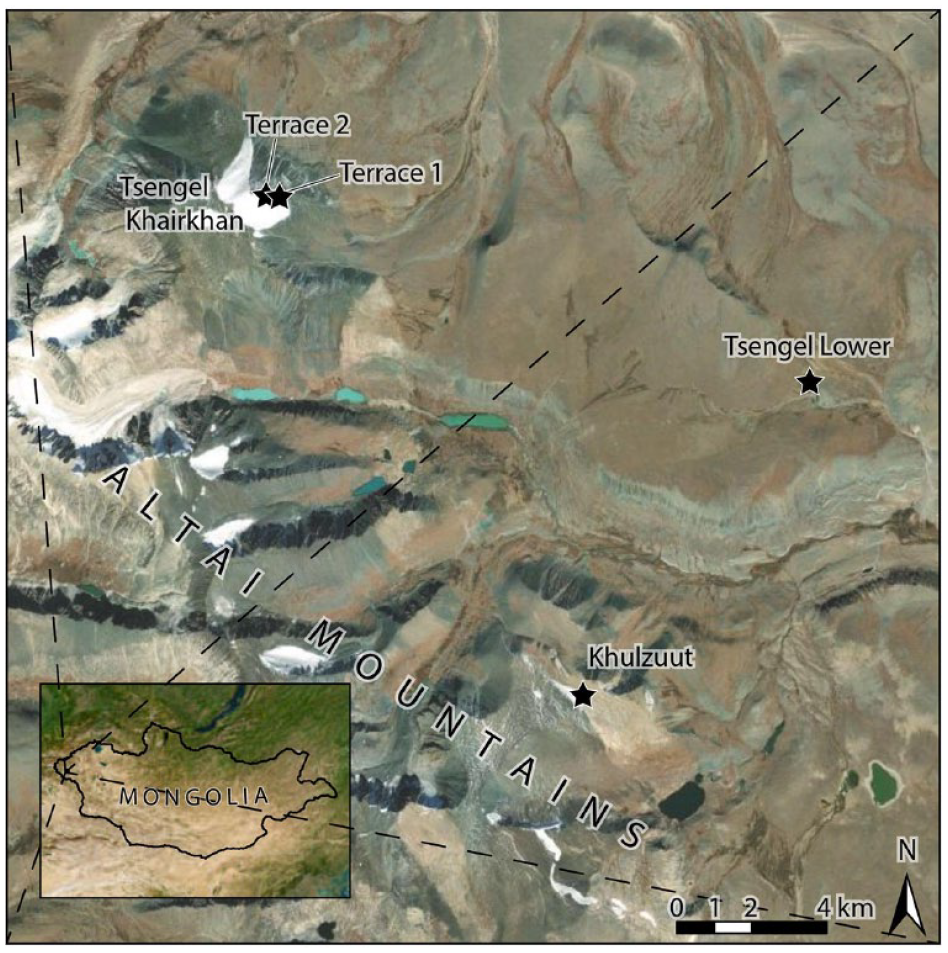
Site locations in the study area in western Mongolia. Sources: Esri, DigitalGlobe, GeoEye, i-cubed, USDA FSA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, and the GIS User Community.

Horse equipment identified during a 2022 survey, including complete horseshoe and partial horseshoes from Tsengel Khairkhan (a and b) and likely hoof pick from Khulzuut (c).
In addition to horse equipment, we identified organic remains of domestic horses at three localities – Tsengel Khairkhan Glacier, Tsengel Khairkhan Terrace 2 and Tsengel Lower Patch 1. These included an intact upper right second premolar (P2) of a juvenile horse (TK Glacier), an apparent horse hoof trimming (TK Terrace 2) and a nearly-complete horse cranium and mandible (Tsengel Lower Patch 1).
Tsengel Lower Patch 1
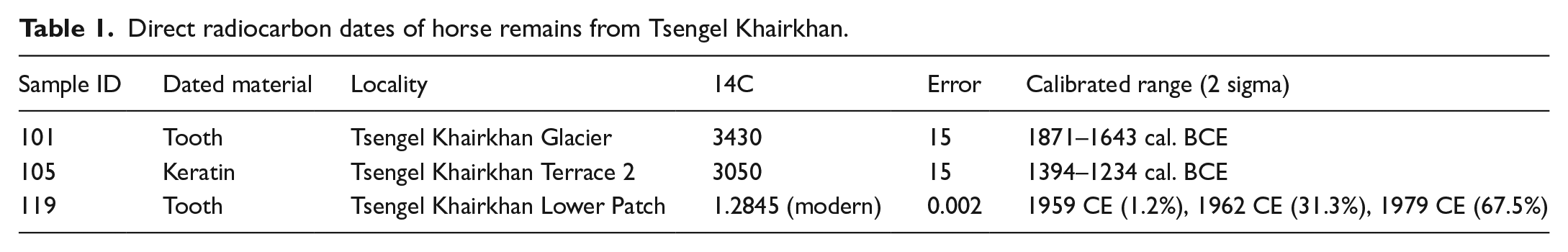
We identified a horse cranium (Figure 3a) along with two-halves of a mandible melting from the ice margin at the lowest-altitude persistent ice patch in the study region (~3000 m elevation) along the first drainage east of Tsengel Khairkhan mountain. Based on the eruption of the teeth, the specimen was an adult male horse, while crown height measurements from the two loose maxillary teeth (P2 and P3, ~28 and 29 mm, respectively) suggest that the animal was between 12 and 15 years at the age of his death (Levine, 1982). Undulations on the occlusal surface of the tooth row show that the horse experienced ‘wave mouth’, a form of malocclusion related to uneven dietary wear, while erosion of the cementum on the P2 points to the use of this horse in transport with a metal bit (Bendrey, 2007). Radiocarbon dating of this specimen demonstrated it to be 20th century in origin, with the measured fraction of modern carbon of 1.2845 ± 0.002 placing the animal’s likely death around either the year 1962 (31.3% probability) or 1979 (67.5%) (Table 1).
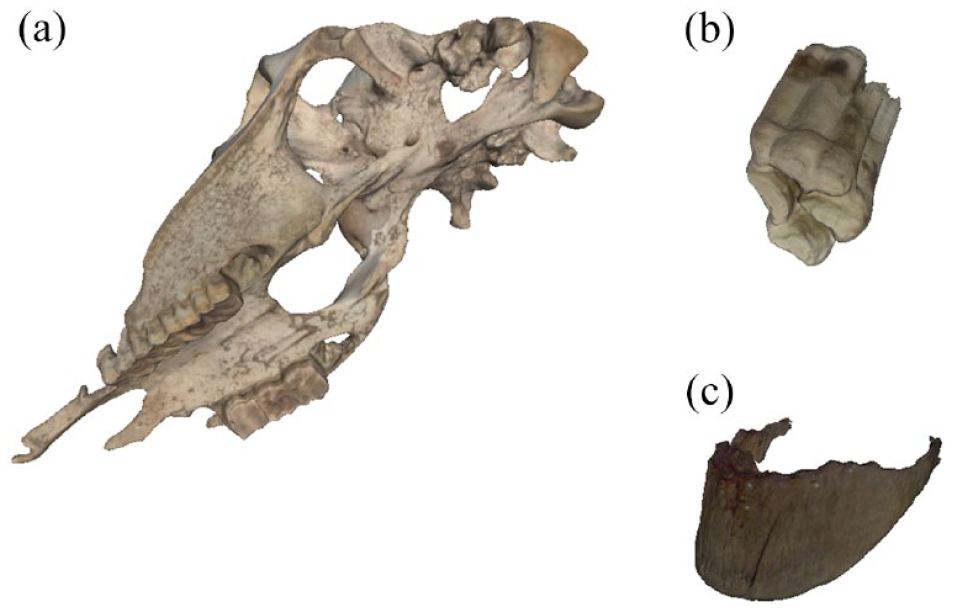
Horse remains identified from high altitude ice patches near Tsengel Khairkhan during 2022 fieldwork, include a horse cranium (a), an isolated premolar (b) and a hoof trimming (c).
Direct radiocarbon dates of horse remains from Tsengel Khairkhan.
Tsengel Khairkhan Glacier
Along the melting western margin of Tsengel Khairkhan glacier, we recovered a single upper second horse premolar (specimen TK101, Figure 3b). The crown height of the specimen (67 mm) indicates that it originates from a horse younger than 5 years old (Levine, 1982), while the lack of dietary wear on the tooth indicates that the animal was likely less than 3 years old at the time of death. Direct radiocarbon dating of this specimen produced a date of ca. 1871–1643 cal. BCE (3430 ± 15 14C years BP), placing it among the oldest finds ever identified from a glacial/ice patch context in Mongolia.
Tsengel Terrace 2
Finally, we recovered a single horse hoof specimen (Figure 3c) from the upper terrace (Terrace 2) at Tsengel Khairkhan, melting from the ice margin alongside other artefacts and ecofacts, including an iron-tipped arrow and remains of wild bovids (O. ammon and several finds of a smaller bovid comparable to Saiga tatarica). Given the mixed nature of most glacial archaeological contexts, it is unlikely that the physical proximity of these objects necessarily represents their cultural association. This hoof specimen shows evidence of cultural modification in form of divots along the proximal edge that truncate the natural surface of the keratin, many of which have a scalloped shape (Figure 5). Under magnification, the shear nature of these modified surfaces suggest that they are culturally-modified rather than the result of natural damage. The most likely explanation for this unique artefact is that it is the leftover trimming from an overgrown or damaged distal section of hoof keratin which was modified by a human visitor to the glacier, with the resulting clipped keratin material ultimately discarded on the ice. Direct radiocarbon dating of this specimen places it in the second half of the second millennium BCE, with a calibrated date range of ca. 1394–1234 cal. BCE (Table 2).
Genomic identifications of sex and species for horse specimens recovered during field survey. Zonkey analyses were limited to the shotgun DNA sequence data generated on the Illumina MiniSeq instrument, during DNA quality screening. The sex identification of sample TK105 was inconclusive, due to the over-representation of some autosomes, especially chr27 and chr31, possibly indicating karyotypic abnormalities in this specimen, disqualifying the use of X-to-autosomal ratios of read alignments as a sex molecular diagnostic. No high-quality read-alignments were obtained against the Y-chromosome scaffolds from Felkel et al. (2019).

Genomic analysis
We conducted DNA sequencing of all ancient horse remains identified during archaeological survey. DNA preservation levels were compatible with the genome characterization of sample TK101 by shotgun sequencing at 0.923-fold average coverage-of-depth, following read quality and duplicate filtering. In order to clarify genetic affinities, the sequence data were compared to a panel including all horse genetic lineages previously identified, consisting of whole genome sequences from 345 ancient and 95 modern horse individuals, plus both an ancient and a modern donkey individual, both used as outgroups (Table S1). Stringent quality filters resulted in the characterization of ~10.4 million transversion SNPs for which at least 80% of the 445 genomes present in the panel were covered. Phylogenetic reconstructions indicated that sample TK101 belonged to a highly-supported mitochondrial clade including both early DOM2 horses (CD1819), Przewalski’s horses as well as horses culturally associated with the Botai and Krokalev cultures (Figure S1A). Additionally, sample TK101 carried a Y-chromosome haplotype belonging to a highly-supported phylogenetic clade consisting of non-DOM2 horses, including, but not restricted, to horses culturally associated with the Botai culture (Figure S1B). Phylogenetic reconstructions based on autosomal DNA sequence variation confirmed the absence of phylogenetic affinities to the DOM2 horse lineage and their closest relatives (CPONT and TURG; Figures 4a and S7). In fact, sample TK101 clustered within a group of horses including Przewalski’s horses, Botai horses, and their close genetic relatives associated with the Krokalev culture and the Borly4, NeoBorly and Novolinka sites (Botai-like, hereafter). Combined, these sites span the third and fourth millennium of Central Asia and SouthWest Siberia. Przewalski’s horses appear phylogenetically the closest, especially one specimen from the 19th century CE. These phylogenetic affinities were confirmed using f3-Outgroup statistics (Figures S2 and S3), in which pairs involving sample TK101 and either Botai-like or Przewalski’s horses showed maximal values (Figure 4b and c). Moreover, the Admixture (Alexander et al., 2009) ancestry profile of sample TK101 was found to only consist of a balanced mixture of two ancestries maximized in Botai-like and Przewalski’s horses (Figures 4d and S4). This sample also projected at an intermediate position between these two groups of horses along the three first Principal Components summarizing the autosomal variation found across our comparative panel (excluding the 2 extremely divergent Late Pleistocene North American horses; Figure 4e and f). Sample TK101 remained close to Botai-like and Przewalski’s horses along the fourth and fifth components (Figure S5A and B). Combined, these analyses establish that TK101 as belonging to the Przewalski’s horse lineage.

Sample TK101 genetic affinities. (a) Neighbour-Joining phylogenetic reconstruction based on autosomal transversion SNPs. Node support was calculated on the basis of 100 bootstrap pseudo-replicates. Labels refer to the sample name, their previously identified population assignment and their age (negative if BCE, Before Common Era, and positive otherwise) (Table S1). The tree was manually edited for maximizing readability, by collapsing the node defining the DOM2 phylogenetic cluster. The full tree is shown in Figure S7. (b) Pairwise f3-Outgroup statistics involving sample TK101 and any other horse present in the comparative genome panel, using 2 donkeys as outgroup. (c) Pairwise f3-Outgroup statistics involving sample TK101 and any other horse present in the comparative genome panel, using 2 divergent Late Pleistocene North-American horse specimens as outgroups. (d) Admixture (Alexander et al., 2009) genetic profiles. The number of genetic components (K = 6) was the best returned amongst those possible values considered using cross-validation (K = 2–10). (e) Principal Component Analysis (PCA) based on autosomal transversion SNPs. The graph reports the PCA projection of sample TK101 on a panel of 440 horse specimens, considering the first 2 principal components. The fraction of the overall variance represented by each axis is reported between parentheses. (f) Same as panel (e), for the second and third principal components.
While genomic preservation of our other two horse samples was not sufficient for high-coverage sequencing, analysis of these specimens using the Zonkey pipeline (Southon et al., 2004) suggests that both horses are caballine horses rather than donkey, mule, or hemione (Table 2), and confirms 2 specimens as males (Supplemental Appendix 1; Table 2). The sex molecular diagnostic was found inconclusive for the remaining specimen (TK105).
Discussion
Results from pedestrian survey provide multiple lines of evidence for the use of wild and domestic caballine horses at high altitude in the Mongolian Altai at each of our three main site clusters, ranging between 3000 to 3700 m above sea level. Genomic data from frozen specimens capture important changes in the human-horse relationship across the late Bronze Age. High altitude zones are a unique place to encounter horse remains – feral domestic horses have been in some cases, observed to avoid high altitudes and unvegetated slopes (Linklater et al., 2000). Although contemporary Mongolian people do practice the tradition of placing horse skulls atop stone cairns in prominent hilltops (Marchina et al., 2017), the difficulty of moving a horse cranium upslope to the glacier and scree of Tsengel Khairkhan makes predator or scavenger transport to these extreme high-mountain zones unlikely. Our genomic identification of a horse specimen belonging to the Przewalski’s horse lineage and dating to the early second millennium BCE at this site therefore suggests that either A) these horse remains were brought to the mountaintop by humans for ceremonial purposes, or that B) the horses themselves made use of high-elevation zones during the early second millennium BCE, where they may have been exploited by human hunters.
Most importantly, our high-altitude survey provides among the earliest evidence for domestic horse use in eastern Eurasia, showing the use of horses in mountain hunting and transport since the 13th/14th centuries BCE. While assemblages at lower altitude sites like Tsengel Lower might reflect herding activities, at higher sites (Khulzuut and Tsengel Khairkhan) the majority of archaeological material is conclusively associated with the hunting of wild game, the primary activity represented in the artefact assemblage (Taylor et al., 2021b). The extreme terrain, low/nonexistent vegetation, and volatile weather conditions of higher elevations at Tsengel Khairkhan are inimical to the pasturing of domestic animals, and even reaching the area on horseback requires extraordinary intentionality. Undateable artefacts such as iron horseshoes from mountain slopes and a hoof pick melting directly from the ice at Khulzuut show that at least in the modern era and perhaps deeper into the Iron Age, hoof maintenance was a critical component of ancient horse transit into these mountain zones, where lost footing on rocky scree and icy slopes can quickly spell disaster.
Based on direct radiocarbon dating, our horse hoof clipping recovered from the ice margin at Terrace 2, at a height of ca. 3700 m, likely predates all other domestic horse remains previously identified in the Mongolian Plateau at ca. 1394–1234 BCE (Figure 5). Previous Bayesian modelling efforts using only direct dates on domestic horses (Esin et al., 2021) modelled the early boundary for the introduction of domestic horses to the region at ca. 1200 BCE. With a measured date of 3050 (±15) radiocarbon years and modification apparently caused by hoof trimming, TK105 is the oldest directly-dated horse specimen with evidence for human management yet known in Mongolia, although other associated dates from human remains without correction for possible reservoir effects have also been reported from the 13th or 14th century BCE (Ventresca Miller et al., 2022; Tumen et al., 2012). Genomic data from this hoof confirm its identification as a caballine horse, though DNA preservation levels were not sufficient to test clustering within the DOM2 genetic lineage. Most importantly, the artefact itself shows direct evidence of human modification and care of horse hooves likely associated with transport activities. This find demonstrates that domestic horses were present in the Mongolian Altai immediately preceding the expansion of DSK horse culture, while modification of the proximal margin of the specimen (Figure 5) shows evidence of human attention to hoof care, and strongly implies the use of the horse in transport.

(a) Scale image of horse hoof clipping from Tsengel Khairkhan (TK105), dating to the 14th/13th centuries BCE. (b) 3D snapshots of modified dorsal surface, showing inset area of 50× magnification in one area of culturally-modified surface with a scoop-like morphology.
Implications for early horse transport and human-horse relations in East Asia
Horse remains from Tsengel Khairkhan have important implications for our understanding of the broader picture of horse transport in prehistory. The first horse transport dates back at least as far as ca. 1950 BCE, to burials of the Sintashta culture in the Russian Trans-Ural region (Lindner, 2020). However, outside of occasional appearances in iconography as scouts or messengers, indicators of the widespread use of horses in high-stress mounted riding activities, like warfare, only appear during the early first millennium BCE (Drews, 2004). In some areas of East Asia, including ancient Chinese states, the initial adoption of cavalry was significantly later, during the late first millennium BCE or beyond (Li et al., 2020). In Mongolia, some archaeological indicators such as petroglyphs and osteological features on horse teeth point to a preference for chariot transport during the late second millennium BCE (Taylor et al., 2021a). However, some scholars hypothesize that Inner Asian pastoralists likely engaged in regular mounted riding long before its first emergence in Chinese historical records (Mair, 2003), and compelling indirect evidence for early mounted riding, including single horse burials with bridle equipment (ca. 1108–893 cal. BCE), and the innovation of trousers with reinforced groyne areas (ca. 1261–1041 cal. BCE), point towards regular mounted riding by pastoralists towards the end of the second millennium BCE (Beck et al., 2014; Wagner et al., 2011).
In context, our finds at Tsengel Khairkhan strongly imply the use of high-mountain horse transport, in the form of packing or mounted riding, prior to the proliferation of horse burials and DSK horse culture in the Mongolian steppe. The steep scree and icy slopes of the study area would render wheeled vehicles or even sledge useless, and so any domestic horses reaching the high mountain slopes must have been either ridden or led up the slopes as pack animals on foot. Careful attention to hoof maintenance is potentially consistent with use of horses for riding, where problems in hoof conformation would have a more immediate impact on humans than when simply pulling horses on a lead. The need to move large animal carcasses downslope after a successful hunt would have provided strong incentive to use horse transport. At the same time, other distractions or stimuli that might increase stress are typically absent in mountain transit. Therefore, it is possible that hunting and packing at high altitudes (Figure 6) was an early application of mounted riding by Inner Asian pastoralists, before the approach was later honed for high-intensity situations such as combat and high-speed, long-distance riding.

Artist’s rendering of a high altitude hunter at Tsengel Khairkhan, using horses for pack transport after successfully harvesting an argali.
Support for the idea of such task-specific applications of mounted horseback riding by early Mongolian pastoralists before the transition to mounted cavalry may also be found in other archaeological datasets. For example, at the end of our 2022 field campaign, we documented a petroglyph in Gobi-Altai province, central Mongolia, that appears to show two chariots alongside a mounted rider (Figure 7). One of the chariots appears equipped with a shield displaying unique chevron images identical to those often found on deer stone carvings, which, assuming the images are part of a single composition, would seem to imply the presence of mounted horsemanship in DSK culture.
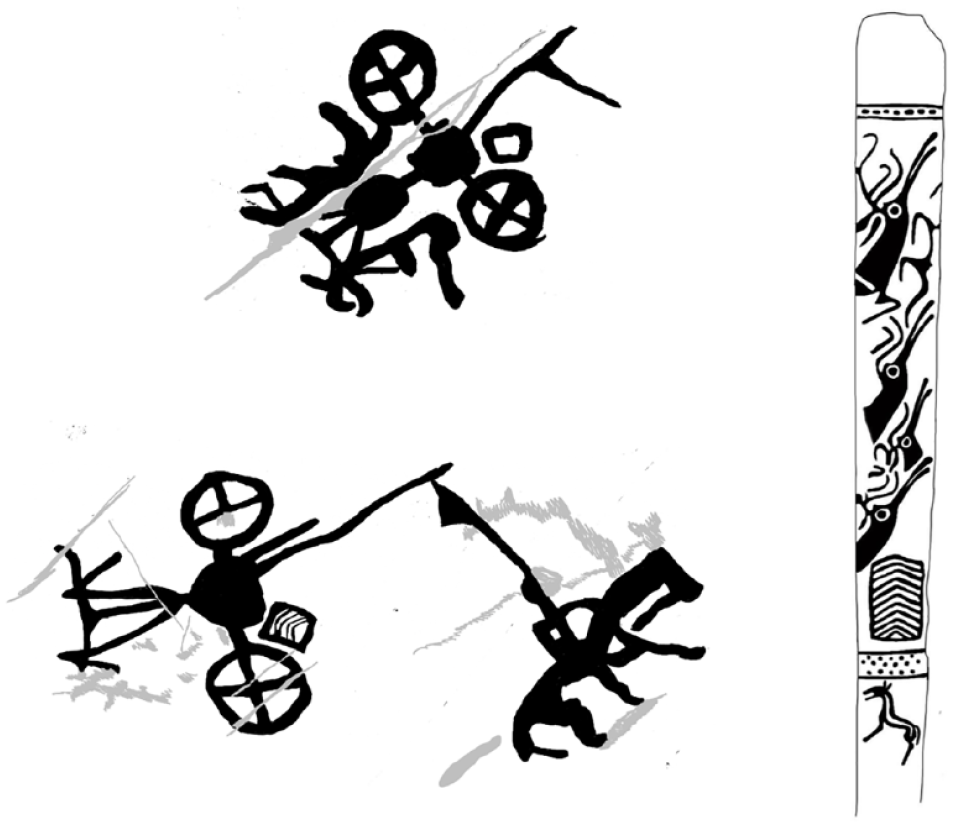
Left: A rock art panel in Gobi-Altai, Mongolia, shows two chariots (associated with the DSK culture through the chevron shield motif) along with a single rider. It is unclear whether the grey damaged area adjoining the rider originally depicted a bow-and-arrow, or simply represents natural taphonomic damage. Right: A deer stone image from Daagan Del, Zavkhan province, Mongolia, showing deer images along with a belt, tools, the ubiquitous DSK shield/chevron motif, horse decorations, and anthropomorphic features such as earrings.
Together, our finds show that mountain ice is a valuable chronicle of changes in the human-horse relationship across the earliest chapters of Inner Asia’s pastoral history, revealing the use of mountain ice by Przewalski’s horse, and later the earliest evidence of domestic horses and mounted horseback riding in the Mongolian Plateau. Future investigation of glaciers and ice patches thus holds the potential to provide a unique window into the early development of the human-horse story in East and Inner Asia.
Conclusion
Archaeological survey of snow and ice provides direct evidence for the use of domestic horses at high altitudes in western Mongolia from the second half of the second millennium BCE through the 20th century. Genomic sequencing of a horse specimen dating to ca. 1700 BCE demonstrates the first genomic evidence of Przewalski’s horse in the eastern steppe, and indicates that this taxon utilized high-altitude zones where it may have been exploited by human hunters alongside argali sheep and other wild game. Recovery of artefacts such as horseshoes, hoof picks, and even hoof trimmings suggest that careful hoof maintenance was required to sustain horse transport on high mountain slopes. Hoof trimmings from high-altitude ice margins represent the earliest indication of domestic horse use in ancient Mongolia yet identified, and suggest that horse transport at high altitude zones may have played an important early role in the evolution of early mounted riding.
Supplemental Material
sj-pdf-1-hol-10.1177_09596836241254484 – Supplemental material for High altitude horse use and early horse transport in eastern Eurasia: New evidence from melting ice
Supplemental material, sj-pdf-1-hol-10.1177_09596836241254484 for High altitude horse use and early horse transport in eastern Eurasia: New evidence from melting ice by William Timothy Treal Taylor, Isaac A Hart, Tumurbaatar Tuvshinjargal, Jamsranjav Bayarsaikhan, Nicholas L Jarman, Peter Bittner, Paula López Calle, Logan A Blakeslee, Muhammad Zahir, Lorelei Chauvey, Gaëtan Tressières, Laure Tonasso-Calvière, Stéphanie Schiavinato, Corinne Cruaud, Jean-Marc Aury, Pedro H Oliveira, Patrick Wincker and Ludovic Orlando in The Holocene
Footnotes
Correction (August 2024):
Article updated to include the missing accession number “PRJEB76474” in the “Data availability” statement.
Author contributions
WT and IH wrote the paper, TT, JB, NJ, PB, PLC, LAB, and MZ provided archaeological data, LC, GT, LTC, and SS carried out the ancient DNA wet-lab work, under LO’s supervision. CC, JMA PO, and PW performed DNA sequencing. PW and LO provided reagents and materials. LO carried out ancient DNA data analyses. LO wrote the Ancient DNA sections, and plotted the corresponding figures.
Data availability
The sequence data generated in this study are available for download at the European Nucleotide Archive (accession no. PRJEB76474). All other data used in the analysis are provided in the manuscript and supplementary materials.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Project fieldwork was funded through awards from the University of Colorado-Boulder Research and Innovation Office and the Council of American Overseas Research Centers (CAORC). The project has received funding from the CNRS, University Paul Sabatier (AnimalFarm IRP), and the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreements 681605-PEGASUS and 101071707-Horse Power). This work is part of the MARENGO project and was supported by the Genoscope, the Commissariat à l’Énergie Atomique et aux Énergies Alternatives (CEA) and France Génomique (ANR-10-INBS-09–08).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.