
Abstract
Over the past decades, numerous studies have examined human ecodynamics, focusing on the trajectories of socio-ecological systems (SES) and the co-evolution of natural and social subsystems. Resilience Theory (RT) provides a conceptual framework for analyzing these dynamics, particularly through the metaphor of adaptive cycles. In this study, we investigate the consumption patterns of agropastoral societies that inhabited northern La Rioja from the 3rd to 17th centuries CE, exploring long-term human adaptation in the drylands of northwestern Argentina. Zooarchaeological analysis revealed shifts in consumption patterns over time. From the 3rd to 7th centuries CE, subsistence strategies were primarily centered on the exploitation of artiodactyls. However, by the 11th century CE, a process of residential site abandonment and dietary diversification emerged. The results indicate significant changes in faunal exploitation, particularly a decline in Artiodactyla consumption, driven mainly by demographic growth and socio-economic adjustments. These transformations led to greater mobility and a shift toward opportunistic resource exploitation. Our findings highlight the resilience and adaptability of human populations in response to socio-economic and demographic challenges. Local societies demonstrated flexibility in adapting to environmental unpredictability and demographic shifts by adjusting their strategies to new conditions.
Introduction
The study of human ecodynamics (HE) focuses on the dynamics of socio-ecological systems (SES) and their irreducible interactions (Fitzhugh et al., 2019; McGlade, 1995). This approach emphasizes long-term processes of stability, resilience, and change in SES relationships. From this perspective, space is socially constructed and time-dependent, causality is non-linear, destabilizing processes function as positive feedback, and the explanations are built upon interpretative and integrative analyses. Predictions are not feasible due to the inherent chaos of non-linear systems (McGlade, 1995).
Resilience Theory (RT) provides a valuable conceptual framework for evaluating how communities respond to external shocks, such as environmental changes. Resilience is commonly defined as the capacity of a system to adapt to changes while maintaining its core functions (Holling, 1973). Some scholars prefer the term “stability landscape” to describe a system in dynamic equilibrium that, when faced with disturbances, develops counteracting forces to restore equilibrium (Scheffer, 2009; Walker et al., 2004). However, destabilizing forces—whether internal or external—can push the system into an alternative basin, leading to a reorganization of its components (Davies et al., 2021; Newton et al., 2024).
An important contribution to resilience analysis is the concept of adaptive cycles, developed by Gunderson and Holling (2002). This model proposes that SES transitions through four main phases. Exploitation (r) corresponds to the phase of rapid colonization of recently disturbed areas; conservation (K) represents the phase of slow accumulation and storage of energy and materials; release (Ω) occurs when the accumulated biomass becomes increasingly fragile and is suddenly released by external agents; and reorganization (α) phase involves the reconfiguration of the system, which may either resemble its predecessor or develop new functional characteristics (Gunderson and Holling, 2002; Redman, 2005). While this model is a useful framework for guiding research, it is important to build models based on appropriate data and to interpret findings in the context of archeological records (Fitzhugh et al., 2019; Gunderson and Holling, 2002; Heitz et al., 2021; Redman, 2005).
In a previous work (Cahiza et al., 2021), we examined the temporal dynamics of occupation in northern La Rioja (Argentina), identifying phases of growth and decline in occupation intensity, as well as an east-west mobility process, particularly after the late 10th century. In this paper, we analyze the consumption patterns of these societies between the 3rd and 17th centuries and the changes observed throughout the temporal sequence. Our hypothesis is that demographic and environmental changes led to adjustments in subsistence patterns related to faunal consumption. The objective is to identify the adaptive and resilience mechanisms related to subsistence practices of small-scale societies in the drylands of northern La Rioja. Zooarchaeology provides a critical framework for understanding the interaction between humans and animals in the past. Here, we present the results of various zooarchaeological analyses that shed light on the adaptive processes of human populations in northern La Rioja.
Eastern sector of the Sierra de Velasco (La Rioja, Argentina)
The research area is located in the northeastern foothills of the Sierra de Velasco, in the northern region of La Rioja province (Figure 1), northwestern Argentina. La Rioja lies within the arid diagonal of South America. The area is characterized as a continental desert, with an average annual precipitation of approximately 200 mm and significant altitudinal gradient variation. The mean temperature is 16.6°C, with extremes ranging from 40°C to −9°C.
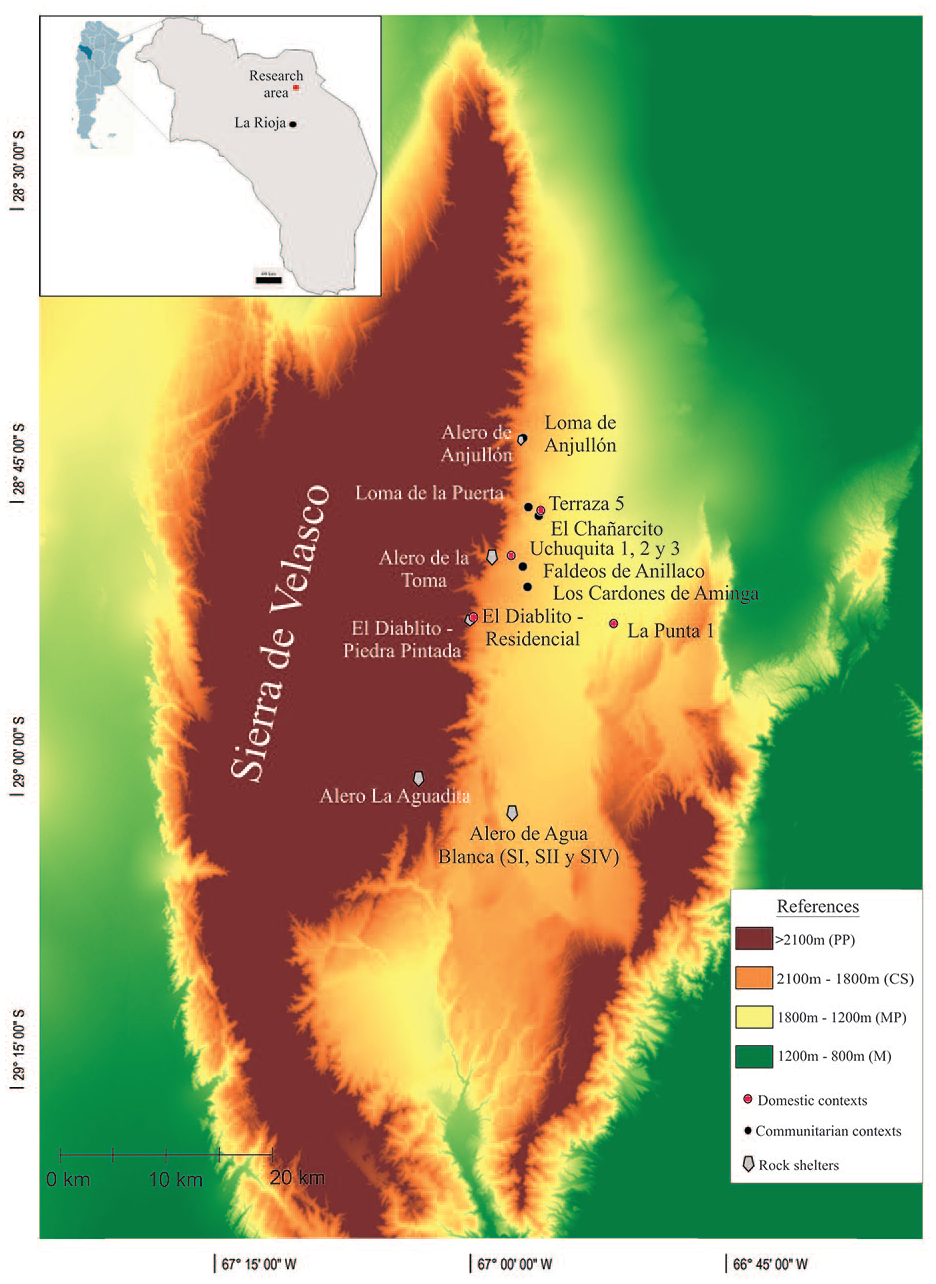
Localization of the research area and archeological sites based on relief characteristics (La Rioja, Argentina).
Relief is the main factor determining climatic conditions in the area. The altitude ranges from 800 to 4000 masl. Between 800 and 1200 masl, the valley floor is characterized by the environment of the Monte (M) phytogeographic province. The vegetation forms a mosaic of two types: a zonal type, represented by shrub steppes or open Larrea cuneifolia shrublands, and an azonal or edaphic type, consisting of open, and aligned Neltuma chilensis (algarrobo) forests, which thrive along the courses of intermittent rivers.
Between 1200 and 1800 masl, the piedmont (MP) develops, encompassing alluvial fans and gently sloping foothills. The vegetation is typical of the Monte, where Larrea scrublands are gradually replaced by open shrublands dominated by Flourensia fiebrigii, alongside Trichocereus terschekii. The aligned forests feature trees such as Celtis ehrenbergiana and Neltuma chilensis.
Above 1800 masl, the Chaco Serrano (CS) exhibits vegetation characteristic of the Chaco phytogeographic province. The steep mountain slopes foster a diversity of environments, including shrublands, forests, and grasslands. The shrubland is dense and hosts various shrub species from the Asteraceae and Verbenaceae families. Rocky slopes are characterized by cushion-forming bromeliads (Deuterocohnia brevifolia), cacti, and ferns. Aligned forests thrive in ravines with permanent rivers, where Parasenegalia visco and Lithraea molleoides are the dominant species.
Finally, on plains above 2100 masl, vegetation corresponds to the Prepuna (PP) and High Andean phytogeographic provinces (Biurrun et al., 2012), with high-altitude grasslands dominated by Jarava ichu.
The faunal assemblage comprises species characteristic of the Monte, Arid Chaco, and Prepuna phytogeographic provinces. South American camelids were the most relevant resource in the subsistence of human communities in northwestern Argentina (NWA) throughout the Holocene (Belotti López de Medina, 2024; López, 2009; Olivera and Grant, 2008; Yacobaccio, 2003). Two wild species (Vicugna [vicuña] and Lama guanicoe[guanaco]) and two domestic species (Vicugna pacos [alpaca] and Lama glama [llama]) inhabited the region. However, alpaca husbandry in NWA may have been ecologically challenging, as noted by several researchers (Franklin, 1983; Mengoni Goñalons and Yacobaccio, 2006; Wheeler, 1995). Currently, just Lama guanicoe was registered in Sierra de Velasco, and there are some indirect archeological evidences of the presence of Lama glama in the past (Garate, 2021; Garate et al., 2024, Garate et al, 2025). There is limited evidence of taruca (Hippocamelus antisensis) in the archeological record, although its presence has been documented to this day in the upper sectors of the Sierra de Velasco (Raviña and Callegari, 1992). In zooarchaeological assemblages, other taxa, like rodents (Lagidium viscacia, Galea sp., Ctenomys sp.), birds (Eudromia elegans and Rhea sp.), and armadillos (Chaetophractus villosus) are frequently present (Garate, 2021; Garate et al., 2024, Garate et al, 2025).
Paleoenvironment, demographic trends, and subsistence strategies in northwestern Argentina
Throughout the Late-Holocene, the environment experienced a series of oscillations between arid and humid phases. On global scale, researchers have recognized two complex phenomena (Bradley, 2015; Bradley et al., 2003; Compagnucci, 2000; Diaz et al., 2011; Ge et al., 2010; Hughes and Diaz, 1994; Kock et al., 2020; Lüning et al., 2019; Morales et al., 2015, 2020; Stine, 1994, 1998). The Medieval Climate Anomaly (MCA) was a climatic perturbation that occurred primarily between 1000 and 1300 CE. Studies suggest that during this period, the climate was hotter and drier than today (Flores-Aqueveque et al., 2024; Kock et al., 2020; Lüning et al., 2019). Some scholars (Cook et al., 2022) have identified episodes of megadroughts in various regions of South America, including Patagonia, central Chile, and central-western Argentina. They recognized some of these events occurring around 1050–1200 CE (coinciding with the estimated period of the MCA), 1250–1400 CE, 1615–1637 CE, 1684–1696 CE, and from 2008 to the present (Cook et al., 2022). The other climatic perturbation is the Little Ice Age (LIA), which spanned from the 15th to the 19th century, imposing colder and wetter conditions, although interspersed with periods of increased aridity (Martel-Cea et al., 2016; Mayta and Maldonado, 2022; Morales et al., 2015; Ortiz et al., 2012; Sampietro Vattuone et al., 2018).
In a previous study (Cahiza et al., 2021), we presented a model of human population trends between the 3rd and 17th centuries. Based on the analysis of radiocarbon dates, we identified three phases. The first temporal component, dated between the 3rd and 7th centuries, corresponds to the processes of colonization, and settlement (Sabatini, 2019; Sabatini and Cahiza, 2021). During this period, people primarily constructed their houses and agricultural infrastructure along the foothills of the Sierra de Velasco. The second temporal component was identified around the 7th century, marked by significant demographic growth in the region. This phase is evidenced by larger and more complex residential sites (Cahiza et al., 2018a, 2018b, 2021; Garate, 2021; Garate et al., 2024). The third temporal component spans from the 11th to the 17th century. A notable shift in the growth trend occurred around 1000 CE, when the study area experienced the abandonment of residential sites. These demographic changes occurred within the context of the megadrought and the events of the MCA (Cahiza et al., 2021; Cook et al., 2022; Kock et al., 2020). Human presence during this period is documented exclusively in rock shelters, suggesting a shift toward increased mobility. From 1500 CE onward, the emergence of sites with defensive features (pukaras) indicates a heightened level of conflict in the region.
In this context, we believe that demographic and environmental changes likely caused shifts in the subsistence patterns of local groups (Bettinger, 2009; Carlson and Edenhamn, 2000; Harrison and Taylor, 1997; Muscio, 2004; Redman, 1999; Winterhalder and Goland, 1993, 1997). In northwestern Argentina (NWA), the economic base of most societies was linked to the development of agricultural and pastoral systems (Acuto, 2007; Berberián and Nielsen, 1988; González, 1955; Izeta, 2007; Laguens, 2006; Núñez Regueiro, 1974; Olivera, 2001; Tarragó, 2000; among others). However, hunting wild species—such as camelids, cervids, rodents, armadillos, and birds—remained a significant activity (Belotti López de Medina, 2010; Dantas, 2011; Figueroa, 2010; Izeta, 2007; Laguens et al., 2013; Mercolli, 2016; Miyano, 2018; Olivera and Grant, 2008; Yacobaccio et al., 1997-1998.
Methodology
Here, we analyzed the archaeofaunal assemblages from 18 sites that are representative of the different temporal, demographic, and ecological patches (Table 1). For this study, residential units were grouped into two categories: domestic scale (simple enclosures with one or two rooms) and community scale (with the presence of public spaces and larger built areas that foster greater social interaction; Cahiza et al., 2018a; Drennan and Peterson, 2005). In addition to the residential sites, rock shelters were included as a third category. A detailed description of each archeological context is provided in Garate (2025).
Summary of Archaeological Sites by Temporal Component and Environmental Context.
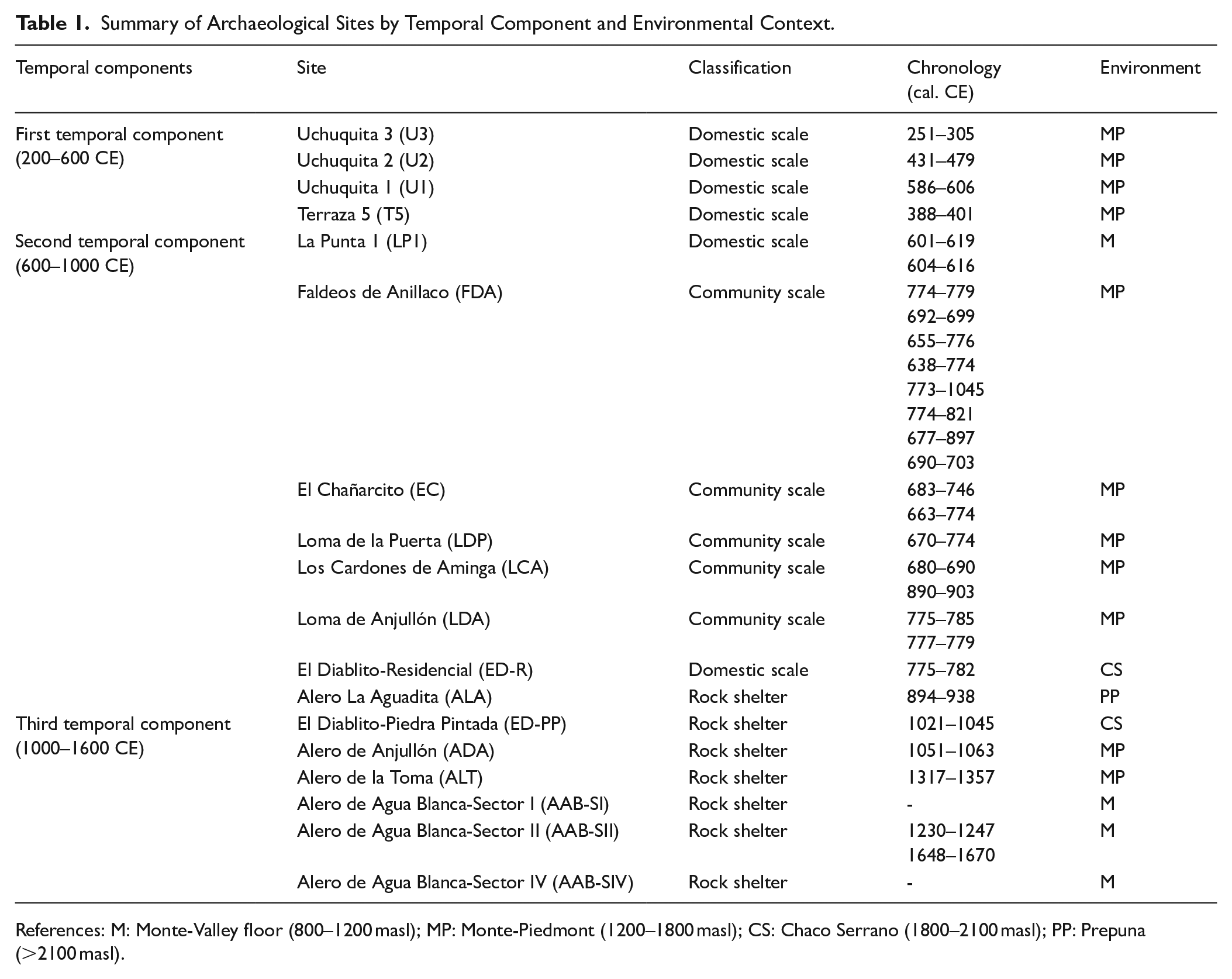
References: M: Monte-Valley floor (800–1200 masl); MP: Monte-Piedmont (1200–1800 masl); CS: Chaco Serrano (1800–2100 masl); PP: Prepuna (>2100 masl).
The zooarchaeological analyses employed a traditional methodology (Lyman, 1994, 2008; Mengoni, 2010), which involved the identification of anatomical and taxonomic characteristics of bone material. The identification of the recovered remains was carried out using comparative reference collections available at Instituto Nacional de Ciencias Humanas, Sociales y Ambientales (INCIHUSA, CONICET-Mendoza) and osteological atlases (Altamirano, 1983; France, 2009; Pacheco Torres et al., 1979; Sierpe González, 2015; among others). For quantification, we employed the Number of Specimens (NSP) to account for the total number of remains; the Number of Identified Specimens (NISP) for those bones that could be assigned to any taxonomic category (Lyman, 1994, 2008). To evaluate taxonomic richness, we used Ntaxa, the Simpson’s diversity index (1/D), the evenness index (V’), and the Artiodactyla index (IA; Broughton, 1994; Grayson, 1991; Lyman, 2008; Mengoni, 2010). For the calculation of diversity indices, we used the most specific taxonomic categories available for each (Grayson, 1984; Lyman, 2008; Reitz and Wing, 2008). Correlations between these indices and altitude (masl) were assessed using Pearson’s correlation coefficient.
We assumed that human populations made rational and adaptive decisions to optimize available resources and maximize their energy returns (Winterhalder, 1981). The exploitation of animals and ecological patches likely followed rational strategies linked to the optimal use of the environment (Boone, 1992; Kaplan and Hill, 1992; Kelly, 1995; Winterhalder and Smith, 1981; among others). In practice, this implies that people incorporated various available resources into their diets based on the energy return rate of those resources (Smith et al., 1983; Winterhalder and Goland, 1997). It has been suggested that there is a positive correlation between animal size and their return rates (Broughton et al., 2011). In northern of Sierra de Velasco, the primary animals in this category are camelids (Lama guanicoe and Lama glama). Consequently, a predominance of Artiodactyla is expected throughout the analyzed temporal sequence. Here, we estimated the age at death of camelid remains to determine mortality patterns (Kaufmann, 2009)
Results
The details of archaeofaunal analysis are presented in Supplemental Material due to the size of the samples considered. We identified that Rodentia (42.6%, NISP = 2393), Artiodactyla (36%, NISP = 2026), Camelidae (13.2%, NISP = 744), Birds (4.2%, NISP = 234) and Dasypodidae (3%, NISP = 170) were the most frequent taxonomic categories in the total assemblage (Figure 2). Rodentia includes only small mammals (micromammals) that could not be identified to genus or species level. Artiodactyla and Camelidae were considered separately due to the presence of Hippocamelus antisensis in the area, but we have limited evidence of their consumption in archeological sites. Therefore, we believe that most of the Artiodactyla remains likely correspond to the Camelidae family.
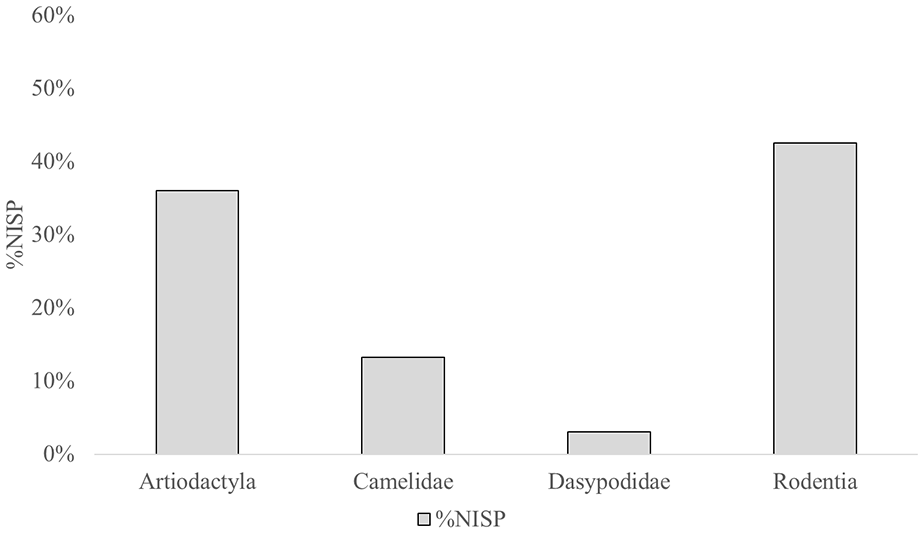
Percentage of identified specimens (NISP%) in the total assemblage.
In the first temporal component, butchery marks (n = 39) were identified exclusively on Artiodactyla (64.1%, n = 25) and Camelidae (35.9%, n = 14) remains. The second temporal component (n = 225) showed a similar pattern, with most butchery marks recorded on Artiodactyla (54.2%, n = 122) and Camelidae (37.3%, n = 84), followed by Birds (3.6%, n = 8) and Rodentia (2.7%, n = 6). In the third temporal component, butchery marks (n = 17) were observed on Artiodactyla (23.5%, n = 4), Camelidae (23.5%, n = 4), Birds (17.6%, n = 3), Dasypodidae (11.8%, n = 2), Rodentia (5.9%, n = 1), Galea musteloides (5.9%, n = 1), and Lagidium sp. (5.9%, n = 1; Figure 3).
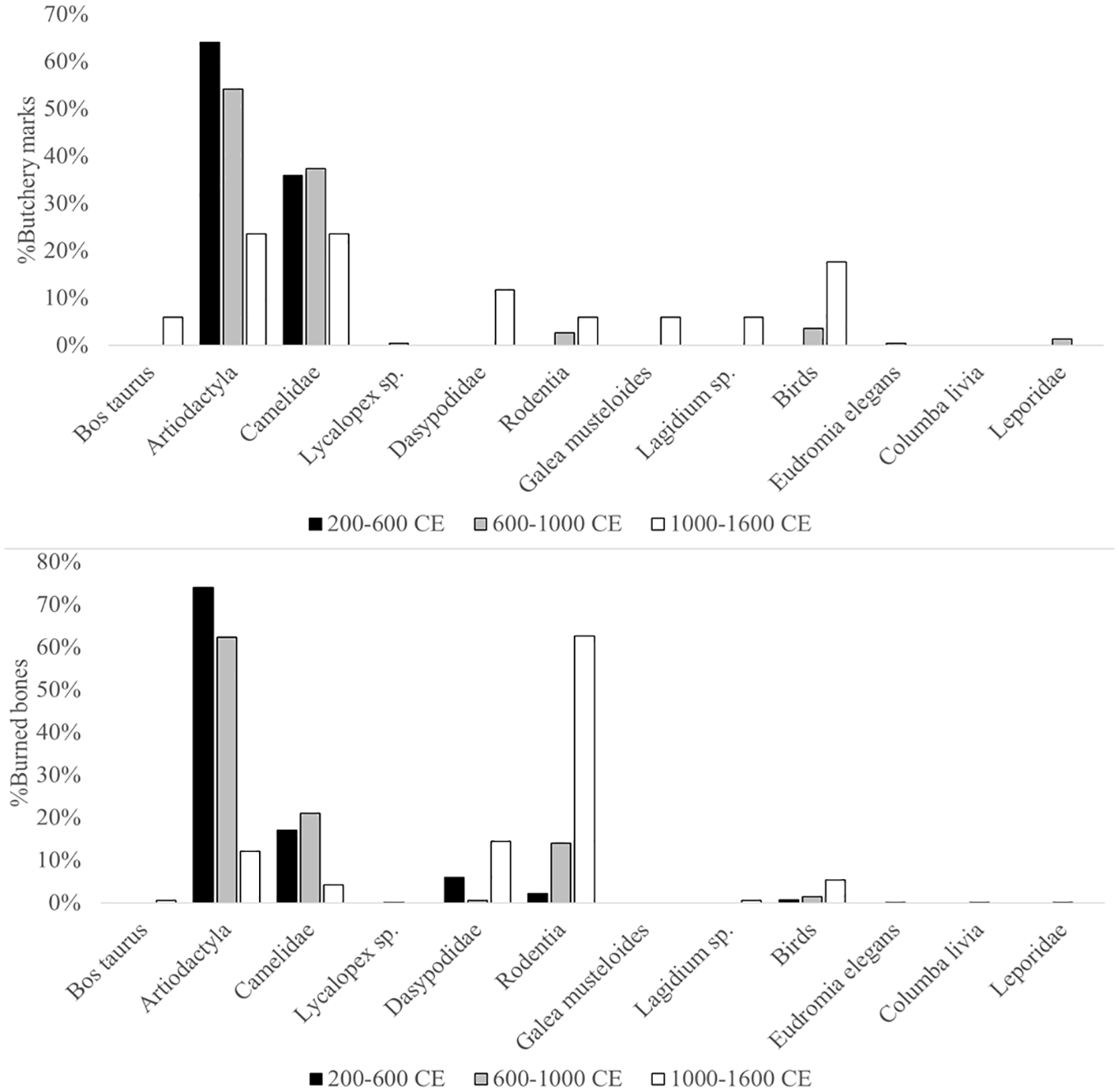
Top: Percentage of butchery marks recorded for each temporal component; Bottom: Percentage of burned bones recorded for each temporal component.
The analysis of burned bones revealed similar trends (Figure 3). In the first temporal component, burned specimens (n = 135) were recorded primarily on Artiodactyla (74.1%, n = 100), followed by Camelidae (17%, n = 23), Dasypodidae (5.9%, n = 8), Rodentia (2.2%, n = 3), and Birds (0.7%, n = 1). During the second temporal component, burned bones (n = 1228) were most frequently associated with Artiodactyla (62.4%, n = 766) and Camelidae (21.1%, n = 259), which together accounted for 83.5% of the total. The remaining specimens corresponded to Rodentia (14.1%, n = 173), Birds (1.5%, n = 18), Dasypodidae (0.7%, n = 8), Lycalopex sp. (0.1%, n = 1), Eudromia elegans (0.1%, n = 1), Columba livia (0.1%, n = 1), and Leporide (0.1%, n = 1). The third temporal component (n = 166) was characterized by a marked predominance of burned Rodentia remains (62.7%, n = 104), followed by Dasypodidae (14.5%, n = 24), Artiodactyla (12%, n = 20), Birds (5.4%, n = 9), Camelidae (4.2%, n = 7), Lagidium sp. (0.6%, n = 1), and Bos taurus (0.6%, n = 1).
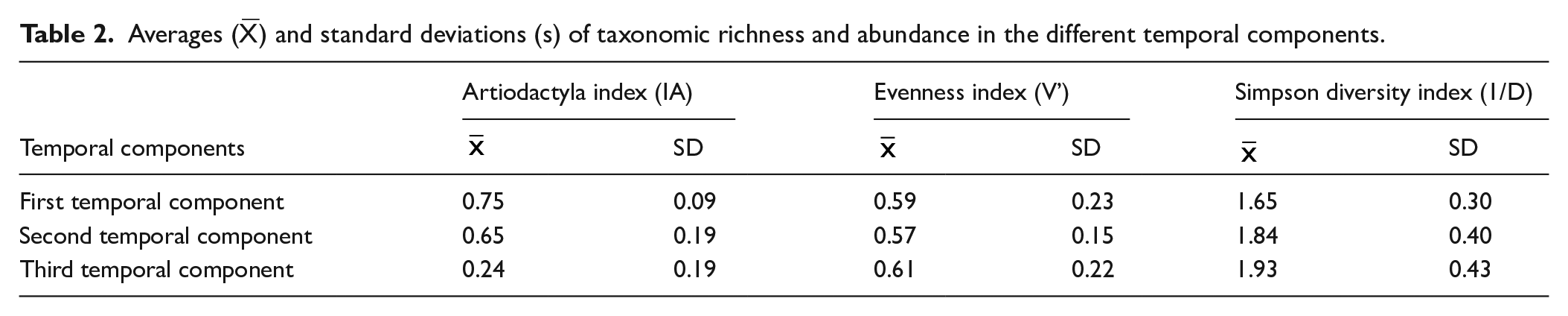
Table 2 summarizes the results of richness and taxonomic indexes for each defined temporal component. The Artiodactyla index shows a predominance of these animals in the assemblages of first and second temporal component but it declines over time. The evenness index presents similar values across the three temporal components. The Simpson index shows an increasing trend over time, indicating greater diversity.
Averages (
The Artiodactyla index shows higher mean values at domestic and community-scale sites (Figure 4). In rock shelter sites this index presents lower values. These results imply that Artiodactyla dominated the archaeofaunal assemblages of residential sites but not in rock shelters.
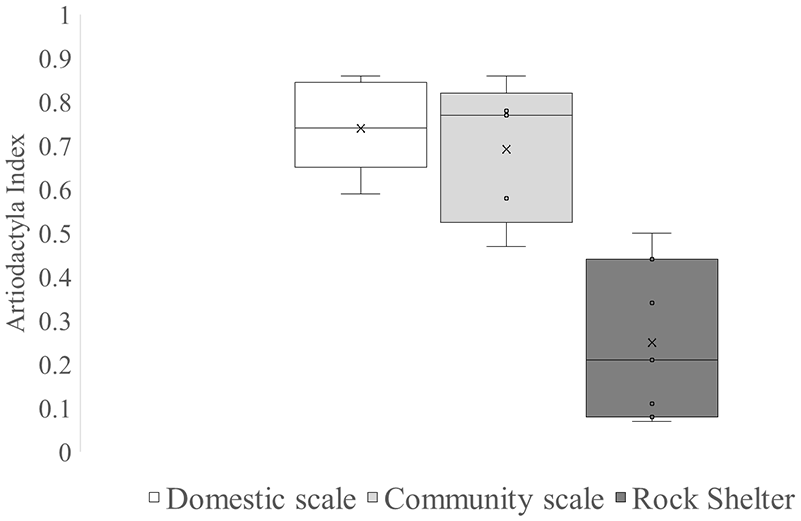
Box plot of Artiodactyla index in domestic scale sites, community scale sites and rock shelters. Boxes show the interquartile range (25th–75th percentiles), with medians as horizontal lines and means as “×.” Whiskers indicate values within 1.5 × IQR. Dots represent individual assemblages.
On the other hand, we detected different trends in the correlation between the Artiodactyla index and altitudinal gradient (masl; Figure 5). Assemblages from the first temporal component showed a negative and non-significant correlation with altitude (r = −0.52, p = 0.48). For the second temporal component we found a negative and significant correlation (r = −0.75, p = 0.03). Finally, for third temporal component was obtained a positive significant correlation (r = 0.84, p = 0.03).
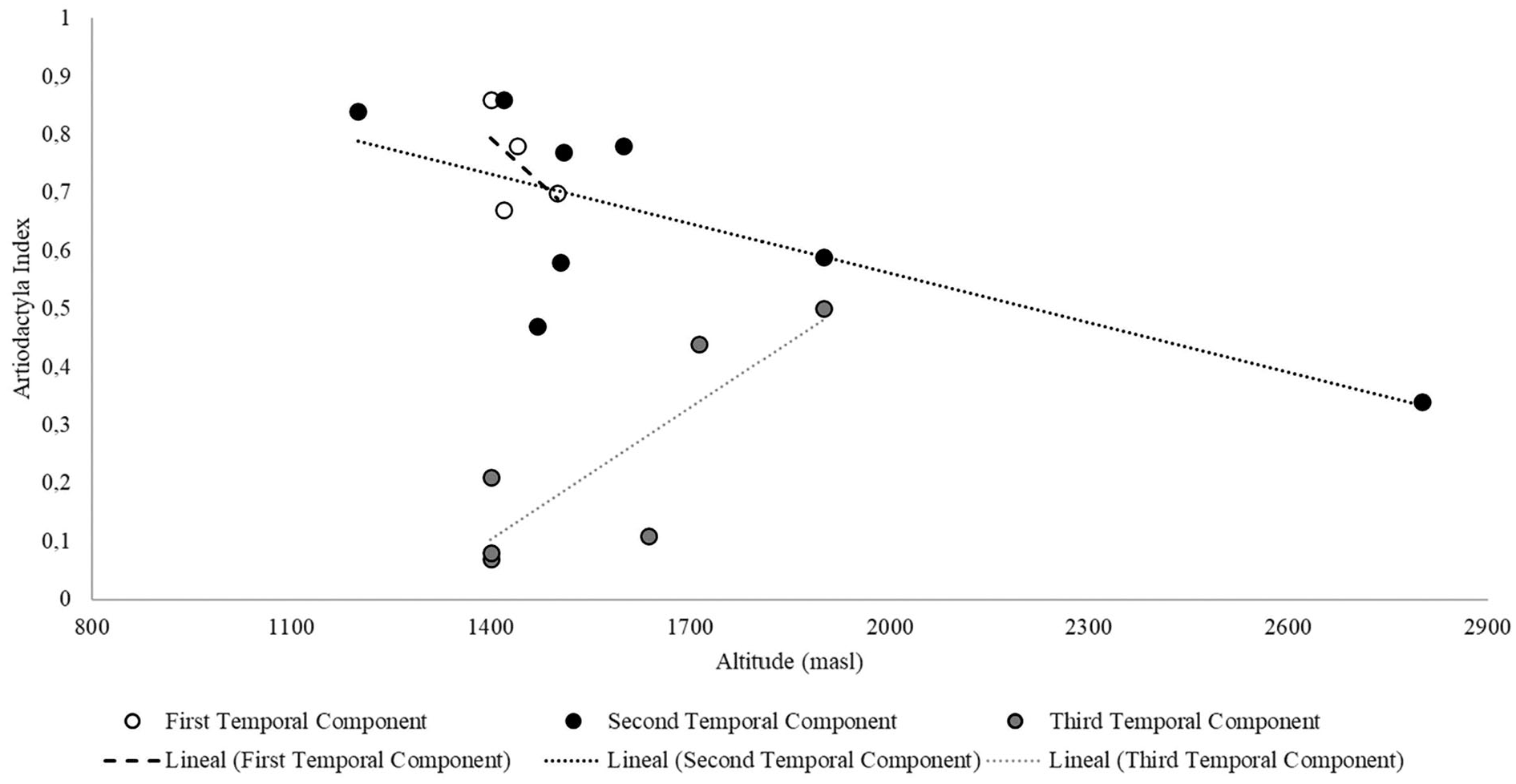
Artiodactyla index over the altitudinal gradient (masl).
The mortality profiles of camelid remains indicated similar patterns for the first and second temporal components. A total of 140 elements with diagnostic zones were recovered, enabling the determination of the age at death of the camelids. Although a smaller number of remains were available for the first temporal component, a strong and significant positive correlation (r = 0.96; p = 0.008) was observed in the different age categories represented for the first and second temporal components. In both cases, adult was the most frequent category (Figure 6). For the third component, few elements with features allowing for the determination of the age at death of the individuals were recovered. However, the results indicate a tendency toward the exploitation of subadults.
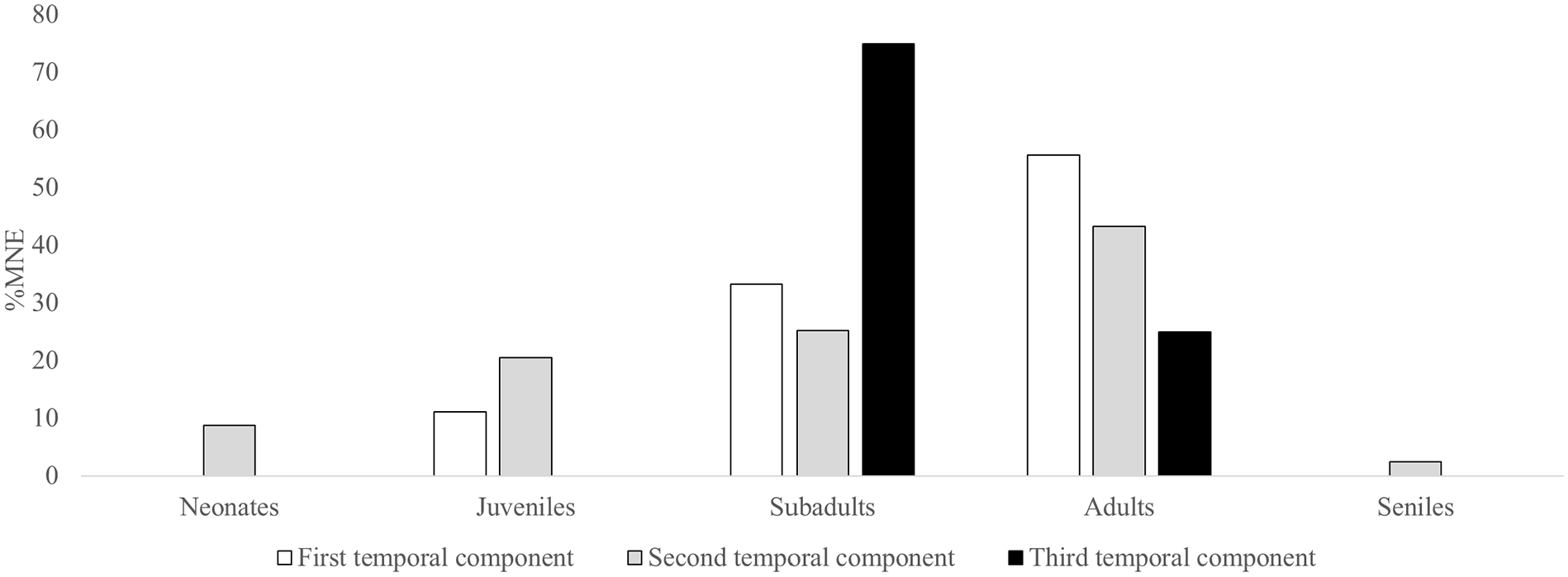
Mortality profiles of camelids across the temporal components.
Residential sites and rock shelters showed differences in consumption patterns. We determined that rock shelters presented highest values of the diversity index (Figure 7), while community-scale sites showed greater diversity than domestic-scale sites.
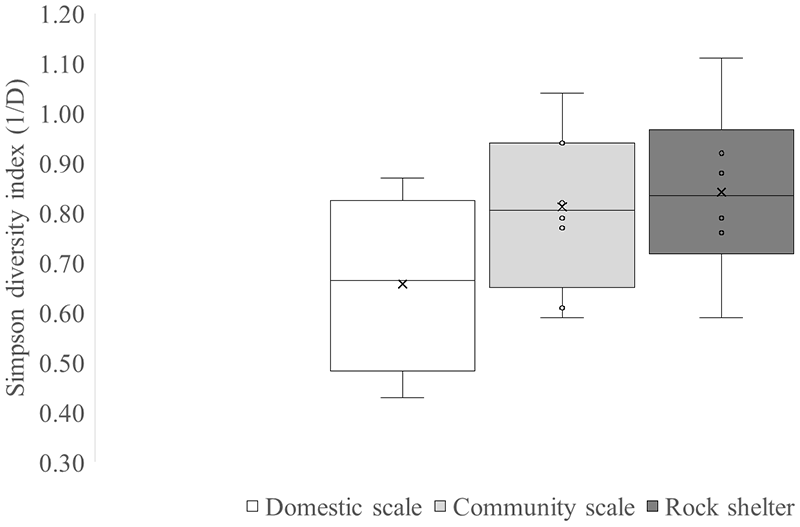
Box plot of Simpson diversity index (1/D) in domestic scale sites, community scale sites and rock shelters. Each boxplot shows the distribution of values per context. The boxes represent the interquartile range (25th–75th percentiles), the horizontal line within each box indicates the median, and the “×” represents the mean. Whiskers extend to the minimum and maximum values within 1.5 times the interquartile range. Individual data points are shown as dots.
Additionally, we evaluated the relationship between the altitudinal gradient (masl) and the Simpson diversity index (Figure 8). A positive and significant correlation was recorded for the first temporal component (r = 0.95, p = 0.05), while a non-significant positive correlation was obtained for the second temporal component (r = 0.17, p = 0.69) and the third temporal component (r = 0.59, p = 0.22).
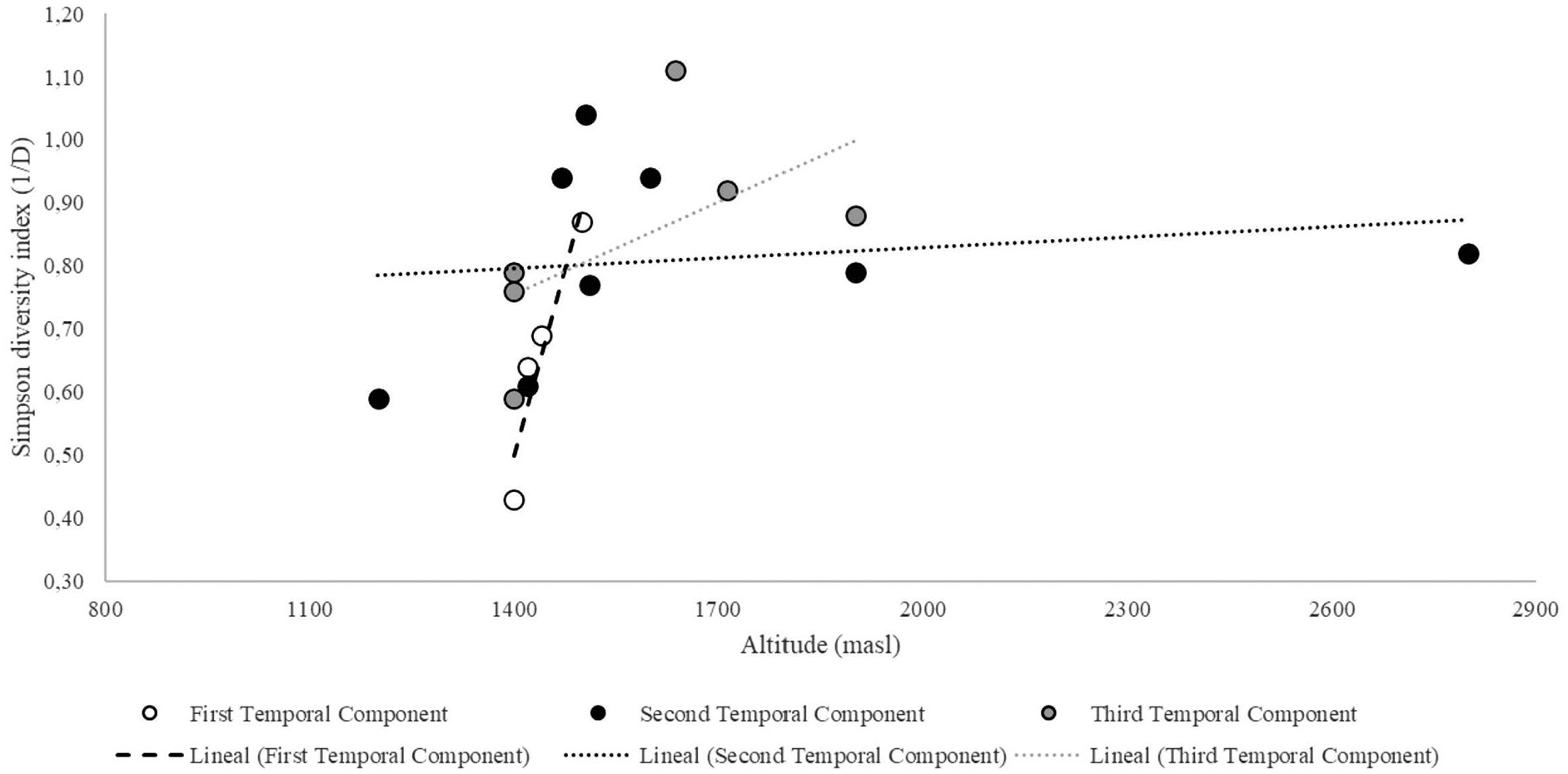
Simpson diversity index (1/D) over the altitudinal gradient (masl).
Discussion
The presented results indicated changes in faunal exploitation over time. A decreasing trend in the consumption of Artiodactyla was recognized (Figure 9). Two processes converged to explain this change: population growth from the 7th century CE onward and climate fluctuations, especially from the 11th century CE onward (Cahiza et al., 2021; Kock et al., 2020). Between the 3rd and 11th centuries, Artiodactyla dominated archaeofaunal assemblages, but since the 7th century, their consumption declined, and people began to incorporate more lower-ranking resources into their diets. In recent works (Garate et al., 2025), based on osteometric and isotopic analysis, we proposed that subsistence strategies at residential sites from the first and second temporal components were characterized by a combination of llama pastoralism and guanaco hunting.
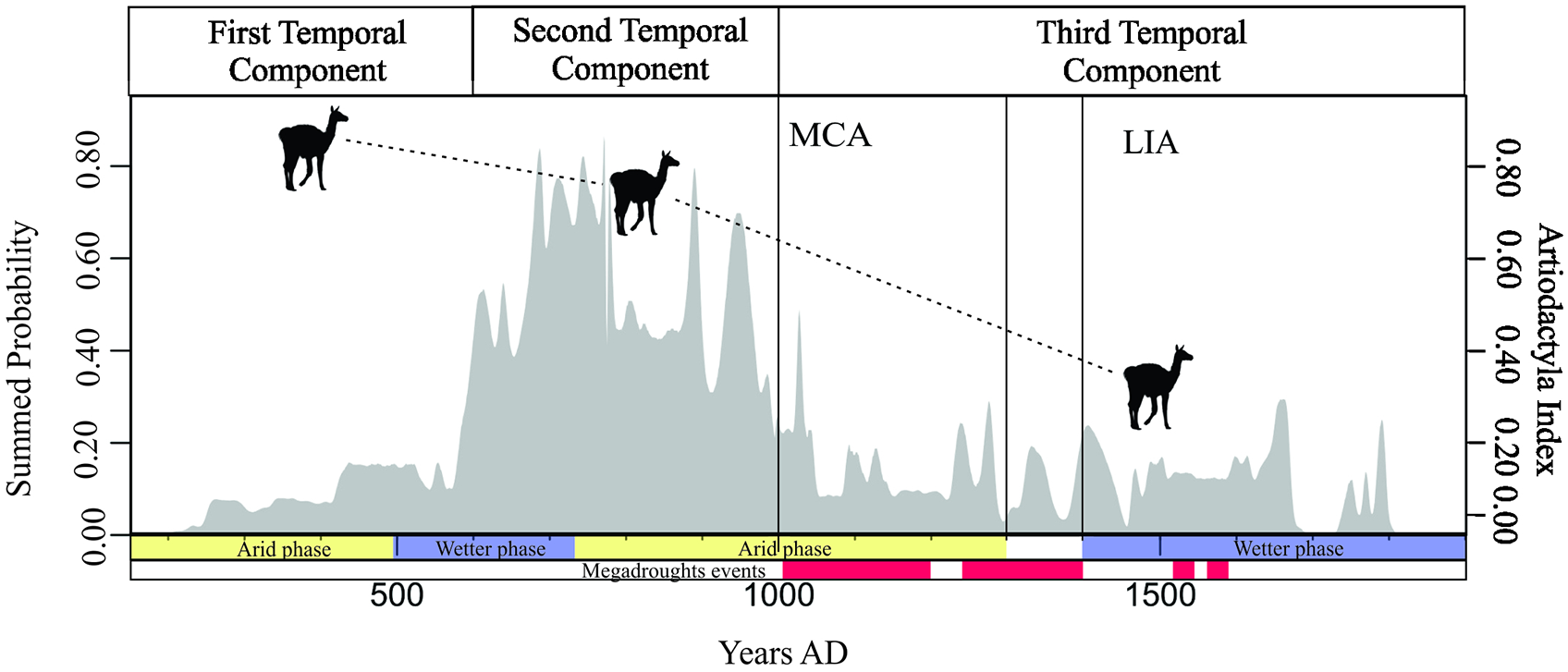
Demographic trends based on summed probability and variations in the Artiodactyla index across centuries. Colors indicate phases of arid (yellow) and wetter (purple) phases according to Kock et al. (2020). Red lines mark megadrought phases in South America (Cook et al., 2022).
From the 11th century CE onward, coinciding with the worsening climatic conditions (Cook et al., 2022; Kock et al., 2020), a process of area abandonment took place (Cahiza et al., 2021), and the Artiodactyla index experienced a sharp decline. The only evidence of human presence for this temporal component was registered in rock shelters, indicating higher mobility than in previous centuries. The obtained results showed an expansion of the diet, with higher proportion of low-ranking taxa (rodents, dasypodids, and birds). Previous taphonomic analysis of these assemblages (Garate, 2025) identified alternating use of rock shelters by humans and natural agents, with some faunal remains likely deposited by noon-anthropic processes. However, the presence of anthropogenic modifications and a significant proportion of burned bones (Figure 3) support the inference that humans intentionally exploited a broader range of taxa (Garate, 2025). A positive and significant correlation between the altitudinal gradient and Artiodactyla index was registered. We deduce that this could be understand as a result of opportunistic hunting. Camelid exploitation increased in upper zones, aligning with natural distribution of these resources. Furthermore, we determined the exploitation of Rheidae, Geoffroea decorticans, and Neltuma spp. in the valley floor. These resources are only available in lower areas of the region, and there is no evidence of their consumption in upper sectors. We argue that between the 11th and 16th centuries, subsistence strategies were oriented toward the opportunistic exploitation of resources based on their natural distribution. This occurred in a context of high mobility in the area.
Finally, from the 15th century onward, some sites with defensive characteristics (pukaras) began to emerge. For example, Loma Pircada is a site located in Chuquis locality, at 1600 masl. Excavations at this site revealed the presence of burnt Zea mays and camelid bones (Carrizo et al., 2001). Unfortunately, we do not have enough excavation work at the site, but we believe that the earliest evidence would indicate a process of reoccupation and the reactivation of traditional agro-pastoral systems in the area.
Based on available evidences, we identified four phases of human strategies linked to demographic and environmental changes. First, the exploitation phase (r) corresponds to the colonization of the area by village populations. This phase coincided with first temporal component (200–600 CE; Cahiza et al., 2021). During this phase, humans developed an agropastoral strategy combined with the hunting of guanacos and, to a lesser extent, small fauna. The management of llama herds involved the complementary use of different altitudinal environments and resources (Garate et al., 2025). Artiodactyla index highlights the central role of these animals in human consumption. This strategy proved effective and facilitated the adaptive success of human populations, which was reflected in a process of demographic growth.
Secondly, the conservation phase (K) corresponds to the period of demographic expansion. This demographic growth has been interpreted as the result of improved local climatic conditions (Cahiza et al., 2021; Kock et al., 2020) and the success of economic strategies (Garate et al., 2025). We found high values of Artiodactyla index in assemblages from the second temporal component (600–1000 CE). These values indicate that the consumption of artiodactyls remained significant. However, a decline in this index was recorded, accompanied by an increase in taxonomic diversity. It is possible that the increasing demographic pressure on high-ranking resources may have led to a progressive decline in the availability of guanacos. Although artiodactyls continued to play a central role in local economies, the increase in hunting of lower-ranking taxa suggests a rise in the procurement and processing costs of higher-ranking resources (Bettinger, 2009; Muscio, 2004; Winterhalder and Goland, 1993, 1997). The exploitation of llamas, on the other hand, would have been oriented toward their preservation until advanced ages, ensuring efficient use of their secondary products. The presence of neonates and seniles in the second temporal component indicates that the camelids were being bred near the sites at that time, reflecting a managed and sustained herd. Similar processes have been observed in various parts of the Argentine Andean region. Examples of this include dietary expansion proposals around 1000 CE in Calchaquí Valleys (Belotti, 2015; Izeta, 2007), Tinogasta (Miyano, 2018), and specialization in llama consumption in Antofagasta de la Sierra (Grant and Escola, 2015; Olivera and Grant, 2008). In the case of northern La Rioja, demographic growth likely drove subsistence adjustments through specialization in llama management combined with a process of dietary diversification. Furthermore, a marked change is observed in the third temporal component, suggesting a shift in camelid exploitation strategies.
The release phase (Ω) was identified from 1000 CE onward. This coincided with the onset of the Medieval Climate Anomaly and the increase in aridity (Kock et al., 2020). Gunderson and Holling (2002) assert that this phase occurs when the accumulated biomass becomes increasingly fragile and is suddenly released by external agents. In the study area, the release phase implied changes in socioecological conditions. Human ecodynamics involved the search for new ecological patches to west, an increase in mobility, the use of rock shelters, and a greater exploitation of wild resources (such as rodents, armadillos, birds, eggs, chañar, and algarrobo trees). This does not mean that the agropastoral strategy was abandoned. It is likely that these practices continued to be developed in the newly colonized areas.
Finally, the reorganization phase (α) began around 15th century, with the reoccupation of the area. Despite the emergence of archeological sites with new architectural characteristics, the earliest evidence suggests the reactivation of the agropastoral system.
Conclusions
This paper examines human ecodynamics between the 3rd and 17th centuries in northern La Rioja, with an emphasis on subsistence strategies. We identified changes in consumption patterns that were related to fluctuations in environmental conditions and demographic trends. Based on archaeofaunal, demographic and environmental we propose that between 200 and 1000 CE, wild and domestic camelids were the most important prey for human populations. However, the adaptive success of these practices, along with improved environmental conditions, facilitated demographic growth. This resulted in increased pressure on wild camelids and a reduction in their availability. People adjusted their strategies by incorporating small fauna into their diet while maintaining the traditional agropastoral system. In a context of worsening conditions and the onset of increased aridity, local populations migrated, leading to the abandonment of the area. In this context of heightened mobility, the consumption of artiodactyls declined, while the hunting of dasypodids, rodents, and birds, the gathering of rheid eggs, and the exploitation of Geoffroea decorticans (chañar) and Neltuma spp. (algarrobo) gained greater significance. Finally, from the 15th century CE onward, a new phase of village occupation began, albeit with distinct characteristics, involving the reactivation of agro-pastoral systems.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836251358699 – Supplemental material for An approach to human ecodynamics and resilience in the economic trajectories of agropastoral societies in northern La Rioja (Argentina) from the 3rd to 17th centuries CE
Supplemental material, sj-xlsx-1-hol-10.1177_09596836251358699 for An approach to human ecodynamics and resilience in the economic trajectories of agropastoral societies in northern La Rioja (Argentina) from the 3rd to 17th centuries CE by Enrique Garate in The Holocene
Footnotes
Acknowledgements
I thank Pablo Cahiza for his support with fieldwork and for reviewing this paper. I also acknowledge CONICET for providing me with a doctoral fellowship to conduct this research.
Author contribution
Data availability statement
Supplemental material for this article is available online.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: ANPCyT through the PICT GRF2018-2138 project, and Secretaría de Investigaciones y Posgrado, Universidad Nacional de Cuyo (06/G787).
Ethical considerations
This article does not contain any studies with human or animal participants.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.