
Abstract
Pastoralism was first established in East Africa in the arid lowlands adjacent to Lake Turkana, northern Kenya, ~4800 years ago. Currently, there are limited palaeoecological records documenting the environmental conditions at the time of this transition, especially for areas east of Lake Turkana. To address this knowledge gap, sedimentary cores were recovered from the Chalbi Basin, Marsabit County, Kenya. These document a detailed environmental history of this arid region derived from evidence from pollen, plant and insect macrofossils, chemical analysis and diatoms, since the end of the Mid-Holocene ~5200 years BP. The high resolution analysis indicates the presence of variable freshwater and saline lake conditions later transitioning to a wetland. Diatom composition from the basin indicates that the water body varied widely in conductivity between ~200 and >20,000 μS cm−1 and 7.5–11.5 pH. The palaeolake and wetlands occasionally exceeded diatom tolerance limits and may have intermittently desiccated. The microfossil record shows that there was a broad trend towards more arid conditions at the end of the Mid-Holocene after ~4650 BP, interrupted by short intermittent wetter conditions, reflecting changes across the wider ecosystem. During the past 3300 years, the wetland changed from more terrestrial conditions with relatively abundant gallery tree taxa (Acalypha, Syzygium and Phoenix) to a Cyperaceae-dominated margin, and more consistent shallow water conditions. Fires were infrequent at the site, with the largest charcoal concentrations observed at 310 cm (3100 cal BP); thereafter, the magnitude of peak charcoal decreased and signs of fire are absent in the most recent sediments.
Keywords
Introduction
The consequences of current environmental change in most arid regions across the globe are highly pronounced and include rapid changes to vegetation, land cover and water resources, through drought, flooding and erosion impacts, all of which are increasingly common. Understanding the societal interactions within these dynamic and changing environments is crucial to determining the potential for agriculture, herding, transhumance patterns, conservation and resource management. A long-term characterisation of environmental change and societal context is important to sustainable resource management and adaptation for groups living in these regions. Equally important is the need to place recent changes and responses in a longer term perspective to determine whether particular environmental patterns are periodic or novel, and also to understand how past societies adapted to climate change and managed resources. The lack of detailed observational records extending beyond approximately the past century across much of sub-Saharan Africa (with some important exceptions) requires other sources of evidence for retrospective analysis of disparate palaeoenvironmental, archaeological and historical records. These records have marked differences in spatial and temporal resolutions, with low geographical coverage and chronological precision.
This is particularly so for the landscapes of northern Kenya, even though several paleoenvironmental studies and related archaeological data sets are available. The former have typically focussed on Quaternary deposits, especially those documenting Holocene lake-level variability (Bloszies et al., 2015; Butzer, 1980; Forman et al., 2014; Garcin et al., 2012; Owen et al., 1982) and palaeoecological conditions around Lake Turkana (Kinyanjui et al., 2026; Mohammed et al., 1996). Archaeological research has also tended to focus on sites and deposits proximal to Lake Turkana, which is recognised as having been an important locus of activity by hunter-fisher-forager groups during the African Humid Period (AHP: c. 14,800–5500 BP). The Lake Turkana region is a key locality for the tracing the initial appearance of food production in East Africa and subsequent development of early herding economies during the Mid-to- Late-Holocene (Barthelme, 1985; Lane, 2013; Marshall and Hildebrand, 2002; Robbins, 2006). Analysis of Holocene hydroclimatic variability and lake level variations have also revealed important human-environment interactions at millennial time scales proximal to the lake (Ashley et al., 2011; Goldstein et al., 2017; Prendergast and Beyin, 2018) and across nearby landscapes (Beyin, 2021; Keding, 2017; Kinyanjui et al., 2026; Lahr et al., 2016; Sawchuk et al., 2019; Wright and Forman, 2011). A recent synthesis of these records for the African Humid Period and beyond around Lake Turkana has also highlighted, in particular, the highly dynamic and spatially variable nature of changes in climate and socioecological strategies ‘across the AHP and immediately thereafter’ (Hildebrand et al., 2022: 1380).
By way of contrast, much of the spatiotemporal complexity of environmental change across the surrounding semi-arid and arid areas of northern Kenya during the Middle-to-Late-Holocene remains largely unexplored (Marchant et al., 2018), compared to other semi-arid and arid regions of Kenya (Ashley et al., 2004; Gillson, 2004; Githumbi et al., 2021a, 2021b; Goman et al., 2020). To address this knowledge gap, five sets of new sedimentary cores were recovered during late 2020 from sites in the Chalbi Basin, Marsabit County, Kenya. The core collection was part of a wider investigation of the long-term history and archaeology of water use and water management by pastoralist communities of northern Kenya and southern Ethiopia (M’Mbogori et al., 2022; Ochungo et al., 2022) and associated knowledge co-production with community members (Lane et al., 2025; M’Mbogori et al., 2024). We report here on the analysis of the longest of these new sediment records, recovered from Erenderi—a wetland site on the eastern side of the Chalbi Desert (Figure 1). The objective of these analyses was twofold. Firstly, to characterise the relatively local-scale variability in the paleoenvironmental record of a wetland ecosystem within the arid Chalbi Basin desert during the regional aridification from the Mid-Holocene to the present. Secondly, to interpret the ecosystem variations to known land use and livelihood changes interpreted from archaeological sources across the wider region.

Inset maps of (a) Africa and (b) East Africa. (c) Study area location map of Marsabit County, northern Kenya, showing archaeological sites, settlements, hand-dug wells and other locations mentioned in the text compiled from: Phillipson (1977), Phillipson and Gifford (1981), Stiles and Munro-Hay (1981), Tiki et al. (2011), Wright and Forman (2011), Wright (2019) and Ochungo et al. (2022). The reconstructed paleoshoreline of paleolake Chalbi along the 410 m asl contour is shown as a white polygon within the Chalbi Basin (based on Nyamweru, 1986: Figure 2). Basemap: Google Earth, 2025 https://earth.google.com/web/). (d) Desert margins near Kalacha. (e) View across Chalbi Playa south of Kalacha. (f) Erenderi 4C coring location. (d) and (f) Photographed July 2022 by Paul Lane; E Photographed by Freda M’Mbogori, August 2023.
Regional setting
The Marsabit region covers a surface area of 70,961 km2 and comprises low-lying arid and semi-arid plains and intermittent highland areas, such as Mount Marsabit and Mount Porr. Highlands are important sources of orographic precipitation (Los et al., 2019) that recharge the groundwater and surficial waters in wetland areas (Ingraham and Matthews, 1988; Sklash and Mwangi, 1991), which are important for fauna and ecosystem services (Galwab et al., 2023). The surface water of the lowlands includes ephemeral streams, incised rivers and intermittent wetlands. The region is characterised by hot weather, water scarcity and ecological variability and has experienced recurring, extended droughts throughout the last century and beyond (Mutua and Runguma, 2020; Oba, 2024; Oba and Tiki, 2000: 43–66). As in other arid regions, pastoralist communities in northern Kenya and southern Ethiopia have developed a range of adaptive strategies (from choice of livestock to water management, and from transhumance patterns to cultural exchanges) to changing conditions, both in recent decades (Cuni-Sanchez et al., 2019; Dabasso and Okomoli, 2015) and over longer, centennial scales (Robertshaw, 2021; Wright, 2019). Correspondingly, pastoralist practices across the county and adjacent areas are typically characterised as being highly dynamic and adaptive, comprising high mobility, choice of hardy livestock, livelihood flexibility in line with a transforming array of daily resources and pressures, and related diversification of preferred subsistence strategies to include foraging and fishing (Robinson, 1985; Robinson and Berkes, 2010; Roth, 1996; Sobania, 1988). Modes of prosocial adaptive strategies may have characterised pastoralist practices in the region since its regional inception c. 5000 years ago (Wright, 2019; Wright et al., 2015). Determining the antiquity of these socio-ecological systems still requires further targeted, integrated palaeoecological and archaeological and paleoenvironmental research.
Chalbi Desert
Located between longitudes 37°05′ and 37°47′ E and latitudes 2°22′ and 3°20′ N, the Chalbi is the world’s lowest latitude desert and marks the location of a former large, shallow, freshwater lake likely in existence during the early Holocene between c. 11,000 and 9500 years BP (Abell and Nyamweru, 1988; Nyamweru, 1986, 1989). The timing of Lake Chalbi’s formation is thus broadly coincident with a phase of increased regional precipitation during the AHP, as evidenced by a rise in Lake Turkana between c. 11.5 and 10.5 kya and overflow from the Suguta Basin to the south (Bloszies et al., 2015; Garcin et al., 2009: 23). Sedimentary records around Maikona and Kargi on the eastern side of the Chalbi also attest to a later, and probably the last, significant pluvial episode after 4.4 ± 0.3 cal ka BP, broadly coincident with the final regression of Lake Turkana (Stinchcomb et al., 2026: 39). Lying within an intermontane basin at an altitude of c. 370 m asl and covering over 1000 km2, the Chalbi today is a flat, intermittently flooded, ‘salt-crusted playa’ that currently functions as ‘an alluvial aggradation surface’ (Nyamweru and Bowman, 1989: 131). It is bounded on its western and eastern sides and at its southern end by volcanic and sedimentary rocks formed during the lower Cretaceous to Quaternary, and except in areas of localised dune formation, the margins are typically delimited by a low, sparsely vegetated, scarp slope strewn with lava boulders. Patches of bedded sands, diatomaceous silts and fossilised shells of freshwater gastropods around the margins of the playa indicate the surface level of the former lake may have reached c. 410 m AOD (Above Ordnance Datum; Nyamweru, 1989: 184). At its northern end, there is a narrow break in the lava scarp where the clay playa grades into rockier, undulating terrain extending to beyond North Horr. Median annual rainfall ranges between 150 and 200 mm and there can be several consecutive years without any rain. Temperatures range from approximately 18°C–36°C, and fierce winds driven by the Turkana Jet are common (Munday et al., 2022; see also
Background archaeological evidence
Compared with the shores of Lake Turkana to the east, the archaeology of the Chalbi Basin has received limited study (Phillipson, 1977; Robbins, 1980; Stiles, 1982). Evidence for human occupation and utilisation of the margins of the Chalbi Desert during the Holocene is attested, nonetheless, by the remains of dense concentrations of different types and forms of stone cairns and the traces of diverse forms of low, stone-walled enclosures. Surface traces of deflated habitation sites, marked by the presence of pottery fragments, stone tools, cores and related flaking debris, fragmentary stone bowls made on pumice and other volcanic rocks, ostrich eggshell beads and occasional animal bones also occur. Several undated sites with rock engravings are also known.
Research around North Horr, c. 90 km east of Lake Turkana and at the northern end of the Chalbi Desert has identified several partially deflated scatters of lithics, pottery, faunal remains, ostrich eggshell beads and other artefactual remains dating from c. 5000 to 600 BP (Phillipson, 1976, 1979; Stiles, 1982). The few excavated sites all exhibit signs of stratigraphic mixing and aeolian deflation, with the best preserved suggesting several distinct phases of occupation initially by Late Stone Age (LSA) foraging communities exploiting a range of wild ungulates including zebra and gazelle and later inclusion of domestic stock (cattle, ovicaprids) associated with the beginnings of the Pastoral Neolithic (PN; Wright, 2019). The lithics from all horizons are fairly characteristic of terminal LSA material found elsewhere in the region, and are dominated by backed microliths, with scrapers being notably rarer. Ceramics were also present, with some in the lower levels bearing similarities to better documented LSA wavy-line forager pottery from around Lake Turkana and areas to the north (Keding, 2017). Subsequent horizons include Nderit-like pottery, typical of the local early PN (Grillo et al., 2022) and stone bowls (also diagnostic of the early PN), as well as late PN Turkwel tradition ceramics (~1800–950 BP), which also occur on other late PN sites proximal to Lake Turkana such as Lopoy and Apeget 1 (Robbins, 1980).
Numerous stone cairns of different forms (mostly simple mound, ring, double ring and kerbed types) occur in their thousands across the Chalbi landscape and have been mapped through a combination of ground survey and remote sensing (Lane et al., 2025). The main concentrations around the Chalbi Desert on the east side are (from south to north) near Korole, Koroni, Maikona and Kalacha and on the west side around Balesa Kulal and Malabot (Stiles and Munro-Hay, 1981; Figure 1(c)). In the 1980s, 10 cairns were excavated on Kokurmatakore, a boulder-covered hill east of Kalacha, with eight shown to contain single inhumation burials, four of which could be sexed and assigned an approximate age (Stiles, 1980, 1981, 1982; Stiles and Munro-Hay, 1981). The available radiocarbon dates suggest the cairns were used over several millennia between c. 4100 and 330 cal. BP, with the earliest type (small mound) likely associated with early PN herders.
Another important feature of this landscape are the clusters of hand-dug wells, concentrated around Balesa, Elhadi, North Horr, Elmuda, Kalacha, Maikona and Kargi (Figure 1(c)). These exhibit considerable variation in form, depth and associated architectural features, but can be broadly differentiated into ‘shallow’ and ‘deep’ types, with various sub-categories and different examples may well be of different origin (Ochungo et al., 2022; Tiki et al., 2011). Available oral histories attest to the initial construction of the deeper wells some 600 or more years ago and attribute their initial construction to a semi-mythical people called ‘Wardai’, who are held to have inhabited this area before the arrival of the current inhabitants (Tablino, 1999).
Currently, the area is inhabited mostly by Gabra, Rendille, and Boran pastoralists (all speakers of related Afro-Asiatic languages), while Samburu, Dassenatch, Waata and Turkana comprise most of the other ethnic groups in the wider area. In common with other parts of arid and semi-arid northern Kenya and southern Ethiopia, the recent settlement history of the region has been characterised by recurrent mobility and the fusion and fracturing of ethnicities. Reconstructed clan histories are distinctly rhizomic in structure (Schlee, 1989), indicative of variable ethno-linguistic affiliations and diverse adaptive responses to shifting climatic conditions and distributions of natural resources, especially water and pasture (M’Mbogori et al., 2022). Camel, sheep and goats are the predominant livestock and all groups practice some form of seasonal mobility. In the recent past many of these groups, such as the Gabra (Stiles, 1984, 1992), were entirely nomadic, exploiting the network of ancient, hand-dug wells located mostly along ephemeral watercourses (also known as lagas or luggas) and oases around the desert margins (Hazard and Adongo, 2015). Many of the latter are now the focus of permanent settlement, as sedentism has increased since the end of colonial rule and continues to be stimulated by the way services are provided by government policies and drought relief interventions (Fratkin and Roth, 2005; Haji and Legesse, 2017; M’Mbogori et al., 2022; Semplici, 2020).
Study site
The Erenderi wetland site lies within the Chalbi Basin, at the centre of which is the Chalbi playa (Figure 1), a bare surface of mud subject to occasional inundation at c. 370 m above sea level (Figure S1, available online). The wetland surface is dominated by semi-continuous patches of Cyperaceae and some Poaceae. Near the wetland Vachellia tortilis (acacia), Salvadora persica, Hyphaene coriacea, Cordia and terrestrial Poaceae and Cyperaceae taxa were observed.
Methods
Fieldwork and sample partitioning
Fieldwork was undertaken from 15 to 19 December 2020, during which five sedimentary cores were taken from different waterlogged sedimentary deposits. The deepest wetland sediment stratigraphy was observed at the Erenderi wetland site. The Erenderi 4C core was collected from the deepest accumulation here (3°06′ 22.34″ N, 37° 22′ 57.46″ E; 367 m asl, WGS84) using a hand-pushed Russian corer with a 50 cm long, 5 cm diameter hemicylindrical chamber (Belokopytkov and Beresnevich, 1955; Jowsey, 1966). The sediment cores were wrapped in plastic and aluminium foil for transportation from the site and were later stored in refrigerators at 4°C. Pollen and diatom analyses on the entire core profile were undertaken at the National Museums of Kenya, Nairobi and an analysis of sieved retained remains (plant, charcoal, chitinous invertebrates) and loss-on-ignition analyses were undertaken on the uppermost stratigraphy at Geoecology, University of Basel, Switzerland (Heiri and von Fumetti, 2022).
Laboratory methods and data processing
Bulk sediment samples or picked organic remains were removed from the core and AMS radiocarbon dated at DirectAMS (Bothwell, WA, USA) or Poznań Radiocarbon Laboratory (Poland). Bulk samples are the most predominant radiocarbon dating source of sediment cores in the region and increasingly, picked organic detritus has been targeted (Courtney-Mustaphi et al., 2026; Courtney Mustaphi and Marchant, 2016). Radiocarbon ages were calibrated with the IntCal20 calibration curve (Reimer et al., 2020) and a linear interpolation age model was developed with the clam package (version 2.4.0; Blaauw, 2010) with R scripts (version 4.1.2; R Core Team, 2021) for the top dates that had the most confidence for representing the accumulation. The entire core was initially analysed for microfossil remains (pollen, diatoms and radiocarbon) and the top half was contiguously analysed for sieved organic remains (plant remains, charcoal, invertebrates) to characterise the local environment during the Late-Holocene.
Microfossils and macroremains
Microbotanical remains (pollen and diatoms) were examined from the entire 625 cm Erenderi 4C core. The pollen study is used here to reconstruct long-term vegetation dynamics that reveal information about past changes in terrestrial and aquatic environments while the diatoms are used to infer water quality changes and basin developmental histories that may have influenced the movement and settlement of inhabitants in the study area.
The top half of the core was also analysed for sieved charcoal to assess local fire dynamics, invertebrate remains and sediment organic content as described below.
Pollen samples were processed from 1 cm3 subsamples (n = 250) at 2.5 cm intervals and were sequentially digested with warm dilute HCl (10%), then KOH (10%) and room temperature HF (48%; Faegri and Iversen, 1989; Moore et al., 1991). Residues were mounted with glycerol onto microscope slides and pollen was identified under optical microscopy at 400× magnification. For each sample, at least 350 pollen grains were counted except in a few samples where preservation was poor. Identification was confirmed by comparison with a reference collection of over 6000 slides of modern East African pollen held at the Department of Earth Sciences (Palynology & Palaeobotany section) at the National Museums of Kenya (NMK). Identifications also made use of a range of publications (Hamilton, 1975, 1982; Marchant, 1997; Vincens et al., 2007) and digital photographs of pollen types obtained from the African pollen database website (
Diatoms preparations used 1 cm3 subsamples (n = 250) at 2.5 cm intervals and involved the use of Calgon sodium hexametaphosphate (Na6[(PO3)6]) solution (5 g L−1) to disaggregate samples over 24 h in warm water. The samples were then washed with distilled water. In a fume cupboard, 10% HCl was added to the samples that were placed in a hot water bath and left overnight to cool and settle, with the samples again washed with distilled water. Samples were then heated in a beaker with 30% hydrogen peroxide (H2O2) with multiple washing in distilled water (Bushozi et al., 2022; Muiruri et al., 2021a, 2021b; Owen et al., 2008). Mounting was done using Naphrax and the diatoms were spread out on the slide and 400 frustules were counted per slide. Identifications were supported by the works of Gasse (1986) and the East Africa training set in the EDDI Diatom Database (http://craticula.ncl.ac.uk/Eddi/jsp/index.jsp). Taxonomic identifications were made at 1000× magnification with reference keys (Gasse, 1986; Krammer and Lange-Bertalot, 1986, 1988, 1991a, 1991b) and nomenclature was updated using taxonomies from Algaebase (Guiry and Guiry, 2021).
In samples where at least 50 diatom valves were counted, diatom abundances were analysed using indirect ordination using Canoco5 software (Šmilauer and Lepš, 2014). Detrended correspondence analysis showed a relatively small gradient present (2.1 standard deviation units) across the Erenderi diatom dataset (Hill and Gauch, 1980). The dataset was subsequently analysed using principal components analysis (PCA) to reveal major patterns. Relative abundance data were square-root transformed to stabilise species variance prior to ordination analyses.
Sieved sediment analysis was done at 2.5 cm interval sampling resolution from 0 to 310 cm depth (n = 132). A calibrated subsampler (Knowledge Core GmbH, Switzerland) was used to extract 1 cm3 (0.997 cm3) of field-wet sediment subsamples that were each sieved with water through a 100 μm mesh. The retained fraction > 100 μm mesh was transferred to a Petri dish with water and manually probed with a metal pick and the total charcoal pieces were counted (Hawthorne et al., 2018; Whitlock and Larsen, 2002). This charcoal size fraction is often applied to macroscopic charcoal records in the region (Courtney Mustaphi et al., 2025; Temoltzin-Loranca et al., 2023a) and in other grasslands (Leys et al., 2015). Anecdotally, subfossil invertebrate remains were picked from the same sample with forceps and mounted on microscope slides with Euparal medium (Brooks et al., 2007).
To estimate sediment organic content, wet sediment subsamples of 1 cm3 (n = 132) were weighed, dried in an oven at 105°C for 24 h, then reweighed, then burned in an oven at 550°C for 6 h, reweighed and reheated at 950°C for 2 h and reweighed again to estimate carbonate content (Heiri et al., 2001; Wang et al., 2011).
Numerical techniques and data visualisation
The percentages of the two sets are presented using the TILIA program 1.7.16 (Grimm, 2011). The results are described below under the various Units I–VI and are plotted in Figures 2 and 3. Pollen types that did not exceed 2% were excluded from the pollen diagram. CONISS was applied using a numerical clustering package within the TILIA programme, with the results identifying six stratigraphic clusters of samples with similar floristic composition.
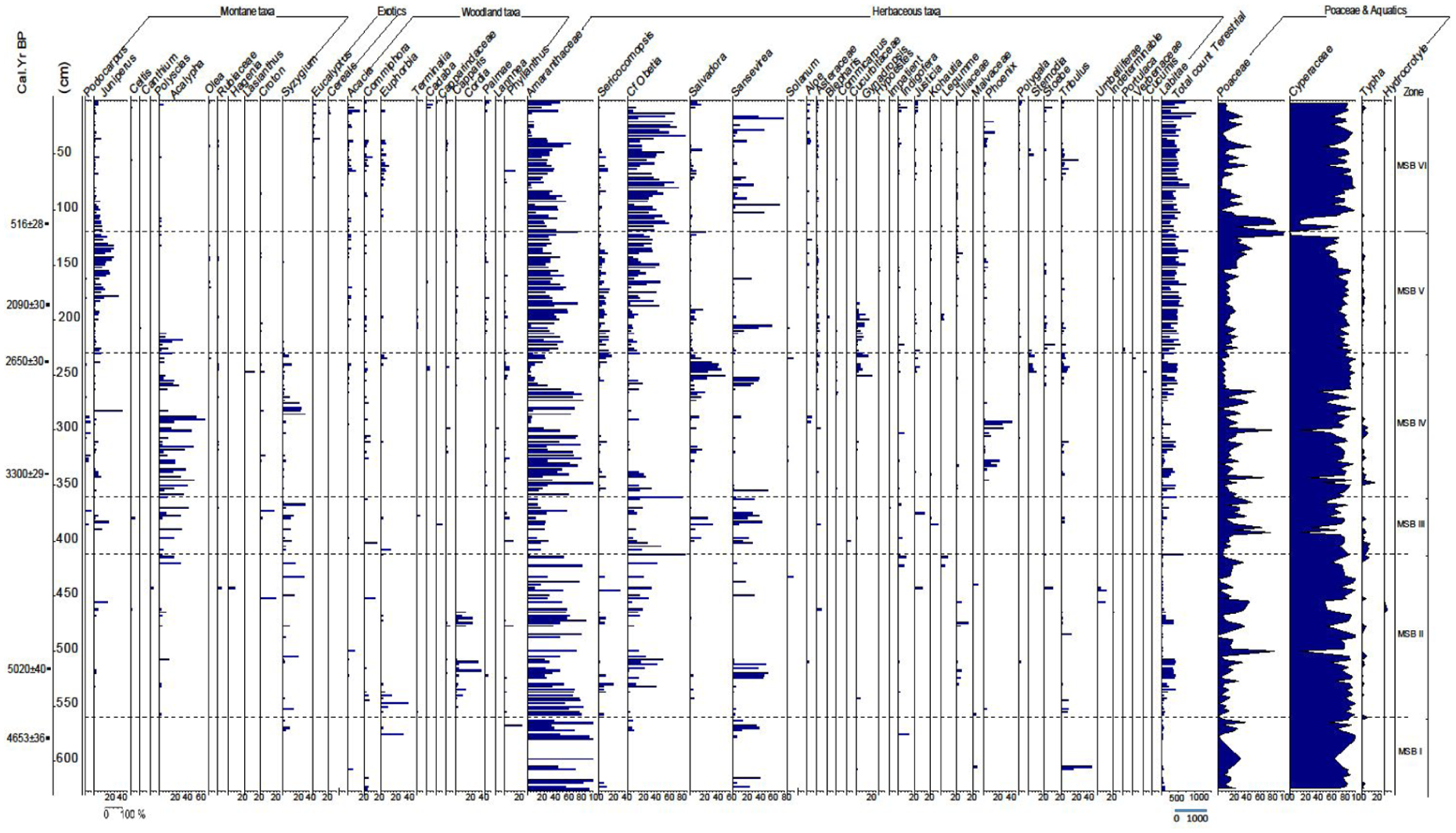
Stratigraphic pollen assemblage of Erenderi (core Eren4C) by relative abundances (%) and grouped taxa of Montane, Exotics, Woodland, Herbaceous, Poaceae and wetland groups (Githumbi et al., 2025; Hamilton, 1982; Muiruri et al., 2021) plotted by depth and radiocarbon age determinations (at left). Pollen assemblage zones were identified using stratigraphically constrained cluster analysis (CONISS, dashed horizontal lines; Bennett, 1996; Grimm, 1987).
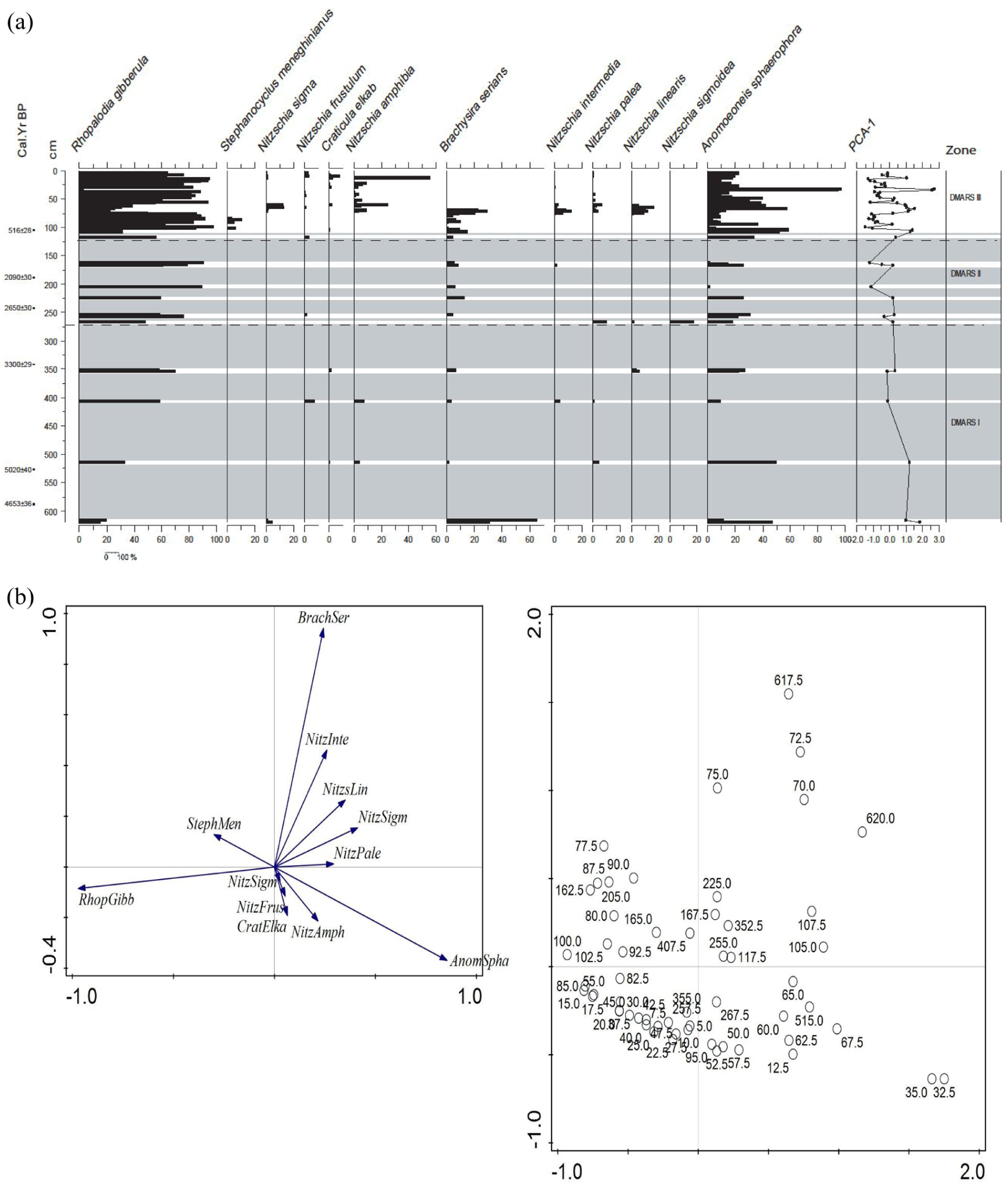
(a) Erenderi (core Eren4C), Diatom stratigraphy, showing down core percentages of diatom assemblages. 14C age determinations and depth to the left. Taxa arranged into habitat preferences according to Gasse (1986). Diatom assemblage zones were identified using stratigraphically constrained cluster analysis (CONISS, dashed horizontal lines; Grimm, 1987; Bennett, 1996). Shaded areas are diatom-free levels. (b) Stastical representation of fossil diatom floras, showing PCA biplot, axis1 and 2 scores.
Results
Stratigraphy and chronology
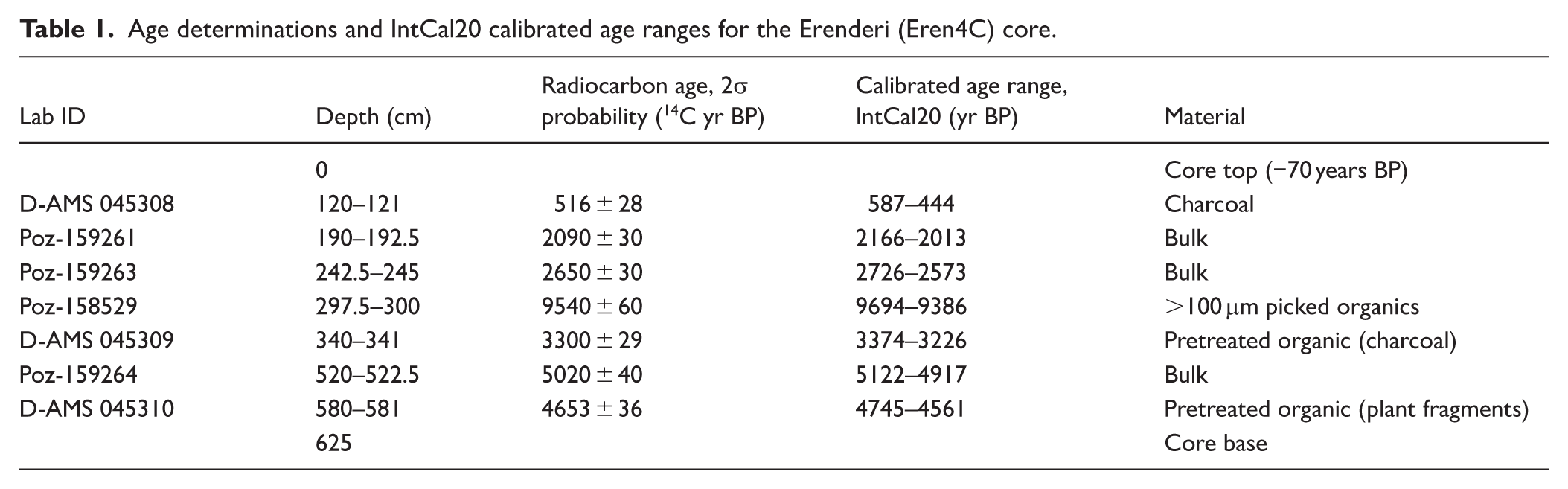
A 625 cm stratigraphy was recovered from Erenderi (core Eren4c). The stratigraphy was complex and had strong apparent changes in colour with several thick and internally structureless (massive texture), more organic brown-to-reddish and brown-to-black beds, interbedded with greyer to greyish-light-brown beds from c. 0 to 314 cm. Thin layers of various grey-to-brown beds were present from 314 to 350 cm, and from 350 to the base of the core, were substantial grey to greyish light brown beds with a partially blackened layer at 580 cm. The reconstructed chronostratigraphy of the core is based on seven radiocarbon samples, and extends from the end of the Mid-Holocene to present (Table 1 and Figure 4) with a mean accumulation rate of 0.12 cm/year. The organic matter at 297.5–300 cm sampled for dating (Poz-158529) appears very old and may reflect old carbon from within the catchment or may have been contaminated during sample preparation. The two deepest dated samples (320 and 580 cm) yielded reversed age determinations, although taken together the dates suggest a Mid-Holocene age for the basal depths of the recovered core. The age-depth model below 340 cm depth is considered to have large age uncertainty and likely to be associated within the latter part of the Mid-Holocene (8200–4200 cal. yr BP).
Age determinations and IntCal20 calibrated age ranges for the Erenderi (Eren4C) core.
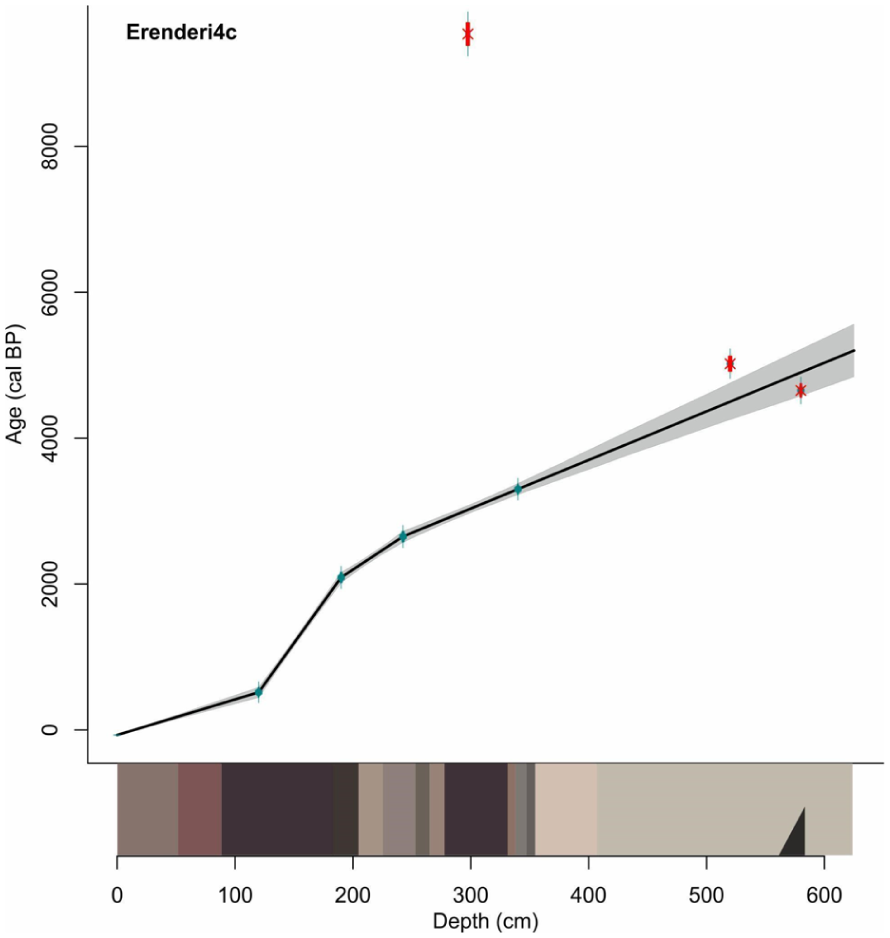
Linear interpolation age-depth model of the Intcal20 (Reimer et al., 2020) calibrated radiocarbon ages (Blaauw, 2010) of the Erenderi core (Eren4C) with stratigraphy at bottom. The upper sediments are continuous accumulation and the deeper sediments, 340–625 cm, are more uncertain and ages for this section are indicative of the latter part of the Mid-Holocene.
Pollen stratigraphy
Pollen and diatom analyses (n = 250 samples each) indicate variations in regional terrestrial vegetation in the catchment and among the local palaeolake aquatic macrophytes. Core Erenderi 4C includes 83 pollen taxa, with 80 belonging to trees, shrubs, lianas and herbs. These can be used to recognise six major vegetation zones, some of which can be further divided into subzones as listed below. Pollen is absent intermittently through the sequence and the descriptions focus on percentage data where pollen are present.
Two sets of fossil pollen sums were calculated. The first consists of the total sum of Poaceae and wetland and immediately local taxa (Cyperaceae, Hydrocotyle and Typha) minus regional taxa or the land pollen and spores. Poaceae was considered as local taxa because of a combination of predominance among the fossil pollen assemblage (local overrepresentation) and the numerous grasses growing within the Erenderi wetland in the Chalbi Basin today (uniformitarian).
The second fossil pollen sum includes regional taxa or the land pollen with unknown and undifferentiated pollen grains included. The trees were grouped together as Afromontane taxa while exotics were grouped together, with shrubs and lianas forming woodland taxa while all the herbs were collated as herbaceous taxa. The three terrestrial groups (land pollen) were regarded as regional, although some of the herbs can also be identified from both local and regional taxa. In this case, they are all included in the pollen sum of the regional taxa.
Pollen zones since the Mid-Holocene
Pollen zone MSBT I (625–560 cm) 5200–4770 cal BP
This zone starts at 625 cm and is distinguished by several hiatuses, where pollen are either few or lacking in sediments. It is also characterised by a relatively high percentage of Cyperaceae, interchangeably occurring with Poaceae, together with a sporadic appearance of herbaceous taxa notably Amaranthaceae, Sansevieria and Tribulus from different levels. Other recorded taxa with less than 30% include, Syzygium, Vachellia/Senegalia (Acacias), Commiphora, Euphorbia, Phyllanthus, Sericomopsis, cf. Obetia, Indigofera and Malvaceae.
Pollen zone MSBT II (560–412 cm) 4770–3780 cal BP
This zone is developed within a wavy bedded clay mud with silicified nodules, an ash layer, organic detritus and carbonate mud. The zone is characterised by the emergence of a variety of species with a higher representation than the previous zone. Cordia, Amaranthaceae, cf. Obetia, Sansevieria distinguish the zone. Montane taxa start to appear from this zone at irregular intervals that is, Acalypha, Hagenia and Syzygium ranged between 5 and 30%. Cyperaceae (20–80%) form a higher percentage than Poaceae (20–60%) and Typha (5–10%) as local taxa. Afromontane taxa includes Syzygium in a greater abundance than observed in the lower zone, forming 5–35% of the flora. Juniperus appeared for the first time (5–10%), while Polyscias, Hagenia and Croton occurred once in this zone. In the woodland taxa group, Cordia this zone followed closely by Euphorbia (5–40%) and a few Commiphora, Vachellia/Senegalia (acacias), Phyllanthus (5–10%) and Palmae (2%) which just appeared once. Herbs, such as Amaranthaceae, Sericomophosis, cf. Obetia and Sansevieria, ranged between 5 and 80% and formed the major component of the vegetation in this zone. Liliaceae, Tribulus and Indigofera appeared in different intervals ranging between 2 and 5%, while Solanum, Asteraceae, Gynandropsis, Justicia, Polygala and Stoebe appeared once with a range of 2–5%.
Pollen zone MSBT III (412–360 cm) 3780–3430 cal BP
The zone includes a significant content of Poaceae and Cyperaceae, which account for about 40–80% each, with a rare Typha (5–10%). The Afromontane taxa Podocarpus appeared for the first time from 5 to 10% of the pollen, while Acalypha and Syzygium dominated with 60 and 40% respectively. Juniperus, Croton constitute about 20% each, Celtis appearing once with 5% at the middle of the zone. Among the woodland taxa, Commiphora, Euphorbia, Terminalia, Capparindaceae and Phyllanthus formed between about 2.5 and 10%. The herbaceous taxa notably include Amaranthaceae (up to 80%), cf. Obetia formed between about 10 and 90%, Salvadora (up to 40%), Sansevieria (up to 50%), Asteraceae, Cucurbitaceae, Indigofera, Justicia, Kohautia and Tribulus formed between about 2 and 20%.
Pollen zone MSBT IV (360–230 cm) 3430–2520 cal BP
The pollen zone is categorised by high percentages of Cyperaceae and Poaceae (10–90%), with Typha ranging to about 2–20% within the local taxa. Within the Afromontane component, Acalypha, Juniperus and Syzygium are common (5–60%) unlike Podocarpus which only appears in a few levels with ⩽5%. Herbaceous taxa that make a common appearance include Amaranthacae, Salvadora, Sansevieria, cf. Obetia, Sericocomopsis, Gynandropsis and Phoenix all between about 10 and 60%. Others were below 10%; of these, common taxa include Solanum, Aloe, Asteraceae, Commicarpus, Indigofera, Justicia, Stemodia and Tribulus.
Pollen zone MSBT V (230–120 cm) 2520–515 cal BP
The pollen zone is distinctive in comparison with the other zones. Cyperaceae are present in slightly higher percentages than Poaceae in multiple levels apart from the top of the zone where they appear interchangeably, with rare Typha and Hydrocotyle. Within the Afromontane component, Juniperus was the most common taxon followed by Acalypha at the base of the zone, while others (Olea, Croton) appeared insignificantly and the same applies for the woodland taxa. Amaranthaceae, cf. Obetia, Sericocomopsis, Salvadora, Sansevieria, Gynandropsis among the herbaceous taxa, were also significant.
Pollen zone MSBT VI (120–2
5 cm) 515 cal BP–present. The pollen zone is distinctive and based on increasing cf. Obetia and Sansevieria from the bottom towards the upper part of the zone and relatively high percentages of Cyperaceae compared to Poaceae apart from the bottom of the zone where they appear interchangeably. Exotic pollen types (Eucalyptus, Cereals) had a notable appearance only in this zone. Other herbs are rare, apart from Indigofera, Phoenix and Tribulus which ranged 2–10%. Vachellia/Senegalia (acacias), Commiphora, Euphorbia, Cadaba, Phyllanthus and some few Palmae appeared regularly as woodland taxa.
Diatom zones since the end of the Mid-Holocene
The number of diatom valves were either too few (less than 50) or absent altogether from the majority of samples, especially those in the lower and middle parts of the core, 625–120 cm (Figure 3(a)). Diatom dissolution (linked to high salinity and very alkaline pH) is very likely the main reason for the absence of diatoms in many samples, rather than no diatom growth at all. However, diatom valves were sufficient in numbers to be counted in all samples, bar five from the top 120 cm and in isolated samples from the lower and middle parts of the core (Figure 3(a)). Only about 30 taxa were enumerated in the core, including those where a precise identification could not be established for example, due to poor preservation. For the purposes of ordination analyses and stratigraphical presentation, only taxa that occurred in samples greater than 5% are shown (n = 12). Two diatom species are present in every core sample where diatoms are preserved in sufficient numbers to be counted: Rhopalodia gibberula (Ehrenberg) O. Müller 1895 and Anomoeoneis sphaerophora Pfitzer 1871.
The first principle component (or axis 1) explains 40% of the variation in the dataset and is mainly characterised by the changing presence of Rhopalodia gibberula, several Nitzschia species and Anomoeoneis sphaerophora (Figure 3(b)). Axis 2 explains just over 20% variation in the data and is largely influenced by the taxon identified as Anomoeoneis sphaerophora var. guntheri. The diatom species have been arranged on the stratigraphy in Figure 3(a) according to their PCA axis1 species scores.
Diatom Zone DMARS I (625–310 cm) ~5200–3100 cal BP
The soft diatomite-muds in the lower part of DMARS I (Figure 3(a)) lack diatoms, but within this, part of the sequence includes dominant benthic species, dominated by Rhopalodia gibberula and Anomoeoneis sphaerophora, with the latter being a shallow water, moderate to high salinity diatom found in spring outflows (Barker, 1990). Notable other taxa with minimal presence include Nitzchia sigma, N. frustulum, N. palea, N. amphibia, N. intermedia, N. linearis, Caloneis bacillum and Navicula elkab.
Diatom Zone DMARS II (310–120 cm) ~3100–515 cal BP
Diatom diversity is low in DMARS II (Figure 3(a)) where there is also a general increase in saline taxa with benthic species, such as Rhopalodia gibberula and Anomoeoneis sphaerophora also increasing but with few Nitzschia palea, N. sigmoidea and rare N. intermedia and N. lenearis. Benthic diatoms are rare or absent in most horizons.
Diatom Zone DMARS III (120–0 cm) ~515 cal BP–present day
This zone is very diverse and contains mixtures of diatoms that inhabit fresh and saline environments. Rhopalodia gibberula dominate (20–90%), interchangeably with the halophyte Anomoensis sphaerophora (20–80%). Other freshwater assemblages include N. amphibia (10–50%), N. intermedia (5–20%), N. linearis (5–20%). Others with <10% include N. palea, Calloneis bacillum, Amophora veneta, Cymbella muelleri while Epithermia sorex, E. adata and E. urgus appeared in only one level with 10%. Planktonic Cyclotella meneghiniana and Rhopalodia gibba present (2–10%).
Sediment organic content
Loss-on-Ignition (LOI) organic values were highly variable (10–65%) and increased with peaks in charcoal, with the exception of the peak values at 40–15 cm depth. Carbonate content was negligible (<2.5%) from 310 to 90 cm and increased to ~4% consistently from 90 to 0 cm (Figure 5).
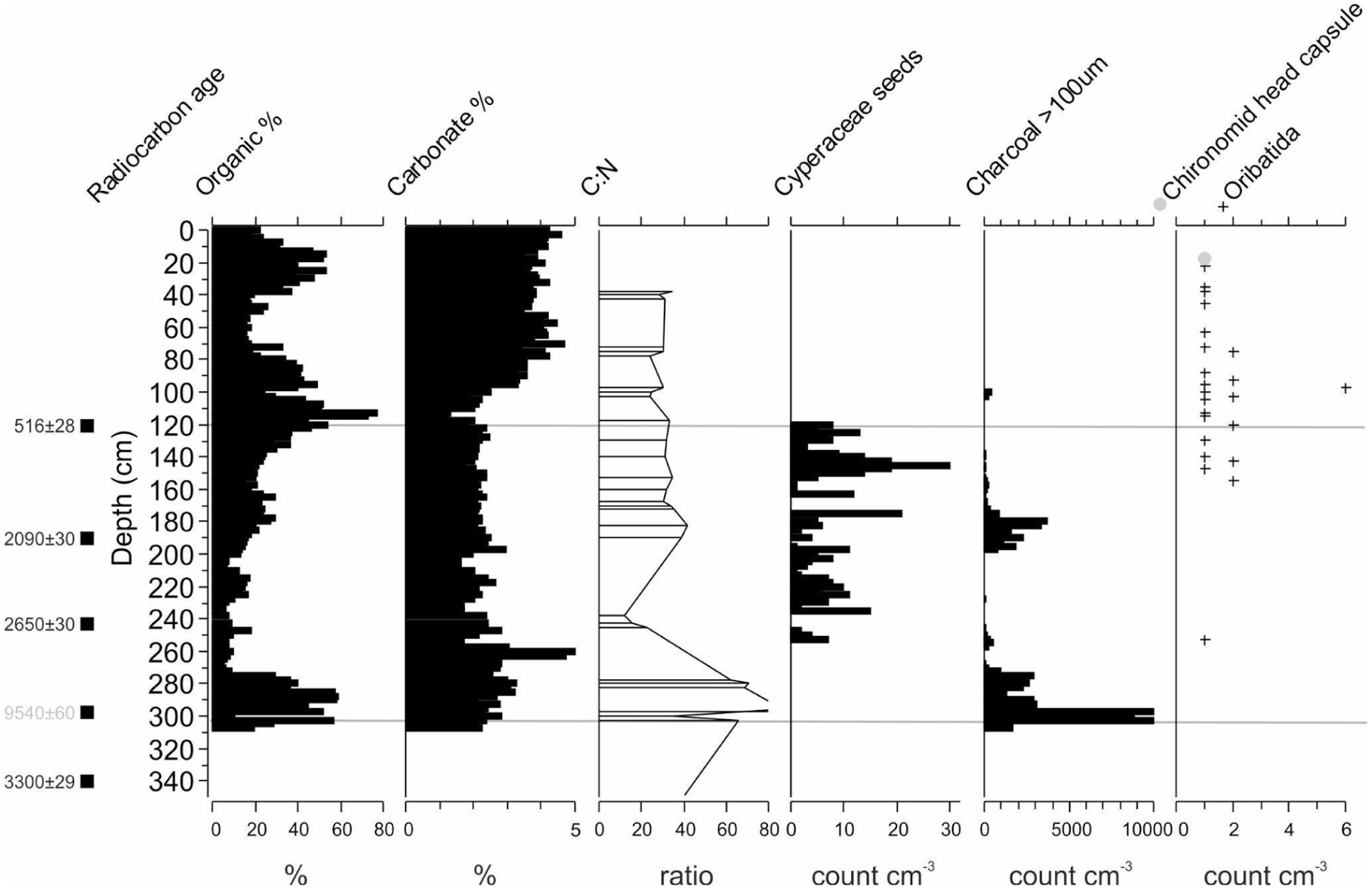
Erenderi (core Eren4C) data for the top 0–310 cm (past ~3300 years to present). Sedimentological data for loss-on-ignition estimates of organic and carbonate content and intermittent carbon-nitrogen ratios (CN). Sieved macro remains >100 μm presented in the text that includes Cyperaceae seeds, charcoal and invertebrate remains (see Figure S2, available online). Horizontal zonation (grey lines) of the diatom assemblages (Figure 4).
Charcoal and other plant and invertebrate remains (>100 μm sieve)
A total of 132 cm3 of sediment was sieved throughout the core, mostly in the top section 0–310 cm depth (3100 cal BP–present) and a few exploratory samples in deeper sections. Total charcoal had extremely high variability that ranged from 0 to 14,730 pieces cm−3 and higher values were concentrated at three depth intervals from 310 to 272.5 cm (3100–2850 cal BP), 200–140 (2200–965 cal BP), and 105–100 cm (440–420 cal BP). Cyperaceae seeds were occasionally found (n = 328) and varied from 0 to 30 seeds cm−3, with the majority found between 255 and 120 cm depth (Figure 5; 2730–515 cal BP). Invertebrate remains (>100 μm) throughout the uppermost stratigraphy of the core predominantly consisted of unidentifiable insect chitinous remains, with slightly more remains towards the top half of the core and varied from 0 to over 40 chitinous fragments cm−3. No carbonate invertebrate remains, such as shells, were observed. Unidentified remains included damaged appendage fragments of insects, Coleoptera elytra and some head parts (Courtney-Mustaphi et al., 2024). Some of the more intact chitinous remains were identified to the order or family taxonomic resolution and included rare occurrences of Insecta: Diptera: Chironomidae (n = 3 head capsules, not plotted), Diptera: Ceratopogonidae (n = 4, not plotted), and Acari: Oribatida mites (n = 35; Figure 5). No other Acari taxa were observed in the record. Diptera: Chironomidae head capsules were only found at two depths in the core and included one Chironomus (22.5–25 cm; ~40 cal BP) and two Paratendipes (517.5–520 cm; 4500 cal BP). Ceratopogonidae head capsules were found 32.5–35 cm (n = 1) and 97.5–100 (n = 3). Oribatida mites varied between 0 and 6 cm−3 and were found intermittently throughout the core with the majority found from 157.5 to 22.5 cm depth (1350–40 cal BP).
Discussion
Mid-Holocene to present palaeoenvironmental change in the wider Chalbi Basin
This section summarises evidence from proxy records from the Chalbi Basin and draws inferences from these data concerning the environmental history of the basin. The section focuses on changes in the composition and distribution of vegetation and suggests possible drivers of these changes. Pollen microfossils were preserved throughout the record unlike the diatoms which appeared intermittently while sediments from 310 cm to the surface were additionally analysed for macro remains and some additional depositional characterisations.
MSBT I (625–560 cm) ~5200–4770 cal BP
Zone MSBT I (Figure 2) is distinguished by the sporadic but high occurrence of Amaranthaceae on the background abundance of Poaceae and Cyperaceae. The pollen data (Figure 2, Zone MSBT I) suggest a series of dry periods given the presence of drought-related herbaceous (Amaranthaceae, Tribulus, Indigofera, Sansevieria) and woodland taxa (Euphorbia, Commiphora and Vachellia; Beentje, 1994). Amaranthaceae, as a family, are adapted to a wide range of climatic factors and soil conditions; they grow best in the sun or in light shade up to 1400 m elevation (Maundu et al., 1999). The Amaranthaceae family includes species such as Amaranthus, which could have contributed to this pollen, thrives in a habitat rich in nitrogen and prefers soils that are rich in nutrients although they can also grow on sandy and saline soils, especially if moist. They are drought-resistant and can even grow under very arid conditions (Maundu et al., 1999) and have been used as a good indicator of arid conditions in pollen palaeoenvironmental studies (Muiruri, 2017; Rucina et al., 2009).
The presence of abundant Cyperaceae and common Poaceae probably reflects the presence of a shallow lake. The Cyperaceae are a herbaceous plant family of terrestrial, semi-aquatic and emergent aquatic taxa with >40 genera in Africa that are mainly associated with the presence of moist temperate to wet tropical regions (Haines and Lye, 1983; Vrijdaghs et al., 2009). They include the sedges, which are mostly perennial or some annual herbs. In East Africa, sedges are often found in wetlands (some are entirely aquatic) and along watercourses, but there are also terrestrial taxa that are associated with moist grasslands and forest margins. In Kenya, the well-known papyrus (Cyperus papyrus) can totally dominate the margins of lakes and wetlands forming an impenetrable, floating mass of plants (Haines and Lye, 1983; Vrijdaghs et al., 2009). Most of the species are also well distributed in moist to wet habitats such as fresh and saltwater marshes, ponds and lakeshores, meadows, bogs, fens and savannahs. They often form an important component in the pollen record but are regarded as local taxa as most are restricted to aquatic ecosystems. A short, wet phase within and around 4600 cal BP, is suggested by an increase in Cyperaceae compared to Poaceae and the presence of Syzygium, Sansevieria and Phyllanthus suggesting nearby shallow water that allowed sedges to grow, which also supported diatom tolerance limit of R. gibberula and Anomoensis sphaerophora. This broadly wet phase gave way to drier conditions as indicated by the presence of drought-related taxa among bushland (Vachellia, Commiphora and Euphorbia), and herbaceous taxa (Tribulus, Indigofera). In the neighbouring Turkana Basin, a previous detailed sedimentologic study of a relict beach sequence in the Mount Porr and Lothagam areas identified up to at least three spillover or near-spillover, events dated ca. 9000, 6200 and 5000 years BP, each of which is bracketed by 30+ m transgressive-regressive phases in the lake elevation that occurred on sub-centennial timescales (Bloszies et al., 2015). The final fall in lake level occurred ca. 5000 years BP, when the lake began to lose significant amounts of water. By 4500 years BP, the lake level had dropped to within 20 m of its current elevation and based on the presence of archaeological sites above +20 m from the modern lake level (Forman et al., 2014; Garcin et al., 2012), did not rise subsequently above this level. The lake level change could explain the absence of diatoms in the remainder of DMARS 1 and II (Figure 3(a)), it could reflect a preservation effect with dissolution taking place in saline water with a high pH (>9.5). Preservation of diatoms in section (diatomite sediment in DMARS 1—see lithology Figure 4), suggest that diatoms did grow in at least parts of the basin during deposition with high pH solutions that might have caused post-mortem dissolution.
Diatoms can be absent from lakes for reasons other than a high pH. For example, diatoms are relatively scarce in sediments from the northern parts of modern Lake Turkana due to high turbidity restricting light penetration into the water column and, consequently, diatom growth. The high input of sediment at Lake Turkana also has a diluting effect on microfossil abundances due to rapid clastic sedimentation which might have been the case in the Chalbi Basin. The waters of Lake Turkana are today moderately saline (~ 2500 mg L−1) and alkaline (pH~9.2; Källqvist et al., 1988) and its sediments contain calcite and high Mg-calcite, partly reflecting the presence of ostracods and chemically precipitated micrite, especially in the south basin (Halfman and Johnson, 1988; Ng’anga et al.’, 1998; Yuretich, 1986). Under these environmental conditions the plankton of the northern lake are dominated by Microcystis and Planktonema with few diatoms. In contrast, diatoms are present in the lower turbidity waters of southern Lake Turkana (Ferguson and Harbott, 1982). Given the general absence of diatoms in DMARS I and a variety of possible explanations for a lack of diatoms, the precise conditions that restricted their growth and deposition remain rather speculative.
MSBT II (560–412 cm) 4770–3780 cal BP
This period coincides with the establishment of riverine forest (Syzygium ~40% and Juniperus ~20%), high density vegetation cover which can be attributed to higher precipitation and the development of river systems as indicated by the presence of a few Acalypha, which normally grow in riverine grassland/forests (Beentje, 1994). Acalypha is common in deciduous bushland and thickets, wooded grassland, riverine grassland and on rocky shores in humid localities from sea level up to 1400 m altitude and are common in grazed areas (Beentje, 1994; Keirungi and Fabricius, 2005; van Wyk and Gericke, 2000), which could have been the case in this region. Together with Juniperus, Acalypha pollen is also reported to be well-dispersed (Hamilton, 1972; Lind and Morrison, 1974) and it is possible that the pollen is widely transported from forests (perhaps on Mount Kulal) to other areas. Over 30 species of Acalypha have been reported in East Africa, most of which occur in forest and forest edges (Radcliffe-Smith, 1987). However, Acalypha fruticosa, for example, is common in grassland and woody savannahs (Beentje, 1994; Kendall, 1969; Lind and Tallantire, 1962) and could have contributed to this record. Commiphora, although a dryland taxon, only flowers when there is high precipitation and its presence is clearly recorded in MSB II together with Euphorbia, suggesting a cooler climate interrupted by moist and dry cycles that encouraged the appearance of Juniperus and Syzygium in the highlands and Cyperaceae wetlands more locally. In this regard, it is interesting to note that sedimentary records from areas south of the sampling site attest to a significant pluvial episode after 4.4 ± 0.3 ka cal BP (Stinchcomb et al., 2026: 39).
The pollen data are also supported by diatom data with freshwater (Nitzschia amphibia, N. palea, N. linearis) taxa and an increasing presence of R. gibberula diatom indicate expansion of wetland co-occurring with saline Anomoeoneis sphaerophora, suggesting alternating wetter and drier phases, possibly on seasonal or longer time scales (Figure 2). The presence of Cyperaceae indicates moisture availability benefitting the local mycoflora (Coniochaeta cf. ligniaria, Curvularia-type, Muiruri personal observation) in the riparian vegetation. Typha, which indicates fresh water, tends to appear intermittently in low percentages towards the top of the zone, supporting diatom evidence for periodic fresher periods.
MSBT III (412–360 cm) 3780–3430 cal BP
Zones MSBIII at core depths of 412–360 cm, indicates a continuation of variable conditions distinguished by a concurrent expansion of Afromontane forest (rare Podocarpus, Juniperus, Celtis, Acalypha, Croton, Syzygium) and few woodland taxa (Commiphora, Euphorbia, Terminalia, Capparidaceae and Phyllanthus) and wetland vegetation (Cyperacae, Typha) at the expense of Poaceae and other taxa. The pollen is present in a variety of sediments that include massive muds with plant roots that probably suggest wet settings, an inference supported by the low presence of Amaranthaceae, Sansevieria, Justicia and Asteraceae. The occurrence of Salvadora and Hyphaene in the same period suggests a mesic environment close to a freshwater point or a riverine environment. Salvadora in most cases occurs along dry riverine beds and lagas and also at springs, while Hyphaene (Palmae) is recorded in seasonal watercourses (Beentje, 1994).
MSBIV (360–230 cm) 3430–2500 cal BP
The pollen present in this Zone suggests a slow establishment of dry montane forest away from the site. This possibly implies a shift to cooler and drier conditions as the more local taxa indicate an absence or low abundance of woody bushes such as, for example, Phyllanthus and Commiphora, but with the presence of herbaceous taxa (Amaranthaceae, Asteraceae, Sansevieria, Aloe). Gynadropsis and Phoenix are common and are indicators of less saline surface water conditions (Beentje, 1994), but decline towards the top of the zone, perhaps demonstrating colder and increasingly wetter conditions during deposition of this part of the sequence. Other taxa (Justicia, Tribulus, Liliaceae and Indigofera) form minor components. Acalypha forms a large component during this period with low Typha, and may also reflect intermittent short wetter phases around gently sloping water margin areas (Kendall, 1969; Kröpelin et al., 2008; Lamb et al., 2003; Temoltzin-Loranca et al., 2023a, 2023b; Wienhues et al., 2024). Intermittently, grasslands expansion is registered together with alkaline friendly Amaranthaceae when the basin becomes shallower forming a wetland environment during a drier climate. The sediments of MSBIV have high (50%) organic content, low to absent carbonate content (3%) and extremely high charcoal content of >10,000 pieces cm−3 (Figure 5).
MSBT V (230–120 cm) 2500–515 cal BP
The absence of the diagnostic Afromontane taxa Syzygium and Podocarpus (deMenocal, 1995), suggests, following Bonnefille’s arguments (Bonnefille, 1976), that forest was no longer proximal to the basin during this period. This is reflected by the low presence of Podocarpus and Olea in the landscape (Figure 2), both long-distance dispersal taxa coming most likely from the Ethiopian highlands. It is also possible that taphonomic processes, such as the fluvial transport of Afromontane taxa from the highlands, may have increased during wet intervals, which might account for at least some of these changes (Lamb et al., 2003). There is a heavier presence of Juniperus in this zone than any other zone in the entire record, which is a characteristic of dry montane forests. Stoebe is recorded and is common in high elevation bushland; co-dominant in a zone on high mountains from 2100 to 4500 m asl (Beentje, 1994), suggesting the possible source is either the Ethiopian Highlands or Mount Kulal as there are no known sites with that elevation recorded in the Chalbi Basin. The increase in Poaceae to 90% between 150 and 120 cm indicates a shift to a grassland dominated landscape from c. 1200–515 cal BP as the environment became drier. Cyperaceae, recorded in the same period (Figure 5), suggests a saline environment though in some instances a number of the Cyperaceae species do occur in savanna grasslands with other herbaceous taxa associated with drier environments as recorded by the presence of Tribulus (Lupien et al., 2018). The persistent occurrence of Cyperaceae and high concentration (up to 30 seed remains cm−3) strongly suggests local presence of Cyperaceae at the coring location, as does the predominant absence of macroscopic charcoal, with the exception of 180–200 cm (Figure 5; 2200–1860 cal BP). Gynadropsis, Phoenix, Polygala and Typha are aquatic species, together with high presence of cf. Obetia, recorded in the same period and their presence is associated with fresh marshy or swampy wetland environments or shorter precipitation periods. Increases in Amaranthaceae indicates the basin became shallower and more saline in the xeric environment. Today, there is a high presence of Amaranthaceae in the Chalbi Basin dominating the drier environment (Mohammed et al., 1996; Riollet and Bonnefille, 1976). Sediment organic content is relatively low (~10%) and carbonate content continues to be low (3%).
MSBT VI (120–0 cm) 515 cal BP–present
Pollen data show similar variability (as in other zones) with increased aquatics often correlating with increased percentages of benthic diatom taxa. The data suggest a variable hydrological environment through time with periodic increases in precipitation, possibly triggering phases of flooding. Additional evidence for drier phases is provided by the intermittent accumulation of beds of pure trona at the 100 cm interval, with trona continuing to accumulate today due to solutes that are delivered by annual floods and permanent springs. Diatoms also appear in all the levels close to the pollen changes (MSBTD III, Figure 3(a)). The taxa present include mixed saline and freshwater species that also suggest seasonally, or periodically, changing basins with mixed floras continuing throughout the zone. Increased benthic taxa (Figure 3(a)) also suggest that the waters may have been periodically shallower and/or that freshwater benthic taxa may have been introduced from streams or littoral areas into the basin. Diatoms would also grow in the fresh waters as long as nutrients were available and prior to evaporative concentration exceeding tolerance limits. Hantzschia amphioxys is an aerophilous species, which grows on moist substrates that dry out frequently, such as conditions that exist at the site today. In the present day, for example, Lake Magadi (southern Kenya), floods annually to form a thin (tens of centimetres) temporary wedge of water over trona, that evaporates within a few months to leave an extensive salt flat.
The near absence of forest taxa with a small recovery of woodland-grassland (Vachellia, Commiphora and Euphorbia) and high presence of cf. Obetia, Sansevieria, Salvadora, Hyphaene and Polygala may reflect changes in effective precipitation and possibly enhanced seasonality and could have caused this drastic change in vegetation composition—a protraction from the previous zones. Obetia are small herbs/shrubs that take advantage of wet conditions and grow on rocky outcrops and hillsides, often in hot and dry places (Beentje, 1994). Protracted rare appearances of Asteraceae, Justicia, Aloe, Phoenix and other species could be supporting similar conditions. Sediment organic content varies from high values of 50–60% and down to ~20% in this zone and carbonate content increases consistently to 5% (Figure 5). Curiously, Cyperaceae seeds (achenes) were absent and may reflect changes to the syn- or post-depositional ecosystem. Speculative mechanisms could either be changes in water flow and grazing that reduced the probability of seed accumulation in the sediments or biodegradation on and in the sediments (Mishra et al., 2016). Cyperaceae phytolith abundances, including phytoliths from achenes, have been observed to vary across the Galana Boi Formation near archaeological sites around Lake Turkana during the Mid-Holocene (Kinyanjui et al., 2026). Oribatida mite remains become more consistently common and may be associated with nearshore aquatic environments. Charcoal is consistently absent, suggesting relatively low burning (Figure 5). Low charcoal content of palustrine sediments is consistent with wetland ecosystems that cross relatively arid ecosystems with sparse vegetation and high grazing pressure on vegetation contribute to reduced fires, such as parts of Amboseli (Githumbi et al., 2018a, 2018b). The rare occurrences of remains of larvae from aquatic Diptera (Chironomidae: Chironomus and Ceratopogonidae) suggest a shallow water level at the site, but quantified estimates are not possible.
Synthesis of Chalbi Basin paleoenvironmental change
High pH and salinity measured in other Kenyan and East African lakes has been linked to extensive diatom dissolution and poor preservation (Hecky and Kilham, 1973), and it is likely that similar processes occurred at Erenderi during the Mid to Late-Holocene, leading to distinct gaps in the diatom palaeoecological record. Where diatoms do preserve, many (if not most) of the major taxa identified in Erenderi are also very common in saline and alkaline lakes throughout East Africa, including Kenya (Cocquyt and De Wever, 2002; Hecky and Kilham, 1973; Owen et al., 2004). These and other studies have been drawn on to help interpret the Erenderi record.
It is clear from the diatom stratigraphy that conditions have generally been unfavourable for diatom preservation for much of the Mid to Late-Holocene, except perhaps for the past 500 years. It is not possible to say definitively why this is the case, but one reason may be that salinity and pH of the wetland has simply been too high for diatom preservation to occur. This may suggest therefore that where there is occasional diatom preservation, perhaps salinity and pH were not as high, linked to, for example, the prevalence of wetter conditions in and around the wetland.
Most of the species identified are benthic, often periphytic (e.g. Rhopalodia gibberula growing on other plants and algae,) or epipelic (e.g. Anomoeoneis sphaerophora, living on mud and sediment surfaces) (Cocquyt and De Wever, 2002; Gasse, 1986). Only Stephanocyclus meneghinianus is planktonic, with optimum growth in open water rather than in littoral habitats. Species including Craticula elkab, R. gibberula, A. sphaerophora, S. meneghinianus and N. frustulum all have very high optima for lake water conductivity, certainly thousands mS/cm (Cocquyt and De Wever, 2002; Gasse, 1986), highlighting that they grow well in aquatic ecosystems with high salinities. For example, in a study of diatoms in wetlands around the Bogoria–Baringo region of Kenya, Owen et al. (2004) found A. sphaerophora and some of its varieties abundant in both hot, spring-fed, very shallow mashy wetlands and in the littoral regions of hypersaline lakes with generally fewer plant macrophytes. R. gibberula on the other hand tended to occur in river and groundwater-fed wetlands with abundant plant macrophytes, alongside N. palea, N. amphibia (Owen et al., 2004) diatom species that grow best in brackish rather than saline ecosystems (Cocquyt and De Wever, 2002; Gasse, 1986). Moreover, it should be noted that R. gibberula is often indicative of nitrogen-poor conditions because it contains N-fixing endosymbiotic bacteria that are able to fix atmospheric nitrogen.
The diatom stratigraphy at Erenderi suggests that the wetland gradually changed through the Mid to Late-Holocene, from perhaps one which was nutrient-rich, hypersaline and very alkaline and with few aquatic macrophytes, to one which became less saline with greater plant macrophyte diversity but was at the same time increasingly nitrogen limited (Figure 3). There is clearly a change in conditions around 500 cal BP allowing better preservation of diatoms. The appearance of the planktonic S. meneghinianus is indicative of deeper water persisting at the site, at least for a short time. The appearance of N. amphibia suggests a further freshening of the Erenderi ecosystem (Gasse, 1986) and potential increased supply of nitrogen, as it is a facultatively nitrogen-heterotrophic taxon (Mertens et al., 2025). Today, the Chalbi Basin is one of the most arid regions of eastern Africa and the saline soil of the desert allows only a few plants to survive, so most of the Chalbi is quite bare of vegetation (Figure 1). Occasionally, oryx, ostrich and zebra can be seen out on the mud, and local Gabbra herders lead their camels across it between watering holes and grazing lands on the desert margins. Apart from this, the Chalbi remains sparsely and extensively occupied as it is one of the least hospitable regions of Kenya. In the past, however, this was not always the case. The evidence suggests that conditions have been moister, and that this area potentially was able to support a larger population of humans and higher plant and animal biomass. This evidence falls into a number of categories: geomorphological (landforms), geological (sediments), zoological (animal remains), botanical (plant remains) and archaeological (remains of human settlements and artefacts). The record herein (plant pollen remains) provides new insight into past ecosystem responses to climate change and fluctuation in the Chalbi Basin over nearly five millennia of increasing regional aridity (Cohen et al., 2016; Bloszies et al., 2015). Pollen, diatoms and charcoal are common, possibly because the geomorphology of the site comprises alluvial fans formed by intermittent flowing water in a laga, with sediments being deposited at a point where the slope of the laga bed decreases. Such fans frequently form along the margins of inland drainage basins that may have been nearly continuously wet leading to bog formation, which enhanced organic preservation. The record shows dominance of Cyperaceae and Amaranthaceae, from ~4650 marking a period when the lake basin was becoming shallower and the water more saline/alkaline due to extreme aridity. This pulse of quite rapid aridity around 4000 years BP has been extensively recorded across East Africa (Marchant and Hooghiemstra, 2004). There then followed a shift to more mesic conditions when Afromontane forest expanded and the site became more riverine until around 2800 BP. This was followed by the onset of cooler and drier conditions that continued through the Late-Holocene.
The charcoal record had the highest sieved charcoal concentration values, known to the authors, for eastern Africa. The three peaks with different amplitudes are very distinct with extremely low charcoal concentrations in between. Peak concentrations may arise from high charcoal influx or intervals of decreased sediment deposition and it is not possible to disentangle these mechanisms with the data presented. The number and depths of radiocarbon dates and non-contiguous analyses of pollen and diatom microfossils limit the interpretation and causes of the charcoal increases. The charcoal peaks and tails may represent time intervals of more local fires and charcoal accumulation and local fire-free or fire-reduced intervals that deposited much less charcoal in the record (Figure 5). Other sediment charcoal records from eastern Africa lowlands, such as Esambu in Amboseli, southern Kenya (Githumbi et al., 2018b), have only one major peak in charcoal during the Late-Holocene. Sediment cores from wetlands in highlands of eastern Africa also have discrete single peaks in charcoal, such as Maua on Kilimanjaro (Courtney-Mustaphi et al., 2021) and Kwasebuge in South Pare (Finch et al., 2017), both in Tanzania.
Correlating these peaks with possible phases of intensification of human activity is challenging, owing to the limited number of radiocarbon dates bracketing each peak, and, as importantly, owing to the limited extent of research undertaken on the Mid to Late-Holocene archaeology of the Chalbi Basin. From the work that has been undertaken around the Basin and from the more extensive sample of sites investigated along the eastern shores of Lake Turkana (and to some extent also referencing evidence from the south-western side), a basic synopsis of the likely nature of settlement around Lake Chalbi and associated livelihood strategies can be attempted.
Faunal remains recovered from sites close to the lake indicate these LSA populations exploited a combination of terrestrial animals (typically medium to small bovids such as oryx, wildebeest, Grant’s gazelle and warthog), fish and, perhaps less frequently, aquatic mammals such as crocodile and hippopotamus (Stewart, 1989: 160–169). Fishing strategies appear to have been opportunistic in nature with an emphasis on Nile perch and the larger cichlids, and to a much lesser extent mudfish (Stewart, 1989: 227–235). Barbed harpoons and points made on bone have been recovered from several sites, including Lothagam (Robbins, 1980) and Lowasera (Phillipson, 1977), as well as some of the earliest evidence in the region for the manufacture and use of ceramics decorated with distinctive wavy line motifs (Keding, 2017).
With the onset of drier conditions towards the end of the early Holocene, reduced rainfall resulted in a fall in lake levels across the region (Garcin et al., 2009, 2012; Kinyanjui et al., 2026; Stager and Johnson, 2000). This period also witnessed the introduction of the first domesticated animals into northern Kenya and the beginning of food production, now dated to ~5000–4800 cal. BP. The lowest pollen zone of the Erenderi core, MSBT I (~5200–4770 cal. BP) thus overlaps this period of transition. Pioneer PN herding communities may have settled around Lake Chalbi, as hinted at by the appearance of diagnostic PN ceramics and stone bowls at North Horr, perhaps also interacting with autochthonous LSA hunting-fishing-foraging communities. Evidence from elsewhere in Marsabit County, notably Ele Bor (Gifford-Gonzalez, 2003) close to the modern border with Ethiopia and Kulchurdo on Mt Marsabit (Phillipson and Gifford, 1981), confirms the continued presence of forager communities in the area well into the Late-Holocene.
The density of stone burial cairns, several with broken stone bowls on their surfaces, around the edges of the Chalbi Desert also suggests the landscape began to assume increasing ritual importance for PN communities from ~4100 to 3380 cal. BP, based on the dating of Cairn 4 on Kokurmatakore Hill, Kalacha, although as this is a date on apatite it should be treated with caution until more securely dated samples are available. If correct, this phase of PN expansion may well have corresponded with pollen zone MSBT II (~4770–3780 cal. BP), a period that experienced higher precipitation than in the immediately previous phase and the development of river systems. By MSBT III (~3780–3430 cal. BP), pastoralism was likely well established around the margins of the Chalbi. At present aside from the deflated sites at North Horr and various undated surface traces elsewhere, there is no securely dated archaeological evidence for this, however. With the onset of cooler and drier conditions and grassland expansion during MSBT IV (~3430–2520 BP), wetland sites such as Erenderi are likely to have been particular foci for settlement, although again the archaeological evidence for this is currently very limited.
The sites on Kokurmatakore Hill attest to new episodes of cairn construction during the latter part of MSTBT V (~2520–515 cal. BP), represented by at least three different forms of burial cairn. The two platform cairns (GdJn 1 and 2) excavated by Stiles and Munro-Hay (1981) may be the earliest of these, although the range of the single available date (from GdJn2) overlaps with some of the available dates for the different types of ring cairn, which are far more numerous in the wider landscape and continued in fashion until the late second millennium CE. Examples of platform cairns similar in form to the two near Kalacha have been noted in southern Ethiopia, and it is possible, as Stiles and Munro-Hay (1981) inferred, that their presence here is associated with initial southward expansion of Southern Oromo speakers around a 1000 years ago. Given the relatively recent dates of the excavated ring cairns, it is conceivable that their presence may mark the appearance of the ancestors of current Gabra and/or Rendille populations. However, other affiliations are possible and, as noted above, far, more Holocene archaeological sites bordering the Chalbi Desert need to be investigated and dated to better understand the trajectory of human-environment interactions over the circa 5000 years of ecological history captured in the Erenderi core.
Conclusions
The new paleoenvironmental record from Erenderi (core Eren4C), Chalbi Basin, Marsabit County, Kenya, preserved pollen and diatom microfossils and charcoal, plant and aquatic invertebrate macroremains, that when combined provide an opportunity to reconstruct patterns of environmental change in this locality from the Mid-Holocene to present. Previous studies have characterised the African Humid Period and subsequent regional aridification. The Erenderi paleoenvironmental records provides a more local-scale pattern of wetland dynamics with variations in the diatom assemblages. During the Mid-Holocene, herding economies were first introduced, supplementing pre-existing hunting, fishing and foraging economies, leading ultimately to the current predominant land uses of today. The Late-Holocene was characterised by a changing wetland ecosystem characterised by terrestrial carbon inputs and potentially intermittent localised burning. From 3300 to 2650 cal. yr BP, the coring site was locally overgrown by Cyperaceae and was likely ephemerally inundated and from 2650 to present shallow water with potential for episodic drying conditions, similar to present time, has prevailed, highlighting the importance of Erenderi to people, plants and animals as a source of surficial and near-surface water. The results suggest that under the regional-scale pattern of aridification, the spatially intermittent wetland ecosystems of the Chalbi Basin were complex with local variability in inundation, water quality, vegetation and fire occurrences.
Footnotes
Acknowledgements
We wish to acknowledge the institutions and individuals who have supported us in one way or the other during all the stages of this research. We thank the GCRF through the British Academy for generously funding this research, the people of northern Kenya who fully participated in this research and showed us a lot of enthusiasm and support throughout the fieldwork. We also wish to thank the National Museums of Kenya, Department of Earth Sciences, Palynology Section for the provision of coring equipment, office and laboratory space and facilities to complete the work. We thank the Marsabit team, Ali and Katelo, for their participation in the field work activities; Alex, for his efforts and involvement in our movement to different places searching for wetland sites; Nik Petek-Sargeant for logistics; Tomasz Goslar at the Poznań Radiocarbon Laboratory; and Pierre Lapellegerie and Lucia Wick for macrofossil identification discussions. The British Institute in Eastern Africa is also acknowledged for providing field equipment and logistical support. We thank anonymous peer reviewers and Anson Mackay whose suggestions improved this study.
ORCID iDs
Author contribution(s)
Conceptualization, PJL, FNM, VM; methodology, VM, RM, CCM; field research: VM, AM; formal analysis, VM, CCM, AM, RM; data curation, VM; writing—original draft preparation, VM; writing—review and editing, RM, CCM, PJL, FNM & VM; visualization, VM, CCM, PJL; project administration, FNM, PJL; funding acquisition, FNM, PJL.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research on which this article is based was funded by the British Academy’s Sustainable Development Programme 2018, supported under the Global Challenges Research Fund – Well-Being: Indigenous Wells, Pastoralist Biocultural Heritage and Community Archaeology for Sustainable Development in Northern Kenya and Southern Ethiopia, PI Freda M’Mbogori, CoI Paul J Lane, Grant No. SDP2\100167.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.