
Abstract
The northern sector of the Western Pampean Dunefield (central Argentina), characterized by Late Quaternary to Late Holocene aeolian morphogenesis and sedimentation under drier conditions than present, hosts numerous shallow lakes developed in the deflation basins of blowout dunes. A multi-proxy analysis, including magnetic susceptibility, charcoal, C and N content, stable isotopes, and pollen and non-pollen palynomorphs, was performed on a sediment core from Laguna Pocitos (33° 58.2’S – 65° 34.3’W) to contribute to paleoenvironmental and paleoecological reconstructions of the region. The integrated evidence allows inferences about environmental variability over the past ~350 years (~1670–2015 CE), including the impact of natural and anthropogenic forcings. The first recorded period (~1670–1758 CE) reveals rising water levels under drier conditions than today, with intense fire events within a grassland-dominated landscape. Frequent water level fluctuations were inferred for the next 100 years (~1760–1870 CE), accompanied by diminished grassland cover and more disturbance taxa associated with an increasing anthropogenic presence in the area. After ~1870 CE, a trend toward lower water levels coincides with widespread droughts that culminate in the historically and geologically recognized Pampas Dust Bowl. Since ~1970 CE, multiple core indicators evidence a lake expansion, consistent with remote sensing data and instrumental records. This phase was characterized by clear water conditions synchronous with an intensification of anthropogenic pressure. Overall, hydroclimatic variability inferred from the Laguna Pocitos record is coherent with historical documents and paleolimnological reconstructions from the Pampa plain, revealing the control of regional climate pattern but including distinctive local modulation which needs further studies.
Keywords
Introduction
Paleolimnological studies on shallow lakes are essential for reconstructing the ecological and climatic conditions in continental settings during the Late Quaternary (Birks and Birks, 1980, 2006) and for assessing natural or anthropogenic landscape changes (e.g. Perez et al., 2021). These reconstructions help understand past Earth system dynamics and inform predictions under ongoing global change (Lee et al., 2021; Steffen et al., 2006). Paleolimnology integrates physical, biological, and chemical indicators preserved in lake sediments (Adrian et al., 2009). In southern South America, the Late Quaternary evolution of the Pampa region, considered the “breadbasket” for Argentina (Baldi and Paruelo, 2008), was mainly reconstructed through aeolian and lacustrine records (Cuña-Rodríguez et al., 2020; Tripaldi et al., 2013; Zárate, 2003 and references therein). In particular, this temperate plain hosts numerous shallow lakes, under varied morpho-structural setting and a climatic gradient in terms of precipitation, which provided insight into the paleoenvironmental and paleoclimatic conditions during the past ~25 ka, especially during the last millennia (Piovano et al., 2025; and references therein).
Paleolimnological reconstructions can be complemented with surface sediment analysis that represent modern conditions in the lake’s catchment area (Li et al., 2022). In the Pampa plain, this approach has been explored through the spatial distribution of pollen (e.g. Frazer et al., 2020; Tonello and Prieto, 2008). Spatial analysis of stable isotopes and major elements on surface sediments provides information about organic matter sources and the processes shaping their isotopic signatures across the landscape (Guo et al., 2020). Although few studies combine surface sediment isotopes with paleolimnological reconstructions (e.g. Cañada-Pasadas et al., 2024; Li et al., 2016), this integration offers an essential calibration framework to better interpret past environmental variability.
In the Western Pampa (~ 38.8° S – 34.5° S, 65.3° W – 65.7° W, San Luis Province, Figure 1), there is an unusual concentration of shallow lakes considering the semiarid-subhumid conditions of this region. This dryland is characterized by a vast (~47,000 km2) cover of stabilized sand dunes (namely Western Pampean Dunefield -WPD- after Zárate and Tripaldi, 2012), that inhibited the development of integrated drainage networks, while the wind processes created numerous deflation basins related to blowout dunes (Tripaldi and Forman, 2025). Between ~33.8°S–34.7°S and ~65.3°W–65.7°W, the WPD presents numerous shallow lakes (an area informally named WPSL- Western Pampean Shallow Lakes), developed due to the combination of an average positive change in the hydrological balance during the past millennium and the presence of deflation basins related to blowout dunes (Tripaldi et al., 2025). The role of anthropogenic impact in this lake formation is still under study, though to the north of this area, the extensive replacement of native woodland to cropland, during the past five decades, reduced evapotranspiration, raised groundwater levels and triggered the appearance of new rivers (Houspanossian et al., 2023; Jobbágy et al., 2021; Santoni et al., 2010).
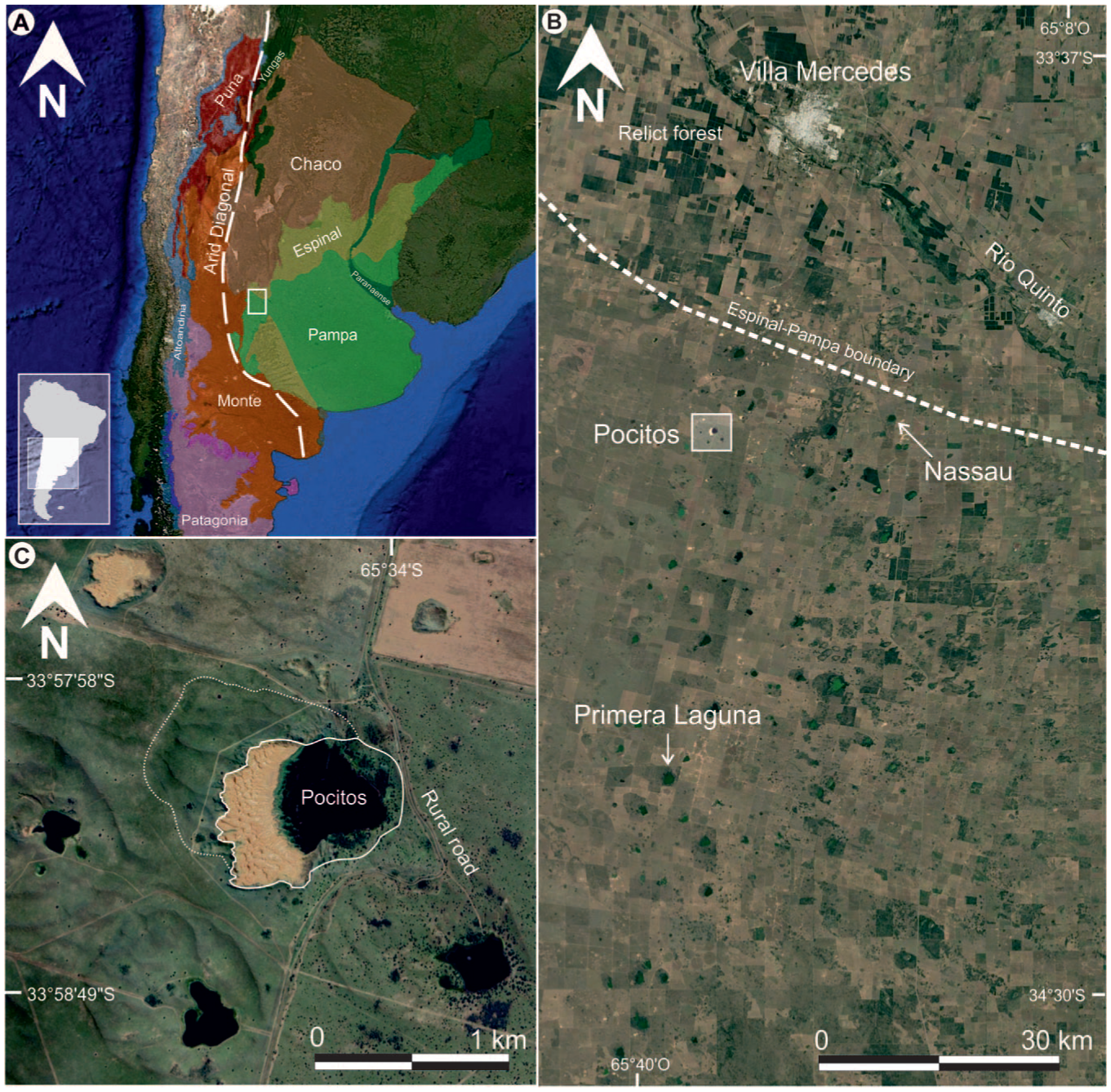
(a) Location of the study area (rectangle) in the western Pampa plain of Argentina, placed at an ecotonal region between the arid–semi-arid Monte to the west and the humid–sub-humid Pampa to the east (phytogeographic provinces after Oyarzabal et al., 2018). The dashed line represents the 250 mm isohyet corresponding to the South American arid diagonal, (b) shallow lake area within the western Pampean dunefield of the Western Pampa, and (c) Laguna Pocitos developed within the deflation basin associated with blowout dunes.
Previous studies in the northern WPD documented a dominant Late Pleistocene–Holocene aeolian sedimentation, accompanied by pedogenic/stabilization phases over the past ~50 ka (Tripaldi and Forman, 2016). Aeolian deposits also contain ~14.7–11.6 Ka old palustrine and lacustrine facies, associated with the blowout dunes deflation basins (Tripaldi et al., 2025). More recent paleolimnological research in the area exposed the growing and evolution of two shallow lakes (namely Laguna Nassau and Primera Laguna, Figure 1b) over the past ~1,400 years (Vilanova et al., 2015, 2022).
With the aims of increasing the number of paleolimnological archives for reconstructing the WPSL hydroclimatic variability, with a particular focus on analyzing the anthropogenic impact over this landscape during the past century, we present a multiproxy study of Laguna Pocitos, located at 33° 58.2’S – 65° 34.3’W (Figure 1). Paleoenvironmental and paleohydrological inferences were made based on the lake sedimentary and pollen record, to then discuss the local controls and compare with regional historical information. A previous regional synthesis of the WPSL shown that Laguna Pocitos followed the general hydroclimatic trends of the Western Pampa record (Maciel et al., 2025). However, the specific role of local geomorphology, sediment dynamics, vegetation, organic matter sources and land-use history on the evolution of this shallow lake remained unexplored. The objective of this research was to identify the role of local controls on the paleoenvironmental and palaeohydrological evolution of Laguna Pocitos during the last ~350 years through a high-resolution multiproxy approach, and to evaluate how these local processes modulated the regional hydroclimatic signal previously recognized for the Western Pampa. Additionally, we focus on Laguna Pocitos as being a more important node in the “rastrilladas” than Laguna Nassau and Primera Laguna. Its use as a stop on the indigenous roads in the 18th and 19th centuries makes the Laguna Pocitos record particularly valuable for exploring human impact before land-use changed to modern agriculture and cattle raising.
Study area
Geomorphology and hydrogeology
The study area lies in the San Luis Province, central-western Argentina, southern South America (Figure 1a). It is included in the Western Pampean Dunefields (WPD, sensu Tripaldi et al., 2025; Zárate and Tripaldi, 2012), part of the Pampean Sand Sea (Iriondo and Kröhling, 1995). The WPD is largely stabilized and covered by native vegetation or cultivated fields, with some patches of active dunes and aeolian sand mantles, and mainly formed by blowout dunes, accompanied by sinuous ridges, aeolian streaks and aeolian sand sheets (Tripaldi and Forman, 2025). These blowout dunes, ranging from 200 to 4000 m in length and 120–2500 m in width, are crater-shaped landforms surrounded by sandy rims and deflation basins over 30 m in depth (Tripaldi and Forman, 2025). The eastern flank of the WPD hosts numerous shallow lakes, like the here studied Laguna Pocitos (Figure 1b and c; Tripaldi et al., 2025). The regional slope in the study area is <1° toward the SE and lacks a well-integrated drainage network, except for the Quinto River, ~35 km north of Laguna Pocitos which drains the Sierras de San Luis.
Sedimentological and luminescence data revealed persistent aggradation interspersed with pedogenic and palustrine phases since, at least, the Late Pleistocene (Tripaldi and Forman, 2016, and references therein). Particularly, increased water availability and the formation of shallow lakes and wetlands marked the Pleistocene-Holocene transition (Tripaldi et al., 2024), while aeolian activity declined significatively over the last millenium (Tripaldi and Forman, 2016), coinciding with renewed shallow lake development (Maciel et al., 2025, and references therein). During the past ~100 years, the WPSL underwent recurrent flooding and desiccation events also recorded in the Pampean plain, linked to both conditions that were related to climatic and anthropogenic factors (Bogino and Jobbágy, 2011; Tripaldi et al., 2013; Viglizzo and Frank, 2006).
Climate
The Western Pampa exhibits interannual to multidecadal climate variability (Compagnucci et al., 2002), oscillating between semi-arid and subhumid conditions due to its proximity to the Arid Diagonal (Bruniard, 1982; Figure 1a), which marks the western limit of the South American Monsoon System (SAMS) influence (Barros et al., 2002; de Carvalho and Jones, 2016; Garreaud et al., 2009). This transitional zone separates arid–semi-arid climates to the west and humid–sub-humid to the east (Garreaud et al., 2009; Vera et al., 2006). The studied area is a temperate dryland with mean annual mean temperature of 15.4°C, daily thermal amplitude exceeding 15°C (Gentile et al., 2020) and mean annual rainfall of 608 mm (period 1903–2024 at Villa Mercedes weather station, Figure 1), about ~70% of which occurs during spring and summer (Figure 2a) under SAMS influence.
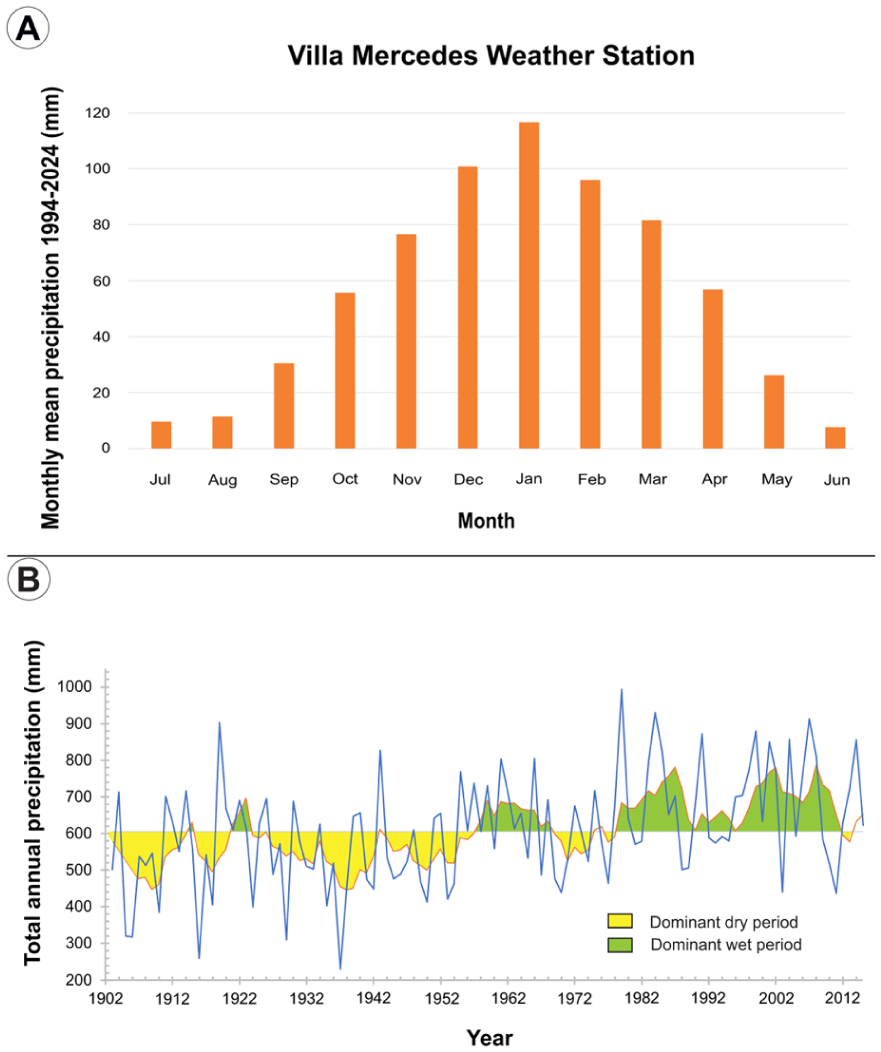
(a) Monthly mean precipitation (from Villa Mercedes INTA weather station; period 1994–2024) and (b) annual precipitation trend during the period 1903–2015 represented by the annual yearly rainfall (solid blue line) and 5 year moving average (orange line) from Villa Mercedes weather station, exposing the predominantly dry and wet periods in the study area during the instrumental record.
Instrumental data from the Villa Mercedes weather station shows strong variability, ranging from 230 to 990 mm for the 1903–2015 period. A dry phase persisted until the 1950s, with the 1930s as the driest decade (mean 515 mm, 230 mm minimum in 1937; Figure 2b). Since 1976/1977, summer rainfall increase was recorded in central Argentina (Agosta and Compagnucci, 2008), also recorded in the study area. The wettest decade at Villa Mercedes was the 2000s (mean 720 mm; Figure 2b).
Vegetation
The vegetation around Laguna Pocitos represents an ecotone between the Pampa grasslands and the Espinal xerophytic forest, shaped by a regional precipitation gradient (Figure 1a). The area lies on the dry western edge of the Pampa Phytogeographic Province (Cabrera, 1976; Oyarzabal et al., 2018) within the Western Interior subregion (León and Anderson, 1983), characterized by steppe and pseudo-steppe grasslands and shrublands. Native grasses such as Sorghastrum pellitum and Elionurus muticus dominate, with species diversity influenced by edaphic factors (e.g. texture, pH) and grazing intensity. Due to limited agricultural development, well-preserved grasslands remain, exhibiting lower disturbance compared to other regions (Baldi and Paruelo, 2008; Demaría et al., 2008).
The grasslands are surrounded by xerophytic vegetation (Espinal and Monte phytogeographic provinces, Figure 1a). Xerophytic woodlands of the Espinal Phytogeographic province, dominated by Neltuma (formerly Prosopis) caldenia, appear to the north and to the south, while to the northwest, the Chaco province features Schinopsis marginata-dominated xerophytic forests (Oyarzabal et al., 2018). These open, deciduous forests thrive where groundwater is accessible (Oyarzabal et al., 2018). Westward lies the Monte Phytogeographic province, a xerophytic shrub steppe dominated by Zygophyllaceae and columnar Cacti (Oyarzabal et al., 2018, 2025).
A shallow water table encourages woody species in low-lying areas, with clusters of Geoffroea decorticans and isolated specimens of N. caldenia, N. alpataco, and Celtis ehrenbergiana (Zuloaga et al., 2019). Since the early 20th century, woody species have expanded, where Espinal and grassland vegetation are mixing in an ecotone (León and Anderson, 1983). Scattered N. caldenia specimens are notable in grasslands, particularly near shallow lakes.
Land-use
Land cover and land-use are fundamental drivers of change in shallow lakes; since the mid-20th century, global land-use transformations have caused both the creation and degradation of numerous lakes and ponds (Brönmark and Hansson, 2002; Meerhoff and Beklioğlu, 2024). The natural Pampean grassland was not the exception: today the region functions as an agricultural anthrome, where intensification of human activity over the past five decades has profoundly altered surface processes and hydrology (Bohn et al., 2016; Ricard et al., 2020).
The dune-lake landscape of the Western Pampa has long been sparsely populated but inhabited by hunter-gatherers since, at least, the Middle Holocene (Heider, 2015; Heider and Rivero, 2018). Despite scarce surface water, evidence of human activity is widespread. These groups displayed high mobility, indicated by the use of lithic resources from diverse regions, including the Central Sierras, the Pampa, and the Andes (Heider et al., 2024). The unvegetated areas of the blowout dunes were particularly attractive for their shallow, good-quality water tables, feeding small, biodiverse water bodies that function as eco-shelters (Heider et al., 2019).
Over the past three centuries, Pampean societies experienced significant cultural and social transformations (Curtoni, 2007; De Jong, 2007, and references therein). The introduction of European livestock, particularly horses and cattle, redefined the social, symbolic, and economic landscape. Interactions with the Argentine state reshaped land-use, especially in the dry Western Pampa, with the development of a network of indigenous roads, known as “rastrilladas” (D’Hiriart and Mollo, 2018). Historical documents and maps from the 18th and 19th centuries highlight the role of water bodies as spatial references for the movement of people and animals, and cultural interactions (Curtoni et al., 2022). During the late 19th century, the so-called “Conquest of the Desert” (i.e. historical and political denomination referring to the “civilization” -or extermination- of native populations) marked a turning point, integrating the region into small-scale agriculture and livestock production that expanded around railway stations by the early 20th century (Tello Cornejo, 2003).
For the past century, native grasslands have been grazed at varying intensities, replaced by Medicago sativa (“alfalfa”) and other drought-tolerant exotic species (Demaría et al., 2008; Viglizzo and Frank, 2006). Livestock intensification led to irreversible biodiversity loss, affecting vulnerable species like Sorghastrum pellitum (Aguilera et al., 1999). Degraded grasslands became prone to the invasion of Geoffroea decorticans clusters (León and Anderson, 1983). Meanwhile, forests and grasslands cover decreased starting in the 1970s, replaced by croplands (Demaría et al., 2008), further facilitating the invasion of exotic species already established in other Pampa regions (Aguilera et al., 1999).
Since the 1980s, higher rainfalls, improved tillage and the use of transgenic crops and agrochemicals, triggered agricultural intensification in Western Pampa (Viglizzo et al., 2011). In the northwestern sector of the WPD, and including the WPSL, remote sensing analyses quantified that the area covered by shallow lakes nearly doubled between 1985 and 2001, while natural grasslands were transformed into cropland or pastures (Demaría et al., 2008).
Laguna Pocitos
Laguna Pocitos (33° 58.2′ S, 65° 34.3’ W, and 485 m asl) is located within one of the numerous deflation basins of the blowout dunes that characterize the WPD (Figure 1b). In particular, at Laguna Pocitos, two large blowout dunes, with different orientations, are superimposed. There is a 1.25 km long dune with a water body to the E-SE and a vegetated depositional lobe, of up to 10 m above the regional plain, oriented to the W-NW (dotted white line in Figure 1c). The reactivation of this blowout dune made the deflation basin deeper and re-shaped a west-oriented depositional lobe (white line in Figure 1c), which is covered by fine-very fine sand with active barchanoid dunes (Figure 3a–c). This younger blowout dune presents a SW-NE asymmetrical relief, with a depositional lobe rising to ~25 m on the western margin and the deflation basin to the east where the Laguna Pocitos was developed (Figure 3e).
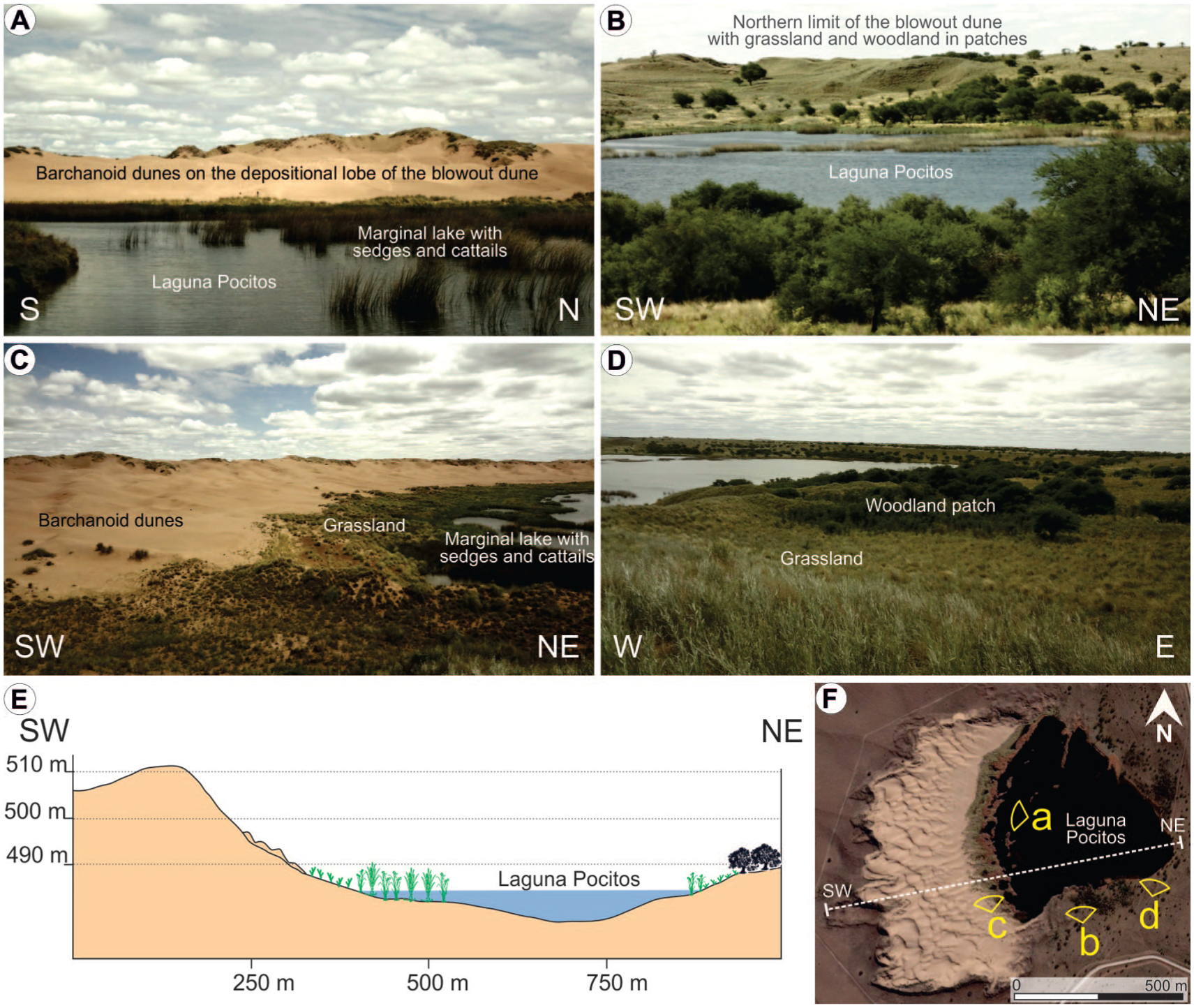
(a–d) Pictures of Laguna Pocitos area with the western sector covered by active dunes, shallow littorals dominated by marsh plants, and grasslands with patches of Caldén woodlands in the surrounding margins, (e) W–E topographic profile of the blowout dune that created the deflation basin that hosts Laguna Pocitos, and (f) the satellite image of Laguna Pocitos indicates the line of the topographic profile and the zones illustrated in the pictures.
The floor of the lake has a shallower slope to the west, where dense emergent hydrophytic vegetation (such as cattails and sedges) is dominant (Figure 3). The northern, eastern and southern margins of Laguna Pocitos are characterized by grasslands and xerophytic woodlands in patches (Figure 3).
In 2015, Laguna Pocitos presented a diameter of about 500 m, with 0.23 km2, an average water-depth of 2 m and a maximum depth of 8 m (Figure 3f). The expansion of Laguna Pocitos was not exception regarding to the regional volume/surface increase of the shallow lakes in the WPD during the past decades, expanding from 0.04 to 0.2 km2 between 1985 and 2022.
Hydro-geochemically, the Laguna Pocitos water was classified as alkaline (mean pH of 9.1) and subsaline (mean TDS of 651 mg/l), with an HCO3--Na+-K+ composition (Echegoyen et al., 2022). Stable isotope analyses indicate continuous inflow and outflow of groundwater, aligned with the regional flow running NW–SE (Vanina Echegoyen et al., 2021).
Regarding land-use, the primary disturbance on the Laguna Pocitos comes from nearby agriculture and livestock, with cattle accessing the lake.
Materials and methods
Paleolimnological data was obtained from a 50 cm-long sediment core (PO15-1) collected in May 2015 (austral autumn) at a depth of 4.1 m by using a manual Eijkelkamp sampler. Lithology was visually described based on texture, sedimentary features, and color (Munsell Soil Color Chart, 2000). Volumetric magnetic susceptibility, expressed in SI units (SI 10-5), was measured with a Bartington MS2E sensor every 0.5 cm in one of the core’s halves. Macroscopic charcoal analysis was conducted with continuous and contiguous sampling every 1 cm, following the methods described in Whitlock et al. (2003). Charcoal concentrations (number of particles cm−3) were converted into charcoal accumulation rates (particles cm−2.yr−1) by using Char-Analysis software (Higuera, 2009), resampling charcoal concentrations and interpolating the record years to the median sample resolution (7 years), spanning from −65 to 264 years cal BP.
For pollen and non-pollen palynomorph extraction (NPP), 1 cm3 subsamples (with the same sampling intervals as charcoal) were filtered through 150 μm mesh sieves, followed by physical-chemical treatments detailed in Faegri and Iversen (1989). A minimum of 300 pollen grains were counted per sample. Pollen percentages were calculated in relation to the total terrestrial pollen sum, excluding marsh plants (e.g. Typha, Cyperaceae) and aquatic plants (e.g. Myriophyllum) due to their local signal. Percentages from these plants and algal remains were calculated based on the total palynomorph sum. Pollen zones were quantitatively determined by the multivariate method CONISS (stratigraphically restricted cluster analysis) considering only terrestrial taxa (Grimm, 1987). Palynological diagrams and dendrograms were plotted using TGView (Grimm, 2004).
Stable isotope analysis of C and N (δ13C and δ15N ‰) and organic C and N contents (wt %) were carried out with a Thermo-Electron dual-inlet mass spectrometer at Baylor University. Sampling in the sediment core was performed every 2–3 cm. Samples were pretreated with acid to remove calcium carbonate followed by rinsing with distilled water.
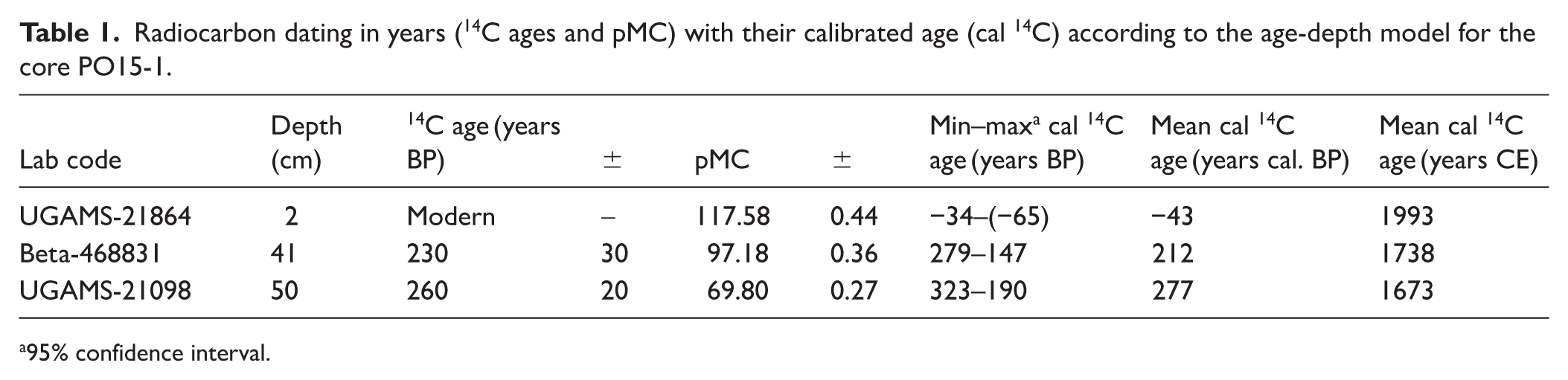
The chronology for the PO15-1 core was developed using three radiocarbon ages from the organic fraction of the sediments. The Accelerator Mass Spectrometry (AMS) technique was employed. Two of the ages were analyzed at the Center for Applied Isotope Studies (University of Georgia), and the third was determined at Beta Analytic Inc. Age calibrations were done using the SHCal20 (Hogg et al., 2020) and post-bomb SH1-2 (Hua et al., 2022) curves and age-depth modeling was performed using the R package Bacon v. 2.5.5 (Blaauw and Christen, 2011).
Also in May 2015, 14 surface sediment samples (top 5 cm) were collected from different biogeomorphological units representing the surface isotopic and major elements conditions. The sample sites were selected based on their geomorphology, and vegetation composition and cover percentage. Treatment and analysis of these samples was the same as previously described for the PO15-1 core samples. To compare the isotopic content of the surface samples, both between different units and within each one, statistical tests (ANOVA, Kruskal-Wallis) were applied by using the base R package v. 4.4.1 (R Core Team, 2024). The same package was also employed to assess the linear relationship between each pair of isotopic variables, both in surface and sub-fossil samples, through Pearson’s correlation analysis.
Results
Chronology
The age-depth model was developed from three radiocarbon dates and including the date of core extraction. No significant gaps were assumed as no sedimentary discontinuities were recognized in the sequence. The average accumulation rates were set to 7 year/cm for a better fit of the two lower/older dates. The memory (dependence of accumulation rate between neighboring depths; Blaauw and Christen, 2011), was set to 0.3. This allowed for greater variability in accumulation rates, providing a better fit for the upper/newer date. According to the average values of the model, core PO15-1 covers the period from 277 to −65 cal BP (1673–2015 CE; Table 1, Figure 4a).
Radiocarbon dating in years (14C ages and pMC) with their calibrated age (cal 14C) according to the age-depth model for the core PO15-1.
95% confidence interval.
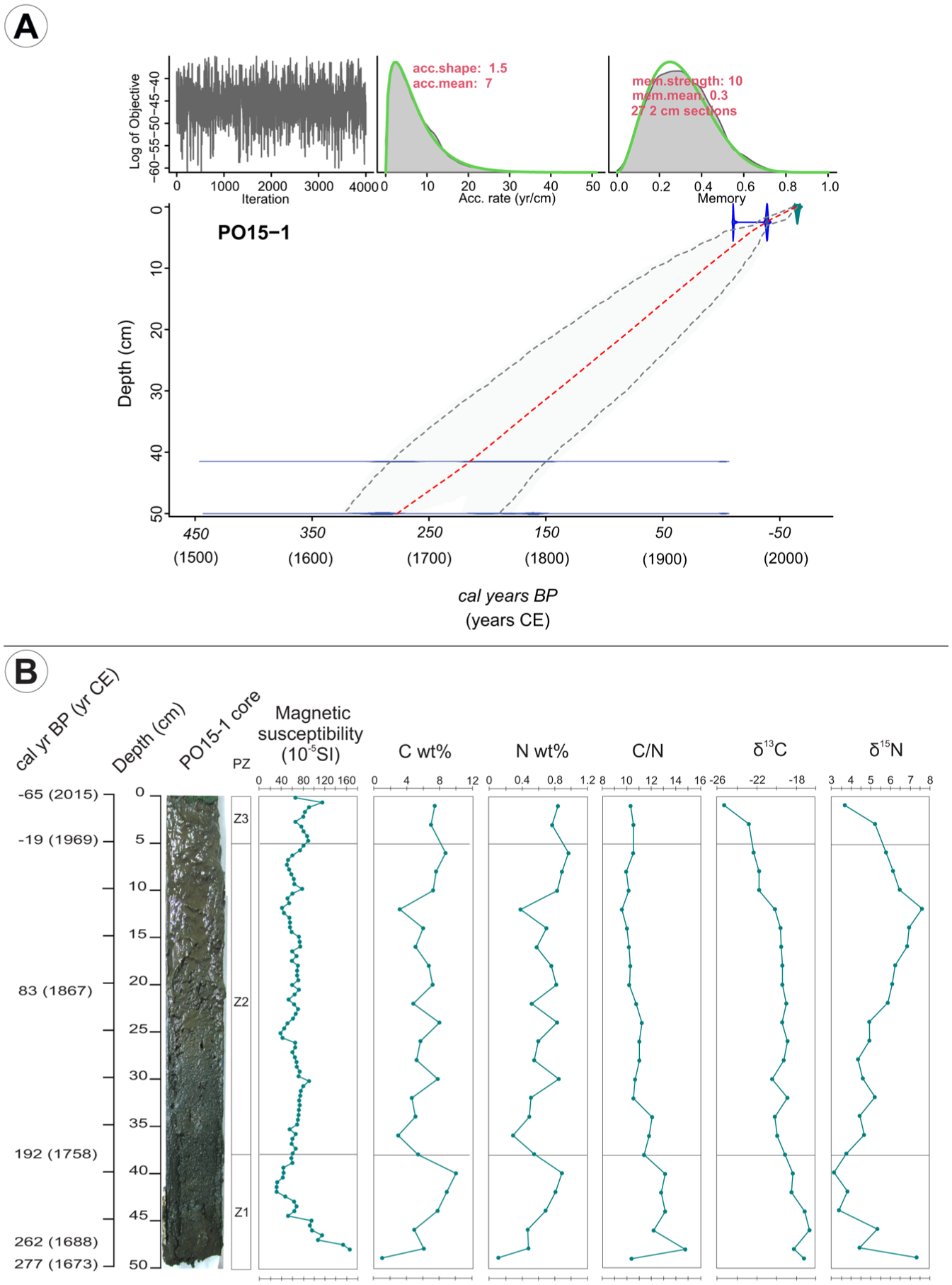
(a) Age model developed for core PO15-1. Red line: mean calibrated age, (b) photograph of PO15-1 core from Laguna Pocitos along with the palynological zones (PZ, see Figure 5) and the comparison of variations in magnetic susceptibility (10-5 SI), content of (c) and N (wt %), C/N ratio, δ13C (‰) and δ15N (‰) stable isotopes ratios.
Lithology and magnetic susceptibility
The PO15-1 core consists of 50 cm of massive, organic, fine to very fine sandy silt. The lowermost section (~50–46 cm) shows a slightly higher sand content than (upper levels) above. Most of the core sediment is black (10 YR 2/1) while in the uppermost ~20 cm the color is very dark gray (7.5 YR 3/1). Along the base of the core (~48–41.5 cm), magnetic susceptibility shows a decreasing trend from the highest value in the entire core (167 SI−5) to the lowest (29 SI−5; Figure 4b). Above this, the MS curve shows fluctuating values between 30 and 87 SI−5 between 42 and 35 cm in depth. From that level to above, it increases to 112 SI−5 before decreasing again in the last centimeter to 65 SI−5.
Charcoal
The average charcoal particle accumulation rate for core PO15-1 is 4.2 particles/cm2.year (Figure 5). The major peak is observed near the base at 222 cal BP, 1728 CE),with 12.6 particles/cm2.year). After this peak, the curve decreases until reaching a minimum at 152 cal BP, 1798 CE) with 1.8 particles/cm2.year. Above this, accumulation rates generally remain within the range of approximately 2–4 particles/cm2.year, with some minor peaks (at 124 and 82 cal BP, 1826 and 1868 CE, with 5.7 and 5.1 particles/cm2.year, respectively). Since −30 cal years BP (1980 CE), accumulation rates fall below 2 particles/cm2.year, with an absolute minimum at the top (0.8 particles/cm2.year at −65 cal year BP/2008 CE).
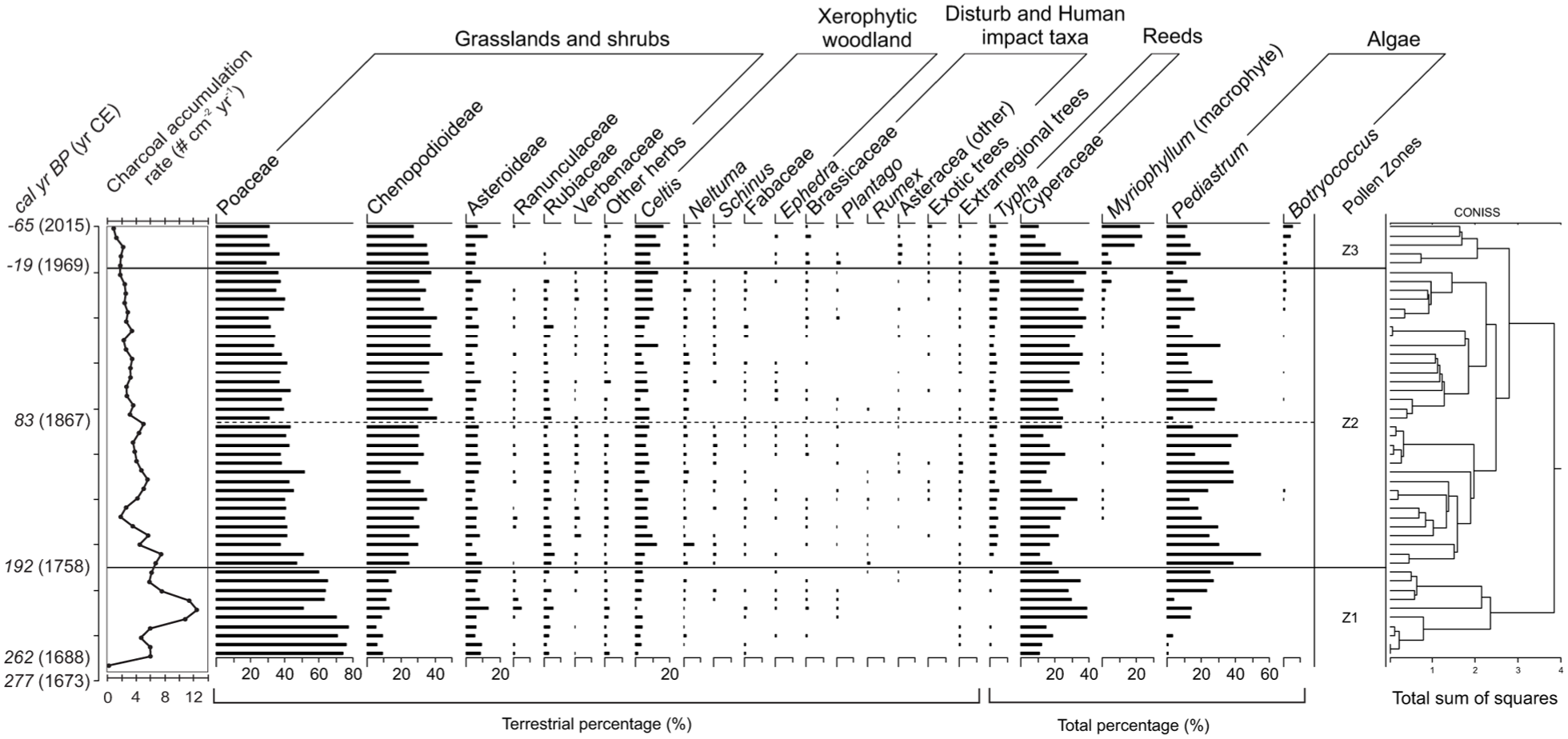
Charcoal accumulation rates (#/cm2.year) and pollen diagram of PO15-1 core from Laguna Pocitos, with the CONISS zonation and the ages of zone limits according to the age model of Figure 4.
Pollen
The pollen spectra from PO15-1 (Figure 5) shows dominance of Pampa grasslands following by taxa from the Espinal Phytogeographic Province (Oyarzabal et al., 2018). In the shrublands the predominant taxa are Poaceae, Chenopodioideae, Asteraceae sf. Asteroideae, and Rubiaceae. There are also species from xerophytic forests and shrubs such as Celtis, Schinus, Neltuma, Ephedra, and species from Fabaceae family. Aquatic and marsh vegetation is characterized by Myriophyllum (macrophyte), Cyperaceae and Typha, along with algae like Pediastrum and Botryococcus, which respond directly to changes in the aquatic system. Among exotic and extra-regional trees, traces of Pinus, Betula, Myrtaceae, Juglans, Alnus (Yungas Phytogeographic Province) and Nothofagus (Subantartic Phytogeographic Province) appear in the record (Oyarzabal et al., 2018).
The pollen spectra were divided in zones by means of CONISS. As a result, we identify three main zones (Figures 4 and 5): Z1 (47–38 cm/277 –192 cal BP), Z2 (38–5 cm/192– (−19) cal BP), and Z3 (5–0 cm/(−19)– (−65) cal BP), with the most significant change occur between zones Z1 and Z2.
Zone Z1 (277–192 cal BP/1673–1758 CE): Z1 is dominated by grasslands (Poaceae), comprising 45%–80% (average 68%) of the recognized terrestrial pollen, followed by Chenopodioideae (6%–17%; average 11%). Asteraceae sf. Asteroidea ranges from 6% to 14% (average 8.7%). Ranunculaceae reaches the highest values in the sequence, up to 4.7% of the terrestrial association. Xerophytic regional forest appears with very low values (0.5%–4.1%), but relatively diverse (e.g. Neltuma, Schinus, Celtis, Mimosoideae, and other Fabaceae). Among marsh vegetation, Cyperaceae dominates, representing 9%–35% of the total palynomorphs (average 24.7%), while Typha appears sporadically and at low levels. Aquatic components, such as the submerged macrophyte Myriophyllum, are absent, and Pediastrum is the only algae identified (average: 13%), showing an increasing trend.
Zone Z2 (192– (−19) cal BP/1758–1969 CE): Poaceae percentages are relatively high, ranging from 30% to 52% (average 40%). Chenopodioideae increases significantly, reaching 20%–45% with an average of 33%. Also, this zone is characterized by Neltuma, Schinus, and Celtis, this last one with values between 3.3% and 12.9%. Species of Verbenaceae family shows the highest values and most consistent presence in the core. Human impact taxa appear in Z2 such as Carduus, Rumex, and Plantago. Cyperaceae varies at 9%–35%, and Pediastrum at 3%–42%, with curves showing about opposite trends. In the upper half of Z2 (from 21 cm, 83 cal BP/1867 CE), Cyperaceae values are consistently higher, while Pediastrum decrease and fluctuate. Myriophyllum and Botryococcus appear intermittently at low values but increase toward the top of this zone. The proportions of Typha remain constant in the record, with low values (average 3%).
Zone Z3 (−19)– (−65) cal BP/1969–2015 CE): In this zone, Poaceae values are slightly lower (29%–35%), while Chenopodiaceae remain constant. Asteraceae sf. Asteroideae shows higher proportions compared to the rest of the core (9%–16%; average 11.66%). Herbaceous taxa appear from Ranunculaceae and Rubiaceae families, while Verbenaceae disappear from the record. Human impact taxa are present with slightly higher percentages, particularly Carduus and Brassicaceae. Xerophytic forest decreases in diversity, mainly represented by Neltuma and Schinus. Celtis reaches higher proportions with a growing trend from 8.5% to 16%. As for Cyperaceae, this family shows a decreasing trend (from 34% to 8%) in contrast to Myriophyllum (from 5% to 23%), while Typha maintains proportions similar to those in Z2. Pediastrum remains with relatively low values, and Botryococcus increases.
Stable isotopes (δ13C and δ15N), C and N content in sub-fossil samples
Throughout the core, δ13C values gradually shift to more negative ratio values toward the top, from −17.2‰ to −25.3‰ (Figure 4b). Throughout most of Z2 (30–14 cm, 141–39 years cal BP/1809–1911 CE), the δ13C curve remains around ~ −19‰, then declines to −25.3‰ toward the top (Z3). For δ15N isotopic proportions, there is an oscillation between values of ~3‰ and 7.5‰. Throughout Z1, the curve shows a significantly decreasing trend within this range, reaching the minimum around 40 cm (205 years cal BP/1745 CE), while in Z2, it increases up to 12 cm (27 years cal BP/1923 CE), then declines again toward the top (Figure 4b).
The weight percentages of C and N (strongly correlated according to the Pearson coefficient, r = 0.941, p < 0.001) show a variable but generally upward trend throughout the core (Figure 4b). In Z1, values increase rapidly (C: 0.9–10%; N: 0.1%–1%), followed by sharp fluctuations in Z2 (C: 2.9%–8%; N: 0.3%–0.9%) until reaching a peak near the top around 6 cm/1962 CE (C: 8.8%; N: 1%). The C/N ratio trend is generally decreasing throughout the PO15-1 core, with a peak near the base in Z1 (14.8 at 48 cm, 262 years cal BP/1688 CE).
Stable isotopes (δ13C and δ15N), C and N content in surface samples
δ13C and δ15N were measured on surface samples in the marginal area of Laguna Pocitos, in different geomorphological contexts and vegetation cover, such as bared sand, dune sand stabilized by semi-aquatic vegetation, grassland and woody vegetation (Figure 6a, Supplemental Table S1). Statistical analyses (ANOVA and Kruskal-Wallis) revealed no significant differences between sectors for any of the evaluated parameters. δ13C values present moderate variation across units, ranging from −30.4‰ (sample #5, sector 1 – littoral; dunes stabilized by semi-aquatic vegetation) to −25.0‰ (sample #11, sector 5 – dunes stabilized by woody vegetation). Although not statistically significant, slightly lower values tend to occur in the littoral sector, suggesting a relatively higher contribution of aquatic organic matter in these environments.
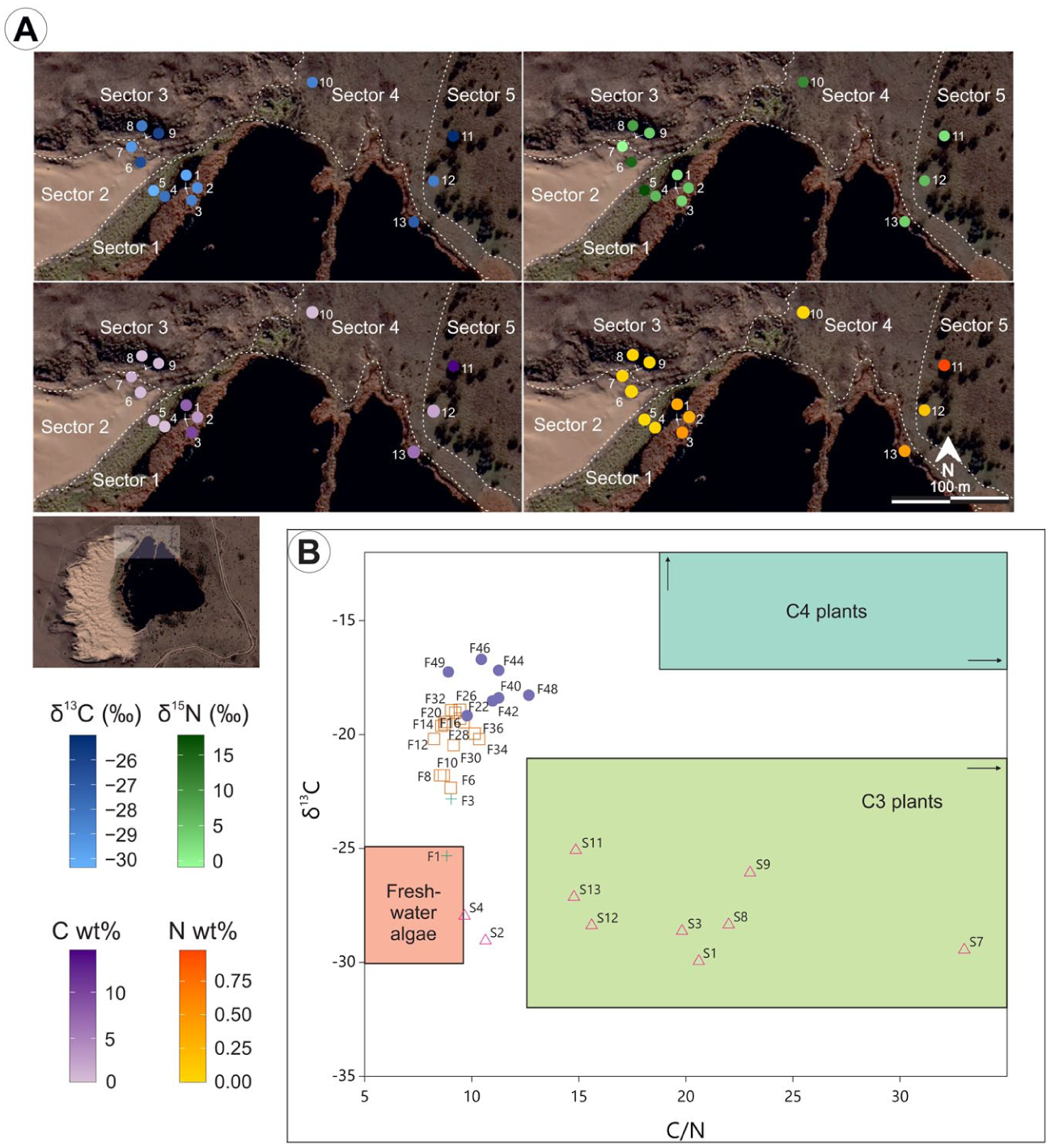
(a) Distribution of δ13C, δ15N, C and N content in surface samples from different biogeomorphological contexts surrounding Laguna Pocitos and (b) scatter plot of δ13C versus /N for surface samples (S) around Laguna Pocitos and from core PO15-1 (F).
Variability is higher for δ15N values than for δ13C, with a range of δ15N from −0.9‰ (#7, sector 2 – dunes with <30% vegetation cover) to 17.8‰ (#5). δ15N values are generally lower in sectors 1 and 5, but this pattern does not reach statistical significance. C contents vary considerably, from 0.29% (#4, sector 1) to 14.56% (#11), while N contents are consistently low, ranging from 0% (#5 and #10, sector 4 – dunes with 90% vegetation cover) to 0.98% (#11). C and N contents show a very strong correlation (Spearman: rho = 0.946; p < 0.001), reaching their maximum concentrations in the littoral zone and woody vegetation patches, reflecting localized accumulation of organic matter rather than sector-wide trends.
Discussion
The obtained results from sediment, pollen and non-pollen palynomorphs, charcoal, magnetic susceptibility and stable isotopes from Laguna Pocitos record provide evidence of variable paleoecological, paleoenvironmental and paleoclimatic conditions at the Western Pampa along the past ~350 years (~1670–2015 CE). We propose an environmental history of Laguna Pocitos based on a multiproxy analysis, which is discussed in terms of local controls and in comparison, with other Pampean paleoenvironmental and paleoclimatic inferences.
Environmental history of Laguna Pocitos (1673–2015 CE)
The analysis of the obtained results allows interpreting the following phases in the environmental evolution of Laguna Pocitos.
Magnetic susceptibility in lake sediments is related to the quantity of magnetic materials, modulated by non-magnetic components. Magnetic particles may be allogenic (windblown, tephra and pollution, and fluvial sediments) or authigenic in origin (bacterial magnetosomes, authigenic iron sulfides and reductive diagenesis; Dearing, 1999). Although some authigenic contribution cannot be ruled out, we interpret the high susceptibility values in the basal 5 cm of the PO15-1 core indicate an enhanced influx of epiclastic sediments from the surrounding dunes. Upward, the decreasing trend suggests dilution of this signal by increasing proportion of biogenic components as the water body expanded.
During this phase, the pollen record was dominated by grasslands (up to 80%), which development may have been favored by the positive hydrological balance, and in accordance with the surrounding Dry Pampa psammophytic grassland (Tonello and Prieto, 2008). A significant proportion of Cyperaceae indicates a lake margin covered by marshes, accompanied by the highest presence of Ranunculaceae, which were mentioned as common elements of wetlands (Tamura, 1993). The increasing proportions of Pediastrum toward the top of Z1 suggest a rising lake level (Medeanic, 2006).
The rates of charcoal accumulation had a peak during this first phase, indicating more frequent fire episodes, which can be related to the widespread coverage of Cyperaceae. Stable isotope signals suggest low lake primary productivity, with higher values compared to subsequent phases, while the highest C/N ratio values show that organic matter accumulation primarily from terrestrial plants (particularly those adapted to water-stressed conditions), rather than from contributions from lacustrine algae. Peaks in C and N almost coincide with charcoal peaks, which could result from an increase of terrestrial OM due to a contraction of the water body that led to higher organic matter concentration.
In situ and remote sensing analyses of the evolution of shallow lakes in the WPD over the past four decades show that the positive hydrological trend promoted wetland formation under palustrine conditions dominated by Cyperaceae. Sustained water availability leads to the development or expansion of shallow lakes. When lake levels drop, marshes widen, though the uneven relief of deflation basins linked to blowout dunes (Tripaldi and Forman, 2025) also creates deeper lakes with marshy margins (as in the present-day Laguna Pocitos). Palynological and lithological data from the base of core PO15-1, taken from the lake’s central sector, indicate that the basin initially experienced palustrine conditions, followed by progressive expansion and deepening of a shallow lake system.
In summary, during this first phase, it is interpreted a shallow lake or wetland with oligo-mesotrophic waters, and a trend of increasing water level. This trend may be a response to a more positive hydrological balance.
During this phase there was a slightly greater presence of components from the xerophytic forest, such as Neltuma and Schinus, while the appearance of exotic species suggests human impact on the area. This increase should be treated with caution, as most of these species are entomophilous (i.e. pollen transported by insects, as opposed by wind). However, it is interpretated that, during this phase, there was a significant seed dispersion of Neltuma due to cattle traffic along the indigenous roads (“rastrilladas,” D’Hiriart and Mollo, 2018). Along the study area, in the WPSL, vestiges of these “rastrilladas” are visualized today as Neltuma tree lines in aerial and satellite images (Curtoni et al., 2022).
In terms of aquatic/semi-aquatic vegetation, Typha appeared as traces, while Cyperaceae and Pediastrum showed variable and opposing trends, reflecting fluctuating water levels that produced variable sizes of the marshy peripheral zone during the second phase. Shallow lake expansion is associated with a smaller area of shallow palustrine margins colonized by Cyperaceae (Laine et al., 2010), while the dilution effect in water parameters such as salinity and trophic state may favor certain algal populations. Charcoal accumulation decreased along this phase, reaching minimum values around 1795 CE which highlights less frequent fires, then increased to a few peaks at ~1850 CE, coinciding with a higher proportion of marsh plants. After this peak, charcoal fluctuated, indicating local fire events.
A progressive contraction of the lake is inferred during the beginning of the second phase (mid-18th century), mainly suggested by the Cyperaceae and Pediastrum curves. During this transition, grasslands were displaced by other herbs from psammophytic/halophytic communities, while the number of xerophytic forest taxa decreased. This condition was followed by lake oscillations during the early to the mid-19th century. Shallow waters are inferred from ~1870 CE until the first half of the 20th century, which coincide with dominant low precipitation (instrumental data from Villa Mercedes weather station) and the reactivation of aeolian processes (Tripaldi et al., 2013). At this interval, terrestrial and marshy vegetation levels are relatively stable. A change in the lake state is recorded around 1923 CE, marked by the increase of the macrophyte Myriophyllum (associated with clearer waters), as well as with low stable isotope signals but an increase in C and N contents. This could indicate an event of isotopic dilution accompanied by a rather fast burial of OM. The short water residence time in Laguna Pocitos would explain the clear waters and relatively low salinities in a highly evaporative environment (Echegoyen et al., 2022). It is also from this date (ca. 1923) that there are higher proportions of Asteraceae sf. Asteroideae and some xerophytic forest taxa, indicating a slight development of shrubby genera in the grassland communities of the southern Pampa (Tonello and Prieto, 2008).
In summary, during the second phase it is interpreted a shallow lake with oligo-mesotrophic waters and greater level variability than before. Initially, it was characterized by fluctuating water levels and then was dominated by low levels since ~1870 CE, when an intensification of arid conditions occurred. Grasslands diminished during this phase, while disturb and exotic taxa increased, indicating a greater anthropogenic impact in the area.
Pediastrum keeps the values observed in the previous phase or slightly diminishing, while Botryococcus increases significantly. Lake’s enlargement could explain both the higher nutrient load and an increase in primary production, indicating some degree of eutrophication, but also the observed clear state could be explained by a dilution effect after a positive water balance. The highest values of Celtis can be associated with rising temperatures (Vilanova et al., 2015), which also favors algal proliferation.
Charcoal accumulation continued at low rates, following the same pattern of the end of the previous phase. For the past ~46 years, the increased number of introduced exotic taxa such as Pinus, Betula, Brassicaceae, and Carduus suggests a renewed anthropogenic pressure on Laguna Pocitos area, as many of these taxa are ornamental and forrager reflecting a different land-use from before. Simultaneously, a lower presence of herbaceous taxa from regional shrublands and forests may indicate the loss or displacement of the native flora due to stronger human activities than during previous lake phases.
The greater anthropogenic signals in the Laguna Pocitos recorded are consistent with the change in land-use in the study area. The expansion of agriculture fields, at the expense of natural grasslands, was mapped in a sector of the WPD which includes the WPSL (Demaría et al., 2008). Between 1985 and 2001, the area covered by natural Pampean grasslands decreased from 92.6% to 43.8%, and the annual conversion rate increased significantly from 1.4% to 10.9% (Baldi and Paruelo, 2008; Demaría et al., 2008).
During the third and final phase, it is interpreted that Laguna Pocitos experienced higher water levels than before, with clear waters but increased autochthonous primary production, along with a more intense anthropogenic pressure on the area.
Stable isotopes interpretation—surface and sub-fossil samples
Stable isotopes and C and N contents help distinguish potential sources of organic matter (OM) in sediments, differentiating between allochtonous and autochthonous material from shallow lakes (Fry, 2006) as well as biochemical processes controlling OM production, recycling and degradation (Meyers, 1994). Soil-derived δ13C proportions remain relatively stable after decomposition, preserving the isotopic signatures of plant sources sediments or soils OM (von Fischer et al., 2008). Interpretation of δ15N ratios is more complex due to large variations in N content and fractionation processes in both terrestrial and aquatic sediments (Talbot, 2001). The strong C-N correlation in the sub-fossil samples indicates both elements are organically bound, suggesting little selective loss or major diagenetic alteration (Talbot and Johannessen, 1992).
In the δ13C versus C/N scatterplot, isotopic distributions correspond with sample groups (sub-fossil pollen zones and surface samples) and form distinct clusters (Figure 6b), reflecting links between dominant components of vegetation and sediment isotopic signals. Higher C/N ratios primarily indicate vascular plant inputs, while lower ratios indicate more algal and macrophyte OM (Meyers, 1994), although redox fluctuations and early diagenesis can also influence C/N (Talbot and Johannessen, 1992).
PO15-1 core sediments contain a mixed composition of allochthonous (C-enriched terrestrial plants) and autochthonous OM (N-enriched algal matter, with minor zooplankton and bacterial contributions), according to reference values (Meyers, 1994; Figure 6b), indicating persistent sub-aquatic conditions over the last ~350 years (Figure 6b). Higher δ13C values in many sub-fossil samples suggest input from C4 terrestrial plants (adapted to water-deficit conditions, δ13C between −9‰ and −17‰), and/or δ13C enrichment in C3 plants as response to water stress (Ladd et al., 2009).
Samples from Z1 and part of Z2, indicate lower water availability, greater allochthonous OM contribution and limited aquatic primary production. In Z2, δ13C remains near –19‰ while C/N decreases and C and N fluctuate, indicating alternating dilution/concentration phases linked to lake-level oscillations. These dilution/concentration shifts are also reflected in δ15N variability. Z3 shows increasingly negative δ13C (to –25‰), low C/N, and rising C and N contents, indicating growing dominance of algal/macrophyte OM under more persistent flooding. Additionally, the advance of exotic pastures at the expense of native C4 grasslands such as S. pellitum may have contributed to the displacement of the isotopic signal. δ15N values reflect a nitrogen cycle controlled mainly by internal biological uptake and recycling.
Surface samples plot within the isotopic range typical of C3 plants (δ13C between −21‰ and −32‰) but show wider variability and higher C/N ratios than sub-fossil samples. Slightly lower δ13C and C/N values in some littoral samples (#2 and #4) reflect greater aquatic OM under shallow-water conditions. The absence of statistically significant differences between sectors suggests a strong small-scale spatial variability in modern sediment OM which masks contrasts between sectors, driven mainly by edaphic factors (soil moisture retention, vegetation cover), due to homogeneous precipitation and temperature.
Sectors with sparse vegetation and steep slopes, and then favoring runoff, are expected to display lower nutrient content than interdunes or areas with denser vegetation (Guevara et al., 2010). In the case of Laguna Pocitos, both bare and vegetated sand show low percentages of C and N, with one sample showing 0% wt. N despite >90% vegetation cover (Figure 6a, Supplemental Table S1).
The wide range of δ15N values observed in surface samples reflects the coexistence of multiple nitrogen cycling pathways rather than variations in a single dominant nitrogen source. Low δ15N likely reflects contributions from N-fixation and minimally altered OM, whereas high δ15N is consistent with fractionating losses (e.g. volatilization, denitrification) under intermittently saturated, evaporative conditions (Kendall, 1998). This situation allows explaining the relatively high δ15N values in geomorphological units with sandy, bare vegetated sediments. In arid and semi-arid ecosystems, woody plants such as Neltuma and other Fabaceae can locally enhance soil nutrients by moderating microclimate conditions (Guevara et al., 2010 and citations therein). This is the case for surface sample #11 (Neltuma forest), which contains the highest weight percentages of C and N.
The PO15-1 core isotopic and elemental evolution reinforce the interpretation of a transition from a terrestrially dominated, hydrologically unstable palustrine system (Z1), through fluctuating shallow-lake conditions (Z2), to a more stable, autochthonous-dominated shallow lake in recent decades (Z3). Spatial analysis with surface samples, as a modern functional analog, shows that edaphic conditions are the main control in isotopic and nutrient composition, but still vegetation patterns and hydrological variability have a correlate in OM source patterns.
Additional sampling (including more surface lake samples), as part of a work in progress, will allow refining these patterns, addressing the spatial heterogeneity in Laguna Pocitos and its catchment area and providing a more robust basis for interpreting past conditions.
Hydroclimatic variability, climate patterns and land-use changes from the Laguna Pocitos record
The multiproxy record obtained from Laguna Pocitos is here considered in the context of paleoclimatic variability in subtropical southern South America over the last ~350 years cal BP (since 1670 CE) and considering the changes in land-use in the study area. Situated in an ecotonal zone between the Arid Diagonal to the west and the humid Pampa grasslands to the east, the WPSL is particularly sensitive to shifts in atmospheric circulation and hydrological balance. Weather stations from the Western Pampa recorded a much lower annual precipitation during the first half of the 20th century than during the second half, which produced a displacement of the isohyets to the west (Berton and Echeverría, 1999; Viglizzo et al., 1995). Different climatological studies indicated that El Niño Southern Oscillation (ENSO) and different configurations of the South American Monsoon System (SAMS) are the most relevant teleconnections associated with this summer precipitation variability in central-western Argentina (Barros et al., 2002; Garreaud et al., 2009 and references therein).
The comparison of the hydroclimatic variability inferred from the Laguna Pocitos record with other paleoclimatic reconstructions reveals some periods of comparable climatic conditions for central Argentina during the past ~350 years (Figure 7). Along this period, different types of anthropogenic signals appear in the Laguna Pocitos record, in accordance with the history of land-use.
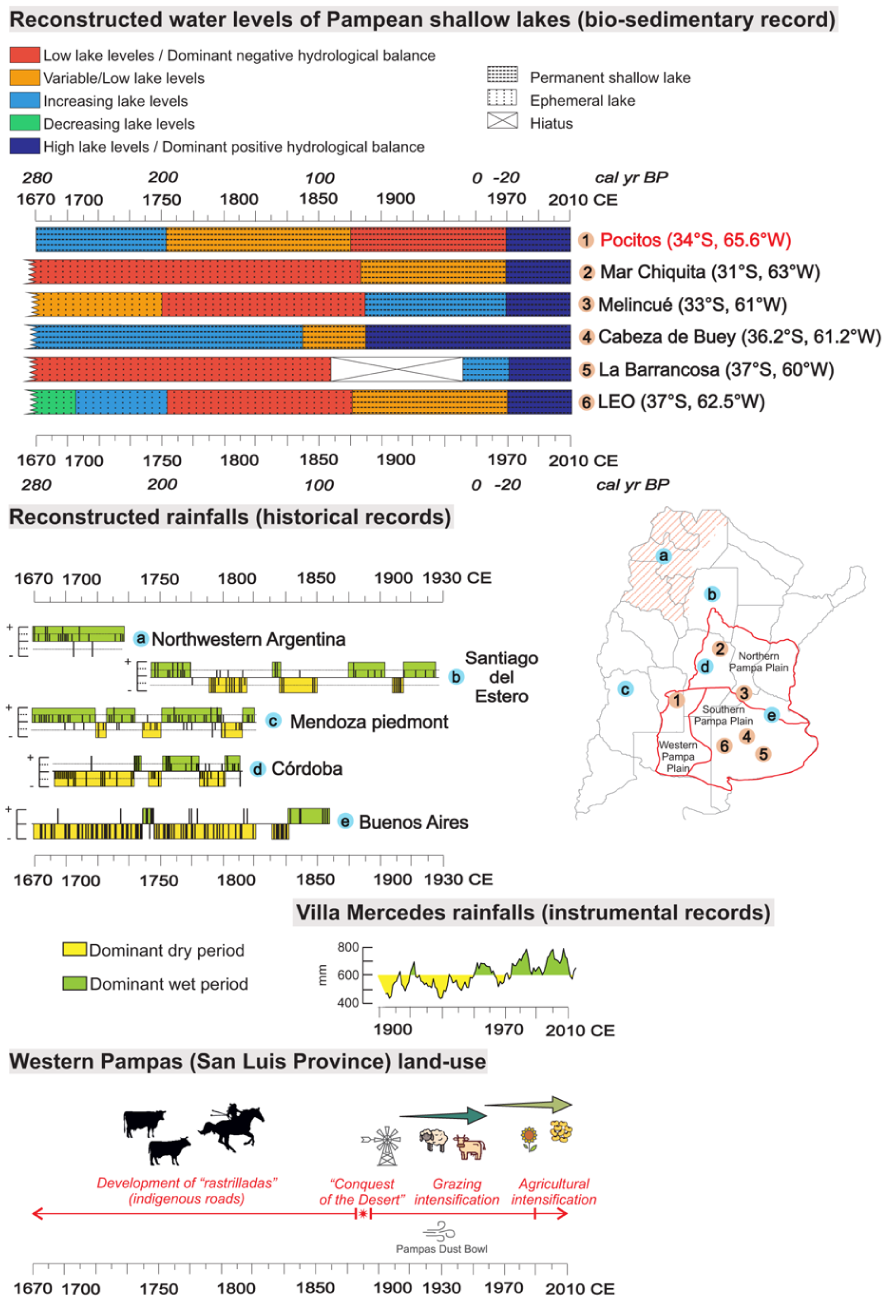
Comparison of some paleolimnological records from the Pampa Plain (subregions after Piovano et al., 2025), documentary source records from Argentina, instrumental records from Villa Mercedes and the changes of land-use in Western Pampas.
Period I (ca. 1670–1750 CE): The initial Laguna Pocitos record (second half of the 17th century) coincides with abnormally wet conditions in localities to the NW and W of the WPSL (ca. 1630–1790 CE). Higher rainfalls than before in Northwestern Argentina, Santiago del Estero and Mendoza piedmont were interpreted from documentary sources of the colonial (mid-16th century–early 19th century) and republican (since early 19th century) historical periods of Argentina (Prieto et al., 2000; Prieto and García Herrera, 2009; Figure 7). Rainfalls in those regions, to the east of the Andes Cordillera, at least during summer months, have a significant correspondence, as the study area, to ENSO and SAMS forcings. This positive hydrological balance likely promoted the formation of a shallow lake or wetland, accumulating mud and organic matter. This period coincides with the Little Ice Age (LIA; 1450–1850 CE; Villalba, 1994), marked by hydroclimatic instability with alternating dry and wet pulses linked to cooler conditions (Córdoba et al., 2014). Such variability may have caused fluctuating water levels within the deflation basin, trending toward higher levels. Similar patterns (frequent desiccation and fire events) were recorded at Laguna Nassau and Primera Laguna (Vilanova et al., 2015, 2022; Figure 1).
Paleoenvironmental reconstructions in shallow lakes of the Pampa plain exposed hydrological heterogeneity during the last millenium, with large water-lake levels fluctuations between wet and dry phases of variable intensity and duration (Córdoba et al., 2025; Figure 7). Northern Pampa records (Mar Chiquita, Melincué) indicate arid conditions, low lake levels, saline waters, and recurrent desiccation (Cuña-Rodríguez et al., 2020; Guerra et al., 2015, 2017, 2025). In contrast, Central–Southern Pampa sites (La Barrancosa, Cabeza de Buey) show spatially variable responses: La Barrancosa remained a small, saline wetland (Plastani et al., 2019, 2025) while Cabeza de Buey indicates humid, turbid freshwater after 1630 CE (Sánchez Vuichard et al., 2021). In the Southwestern Pampa (Lagunas Encadenadas del Oeste, LEO system), this late inference coincides with a humid pulse since 1530 CE that led to low- intermediate water levels (Córdoba et al., 2014, 2025).
Period II (ca. 1760–1870 CE): From the late 18th century, Laguna Pocitos shows fluctuating water levels, consistent with widespread drought–flood cycles across Northwest, West and Central Argentina, when drier conditions intensified since ca. 1780/1790 (Prieto et al., 2000). “The Great Drought” (1800–1840 CE; Tapia and Dussart, 2013; Tonni, 2023), noted by Darwin (1846), marked prolonged aridity in Western Pampa, while wetter years increased in Eastern Pampa after 1840 (Scarpati and Capriolo, 2013).
Paleolimnological records confirm this regional dryness: nearly desiccated lakes in Northern Pampa (Mar Chiquita, Melincué; Cuña-Rodríguez et al., 2020; Guerra et al., 2017), negative water balances in Central Pampa (Cabeza de Buey; Sánchez Vuichard et al., 2021; La Barrancosa; Plastani et al., 2019, 2025), and low lake levels in the LEO system (Córdoba et al., 2014, 2025; Figure 7).
At Laguna Pocitos area, pollen data suggest growing anthropogenic influence. The spread of horses and cattle during the 18th–19th centuries transformed indigenous (Rankülche) land-use, increasing grazing and transit (Curtoni, 2007). Movement along “rastrilladas” likely promoted fires and grassland reduction, as well potentially impacted soil cover and windborne sediment accumulation (León and Anderson, 1983). The decline in Poaceae and rise of Chenopodioideae after 1760 CE indicate such impacts, possibly accompanied by xerophytic taxa dispersal via cattle (D’Hiriart and Mollo, 2018). Variable charcoal accumulation in lower Z2 reflects these drier conditions and more extensive marshes.
An archeological report, which includes interviews with local inhabitants of the WPSL area, highlighted that numerous fires affected this landscape during the 19th century (Greslebin, 1961). The testimony of a resident in 1923 indicates that his father, who died at about 90 years old in ~1874, remembered this landscape as “a beautiful grassland and impenetrable woodlands,” and that there were successive, large fires (Greslebin, 1961).
Period III (1870–1970 CE): By the late 19th century, most Pampean lake records show marked ecological shifts and increasing anthropogenic pressure. In Laguna Pocitos, water levels stabilized but remained lower than before, and charcoal decreased. Intensified human activity is suggested in Western Pampa since the end of the 18th century, when the inhabitants of these lands moved more frequently between Chile and Argentina, leading to an increase in fire frequency in La Pampa Province (Sastre et al., 2018; Tapia and Dussart, 2013). In the dune field area, following the so-called “Conquest of the Desert” at about the onset of this period (1880 CE), the indigenous roads fell into disuse (D’Hiriart and Mollo, 2018), which could explain the lower fire frequency in Laguna Pocitos area despite increased aridity. Regionally, native vegetation was nearly replaced by cereal agriculture (Viglizzo et al., 2011), and agricultural expansion reached the northern WPD by the late 20th century (Baldi and Paruelo, 2008; Demaría et al., 2008).
During the first half of the 20th century, instrumental data record dry decades (Figure 2b), notably the 1930s “Pampas Dust Bowl” (Viglizzo and Frank, 2006), with rainfall reductions between 33% and 62% (Tripaldi et al., 2013). This dry spell, amplified by anthropogenic factors, triggered aeolian reactivations in dunefields of Western Pampa (San Luis and La Pampa provinces; Tripaldi et al., 2013). Laguna Pocitos (Figure 4b) and the nearby Primera Laguna (Figure 1b; Vilanova et al., 2022, Figure 5) registered peaks of fine sand related to this dune reactivations.
Paleolimnological records show predominantly low-intermediate water levels and recurring droughts, especially in Western Pampa (Córdoba et al., 2025; Figure 7). In Northern Pampa, very low lake levels remained in Mar Chiquita, interrupted by short humid pulses (Cuña-Rodríguez et al., 2020; Piovano et al., 2009), while a shift to permanent, saline to subsaline, eutrophic waters was inferred for Melincué (Guerra et al., 2017). In Central Southern Pampa, humid conditions with higher than before water levels were inferred at Cabeza de Buey (Sánchez Vuichard et al., 2021). In La Barrancosa a hiatus at 1860–1940 CE was interpreted as the Pampas Dust Bowl record, followed by the onset of permanent, subsaline conditions (Plastani et al., 2019, 2025). Southwestern Pampa records indicate a shift to permanent conditions, with intermediate levels in LEO lakes (Córdoba et al., 2014, 2025).
Period IV (1970–2015 CE): Since the 1970s, increasing precipitation in the Pampean plains, linked to the Current Warm Period and enhanced SAMS activity (Córdoba et al., 2014) produced a marked rise in Laguna Pocitos water levels, registered in the biosedimentary succession, and the expansion of lake area, evidenced by satellite images since 1985; this enlargement caused the relocation of a former road (M. Roca, pers. comm., 2008; Figure 1c).
Similar positive trends occurred in several Pampean lakes (e.g. Mar Chiquita, Melincué, La Barrancosa, Cabeza de Buey, LEO, Nassau, Primera Laguna; Figure 7). Although many of these lakes shifted to turbid, eutrophic states due to high levels of nutrient input, Laguna Pocitos has remained clear, likely owing to its deep deflation basin, groundwater inflow, dune recharge, and relatively limited agricultural pressure in the very near pasture fields.
Conclusions
The multiproxy record from Laguna Pocitos, in the northern WPD, documents ~350 years of hydroclimatic variability and anthropogenic interactions in the ecotonal zone of the Western Pampa. The inferred changes reflect the local expression of complex interactions between regional climate dynamics (particularly the SAMS) and global phases such as the Little Ice Age, under increasing anthropogenic influence.
Three main lake phases were recognized based on vegetation, magnetic susceptibility, charcoal, C and N content, and isotopic data. During the first phase (~1670–1758 CE), and despite regionally arid conditions (Cuña-Rodríguez et al., 2020 and references therein), a shallow lake or wetland was well established, with rising water levels, oligo-mesotrophic conditions, and grassland-dominated vegetation. The second phase (~1758–1969 CE) shows grassland decline and expansion of disturbance and exotic taxa, indicating growing anthropogenic impact. At first, the oligo-mesotrophic water body was characterized by fluctuating levels (~1758–1867 CE), and lower levels than before were inferred afterward, consistent with reduced regional precipitation (Cuña-Rodríguez et al., 2020 and references therein). The final phase (~1969–2015 CE) marks the expansion and deepening of Laguna Pocitos, clearer waters dominated by Myriophyllum, and increased anthropogenic disturbance through land cultivation and loss of native flora. Isotopic evidence suggests greater input from algal and macrophyte organic matter, reflecting lake’s enlargement and eutrophication.
The main hydroclimatic inferences for the Laguna Pocitos record align with other climatic reconstructions from central and northern Argentina based on lake cores and documentary sources. Laguna Pocitos record reflects the local impacts of major climatic phases such as the fluctuating moisture availability and intermittent arid conditions during the 18th and 19th centuries (i.e. the end of the LIA). However, despite frequent droughts, many Pampean shallow lakes, including Laguna Pocitos, persisted as water bodies, with episodes of relative stability or deepening linked to transient increases in precipitation. Anthropogenic impact (horse herds and cattle movement during the 18th and 19th centuries, early 20th century pasture cultivation and cattle growing, and the past four decades of agricultural intensification) have contributed to vegetation changes, lake water conditions, and sediment dynamics (e.g. the “Pampas Dust Bowl”). Rising water levels in recent decades responded to enhanced SAMS activity and regional warming corresponding with broader patterns across the Pampean plain. Overall, the Laguna Pocitos record supports the importance of integrating paleoenvironmental and historical records to interpret climate-landscape-human relations in climatically sensitive zones.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836261450817 – Supplemental material for Multiproxy study of past three centuries of climatic variability and anthropogenic impact on Laguna Pocitos, western Pampa of Argentina
Supplemental material, sj-xlsx-1-hol-10.1177_09596836261450817 for Multiproxy study of past three centuries of climatic variability and anthropogenic impact on Laguna Pocitos, western Pampa of Argentina by Vera Maciel, Isabel Vilanova, Guillermo Heider, Karsten Schittek and Alfonsina Tripaldi in The Holocene
Footnotes
Acknowledgements
Several analyses performed in this research would not have been possible without the information generated and shared by the Argentinian public institutions Servicio Meteorológico Nacional (SMN), Instituto Nacional de Tecnología Agropecuaria (INTA), Museo Argentino de Ciencias Naturales (MACN), Universidad de Buenos Aires (UBA), Facultad de Ciencias Exactas y Naturales (FCEN, UBA), and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). The authors are deeply grateful to Dr. Steven Forman, Dr. Eduardo Piovano, and Dr. Esteban Jobbagy for their invaluable academic support, to Dr. Leandro Rojo for helping with palynological determination and laboratory facilities, and to Gastón Angelini, Kathy Breen, and Ricardo Paéz for their assistance in the field. We are grateful to the many farmers and inhabitants of the San Luis Province whose generosity and hospitality made this study possible.
Ethical considerations
This article was not required of ethical approval.
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research is a contribution to the grants PUE-IGEBA-CONICET 22920160100030CO and PIP-CONICET 11220210100052CO to AT.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Raw data of this paper can be found in the supplemental material file and will be opportunely shared in a public repository.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.