
Abstract
Kretzschmaria deusta is a wood-parasitic ascomycete affecting a range of broadleaf trees in temperate regions, including Europe, causing wood decay. Its ascospores, as an example of Non-Pollen Palynomorphs (NPPs), are preserved in Quaternary sediments, providing a rare record of past host-pathogen associations. This study synthesizes the Holocene palaeoecological record of Kretzschmaria deusta, examining its spatial and temporal distribution, host preferences, and ecological impacts. Early Holocene occurrences are sparse in Europe, whereas Middle and Late-Holocene spores indicate widespread presence, coinciding with the expansion of deciduous forests. Throughout the Holocene, Kretzschmaria deusta acts primarily as an opportunistic pathogen, exploiting trees weakened by natural and anthropogenic factors. Elevated spore abundances may reflect periods of increased tree damage rather than primary forest decline. Host responses vary among taxa. Tilia tolerates repeated infection, whereas Fagus, Abies, and Ulmus are more sensitive, with higher fungal abundances corresponding to possible host decline. Warm temperatures during the Holocene further facilitated fungal growth and infection success. The predominantly local dispersal of spores suggests that this NPP record reflects nearby woodland conditions, although episodic transport by wind or water may cause temporary increases. The Holocene findings of Kretzschmaria deusta enhances our understanding of long-term tree-fungus associations and highlights the potential role of this fungus in forest decline under future warming and intensified anthropogenic pressure.
Introduction
Parasitism is one of the most widespread modes of existence in nature. Parasites exploit hosts for nutrients, reproduction, and dispersal, while conferring no benefits and often imposing biological costs (Perlin et al., 2025). Virtually every organism, including parasites themselves, may serve as a habitat and target of exploitation. Host-parasite interactions involve reciprocal responses: hosts limit parasite activity, while parasites must evade or suppress host defences (Frank, 1996; Schmid-Hempel, 2008). Consequently, these interactions often lead to relatively stable associations, in which parasites avoid completely destroying their hosts to maintain a reliable source of resources and habitat, while host defences restrict exploitation to levels that do not result in the host’s mortality (Clayton et al., 2019). Such relationships allow parasites to access resources of higher quality than most forms of dead organic matter (Frank, 1996; Schmid-Hempel, 2008).
Among parasitic organisms, fungi constitute an important group. This group is particularly interesting because a fungus itself is inherently part of a relationship with another organism. Its absorptive mode of nutrition ensures that such an association always exists, whether in symbiosis, saprotrophy, or parasitism (Rossman, 2009). However, only in parasitism does the fungus engage in a relationship in which it exploits a living host, often resulting in the host’s weakening or disease. As parasites, fungi employ diverse strategies for host colonization, including spores, hyphal invasion, and toxin production (Liu et al., 2009).
Plant parasites constitute a major group of parasitic fungi. Characteristic indicators of their presence on a host plant include the mycelium and various fungal structures, such as fruiting bodies and spores. Parasitic fungi exploiting living host plants may cause visible disease symptoms depending on the host-pathogen interaction. The most common symptoms include spots and deformities resulting from tissue responses to fungal infection (Marcinkowska, 2012). Fungi induce necrosis of plant tissues, resulting in spots, cankers, and rots of various shapes, sizes, and colours. Certain disease symptoms are characteristic of specific groups of fungal pathogens.
Much is known about contemporary parasites; however, their presence and functioning in the past remain poorly understood. Most studies on the evolution of parasites are based on phylogenies of extant species constructed from morphological and molecular data. Unfortunately, these approaches provide only an incomplete picture of parasite-host interactions (Leung, 2017). Direct evidence of parasitic species in past environments is therefore particularly valuable, but fossil records are generally very sparse. In Quaternary sediments, their presence can be inferred from so-called Non-Pollen Palynomorphs (NPPs), microfossils that, as the name indicates, are not pollen grains. Although distinct from pollen, their identification is closely associated with palynological analysis, as they are recovered from the samples. NPPs include green algae, cyanobacteria, remains of other algae, invertebrates, microscopic plant structures, testate amoebae, fungal spores, as well as structures of unknown origin. One of the parasites whose past presence can be detected on the basis of NPPs is the fungus Kretzschmaria deusta. Its spores were first identified and described from organic sediments by Van Geel (1978). Currently, Kretzschmaria deusta of Xylariaceae is known as a parasitic fungus associated with a wide range of broadleaf tree taxa, where it is linked to root, butt, and trunk decay as well as canker formation. The decay caused by it involves necrosis of extensive tissue areas, and sometimes even entire organs, resulting from enzymatic breakdown of cells. This process is classified as a dry-type wood decay. The fungus continues the decomposition of wood after the host tree has died (Rogers and Ju, 1998). In temperate regions, K. deusta has been reported as one of the main causes of reduced stability and windthrow in Acer spp., Aesculus spp., Fagus spp., Tilia spp., and Ulmus spp. trees (Terho and Hallaksela, 2008).
It is well established that parasites currently constitute important drivers of ecosystem dynamics, exerting far-reaching effects on their structure and functioning. Parasitism constitutes an important evolutionary driver, which can promote genetic diversity in host populations and speciation (Evison et al., 2013). It is highly likely that this was also the case in the past. Therefore, understanding the role of parasitism in the past may be crucial for reconstructing the complexity of former ecosystems (Harper and Krings, 2021).
The aim of this paper is to provide a comprehensive synthesis of the Holocene record of the wood-parasitic fungus Kretzschmaria deusta in Europe, with particular emphasis on its spatial and temporal distribution. Additionally, the study aims to compile and summarize available evidence on the fungus’s potential host preferences and ecological impacts, thereby contributing to a better understanding of long-term forest–pathogen interactions, changes in forest cover, and the role of K. deusta in tree functioning.
Spores of Kretzschmaria deusta as non-pollen palynomorph (NPP)
These are ascospores that can be found alongside pollen during palynological analysis. They represent one of over 1300 currently recognized non-pollen palynomorphs (NPPs; Miola, 2012). They were first described by Van Geel (1978) as “Type 44,” considered at the time to represent a xylariaceous ascomycete. Their association with the species Ustulina deusta (the former name of Kretzschmaria deusta) was established by Pals et al. (1980), who referred to them as “Type HdV-117.” Today, these spores are most commonly identified simply as K. deusta, occasionally with the designation HdV-44. They are fusiform, smooth, non-septate, about 35 × 8 μm, with acuate, thick-walled ends (Figure 1). One side of the spores is flattened, bearing a longitudinal germ slit, much shorter than the total spore length (van Geel et al., 2013). Studies of contemporary K. deusta indicate that, although the fungus is also known to produce asexual conidia, infection of host trees predominantly occurs via sexual spores (ascospores). Regarding their dispersal, ascospores are typically transported over shorter distances than pollen grains. This is because fruiting bodies develop at the base of tree trunks, near the forest floor, where wind dispersal is relatively inefficient. Therefore, the presence of ascospores in sediments is more likely to reflect local transport and may indicate the close proximity of a forest. This was highlighted by Van Geel and Andersen (1988), who analysed Eemian and Holocene records of these spores. Root infection via vegetative mycelium is also possible (Guglielmo et al., 2012). However, from a palaeoecological perspective, neither conidia nor mycelium are useful for identification. Asexual spores lack a characteristic morphology suitable for recognition in pollen slides (van Geel et al., 2013). Similarly, vegetative mycelium cannot be used as an NPP because, unlike Clasterosporium caricinum, whose hyphopodia are recognized as HdV-126, no diagnostic characters corresponding to the hyphopodia of K. deusta have been recognized (see Miola, 2012; van Geel and Aptroot, 2006).
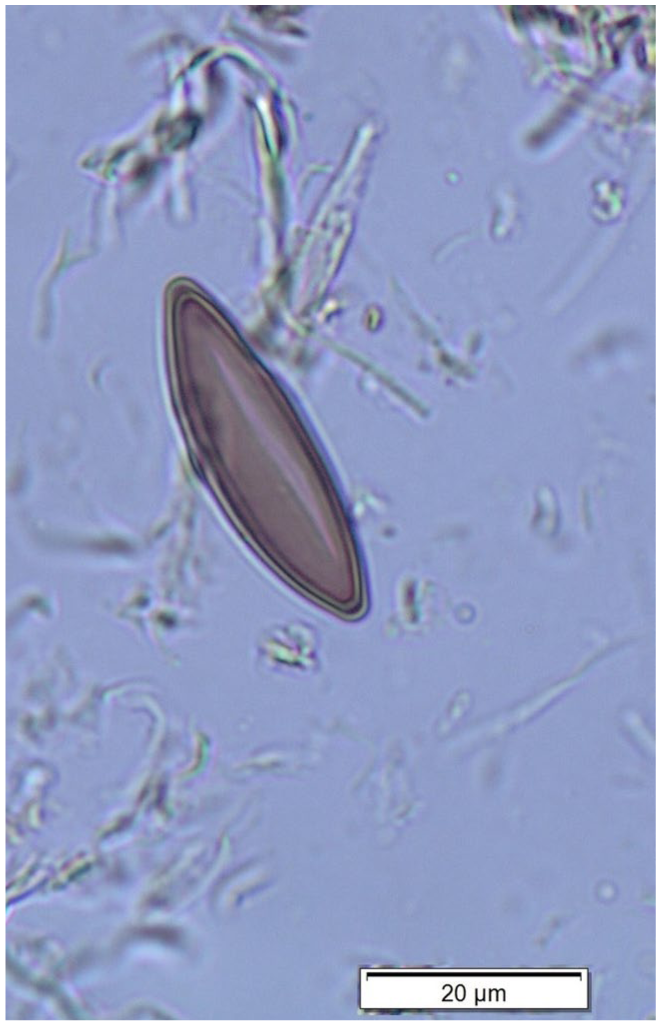
Ascospore of Kretzschmaria deusta. Picture taken by D. Drzymulska from Poganowo microfossil slide. Material published by Szal et al. (2015).
Holocene microfossils of Kretzschmaria deusta in Europe
The fossil record of Kretzschmaria deusta has been compiled using a systematic literature synthesis. A search was conducted in Web of Science, Scopus, and Google Scholar applying the species name (Kretzschmaria deusta or Ustulina deusta) combined with keywords related to past occurrences, including “Holocene,” “subfossil spores,” “fossil spores,” and “palaeoecology.” Additional search terms included “peat,” “lacustrine,” and “sediment,” which were combined with the species names using Boolean operators. A few relevant publications were identified through reference list screening. Retrieved records underwent a two-stage selection process. In the first stage, abstracts were assessed, and studies not referring to fossil or subfossil records of K. deusta/U. deusta, the Holocene, or palaeoecological contexts were excluded. In the second stage, full texts were evaluated according to predefined inclusion and exclusion criteria. Inclusion criteria comprised studies reporting palynological data, non-pollen palynomorphs (NPPs), or sediment-based analyses from Holocene deposits in Europe. Exclusion criteria included non-Holocene records, non-European studies, and modern-only datasets. Data were assembled specifying the sites from which the remains were recovered, sediment characteristics, the subepochs of Holocene used in the quoted paper, and estimated time depth of ascospore-bearing sediments (Table 1). Figure 2 shows the location of the Kretzschmaria deusta microfossil finds, with records of different ages indicated with different symbols. The names of the Holocene subepochs (subseries) were used by Walker et al. (2019).
Records of Holocene spores of Kretzschmaria deusta in Europe, where + indicates occurrence, na1 – not assessed due to insufficient spore occurrence (single/sporadic records), na2 – not assessed due to insufficient age data.
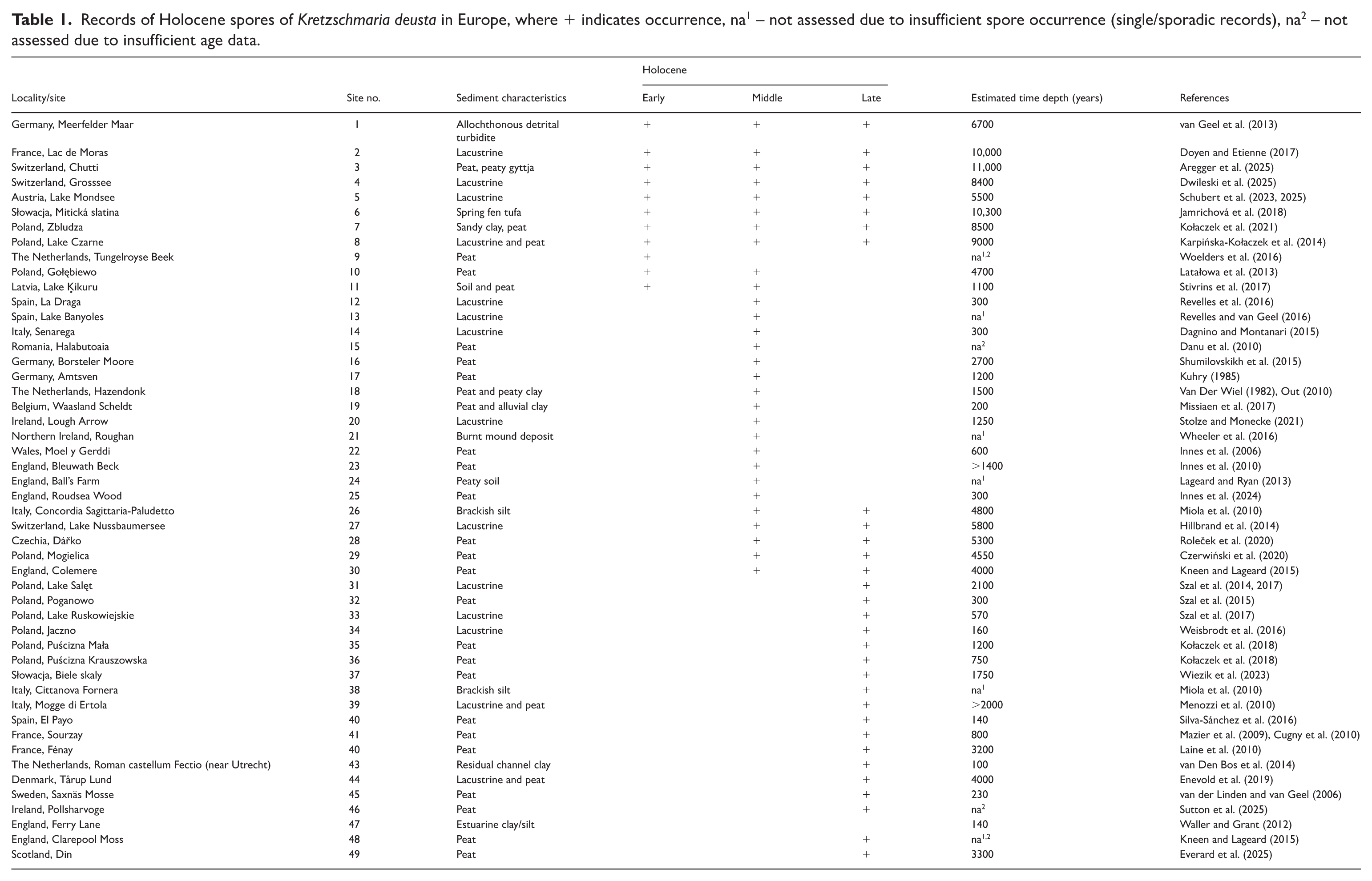
Map of sites with Holocene Kretzschmaria deusta spores in Europe. Numbers as in Table 1.
Spores of K. deusta are known from nearly all parts of the European continent, predominantly from Middle and Late-Holocene sites (Figures 2 and 3). Only eight locations have been identified in which the spores occurred in all three Holocene subseries, namely Meerfelder Maar in Germany (van Geel et al., 2013), Lac de Moras in France (Doyen and Etienne, 2017), Chutti (Aregger et al., 2025) and Grosssee, both in Switzerland (Dwileski et al., 2025), Lake Mondsee in Austria (Schubert et al., 2023, 2025), Mitická slatina in Slovakia (Jamrichová et al., 2018) and in Poland – Lake Czarne (Karpińska-Kołaczek et al., 2014) and Zbludza (Kołaczek et al., 2021). In the Early Holocene, the spores were recorded from the Tungelroyse Beek valley in the Netherlands (Woelders et al., 2016), from Gołębiewo in Poland (Latałowa et al., 2013) and from Lake Ķikuru in Latvia (Stivrins et al., 2017).
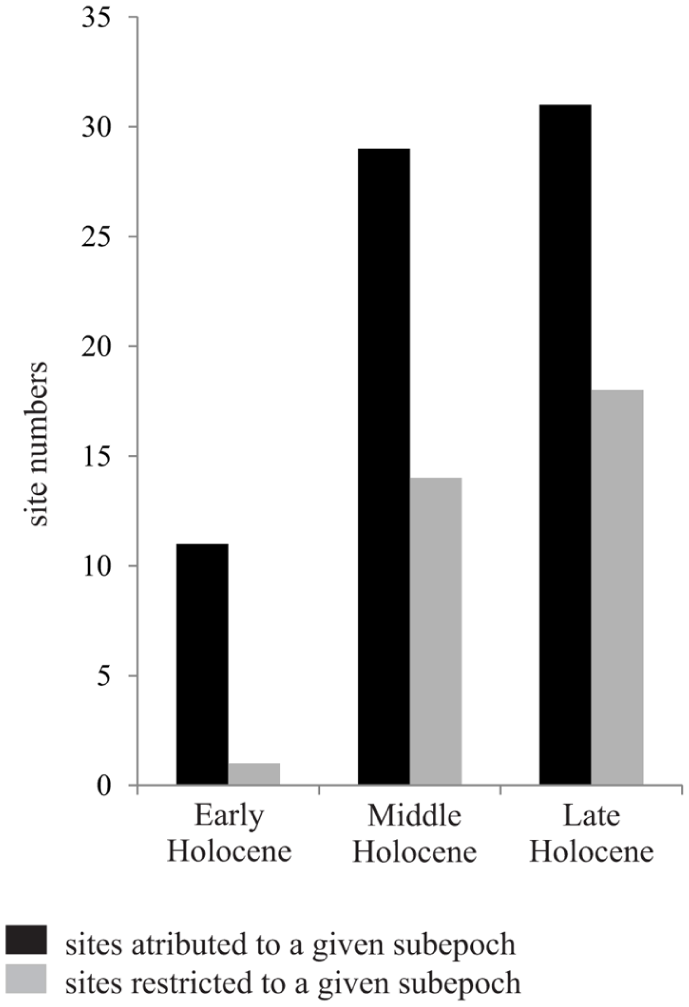
Holocene record of Kretzschmaria deusta spores.
During the Middle Holocene, microfossils of K. deusta were recorded across various regions of the European continent. The southernmost occurrence is documented in Spain at La Draga (Revelles et al., 2016) and Lake Banyoles (Revelles and van Geel, 2016). Further north, records are known from Senarega in Italy (Dagnino and Montanari, 2015) and Halabutoaia in Romania (Danu et al., 2010). In Germany, spores were identified in sediments from Borsteler Moor (Shumilovskikh et al., 2015) and Amtsven (Kuhry, 1985). Additional Middle Holocene records include Hazendonk in the Netherlands, initially investigated by Van Der Wiel (1982) and later by Out (2010), the Waasland Scheldt area in Belgium (Missiaen et al., 2017), and the previously mentioned Gołębiewo locality in northern Poland (Latałowa et al., 2013). On the British Isles, HdV-44 findings have been reported from Middle Holocene deposits in Ireland at Lough Arrow (Stolze and Monecke, 2021), in Northern Ireland at Roughan (Wheeler et al., 2016), and at Moel y Gerddi in Wales (Innes et al., 2006). In England, occurrences have been documented at Bleuwath Beck (Innes et al., 2010), Ball’s Farm (Lageard and Ryan, 2013), and Roudsea Wood (Innes et al., 2024).
Both Middle and Late-Holocene records of K. deusta spores are distributed across Europe. In Italy, they have been reported from Concordia Sagittaria-Paludetto (Miola et al., 2010), while in Switzerland they are known from Lake Nussbaumersee (Hillbrand et al., 2014). Further north, occurrences have been identified at Dářko in Czechia (Roleček et al., 2020) and Mogielica in Poland (Czerwiński et al., 2020). On the British Isles, a record from Colemere in England has also been documented (Kneen and Lageard, 2015).
In the Late-Holocene, the presence of K. deusta spores has been confirmed at numerous localities across Europe, with the highest concentration reported from Poland, including Lake Salęt, Poganowo, and Lake Ruskowiejskie (Szal et al., 2014, 2015, 2017, respectively), Jaczno (Weisbrodt et al., 2016), and Puścizna Mała and Puścizna Krauszowska (Kołaczek et al., 2018). A slightly further south location is Biele skaly in Slovakia (Wiezik et al., 2023). In southern Europe, HdV-44 occurrences were identified in Italy at Cittanova Fornera (Miola et al., 2010) and Mogge di Ertola (Menozzi et al., 2010), in Spain at El Payo (Silva-Sánchez et al. (2016), and in France at Sourzay (Cugny et al., 2010; Mazier et al., 2009). Further north, one more locality tknown from France is – Fénay (Laine et al., 2010). In the Netherlands spores of K. deusta were found at Roman castellum Fectio near Utrecht (van Den Bos et al., 2014). In Denmark these finds were recorded at Tårup Lund (Enevold et al., 2019) and in Sweden at Saxnäs Mosse (van der Linden and van Geel (2006). In Ireland, a Late-Holocene record is known from Pollsharvoge (Sutton et al., 2025), and in England from Ferry Lane (Waller and Grant, 2012) and Clarepool Moss (Kneen and Lageard, 2015). The northernmost occurrence has been documented at Din in Scotland (Everard et al., 2025).
On the palaeoecology of Kretzschmaria deusta
Data on the ecological preferences of this ascomycete fungus during the Early Holocene remain very limited. Schubert et al. (2023) reported a sensitive response of K. deusta in the Lake Mondsee sediment record to the cooling event around 8200 cal yr BP, which was manifested by a pronounced minimum in ascospore concentrations. This finding suggests that the ascospore record of K. deusta may potentially provide useful information for reconstructing Holocene environmental variability, although its value as a proxy should be interpreted with caution due to potential effects of dispersal and depositional setting. It should be noted that the interpretation of ascospore abundances must always consider the potential influence of long-distance dispersal, transport processes (wind, water), and site-specific depositional conditions. These processes may introduce allochthonous material and potentially weaken the link between spore abundances and strictly local ecological conditions. By contrast, spores preserved in peat deposits are generally more likely to reflect a local signal, although some degree of regional input cannot be excluded. With regard to possible host trees, K. deusta may have been associated during the Early Holocene with Tilia. Aregger et al. (2025) noted a clear positive correlation between the percentage curves of K. deusta spores and Tilia pollen at the Chutti site in Switzerland, with both increasing synchronously. A similar synchronous increase was also observed at Mitická slatina. However, the authors interpreted this pattern there as a response to increasing climatic humidity (Jamrichová et al., 2018). A possible susceptibility to infection by K. deusta has also been suggested for taxa other than Tilia, including Salix, Ulmus and, in particular, Alnus, as recognized in the Tungelroyse Beek valley by Woelders et al. (2016). However, the very low abundance of spores in the sediment, along with the difficulty in precisely constraining their time depth, should be considered when interpreting the results. On the other hand, it has been established that Alnus occurred in situ, as indicated by numerous macroremains in the deposit, probably representing Alnus roots. At present, alder is also known to act as a host for K. deusta in temperate regions of the Northern Hemisphere (van Geel and Aptroot, 2006).
From the Middle Holocene onwards, this fungal species may have acted as an additional stress factor contributing to tree mortality driven by climatic change, shifts in land use, slope processes, erosion and fire. Such interactions have been documented in the eastern Swiss Alps at Grosssee, where around 6800 cal yr BP a marked decline in arboreal Alnus glutinosa and a concurrent expansion of the shrub Alnus viridis were recorded (Dwileski et al., 2025). Fungal indicators may therefore point to a role of fungi in the formation of gaps in the forest canopy. An example closely associated with the microfossil fungal record, including the presence of Kretzschmaria deusta spores, is the mortality of prehistoric bog oak woodland at Ball’s Farm, England (Lageard and Ryan, 2013). Comparable patterns of mid-Holocene woodland decline have also been identified at other sites in the British Isles. A coincidence between declining Ulmus pollen frequencies and increasing abundances of ascospores of this fungus was observed during the Late Mesolithic at Bluewath Beck (Innes et al., 2010) and was even more pronounced during the Early Neolithic at Meol y Gerddi (Innes et al., 2006). Forest farming practices, probably in the form of tree girdling, were likely used at that time as a method of exploiting mixed deciduous woodland. In response to girdling, trees produced numerous sprouts prior to their death, which were subsequently used for free-ranging cattle within the woodland. Trees damaged in this way by human activity became ideal substrates for infection and rapid wood decomposition by Kretzschmaria deusta. Therefore, peaks in K. deusta and the decline of Ulmus could potentially be interpreted as a proxy for local human land-use associated with girdling and stock browsing. This supports the ecological interpretation of this ascomycete as being favoured by certain forms of woodland exploitation. K. deusta is an opportunist that takes advantage of damaged trees, requiring open wounds for major fungal expansion. Similar conclusions were drawn by Van Der Wiel (1982), who suggested that the increased frequency of K. deusta spores in the Hazendonk area (the Netherlands) most likely reflected the opening and degradation of deciduous forests, including oak woodlands, associated with human activity, which promoted the occurrence of injured and dying trees. A similar role of human activity in facilitating fungal infection was reported by Stolze and Monecke (2021) based on studies at Lough Arrow, Ireland. According to Kuhry (1985), a high and consistent occurrence of fungal spores in Quercus- and Tilia-dominated forests in Amtsven (northwestern Germany) may indicate not only anthropogenic processes, such as forest clearance, but also natural changes, including wood paludification and its gradual replacement by bog. This suggests that elevated K. deusta frequencies could be linked to woodland stress and decline resulting from a range of processes.
On the other hand, positive correlations have been reported during the Middle Holocene between increasing numbers of K. deusta spores and the course of tree pollen curves. For example, Shumilovskikh et al. (2015) recorded increased numbers of fungal spores in the presence of oaks at Borsteler Moor, Germany. Similarly, Hillbrand et al. (2014) showed a K. deusta curve resembling that of F. sylvatica, with both increasing concurrently, in the subfossil record from the vicinity of Lake Constance, Switzerland. Missiaen et al. (2017), in turn, observed such a relationship between the fungus and lime (Tilia) at a site in northwestern Belgium. Very high spore concentrations, positively correlated with pollen curves of Tilia spp., as well as Ulmus and Fraxinus, during the Holocene Thermal Maximum were reported at Gołębiewo, Poland (Latałowa et al., 2013). Interestingly, these tree taxa are considered more susceptible to infection than, for example, Quercus and Corylus (Baum and Schwarze, 2002). A particularly long-lasting, ca. 2700-year, positive correlation was observed between K. deusta and Tilia, indicating that the fungus did not have a negative effect on the lime population. Latałowa et al. (2013) have proposed that the infection may have contributed to the dominance of lime in the forest. Herbivorous animals, which used the open mire as a grazing area, may also have played a role by damaging trees through browsing and bark stripping, thereby facilitating fungal spread. Similar Middle Holocene evidence for the role of herbivores in exerting pressure on trees, facilitating fungal infection, comes from studies in the Lake Czarne region (Karpińska-Kołaczek et al., 2014). At the same time, human activity during the long-lasting Holocene Thermal Maximum, such as selective cutting of trees for wood quality, may have influenced forest composition, acting in parallel with herbivore pressure and other natural factors. In the case of lime, this pressure triggered intensive clonal reproduction (Latałowa et al., 2013). The fungus was also favoured by the climate during the Holocene Thermal Maximum, as K. deusta is known to have relatively high temperature requirements. In addition, summer drought caused by high temperatures was likely important for the successful infection of weakened Tilia trees. The effect of temperature on mycelial growth of K. deusta, experimentally studied by Cordin et al. (2021), showed optimum growth at 25°C, whereas growth was reduced at 10°C.
At the end of the Middle Holocene and during the first half of the Late-Holocene, a positive correlation between the presence of Kretzschmaria deusta and pollen of its potential host, in this case Fagus, was also observed at Lac de Moras in eastern France (Doyen and Etienne, 2017). Around 3000 cal yr BP, however, a decline in the abundance of K. deusta and several other wood- and plant-associated NPP morphotypes was recorded, accompanied by an increase in coprophilous spores, suggesting anthropogenic ecosystem transformations. A similar negative effect of woodland transformations on the occurrence of parasitic K. deusta was reported by Karpińska-Kołaczek et al. (2014) in the Lake Czarne region, northeastern Poland, from ca. 2400 ± 40 cal yr BP. These transformations included the expansion of Betula and Carpinus in areas created by human forest burning to enlarge the area for agricultural activity. Trees whose abundance declined at that time included Ulmus, Fraxinus, Picea, Tilia, and Corylus, probably reducing suitable hosts for K. deusta.
In contrast, a marked increase in K. deusta spores in beech-fir forests from 4070 cal yr BP was observed in the Beskid Wyspowy Mountains at Mogielica, southern Poland (Czerwiński et al., 2020). Here, the retreat of Fagus between 2630 and 2070cal yr BP did not lead to a decrease in spore abundance; on the contrary, percentages increased. This contrasts with observations at Lac de Moras (Doyen and Etienne, 2017) and Lake Czarne (Karpińska-Kołaczek et al., 2014). Tree damage caused by human activity, such as the removal of beech for livestock fodder, may have stimulated fungal infection, indicating that anthropogenic factors could act as a promoter for K. deusta. Unlike Tilia, however, beech may not have tolerated such damage well, resulting in its local retreat. Similarly, van Geel et al. (2013) reported a possible impact of K. deusta infections on the weakening of beech at Meerfelder Maar, Germany, reflected in a decline of Fagus pollen at 3285 varve yr BP. It is not excluded that Fagus may also have served as a host for the fungus at Saxnäs Mosse in southern Sweden, an area representing the northern distribution limit of this tree. Such a possible co-occurrence of both taxa may have persisted there for approximately 230 years.
In the Late-Holocene, as in the Middle Holocene, K. deusta was also associated with Quercus. Laine et al. (2010) observed a relationship between increases in K. deusta spores and Quercus pollen at a site in Burgundy, France. Szal et al. (2015) reported synchronous changes in the abundances of oak and the fungus in the top layer of the profile at Poganowo, northeastern Poland, suggesting that this fungus may have infected oaks more frequently than other trees. At this site, tree damage caused by livestock, which scraped bark from roots and lower trunks, could have promoted infections. High abundances of coprophilous fungi in the sediments indicate a dense population of livestock in the surrounding area.
Possible associations of K. deusta have also been documented in the Late-Holocene with the occurrence of other tree taxa. At Ferry Lane in England, an initial increasing trend was observed in the curves of fungal spores and Alnus pollen; subsequently, the spore peak coincided with a decline in this tree pollen (Waller and Grant, 2012). However, this pattern does not necessarily imply a parasitic causal relationship. The decline of alder may instead have resulted from tidal flooding in the estuarine environment of the River Thames, where the site is located. This interpretation is supported by the presence of standing dead trees preserved within the sediment. At El Payo, Spain, a decline in Betula was similarly correlated with the occurrence of K. deusta spores (Silva-Sánchez et al., 2016). The relationship between K. deusta and forest cover was further demonstrated at the Roman castellum Fectio in the Netherlands (van Den Bos et al., 2014), where a synchronous decline in fungal spores and pollen of potential tree hosts was recorded. This large-scale deforestation was reflected in fungal assemblages, which shifted from a community dominated by spores of the tree-associated parasite K. deusta to assemblages dominated by fungi associated with herbaceous vegetation and dung. The primary driver of the observed forest decline was anthropogenic activity, linked to the expansion of Roman army and surrounding settlements. In this context, K. deusta itself appears to have been indirectly affected by deforestation, experiencing a reduction in available tree hosts.
Palaeoecological data indicate a role for this fungus as a potential broadleaf tree pathogen (Everard et al., 2025), as correlations between fungal spores and deciduous tree pollen are commonly recorded in sediments (Pals et al., 1980). At the same time, it is less frequently associated with conifers (Dietre et al., 2012). However, this does not exclude conifer species from being suitable hosts for this parasite. For instance, Menozzi et al. (2010) suggested that K. deusta could have contributed to the abrupt decline of Abies in the Ligurian Apennines during historical times, while noting that other factors, such as additional fungal pathogens, insects, or even Viscum album, may have also played a role. In the Beskid Wyspowy Mountains, the decline of fir was similarly found to correlate temporally with increases in K. deusta spores (Czerwiński et al., 2020). Szal et al. (2015), based on statistical analyses, did not rule out a relationship between this ascomycete and Picea abies, reporting a distinct positive correlation between the pollen record of spruce and spores of this parasitic fungus. Experimental studies by Schwarze (2007) confirmed that such infections are possible in living spruce trees.
Only a few studies have addressed spore transport in the past. Evidence for the close presence of mixed forests based on the occurrence of K. deusta spores has been reported by Kołaczek et al. (2018) in the Western Carpathians. Similarly, Innes et al. (2024) inferred the presence of terrestrial vegetation close to the coring site at Roudsea Wood, England, based on the occurrence of coprophilous fungi and K. deusta spores. The interpretation of spores as a local signal is further supported by the type of the sediment at both sites, which was peat. On the other hand, Cugny et al. (2010) emphasized the need for caution when interpreting spore presence as an indicator of nearby forests. In modern datasets, ascospores have been found at sites more than 100 m from beech forests, suggesting that long-distance transport by wind or water is possible. The potential role of water, driven by heavy rainfall, in transporting large quantities of K. deusta spores is supported by subfossil evidence from Meerfelder Maar, Germany (van Geel et al., 2013). Short-term peaks in K. deusta abundance may reflect rainstorms events that could have caused erosion of the litter layer and subsequent transported of material from the forest floor into the lake via stormwater runoff and strong winds. Additionally, tannins in the litter of the beech forest surrounding Meerfelder Maar during the Late-Holocene inhibited understory growth, which facilitated the washing of litter containing spores into the lake during rainfall events. The steep slopes around the lake, covered by beech forest, further enhanced surface runoff. It is also possible that heavy rains, by damaging tree trunks, contributed to fungal infection. An increase in K. deusta influx at Lake Mondsee, Austria, was likewise suggested to possibly indicate erosion, which may have been related to enhanced surface runoff during the Late-Holocene (Schubert et al., 2025).
Conclusions
The compiled fossil record shows that Kretzschmaria deusta was widely distributed across Europe throughout the Holocene, with the majority of records originating from the Middle and Late-Holocene. The relatively sparse Early Holocene occurrences contrast with the markedly broader spatial and temporal presence of spores after ca. 8.2 ka BP, suggesting an expansion linked to the development of mature deciduous forests.
Kretzschmaria deusta functioned throughout the Holocene primarily as an opportunistic pathogen, exploiting trees weakened by climatic stress, herbivory, geomorphological processes, or human activities such as woodland exploitation, free-ranging livestock, and forest clearance. Elevated spore abundances may therefore be interpreted as reflecting periods of increased tree damage and reduced forest resilience rather than primary forest decline. Host-tree responses varied among taxa, reflecting differences in physiological tolerance, regenerative capacity, and sensitivity to repeated disturbance.
Interpretations based on ascospore records are subject to uncertainty related to processes including differential dispersal, transport, and preservation. In particular, spore abundances may be influenced by non-local inputs and depositional dynamics, which can complicate direct ecological attribution. Therefore, the results presented should be regarded as reflecting environmental associations rather than direct causal relationships.
In lime-dominated forests, high abundances of K. deusta spores coincided with stable or even expanding populations of Tilia. This suggests that lime recovered from repeated infection and related stress mainly via vigorous clonal regeneration following browsing or mechanical damage. In contrast, Fagus and Abies appear to have been more sensitive to sustained fungal pressure, with increases in K. deusta spore abundances commonly corresponding to phases of host decline, particularly where anthropogenic disturbance intensified.
Climatic factors further shaped the distribution and abundance of K. deusta during the Holocene. Associations between high spore concentrations and the Holocene Thermal Maximum indicate that warmer temperatures and episodic summer droughts enhanced fungal growth and facilitated infection of physiologically stressed trees. Experimental evidence demonstrating higher mycelial growth rates at elevated temperatures is consistent with these palaeoecological observations.
The predominantly local dispersal of ascospores suggests that fossil records of K. deusta generally reflect nearby woodland conditions. However, episodic long-distance transport by wind or water, particularly during heavy rainfall events and enhanced surface runoff, may have contributed to short-term peaks in spore abundance in lacustrine sediments.
Overall, the Holocene palaeoecological record identifies K. deusta as a sensitive indicator of woodland stress and disturbance rather than a direct driver of forest decline. The patterns observed during the Holocene closely resemble processes documented in modern temperate forests, where ongoing climatic warming, prolonged summer droughts, and increasing human pressure are associated with reduced tree resilience. Integrating fossil and modern data may allow improved understanding of the long-term dynamics of tree-fungus interactions and suggests that K. deusta may be linked to forest decline under conditions of climatic warming and intensified human impact.
Footnotes
Author contribution(s)
Funding
The author received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.