
Abstract
A multiproxy modern surface sediments sample analogue including pollen, non-pollen palynomorphs (NPPs), Total organic carbon/Total nitrogen (TOC/TN), and stable carbon isotopes (δ13C values) were generated from the Bakhira Lake, Central Ganga Plain (CGP) to delineate the modern climate-vegetation relationship and human-environment interactions. Pollen data reveal a dominance of non-arboreals (67.4%) over arboreals (32.6%) along the lake margins, with increased arboreal representation (e.g. Acacia, Madhuca indica, Anacardiaceae, Trewia nudiflora, Peltophorum, Arecaceae) in the lake centre. This reflects both land-use intensity and pollen production biases. The assemblage indicates a mixed deciduous forest under warm, humid conditions. The occurrence of marker pollen taxa, such as Cerealia, Alternanthera, Asteraceae, Chenopodiaceae/Amaranthaceae (Cheno/Am), Brassicaceae and NPPs along with coprophilous fungi assemblage (viz. Sordaria, Podospora, Delitschia, and Ascodesmis) shows the anthropogenic activities and intensively-grazed conditions over the sampling site. The relatively higher δ13C values (−19.7 to −21.7‰) indicate C3 dominated to mixed C3−C4 vegetation input to the sedimentary organic matter at the lake centre. The NW and SE flanks, together with cropland, have relatively lower δ13C values ranging from −21.2‰ to −28.9‰, except for the sample number S23 (−19.3‰), indicating a higher contribution of organic matter from C3 vegetation. Ordination analysis of pollen taxa distributions in surface sediments samples revealed correlations among sampling locations, modern pollen deposition, and associated environmental factors. This multiproxy modern dataset provides a robust baseline for interpreting Holocene palaeoecological and palaeoclimatic dynamics across tropical lacustrine systems in the central Indian subcontinent.
Keywords
Introduction
Reconstructing past vegetation dynamics and climate variability relies fundamentally on understanding the relationship between modern pollen assemblages and extant vegetation. Modern pollen-vegetation calibration studies provide the essential interpretative framework for distinguishing between climatic, hydrological, and anthropogenic signals preserved in sedimentary archives. However, pollen assemblages preserved in sediments do not represent vegetation in a simple linear manner, as they are influenced by differences in pollen productivity, dispersal, depositional processes, and preservation biases (Barboni and Bonnefille, 2001; Barreto et al., 2024; Bunting et al., 2004; Morales-Molino et al., 2020; Sugita, 2007a; Trivedi et al., 2024; Xu et al., 2016). Establishing robust modern analogues is therefore essential, particularly in monsoon-dominated tropical regions where vegetation structure is sensitive to hydroclimatic variability and human land-use intensity. In the Indian subcontinent, pollen-based reconstructions have significantly advanced our understanding of monsoon variability and vegetation dynamics (Dubey et al., 2018; Prasad et al., 2014; Saxena et al., 2017; Tripathi et al., 2023). However, modern calibration studies from the Central Ganga Plain (CGP) remain comparatively limited, despite the regions climatic sensitivity and long history of human occupation. The CGP, specifically the Bakhira Lake area, has a substantial collection of sedimentary records, rendering it an optimal site for high-resolution palaeoclimatic research. This study incorporates modern analogues using surface sediments pollen, non-pollen palynomorphs (NPPs), and stable carbon isotope data to interpret past climae conditions and vegetation dynamics. Surface sediments pollen analysis is fundamental to palaeoecological research, offering insights into past vegetation and climate by examining pollen grains preserved in sediments. Pollen analysis indicates the composition of contemporaneous vegetation during deposition, providing direct evidence of past plant communities and related climatic conditions (Anupama et al., 2000; Birks and Birks, 1980; Bonnefille et al., 1999; Flenley, 1973; Fraegri and Iverson, 1975). This approach has demonstrated its utility in reconstructing monsoonal variations and vegetation changes in tropical regions. Prior investigations in this area have illustrated the efficacy of pollen analysis in delineating historical variations in SW monsoon strength and its effects on indigenous vegetation (Farooqui and Sekhar, 2011; Kar and Quamar, 2019; Kumar et al., 2022; Mandaokar et al., 2008; Saxena et al., 2017; Tripathi et al., 2023; Trivedi et al., 2014, 2016, 2019).
Recent advances in palaeoecology emphasise the integration of non-pollen palynomorphs (NPPs) and geochemical proxies with pollen data to improve environmental interpretation. NPPs, including coprophilous fungal spores, algae, and soil derived fungal taxa, provide independent evidence of grazing intensity, hydrological variability, soil disturbance, and land-use change (Basumatary et al., 2024; Birks, 2019; Ejarque et al., 2011; Ekblom and Gillson, 2010; Lee et al., 2022; Shumilovskikh et al., 2021; van Geel and Aptroot, 2006; Zhang et al., 2025). The integration of these microfossil indicators with pollen data strengthens the reconstruction of human-environment interactions in lacustrine systems (Jun et al., 2024; Kiage, 2025; Kramer et al., 2010; Leroy et al., 2021; Montoya et al., 2010). Along with pollen and NPPs, the study of stable carbon isotopes (δ13C values) in organic matter associated with sediments (SOM) serves as a reliable proxy for past vegetation types and carbon cycling processes, aiding in the comprehension of terrestrial ecosystems (Limaye et al., 2007; McAndrews and Turton, 2010; Meyers, 1994). The δ13C values in the vegetation vary based on the photosynthetic pathways (C3 and C4 vegetation) (Agrawal et al., 2012; Basu et al., 2015; Deines, 1980; Kohn, 2010; O’Leary et al., 1981). Environmental variables such as precipitation and temperature affect carbon isotope fractionation during photosynthesis in plants, hence influencing their δ13C values. The δ13C values for C3 plants vary from −34 to −22‰, with an avg. of −27‰; for C4 plants, the range is −16–−10‰, avg. −12.5‰ (Agrawal et al., 2012; Basu et al., 2015; Cerling et al., 1997). The difference in δ13C values between C3 and C4 vegetation offers a significant opportunity to reconstruct vegetation community dynamics. C3 vegetation, prevalent in cooler, wetter habitats, display distinct δ13C values relative to C4 plants, which flourish in warmer, drier conditions (Cerling and Quade, 1989; Farquhar et al., 1989). Examining δ13C values in surface sediments from the CGP helps elucidate changes in vegetation composition and associated climatic conditions (Kumar et al., 2022; Sharma et al., 2004; Tripathi et al., 2023; Trivedi et al., 2024). This isotopic approach, in conjunction with pollen and NPPs data, offers a robust framework for reconstructing past climate variations (Chen et al., 2024; Kumar et al., 2022; Meyers, 1994; Sage, 1999).
Despite these methodological advances, comprehensive multiproxy modern analogue studies combining pollen, NPPs, and δ13C values from surface sediments are scarce in the CGP. In particular, oxbow lake systems such as Bakhira Lake remain underexplored, even though they are highly sensitive to hydrological fluctuations, catchment processes, and land-use change. The present study addresses this gap through a multiproxy investigation of surface sediments collected along transect across Bakhira Lake and adjacent croplands in the CGP. By integrating pollen, NPPs, and δ13C values, we aim to establish a modern ecological baseline for interpreting fossil records from the region. The specific objectives of this study are to: (a) delineate the modern pollen-vegetation relationship across lacustrine and adjoining cropland environments and characterise associated organic matter sources (C3–C4 vegetation) using δ13C values; (b) evaluate the ecological significance of NPPs assemblages, including coprophilous fungal spores and algal remains; and (c) investigate human-environment interactions and their impact on land cover. To achieve these objectives, the present study addresses the following key research question: (i) how effectively do modern multiproxy signals integrating surface pollen assemblages, NPPs, and δ13C values reflect contemporary vegetation patterns, hydrological conditions, and anthropogenic land use around Bakhira Lake, and (ii) how can these relationships improve the interpretation of Holocene palaeoclimatic and palaeoecological records from the monsoon-dominated Indian subcontinent? By developing an integrated modern multiproxy framework, this study provides a robust baseline for future palaeoclimatic and palaeoecological investigations in the CGP and contributes to a better understanding of climate-vegetation-human interactions in monsoon-dominated lacustrine systems of the Indian subcontinent.
Study area
The Ganga plain forms part of the Indo-Gangetic foreland basin and composed predominantly of thick Quaternary alluvial deposits overlying Precambrian basement rocks and derived mainly from erosion of the Himalayan hinterland and the Peninsular Indian shield (Singh, 1996; Valdiya, 1976). These sediments consist of alternating layers of sand, silt and clay deposited through repeated phases of channel migration, overbank flooding and floodplain aggradation. The present gently sloping and low-relief topography of the plain has also been influenced by neotectonic activity along structural features such as the Faizabad ridge (Singh, 2001). Geomorphologically, the CGP represents a tectonically active peripheral foreland basin characterised by a variety of fluvial landforms including channel deposits, muddy overbank sediments, crevasse splays, floodplains, meander scars, palaeochannels, point bars and numerous oxbow lakes formed through lateral channel migration (Singh, 2000; Srivastava et al., 2003). The present floodplain landscape is largely shaped by meandering river systems whose migration has produced several abandoned channels, wetlands and oxbow lakes across the basin. Bakhira Lake (26°54’ 31.0”N; 83°6’22.1”E) is a shallow, perennial oxbow lake located west of the Rapti River in the CGP of northern India (Figures 1 and 2). The lake covers an area of ~29 km2 and is recognised as the largest natural wetland in Uttar Pradesh and a Ramsar site (2022). It formed through the lateral migration and subsequent cut-off of a meander of the Rapti River, and forms part of the Ghaghara-Rapti plain, representing a typical floodplain wetland system of the Terai region. The surrounding landscape comprises agricultural fields, grasslands, scrub vegetation and aquatic habitats typical of the Terai region. The soils in the study area are primarily recent alluvial soils, broadly classified as Entisols and Inceptisols, which are generally silty to clayey near the lake margins and relatively loamy in cultivated interfluve areas (National Bureau of Soil Survey and Land Use Planning [NBSSLUP], 2002; Srivastava et al., 2015). Such oxbow lakes within the CGP act as natural sediment traps and provide valuable archives for reconstructing past environmental and climatic changes in the monsoon-dominated landscape.
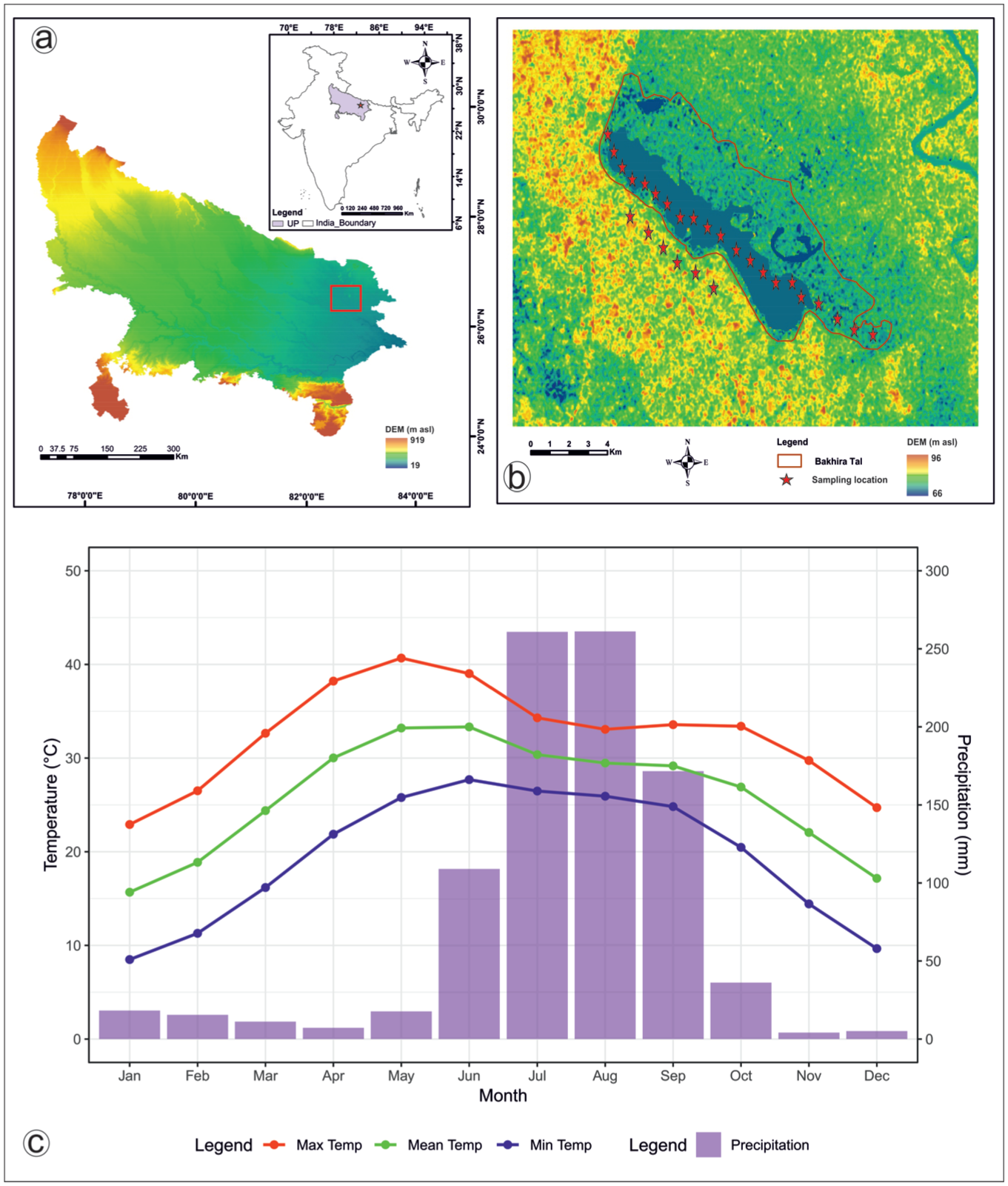
Study area location and sampling sites: (a) DEM showing Uttar Pradesh and the location of Bakhira Lake in the Central Ganga Plain (red rectangle), (b) DEM-based detailed map of Bakhira lake, illustrating sampling locations (red stars), and (c) plot showing the monthly climatology of avg. minimum surface air temperature, avg. mean surface air temperature, avg. maximum surface air temperature and precipitation 1991–2020; Uttar Pradesh, India. Vegetation.

Field photographs depicting (a) sample collection process, (b) surrounding marshy vegetation, (c) open water body, (d) cropland area adjacent to the lake, and (e and f) diverse aquatic and marsh vegetation habitats across the lake.
The study area lies within the subtropical deciduous forest belt of northern India and represents a mosaic of moist deciduous, dry deciduous, scrub, grassland, and wetland vegetation types (Champion and Seth, 1968; Farooqui and Sekhar, 2011; Trivedi et al., 2019). According to the forest classification of Champion and Seth (1968), the broader region of the CGP and adjoining Terai supports both tropical moist deciduous forests characterised by species such as Shorea robusta (Sal) and tropical dry deciduous forests, which include Tectona grandis along with other associated taxa. Although large continuous forest tracts are now fragmented due to agricultural expansion, remnants of these forest types persist in patches within the landscape. At present, the vegetation surrounding Bakhira Lake exhibits a heterogeneous pattern dominated by scrub forests, open grasslands, and marshy habitats. The important tree species such as Acacia chundra, Acacia nilotica, Adina cordifolia, Ailanthus excelsa, Bauhinia variegata, Butea monosperma, Dalbergia sissoo, Emblica officinalis, Holoptelea integrifolia, Madhuca indica, Moringa oleifera, Syzygium cumini, and Terminalia bellirica. Though often sparsely distributed, these taxa represent remnants of the broader deciduous forest vegetation of the region. Shrub communities include Abrus precatorius, Adhatoda vasica, Carissa opaca, Indigofera himalayensis, Rhamnus sp., Strobilanthes angustifrons, Vitex negundo, and Ziziphus mauritiana, contributing to structural complexity and providing habitat for fauna. Ground vegetation is dominated by grasses and herbaceous taxa, with significant representation from Asteraceae, Cheno/Am, Euphorbiaceae, Ranunculaceae, and Solanaceae. These open vegetation elements reflect both natural edaphic conditions and anthropogenic land-use intensity. Marshy habitats near the lake margins are characterised by Cyperaceae and Polygonaceae, along with semi-aquatic reed-swamp species such as Phragmites karka and Typha latifolia, which form dense stands. Floating and submerged macrophytes include Jussiaea perennis, Nelumbo nucifera, Nymphoides cristata, Potamogeton cristatus, and Trapa natans. A dense cover of Eichhornia crassipes, an invasive aquatic species widely distributed across the Ganga Plain, frequently blankets parts of the lake surface. Overall, the vegetation of the study area represents a dynamic interaction between tropical deciduous forest elements, wetland communities, grasslands, and agricultural landscapes (Supplemental figures 1–3). This heterogeneous vegetation mosaic provides the ecological framework necessary for interpreting modern pollen and NPPs assemblages and for reconstructing past vegetation and climate variability in the CGP.
Climate
The study area experiences a humid subtropical monsoon climate controlled primarily by the Southwest (SW) monsoon (Chauhan and Bera, 1990; Trivedi et al., 2019). According to Köppen’s climate classification system (Köppen and Alt, 1936) and the Köppen-Geiger climate classification (Geiger, 1961), the region falls under the Cwa category, characterised as a humid subtropical climate with a dry winter and hot summer. The monsoon season extends from mid-June to mid-September and contributes ~75% of the annual rainfall, which avg. 100–120 cm. Western disturbances bring occasional winter rainfall and hailstorms during January–February. Winter season (November–February) are cool, with avg. minimum and maximum temperatures of ~7.6°C and 21°C, respectively and occasional lows of 2°C–3°C in December–January. Spring (March–mid-April) is dry and mild, with temperatures of 15°C–25°C. Summer season (mid-April–June) are hot, with mean temperatures of ~27°C–32.5°C and extremes up to 46°C in June. The monsoon period is warm and humid, followed by autumn (October–early November), which is marked by declining rainfall and moderate temperatures (20°C–30°C; Figure 1 and Supplemental figure 4).
Material and methods
Samples: Collection, extraction and identification of pollen/non-pollen palynomorphs
A total of 29 surface sediment samples were collected for multiproxy analysis. To examine the modern pollen-vegetation relationship, we collected 23 samples from a linear transect and 6 samples from the cropland field at intervals of 500 and 200 m, respectively (Figures 1 and 2). To understand how the dominant forest elements are cumulatively represented in the regional vegetation, we designed a sampling strategy using linear transects and adjoining cropland areas. The transect follows the hydrological gradient from the northwest flank, through the central basin, to the southeast flank of Bakhira Lake. Modern sediments that contain preserved pollen and spores help understand the vegetation assemblage, enabling scientists to help changes in the climate-vegetation equilibrium over time. Samples were oven-dried at 50°C overnight. Subsequently, 10 g of each sample was treated with a 10% aqueous potassium hydroxide (KOH) solution and boiled to remove humic substances and facilitate the disaggregation of pollen and spores from the sediment matrix. This was followed by treatment with 40% hydrofluoric acid (HF) to remove siliceous components. The standard method of acetolysis outlined by Erdtman (1943) uses an acetolysis mixture (9:1 ratio of acetic anhydride and concentrated sulphuric acid) for removing cellulose and humic debris, while fine sieving is used to remove clay-sized particles. Finally, the processed residues were stored in a 50% glycerine solution prior to microscopic examination.
Palynomorphs identification and counting were carried out using a Leica DM3000 LED Digital Microscope. Photographs and observations were made at 40X/0.80 CORR magnifications (Figures 3 and 4, and Supplemental figure 5). Conventionally, ∼200–305 pollen grains per sample were counted that are considered as total palynomorph count. Pollen percentages were calculated on the basis of the total pollen sum (TPS), which includes arboreal and non-arboreal terrestrial pollen taxa, while aquatic and marsh taxa were excluded from the TPS (Supplemental figure 6). The pollen taxa were categorised into arboreal taxa (trees and shrubs), non-arboreal taxa (terrestrial, anthropophytes/cultural, wetland and aquatics), extra-regional taxa, and ferns allies in the pollen spectra. NPPs were counted from the same pollen slides, with 100–288 NPPs counted per sample, excluding pollen grains. NPPs percentages were calculated relative to the total pollen sum. NPPs were categorised into algal remains, coprophilous fungi, non-coprophilous fungi, zoological remains (zooplankton), and unidentified morphotypes (Figures 5 and 6). Identification of the palynomorphs was achieved by comparison with reference pollen slides available at Birbal Sahni Institute of Palaeosciences herbarium as well as the pollen and fungal spore photographs from various published literature (Basumatary and McDonald, 2017; Chauhan and Bera, 1990; Nayar, 1990; Shumilovskikh et al., 2021; Trivedi et al., 2015, 2019; van Geel et al., 2011; van Geel and Aptroot, 2006). TILIA and TILIA graph software (Grimm, 1992) was used for the preparation of pollen, NPPs spectra and cluster analysis (CONISS). The cluster analysis helped to delineate compositional groupings corresponding to the NW flank, Centre, SE flank, and cropland site.
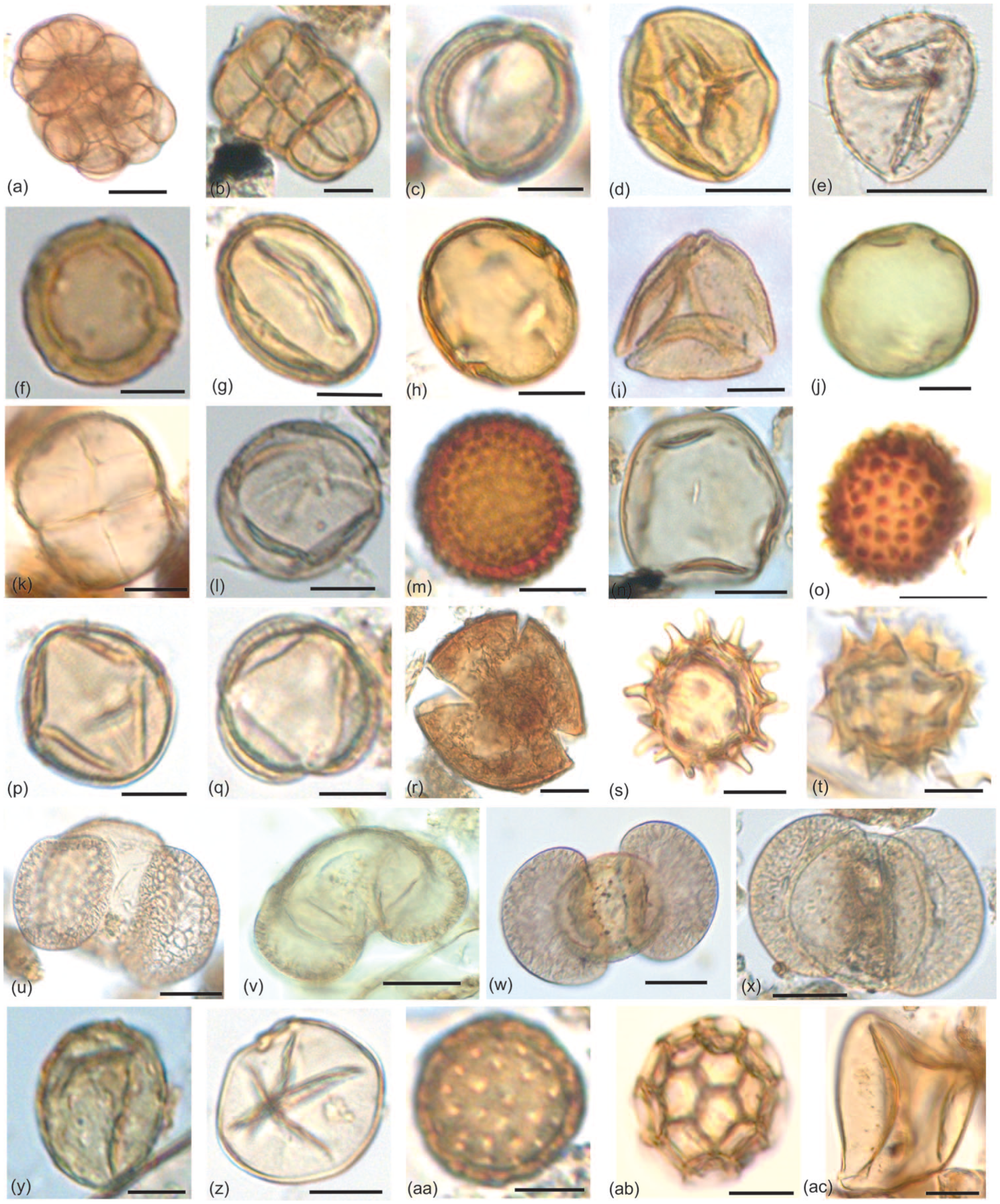
Key pollen taxa recovered from the surface samples of Bakhira Lake: (a and b) Acacia sp., (c) Aegle marmelos, (d) Anacardiaceae, (e) Arecaceae, (f) Holoptelea integrifolia, (g) Terminalia sp., (h) Madhuca indica, (i) Schleichera oleosa, (j) Trewia nudiflora. (k) Fabaceae, (l) Dodonaea, (m) Croton, (n) Poaceae, (o) Malvaceae, (p) Solanum, (q and r) Convolvulaceae, (s and t) Asteraceae, (u to x) Pinus sp., (y) Juglans, (z) Cerealia, (aa) Cheno/Am, (ab) Alternanthera, (ac) Cyperaceae.
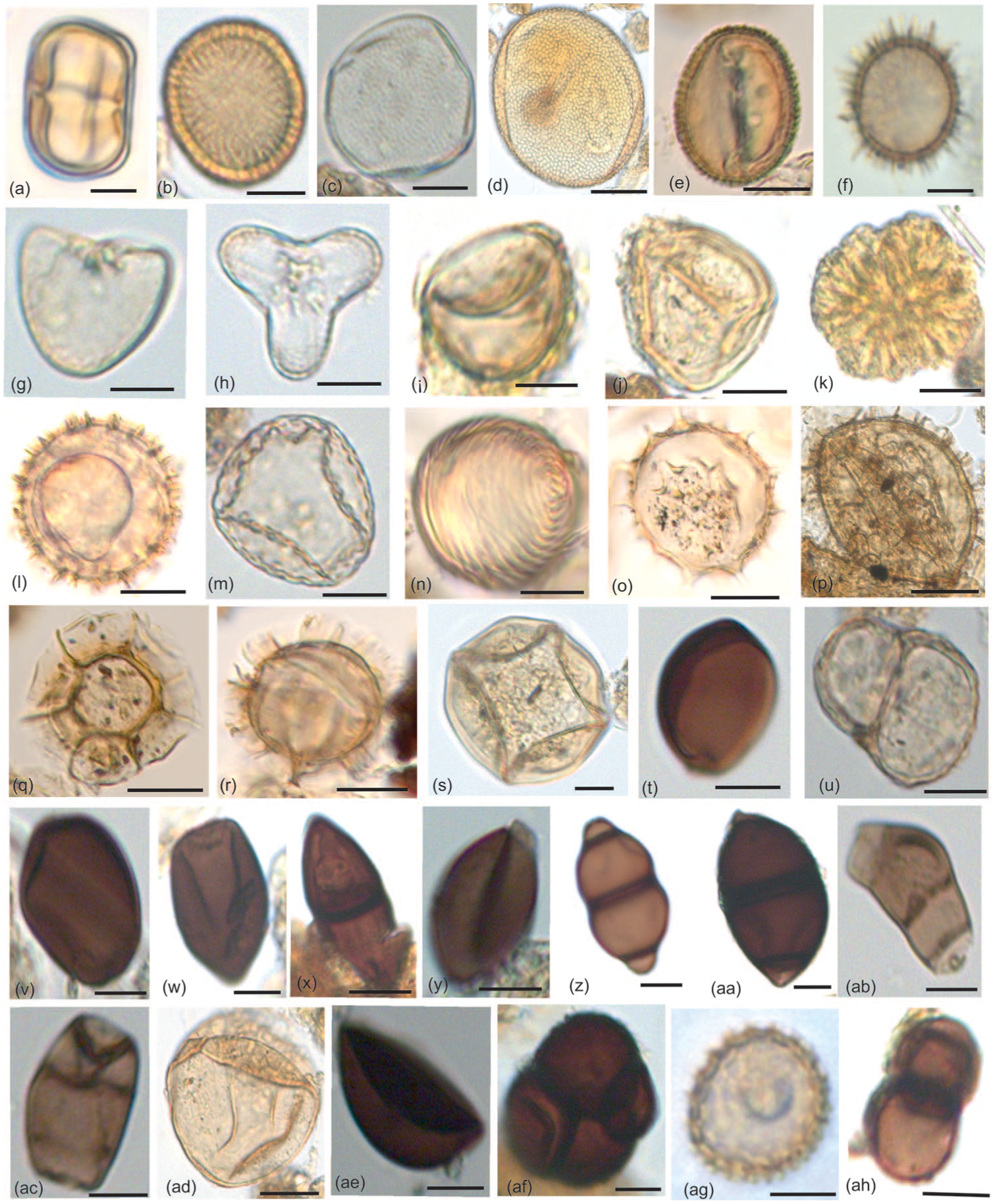
Key palynomorphs recovered from the surface samples of Bakhira Lake: (a) Polygonum plebeium, (b) Polygonaceae, (c) Typha, (d) Iris pseudacorus, (e) Cressa cretica, (f) Lemna, (g and h) Adiantum sp., (i) Fern (monolete; j) Fern (trilete), (k) Botryococcus, (l) Debarya zygospore, (m) Zygnema zygospore, (n) Concentricystis, (o and r) Dinoflagellates cysts, (s) Mougeotia type (zygospore), (t) Sordaria, (u) Delitschia, (v and w) Podospora, (x) Cookeina, (y) Arthrinium, (z)–(aa) Valsaria, (ab)–(ac) Curvularia, (ad) Rhizopus sp., (ae) Xylariaceae, (af) Spegazzinia tessarthra, (ag) Laccaria, (ah) Diplodia.

Percentage pollen diagram showing the relative abundance of pollen taxa in surface samples from Bakhira Lake, CGP.
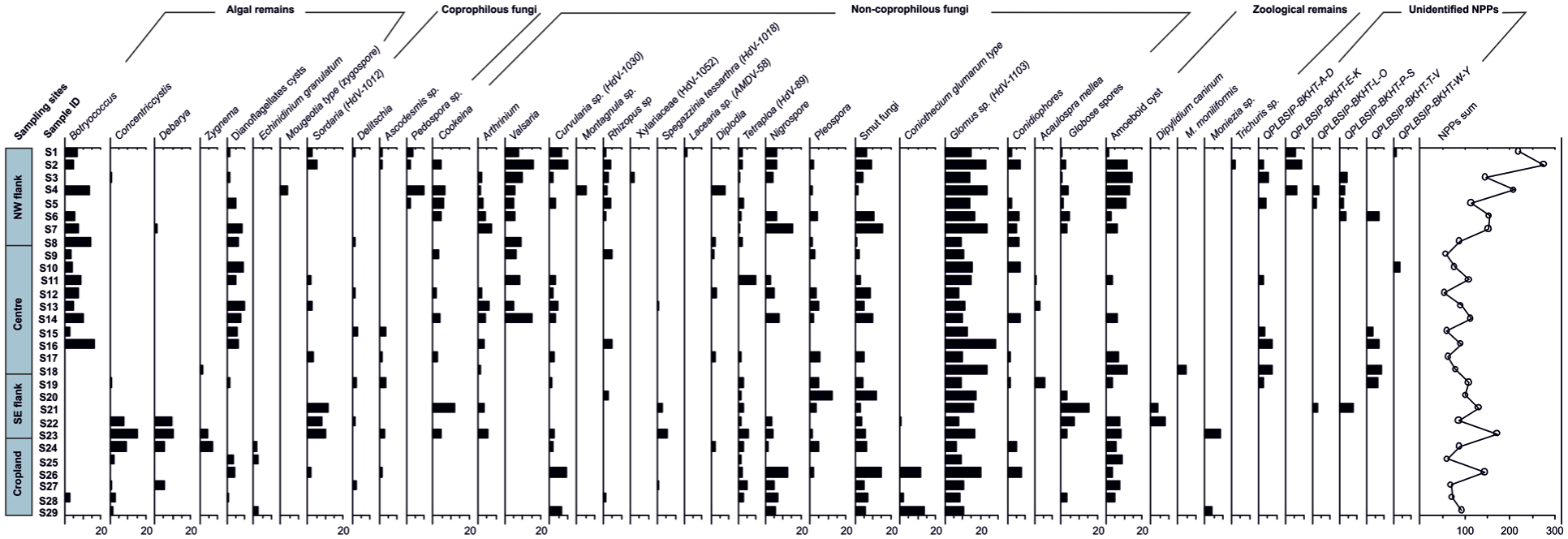
Percentage diagram of non-pollen palynomorphs recorded from surface samples of Bakhira Lake, CGP.
Sample preparation and measurements of total organic carbon content (TOC), total nitrogen (TN) and carbon isotope (δ13C values) of surface sediments
To analyse the δ13C values in organic matter associated with sediments (SOM), individual samples were powdered and thereafter decarbonated using 0.5 N HCl thrice to remove carbonate. Acid and soluble salts were removed by centrifuging (∼3000 rpm) 5-6 times with milli-Q water. After centrifuging, the decarbonated samples were dried in oven at about 60°C temperature overnight. The dried samples were crushed to make fine powder. Powdered sample of 15–25 mg was loaded into a tin capsule and introduced into pre-filled and conditioned reactor of Elemental Analyser (Flash EA 2000 HT) through an auto sampler for isotopic (δ13C values) analysis, following the sample preparation protocol outlined in Agrawal et al. (2015). Total organic carbon (TOC) in the decarbonated samples and total nitrogen (TN) in the bulk samples were calculated from the peak area obtained from the sum of the integrated m/z 44, 45 and 46, and 28 and 29 signal measured in the CFIRMS, respectively (Jensen, 1991). Analytical reproducibility, checked by repeat measurements of selected samples, ranged between 0.1‰ and 0.4‰ for δ13C value. Similarly, routine repeat measurements of TOC content show 0.02%–0.5 % variability, depending on the organic carbon content in the samples. Stable isotope analyses of all samples were conducted using isotope ratio mass spectrometry (IRMS) at the Birbal Sahni Institute of Palaeosciences (BSIP), Lucknow.
Ordination analysis
Ordination of surface palynological samples and associated pollen taxa is essential for the quantitative interpretation of pollen assemblage data. This approach also aids in identifying key correlations between pollen and non-pollen palynomorphs assemblages and environmental variables such as vegetation composition, climatic conditions, and land cover (Birks and Gordon, 1985; Chaloner, 1975). To determine the appropriate ordination method, a preliminary Detrended Correspondence Analysis (DCA) was conducted. The analysis yielded a gradient length of less than 3 standard deviations (SD) along the first axis, indicating a relatively short ecological gradient. As such, a linear response model was deemed suitable, and Principal Component Analysis (PCA) was applied to discern the primary factors influencing the distribution of pollen taxa and sample composition. Prior to ordination, the pollen percentage data were subjected to Hellinger transformation, which reduces the influence of dominant taxa and renders compositional data suitable for Euclidean-based ordination methods such as PCA. In the ordination diagram, the length of each pollen taxa vector represents the degree to which its variability is captured within the reduced-dimensional space. PCA allows for the assessment of variability among depositional settings and effectively captures linear relationships present in the dataset. All numerical analyses were conducted using R statistical software (ver.4.3.2, ucrt R foundation, 2023). Ordination analysis was performed using the vegan package (Oksanen et al., 2012).
Results
Pollen assemblages
The pollen spectra indicate that arboreal taxa account for 32.6% of the total assemblage, whereas non-arboreal taxa dominate with 67.4% (Figure 5; Supplemental Table 1). In NPPs; ferns allies, algal remnants, coprophilous fungi, non-coprophilous fungi were documented with considerable frequency, while zooplankton occurrences were noted less frequently (Figure 6; Supplemental Table 2). Additionally, unidentified fungal spores and cysts were reported. Arboreal pollen (AP) mostly consists of Acacia sp., Aegle marmelos, Madhuca indica, Pinus sp., Trewia nudiflora, Terminalia sp., Arecaceae, Holoptelea integrifolia, Croton, Aspidopteris, Dodonaea. Conversely, non-arboreal pollen (NAP) comprises Poaceae, Cerealia, Alternanthera, Convolvulaceae, Cheno/Am, Solanum, Asteraceae, Polygala, Cyperaceae, Polygonaceae, Potamogeton, Typha, Iris pseudacorus, Cressa cretica. Our study region is categorised as the NW flank, Centre, SE flank, and Cropland based on sample sites and recovered palynological assemblages (Figure 5 and Table 1).
The δ13C (VPDB‰) and TOC/TN values in the lake sediments.
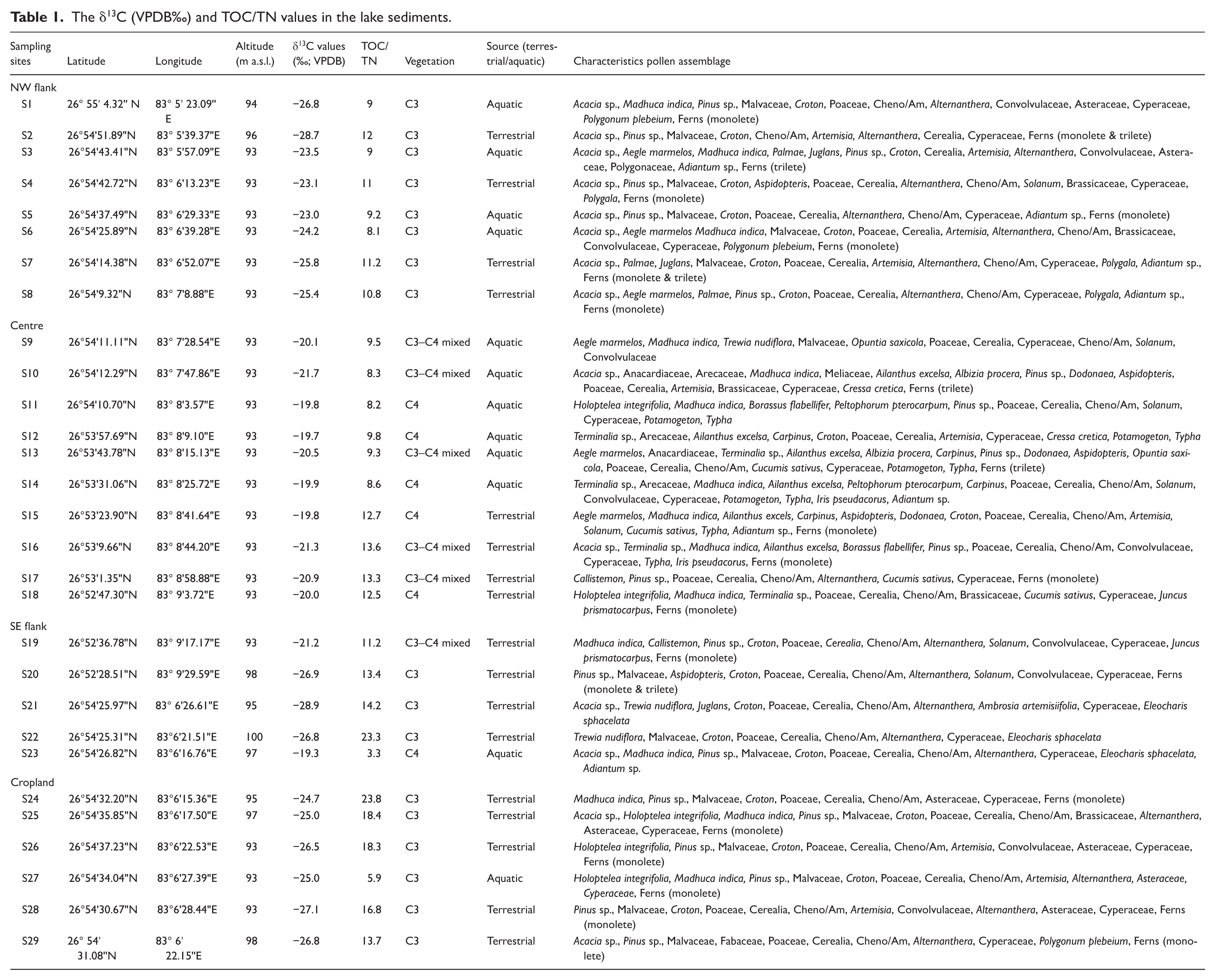
NW flank site (Acacia-croton-poaceae-alternanthera-cerealia-cheno/am-cyperaceae assemblage)
This location is predominantly represented by non-arboreal (61.6%) compared to arboreal (38.4%). Among arboreal; tree taxa such as Acacia sp. (7%−23.5%) are the dominant taxon followed by extra-regional taxa viz. Pinus sp. (3%−14.5%). Other arboreal taxa are recorded in moderate values such as Aegle marmelos (5.2%−7.4%), Palmae (4.4%−6.8%), Holoptelea integrifolia (5.6%) followed by Juglans (5.2%−5.7%), Anacardiaceae (2.4%−5.5%), Meliaceae (4.5%), and Arecaceae (3%). Shrubby elements are represented by Croton (2.4%−12.3%), and Aspidopteris (2.4%−8.6%). Among the non-arboreal, terrestrial herbs such as Poaceae (5.2%–21%) is the predominant taxa, followed by Asteraceae (4.2%−7.3%), Malvaceae (3.8%−7.3%), and Convolvulaceae (2.4%−4.7%). Anthropophytes/cultural taxa such as Alternanthera (6.6%−20.5%), Cerealia (3.5%−18.7%), and Cheno/Am (5.2%−15%) are the predominant taxa, followed by Brassicaceae (4.2%−8.2%). Other anthropophytes are recorded in moderate values viz., Artemisia (2.4%−6.3%) followed by Solanum (5.2%). The wetland taxa include Cyperaceae (4.5%−17.6%) is recorded in abundance followed by Polygonaceae (4.7%), Polygala (1.6%−3.8%) and Polygonum plebeium (1%−3.3%). The aquatic is represented by Lemna accounting 2.3%−4.3%. Ferns allies, including Adiantum sp., monolete, and trilete spores, were found in all samples, with an avg. frequency of 11%.
Centre site (Madhuca indica-acacia-poaceae-cerealia-cyperaceae assemblage)
This site is characterised by relatively increasing arboreal taxa (49.5%) and decreasing non-arboreal taxa (50.5%) compared to other sites. Among the arboreal, tree taxa such as Madhuca indica (8.3%−20.6%), Acacia sp. (6.6%−20.4%), Peltophorum pterocarpum (10%−10.8%), Arecaceae and Anacardiaceae (5.1%−10.8% each) are the predominant taxa followed by Borassus flabellifer (8.4%−9.7%), Trewia nudiflora (6.4%−9.1%), Aegle marmelos (5.7%−8.4%), Callistemon (8.2%), and Holoptelea integrifolia (7%) are recorded at moderate values. Albizzia procera (4.5%−6.1%), Terminalia sp. (6%), and Ailanthus excelsa (4.8%), are represented in low values. Extra-regional taxa viz. Carpinus (6.2%−10.7%), and Pinus sp. (4.5%−9.6%) are also recorded in appreciable proportion. Croton (7.3%−15.3%), and Aspidopteris (10.2%−11.22%) are the dominant shrubs followed by Dodonaea (5.6%−9%), and Opuntia saxicola (5.4%−8.3%). Among NAP, terrestrial herbs such as Poaceae (7.5%−19.8%), Malvaceae (4.2%−14.2%), are the dominant taxa followed by Convolvulaceae (7%−9%). Anthropophytes/cultural taxa such as Cerealia (6.2%−20.4%), Cucumis sativus (6%−18%), Cheno/Am (5%−16.2%), and Solanum (6.2%−10.8%), are the dominant taxa followed by Artemisia (5.3%−8.4%), Ambrosia artemisiifolia (5%−6.8%), Alternanthera (7.5%), and Brassicaceae (4.6%−6%) are recorded moderately. The dominant wetland taxon is Cyperaceae (2.8%–15%) followed by Cressa cretica, Polygala and Juncus prismatocarpus (3% each). Aquatics such as Potamogeton (3.6%−11.5%), Iris pseudacorus (6.2%−11.4%), Typha (4.5%−11.3%), are the dominant taxa whereas, Lemna (1.2%–2%) represented in low values. Ferns allies (Adiantum sp., monolete, and trilete) occurred in all samples with an avg. frequency of 6.5%.
SE flank site (Acacia-malvaceae-poaceae-cerealia-cheno/am-alternanthera assemblage)
This location is predominantly occupied by non-arboreal species (67.8%) compared to arboreal taxa (32.2%). Among arboreal, tree taxa such as Acacia sp. (6.7%−19.6%) and Ailanthus excelsa (11%) are the dominant taxon followed by Pinus sp. (9.5%–10%) and Juglans (7.1%). Terminalia sp. (6%), Callistemon (5.6%) and Anacardiaceae (5%−5.5%), is represented in low values. Shrubs taxa include Croton (6%−11.7%) is the dominant taxon followed by Aspidopteris (7.4%). Among non-arboreal taxa, the major terrestrial taxa include Malvaceae (7%−19.5%) and Poaceae (9%−17%), followed by Convolvulaceae (5%−5.6%), and Asteraceae (5.4%) are recorded in low values. Anthropophytes/cultural taxa such as Cerealia (11.3%–21%), Cheno/Am (8.6% to 20.2%), Alternanthera (7.4%–20%), are recorded in high values followed by Solanum (5%−8.3%) and Ambrosia artemisiifolia (8.4%). The dominating wetland taxa are Cyperaceae (5%−19%), followed by Eleocharis sphacelata (3.4%−9.4%) and Juncus prismatocarpus (1%) is represented in low values, whereas aquatics are absent from the SE flank sample. A consistent presence of ferns allies (Adiantum sp., monolete, and trilete spores) was recorded across all samples, with an avg. frequency of 4.5%.
Cropland site (Acacia-asteraceae-poaceae-alternanthera-cerealia-cheno/am assemblage)
This location is predominantly occupied by non-arboreal species (71.7%) compared to arboreal taxa (28.6%). Among arboreal, tree taxa such as Acacia sp. (4.5%–27%) is a dominant taxon followed by Terminalia sp. (4%−8.6%), Anacardiaceae (6.8%), and Ailanthus excelsa (3.6%) is represented in low values. Extra-regional taxon, Pinus sp. is found in significant range (4.3%−10.7%). Shrubby taxa such as Croton (6.7%−11.7%) is the dominant taxa followed by Fabaceae (7%). Among non-arboreal taxa, dominant terrestrial herbs include Asteraceae (8.2%−22.3%), Poaceae (6.7%−14.5%), and Malvaceae (7%−12%) followed by Convolvulaceae (5%−6.4%). Anthropophytes/cultural taxa such as Alternanthera (7.6%–22%), Cerealia (10.4%−18.4%), and Cheno/Am (5.4%−13.6%), are recorded in high values followed by Artemisia (3%−8.2%) and Brassicaceae (2.7%) is represented in low values. The dominant wetland taxon is Cyperaceae (6%−15%), and Polygonum plebeium (1.6%) is recoded in low values, whereas aquatic herbs are absent from the cropland site sample. Monolete spores occurred in all samples with frequencies 5.6%.
Despite the occurrence of tropical moist and dry deciduous forest taxa such as Shorea robusta and Tectona grandis in parts of the broader Ganga Plain (Champion and Seth, 1968), their pollen was not recorded in the surface sediment samples. This absence likely reflects the fragmented distribution of forest patches near the sampling sites and the dominance of agricultural and scrub vegetation around Bakhira Lake. In addition, Tectona grandis and Shorea robusta are generally under-represented in the pollen assemblage due to their low pollen productivity and limited dispersal efficiency (Chauhan, 1994; Chauhan et al., 2013; Kar and Quamar, 2019; Prasad and Quamar, 2023; Quamar and Bera, 2014). Even under peak flowering conditions, their pollen representation remains low, averaging ~10% within forest moss polsters/surface soils and about 4% in open areas ~100 m away from the source vegetation. This reduced representation is further compounded by post-depositional processes, including microbial and chemical degradation of pollen grains within sediments, which significantly diminish their preservation potential (Bera, 1990; Chauhan, 1994; Chauhan et al., 2013; Fraegri and Iversen, 1975). Consequently, the absence representation of teak and Shorea pollen in the present record does not necessarily indicate their absence in the regional vegetation.
Non-pollen assemblages
NW flank site (Botryococcus-glomus-nigrospora-valsaria-podospora-sordaria assemblage)
Among the NPPs assemblage; algal morphs such as Botryococcus (4.6%−14.2%) is recorded in high frequency and dinoflagellates cysts (1.4%–8.8%) is noted in moderate value. Other algae, including Mougeotia type (zygospore), Debarya, and Concentricystis are recorded with low values 1%−3.5%. Coprophilous fungal spores are dominated by Podospora, and Sordaria (HdV-1012), with value ranges of 2.5%–9.5%, with additional contributions from Delitschia and Ascodesmis reaching 2%. Non-coprophilous fungal spore such as Glomus (HdV-1103; 9.3%–24%) and Nigrospora (4.7%−15.7%) are dominant the assemblage followed by Valsaria (4.8%–16%). Other non-coprophilous fungal spores such as Arthrinium, Conidiophore, Curvularia, smut fungi, Cookeina, and Diplodia occur in moderate values (1%−10%) while Pleospora, Tetraploa, Xylariaceae (HdV-1052), Laccaria, Rhizopus sp., and Globose spores, Montagnula sp., are recorded in low frequency (<4%). Zooplankton indicators are also present, with Amoeboid cysts reaching high values (1.7%–14%) and Trichuris sp. detected in low values (1.8%).
Centre site (Botryococcus-dinoflagellate cysts-glomus-valsaria-tetraploa assemblage)
The central basin exhibits a moderately diverse algal signal, with Botryococcus (2.3%–16%) and dinoflagellates cysts (4.8%–9.6%) being moderately represented. Coprophilous fungal spores are recorded with relatively low value of 1.4%–4% includes Delitschia, Sordaria, and Ascodesmis. Among non-coprophilous fungal spores, Glomus (7.6%–28%) and Valsaria (5%–15.4%) are dominant. Tetraploa (1.4%–9.6%) and Conidiophores (1.4%–7%) are recorded in moderate values. Other non-coprophilous fungal spores such as Arthrinium, Rhizopus sp., Nigrospora, Pleospora, Curvularia, Cookeina, Acaulospora mellea, Diplodia, and smut fungi are recorded with moderate values of 1.5%−7.7% while Spegazzinia tessarthra (HdV-1018) is rare (0.5%). Zooplankton remains include Amoeboid cysts (3.6%–12%) and M. moniliformis (4.5%).
SE flank site (Concentricystis-debarya-sordaria-smut fungi-glomus assemblage)
Among the NPPs; algal morphs such as Concentricystis (1%−15.5%) and Debarya (9.4%–10%) are moderately abundant, while Zygnema and dinoflagellates cysts occur at lower frequency (1.4%–4%). Coprophilous fungal spores, particularly Sordaria is prominent (7.8%–11.7%), with Delitschia and Ascodesmis reaching up to 3.7%. Non-coprophilous fungal spores such as smut fungi (17.4%−19.5%), Glomus (7.8%−17.2%) and Pleospora (1.3%−12.3%) are dominant, followed by Globose spores (3.4%−15.6%). Other fungal spores such as Cookeina, Acaulospora mellea, Arthrinium, Spegazzinia tessarthra, Tetraploa, Rhizopus sp., and Nigrospora are recorded with moderate values of 1.5%−11.7% while Curvularia, Conidiophores, Coniothecium glumarum type are recorded in low frequency (0.5%−2.7%). Zooplanktons such as Amoeboid cyst, Dipylidium caninum and Moniezia are recorded in moderate frequency (3.7%−8.7%).
Cropland site (Glomus-smut fungi-coniothecium glumarum-nigrospora-botryococcus assemblage)
At the cropland site, algal remains such as Botryococcus, Concentricystis, Zygnema, Debarya, and dinoflagellates cysts, are moderately represented (1.2%–8.8%), with Echinidinium granulatum recorded at low frequency (2%). Coprophilous fungi including Delitschia, Podospora, Sordaria, and Ascodesmis are relatively sparse (1%–2.5%). Non-coprophilous fungal such as Glomus (6.8%−20.4%) and smut fungi (3%−20%) are dominant the assemblage followed by Coniothecium glumarum (2%−13.4%), Nigrospora (2%–12.7%). Moderate values (2%–9.6%) are recorded for Curvularia, Conidiophores, Tetraploa, Pleospora, and Globose spores, while Diplodia, Rhizopus sp., and Spegazzinia tessarthra remain low (0.5%–1.5%). Zooplanktons such as Amoeboid cyst (3.2%–9%) and Moniezia (3.8%) are recorded in moderate frequency.
TOC and δ13C values
The δ13C values varied between −19.3‰ and −28.7‰ with an avg. value of −22.9‰ and shows decreasing trend from NW flank towards centre to SE flank of the Bakhira Lake. On the other hand, at the cropland area, δ13C values range between −24.7‰ and −27.1‰ with an avg. value of −25.9‰. The TOC/TN ratio range from 8 to 14, with an avg. value of 11, exhibiting a gradual increasing trend from NW flank of lake towards centre to the SE flank of the Bakhira Lake. In contrast, the cropland area recorded a relatively higher TOC/TN ratio, avg. 16, with values ranging from 6 to 24 (Table 1).
Ordination analysis of the pollen data
PCA was applied to investigate spatial variations in pollen composition across the sampled sites and to assess relationships among pollen taxa. Analysis of 47 pollen types from 29 surface lake sediment samples yielded a total inertia of 50.94. The first and second PCA axes accounted for eigenvalues of 7.86 and 5.38, respectively, together explaining 26.5% of the total variance within the dataset (Table 2). These ordination results provide insight into patterns of compositional variability among both sampling locations and pollen types. PCA axis 1 explained 15.7% of the variation and was primarily associated with pollen taxa such as Alternanthera, Malvaceae, Cerealia, Croton, Asteraceae, Acacia sp., Cheno/Am, Cyperaceae, and Brassicaceae that contribute to the high positive score while Holoptelea integrifolia, Aspidopteris, Trewia nudiflora, Borassus flabellifer and Dodonaea that contribute to the high negative score of the PCA axis 1. PCA axis 2 explained 10.8% of the variation and was primarily associated with pollen taxa such as Peltophorum pterocarpum, Iris pseudacorus, Typha, Potamogeton, and Madhuca indica that contribute to the high positive score while Holoptelea integrifolia, Aspidopteris, Trewia nudiflora, Borassus flabellifer and Dodonaea that contribute to the high negative score of the PCA axis 2 (Figure 7 and Supplemental Table 3).
Summary statistics of principal component analysis (PCA) and total inertia for the 29 samples location and 47 pollen types.
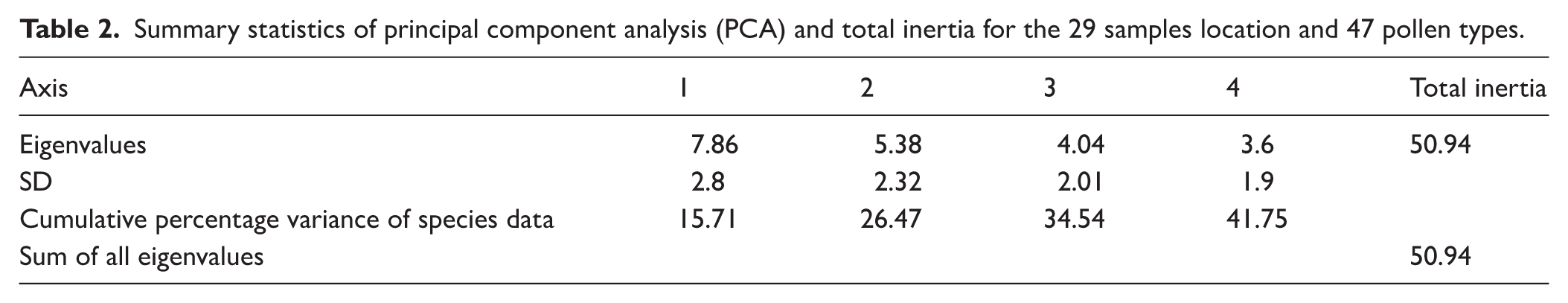
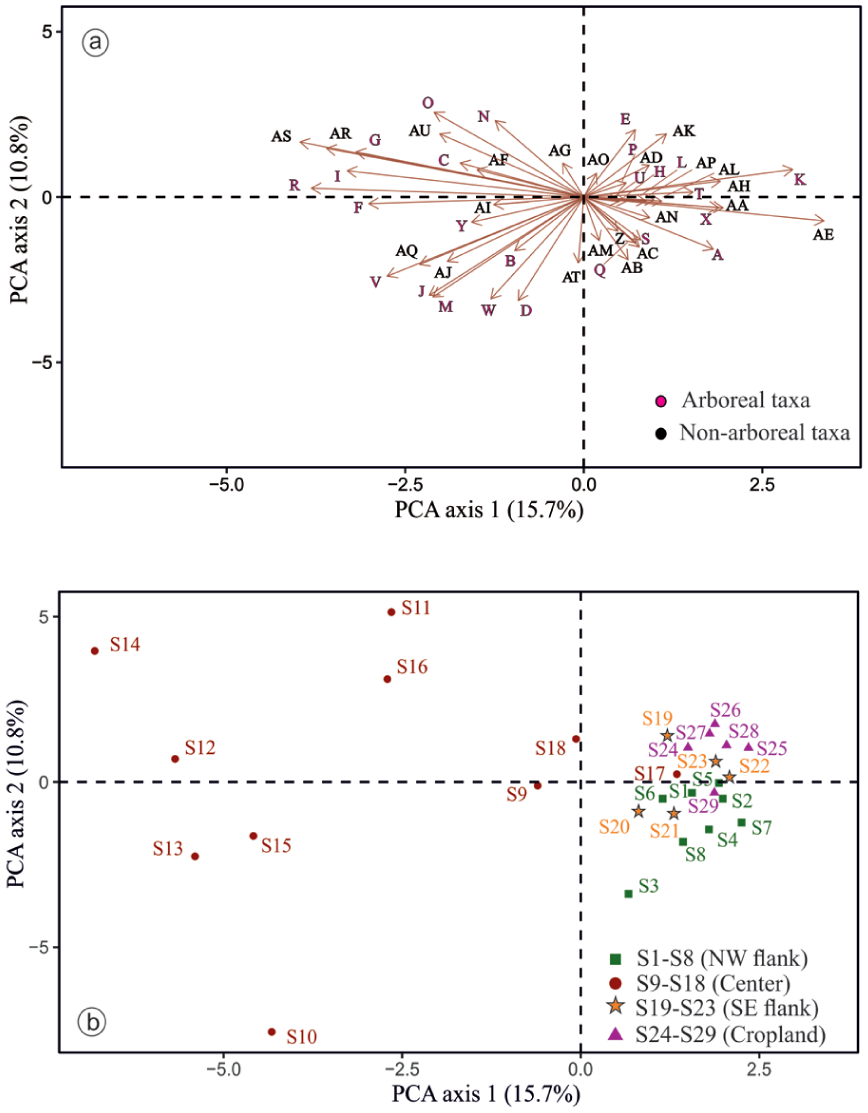
(a and b) PCA results of the surface pollen data from the Bakhira Lake, CGP.
In alignment with the ordination of pollen taxa, the PCA of surface pollen samples reveals distinct spatial structuring. PCA Axis 1 effectively differentiates the central lake surface samples from those collected along the NW and SE flanks and adjacent cropland, except for sample S17, which clusters with marginal sites due to high proportions of Acacia sp., Cerealia, Cyperaceae, Cheno/Am, and Poaceae. PCA Axis 2 further separates the NW flank samples from those of the cropland and most SE flank sites. Along Axis 1, sample S25 exhibits the highest positive score, whereas samples S13 and S15 record the lowest negative values. On Axis 2, sample S11 displays the highest positive score, while S10 records the lowest negative score.
Discussion
Modern multiproxy-vegetation relationships in a monsoon-dominated landscape
The integrated analysis of pollen, NPPs, δ13C values, and TOC/TN ratios from surface sediments of Bakhira Lake provides a coherent picture of modern vegetation structure, hydrological gradients, and anthropogenic influence in the CGP. Rather than operating independently, these proxies collectively capture the dynamic interaction between vegetation composition, land-use intensity, and monsoonal hydrology (Davis et al., 2020; Deng et al., 2025; He et al., 2024; Tripathi et al., 2023; Trivedi et al., 2014, 2015). The modern pollen-rain dataset of 29 surface samples indicated a predominance of non-arboreal pollen (avg. 67.4%), reflecting the present-day landscape dominated by open grasslands, croplands, and marshy vegetation. In contrast, arboreal pollen (avg. 32.6%) is relatively higher in the lake centre, likely due to reduced disturbance and enhanced preservation under more stable, anaerobic depositional conditions, which favour the retention of regionally transported pollen relative to the more disturbed marginal zones. (Bennett and Willis, 2001; Bradshaw and Webb, 1985; Jacobson and Bradshaw, 1981; Moore et al., 1991; Seppä, 2013). This spatial differentiation highlights the strong influence of local vegetation and depositional processes on pollen rain in oxbow lake systems.
Pollen spectra (Figure 5) show that among arboreal taxa, Acacia sp. shows the highest and most consistent representation (avg. 7%), reaching up to 14% in the NW flank, reflecting the dominance of Acacia scrub vegetation adjacent to the lake. Moderate frequencies of Croton (avg. 5.6%), Malvaceae (avg. 5.2%), Anacardiaceae (avg. 2.5%), Aspidopteris (avg. 1.8%), and Madhuca indica (avg. 1.5%) correspond to their local occurrence in scrub dominated and cultivated landscapes. In contrast, several regionally common tropical deciduous taxa (Aegle marmelos, Holoptelea integrifolia, Terminalia, Arecaceae, Trewia nudiflora, and Ailanthus excelsa) occur at low and sporadic frequencies. Such underrepresentation of tropical tree taxa in modern pollen rain has been widely reported from the CGP, including records from Lashoda Tal and Chaudhary-ka-Tal (Trivedi et al., 2015; Trivedi and Chauhan, 2011), Jalesar Lake (Trivedi et al., 2013), and Bari Tal (Trivedi et al., 2014), Kukrail Reserve Forest (Tripathi et al., 2025) as well as from the Ganga-Ghaghara-Gandak interfluves (Tripathi et al., 2023) and the Ganges-Brahmaputra-Meghna floodplain (Kumar et al., 2019). This pattern mainly reflects differences in pollen productivity, dispersal efficiency, and preservation potential among taxa (Birks and Gordon, 1985; Tripathi et al., 2023; Trivedi et al., 2014; Xu et al., 2016). Similar observations have also been reported from tropical deciduous forests of central India, where dominant tree taxa are often poorly represented in modern pollen spectra due to low pollen productivity, insect pollination, and differential preservation in sediments (Chauhan, 1994, 2008). Studies from the northeastern Indian region further demonstrate that many forest taxa occur only sporadically in pollen assemblages despite their presence in the surrounding vegetation, highlighting the strong influence of pollen dispersal and depositional processes in monsoon-dominated environments (Basumatary et al., 2015; Kar and Quamar, 2019; Tripathi et al., 2023). Comparable patterns have also been documented from monsoon-influenced regions of southern China and Southeast Asia, where many tropical tree taxa are weakly represented in modern pollen assemblages due to differences in pollen productivity and dispersal characteristics (Cao et al., 2022; Cheng et al., 2020; Dai et al., 2023; Gosling et al., 2009; Huang et al., 2023; Zheng et al., 2014). The representation of non-arboreal largely aligns with the actual occurrence of ground vegetation in the area, as evidenced by the predominant fraction of avg. 67.4% pollen concentration (Figure 5). Herbaceous vegetation is dominated by grasses (Poaceae), which are consistently well represented in the pollen assemblage, avg. 10.2% across samples and reaching a maximum of 22% in the central part of the lake, indicating open grassland and agricultural landscapes. The combined presence of anthropophytes/cultural taxa viz. Cerealia, Cheno/Am, Brassicaceae, and Alternanthera (avg. 30%) strongly reflects intensive cultivation and anthropogenic disturbance in the lake periphery (Ge et al., 2021; He et al., 2024; Trivedi et al., 2024; Xu et al., 2016). Other anthropophytes taxa such as Cucumis sativus, Asteraceae, and Solanum, occur in lower frequencies (avg. 6%), indicating localised anthropogenic disturbance and seasonal vegetation shifts. Similarly, the dominance of non-arboreal pollen has been widely reported in modern surface samples across the CGP, where herbaceous taxa and anthropophytes are abundant, reflecting significant anthropogenic influence through agricultural activities and land-use practices (Tripathi et al., 2023; Trivedi et al., 2013, 2014, 2015; Trivedi and Chauhan, 2011). Comparable patterns have also been documented in northeastern India, where modern pollen assemblages are frequently dominated by non-arboreal taxa under human-modified landscapes (Basumatary et al., 2015; Tripathi and Pandey, 2023). Similar trends of high non-arboreal pollen representation associated with agricultural landscapes have also been reported from several monsoon-influenced regions of Asia. Modern pollen studies from southern China and Southeast Asia indicate that herbaceous pollen often dominates surface assemblages in cultivated landscapes, frequently exceeding the total pollen sum (Dehghani et al., 2017; He et al., 2024; Xu et al., 2016). Wetland taxa, including Cyperaceae, Polygonaceae, Polygala, Polygonum plebeium, Cressa cretica, and Juncus prismatocarpus (avg. 9%), indicate intermittent marsh conditions along the lake margins. Aquatics, such as Potamogeton, Typha, Lemna and Iris pseudacorus further confirm proximity to permanent water bodies. The presence of ferns allies (Adiantum sp., monolete and trilete) signifies a humid and shaded habitat, consistent with localised moisture retention (Bergeron and Pellerin, 2014; Page, 2002; Tryon and Tryon, 2012). The consistent occurrence of extra-regional taxa such as Pinus, Carpinus, and Juglans, with Pinus avg. 4.5% and the others appearing sporadically, indicates strong wind dynamics and substantial fluvial transport from the adjacent Himalayan highlands, facilitated by their high pollen productivity, buoyancy, and strong preservation potential (Andersen, 1970; Agnieszka Pidek et al., 2010; Bajpai and Kar, 2018; Bohne et al., 2005; Kumar et al., 2019; Matsushita and Sanukida, 1986; Tripathi et al., 2023; Xu et al., 2016). Principal Component Analysis (PCA) results indicate that Alternanthera showed the highest positive score on Axis 1, whereas Holoptelea integrifolia exhibits the lowest. Other taxa such as Aspidopteris, Trewia nudiflora, Borassus flabellifer and Dodonaea show negative score along Axis 1, while Malvaceae, Cerealia, Croton, Asteraceae, Acacia sp., Cheno/Am, Cyperaceae, and Brassicaceae showed positive scores (Figure 7). Axis 1 reflects a compositional gradient driven by differences in the relative abundance of arboreal taxa in the lake centre compared to a greater representation of cultivated and open-land elements along the flanks and adjoining cropland. Although the positive scores of cultivated and open-land taxa indicate the imprint of contemporary land-use activities, this axis primarily captures spatial heterogeneity in vegetation composition and landscape structure and may also reflect anthropogenic influence. Along PCA Axis 2, Peltophorum pterocarpum shows the highest positive score, while Holoptelea integrifolia and Aspidopteris record the most negative values. Additional taxa such as Iris pseudacorus, Typha, Potamogeton, and Madhuca indica are positively associated, whereas Albizia procera, Trewia nudiflora, Borassus flabellifer and Dodonaea are negatively associated. This axis is interpreted as reflecting hydrological control, particularly water-mediated transport and depositional processes of palynomorphs. The spatial distribution patterns further demonstrates that arboreal taxa increase towards the lake centre, whereas non-arboreal taxa are relatively more abundant along the margins, indicating that hydrodynamic effects play a dominant role in palynomorph dispersal. Notably, sample S25, with the highest score on Axis 1, likely reflects the driest microenvironment, while samples S13 and S15, with the lowest scores, are associated with wetter conditions. On Axis 2, S11 has the highest positive score and S10 the lowest, possibly reflecting spatial variability in recent SW monsoonal activity within the CGP.
The NPPs assemblages provide process-based insights into hydrology, grazing intensity, soil disturbance, and anthropogenic activity across Bakhira Lake (Etienne et al., 2013; Graf and Chmura, 2006; Parker and Williams, 2012; Raper and Bush, 2009; van Geel et al., 2003). Their spatial distribution across the NW flank, Centre, SE flank, and Cropland sites reflects the interaction between local environmental gradients and land-use pressure (Figure 6). Algal remains, including Botryococcus, Concentricystis, and Zygnemataceae, confirm the persistence of aquatic and semi-aquatic conditions, particularly in central and marginal wetland zones (van Geel, 2001; van Geel et al., 2011). Their abundance indicates standing or slow-moving water and supports the hydrological interpretations derived from pollen data. Coprophilous fungal spores including Sordaria, Ascodesmis, Podospora, and Delitschia are particularly abundant in the NW and SE flanks, indicating active grazing pressure in these zones (Basumatary and McDonald, 2017; Burney et al., 2003; Davis, 1987; Etienne et al., 2013; Lee et al., 2022; Mazier et al., 2006; Schlütz and Shumilovskikh, 2017; van Asperen et al., 2021; Wang et al., 2025; Zhang et al., 2025). These fungi develop on herbivore dung, and their elevated frequencies suggest sustained livestock activity or natural herbivory. In contrast, relatively lower abundances in cropland samples likely reflect reduced open grazing due to agricultural land management. The consistent occurrence of coprophilous taxa further implies warm and humid conditions, as adequate moisture is required for sporulation and dispersal (Dix, 2012; Kuthubutheen and Webster, 1986; van Geel et al., 2011). Saprophytic fungi viz. Curvularia, Valsaria, Cookeina, Arthrinium, Rhizopus, and Xylariaceae indicate organic matter decomposition under humid conditions, whereas plant pathogenic fungi viz. Diplodia, Tetraploa, Nigrospora, Pleospora, and smut fungi are more frequent in cropland and marginal sites (Gelorini et al., 2011; van Geel et al., 2011; Gou et al., 2022). The occurrence of smut fungi particularly supports active cereal cultivation and crop-associated fungal infection in agricultural areas (Carris et al., 2006; Kumar and Attri, 2016; Ramazanova et al., 2024). Glomus spores occur consistently and in relatively high frequencies across sites. Glomus, an arbuscular mycorrhizal fungus, is widely recognised as an indicator of soil erosion and catchment disturbance because its spores are transported into lake sediments via surface runoff (Kołaczek et al., 2013; van Geel, 1986; van Geel et al., 2011). Elevated Glomus frequencies in flank and cropland sites suggest enhanced soil erosion linked to grazing, cultivation, and monsoon-driven runoff. This pattern underscores the sensitivity of the oxbow lake system to catchment-scale land-use modification and seasonal hydrological processes (Limaye et al., 2007; Masetto and Lorscheitter, 2014; Musotto et al., 2012). The presence of amoeboid cysts, especially abundant in the NW flank (7.5%), suggests stagnant or slow-moving water bodies that favour protozoan proliferation (Booth, 2002; Neville et al., 2010; Farooqui et al., 2020; Mitchell et al., 1999; Nolan et al., 2019; Trivedi et al., 2014). Additionally, parasitic helminth remains such as Dipylidium caninum and Moniezia indicate faecal contamination from domestic or wild herbivores and further corroborate grazing activity and human-animal interactions in the lake catchment (Okoronkwo and Onwuliri, 2001; Reinhard et al., 1986; Shumilovskikh et al., 2021).
Stable carbon isotope (δ13C) values of sedimentary organic matter provide a robust indicator of dominant photosynthetic pathways (C3 vs C4 vegetation) within the catchment (Agrawal et al., 2015; Cerling et al., 1997; Ehleringer et al., 1997; Kumar et al., 2022). The C3 and C4 vegetation exhibit distinct isotopic signatures, δ13C values serve as an effective tracer of vegetation composition and associated climatic controls such as temperature and moisture availability (Basu et al., 2015; Boom et al., 2001). In contrast to pollen, which is subject to differential production, dispersal, and preservation biases, sedimentary organic matter integrates bulk vegetation input and is generally assumed to retain the mean isotopic signal of the source vegetation with minimal fractionation (Cerling et al., 1997; Whitbread, 1995). The positive correlation between TOC and TN in both peripheral (NW and SE flanks, cropland) and central lake samples suggest broadly similar organic sources across the basin (Hedges et al., 1986). However, the stronger correlation in the flanks and cropland sites (R2 = 0.4) compared to the lake centre (R2 = 0.1) indicates greater variability in nitrogen-rich inputs in marginal zones, likely influenced by vegetation type, agricultural activity, and soil processes (Supplemental figure 7). TOC/TN ratios further constrain organic matter sources. Values <10 typically indicate algal or aquatic input, whereas ratios >10 reflect terrestrial plant contributions (Meyers and Teranes, 2002). In the present study, TOC/TN ratios range from 6 to 24 across peripheral sites, indicating mixed terrestrial and aquatic inputs. The notably low ratio in sample S23 (TOC/TN = 3) suggests enhanced aquatic contribution or localised nitrogen enrichment. Central lake samples show comparatively lower and less variable TOC/TN ratios (8–14), consistent with greater aquatic influence (Figure 8). The relatively low δ13C values (ranging from –21.2‰ to –28.7‰) indicate dominant C3 vegetation input across peripheral sites (Agrawal et al., 2012; Basu et al., 2015; Trivedi et al., 2024). An exception is sample S23 from the SE flank (−19.23‰), which suggests localised C4 input, potentially linked to agricultural activity, open grassland vegetation, or specific hydrological pathways transporting C4 derived organic matter into this depositional zone. When integrated with pollen data, peripheral sites exhibit strong coupling between non-arboreal pollen dominance (avg. 66.9%), variable TOC/TN ratios, and predominantly C3 isotopic signatures, reflecting intensive land use and mixed terrestrial-aquatic inputs. In contrast, central lake samples show increased arboreal pollen (avg. 38.9%), relatively higher δ13C values (ranging from −19.7‰ to −21.7‰), and moderate TOC/TN ratios, indicating that the organic matter at the central site is derived from C3 dominated to mixed C3–C4 vegetation source relative to peripheral zones (Figure 9 and Table 1). This mixed isotopic signature reflects variability in C3−C4 vegetation dynamics, likely driven by fluctuating SW monsoonal condition in the CGP (Basu et al., 2015; Tripathi et al., 2023). The observed variation in δ13C values and TOC/TN ratio at the lake centre, relative to peripheral regions, is primarily controlled by hydrological changes within the lake during the SW monsoon season, which influence both organic matter input and preservation.
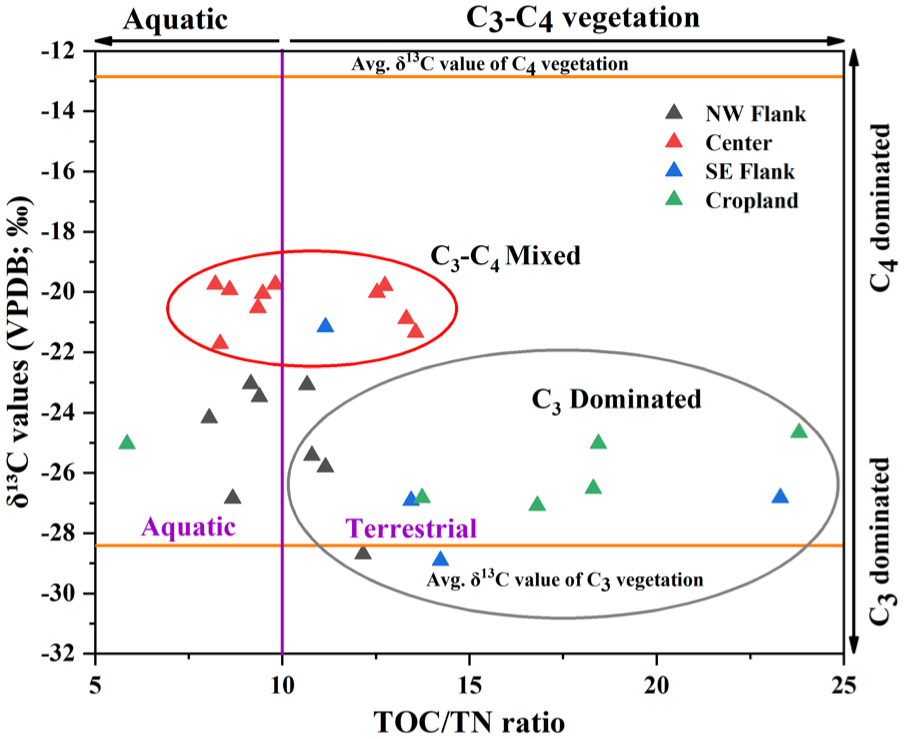
Cross-plots between δ13C and TOC/TN values representing the sources of organic matter contributions in the surface sediment samples.
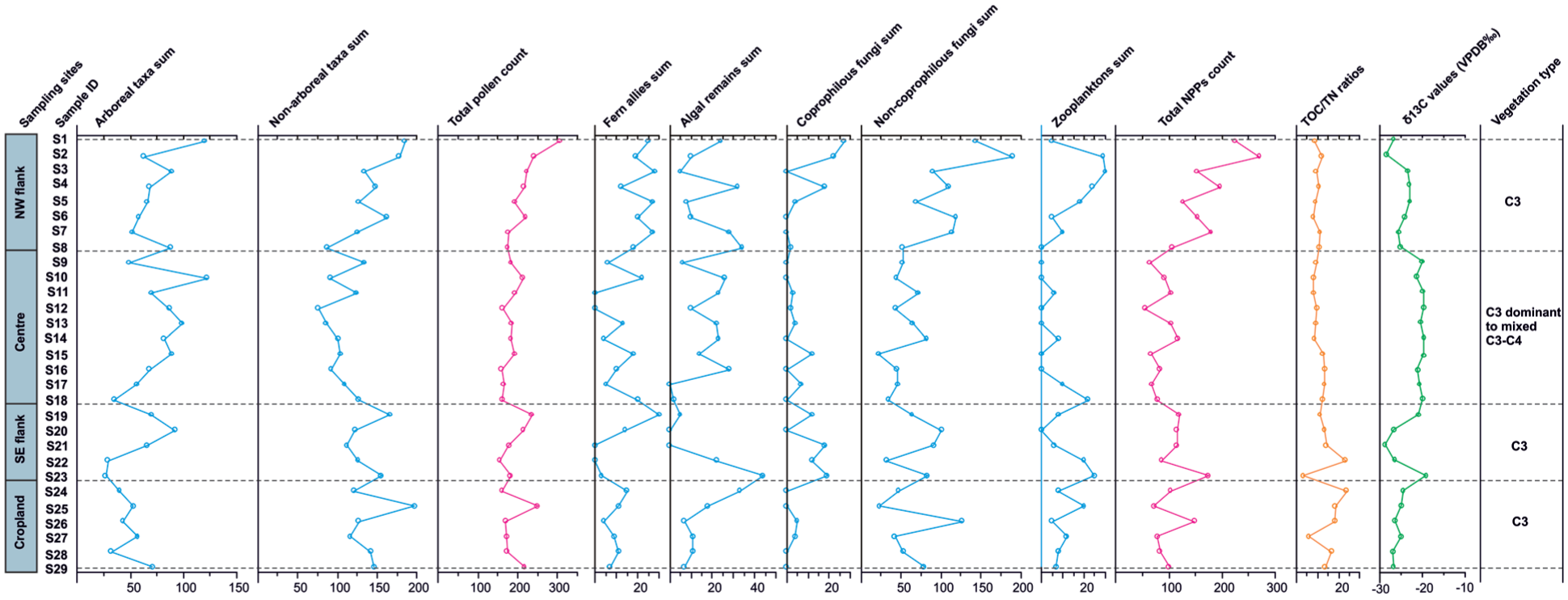
Spatial distribution of total pollen and NPPs sums together with TOC/TN ratios and δ13C values derived from the surface sediments of Bakhira Lake.
Overall, the integrated pollen, NPPs, and stable carbon isotope data from Bakhira Lake demonstrate that modern surface sediments archive the coupled effects of vegetation composition, land-use intensity, and monsoon-driven hydrological processes in the CGP. Pollen assemblages capture spatial vegetation gradients and depositional dynamics, NPPs provide process-based insights into grazing pressure, soil erosion, and aquatic conditions, while δ13C values and TOC/TN ratios constrain organic matter sources and dominant photosynthetic pathways. Together, these multiproxy datasets establish a robust modern analogue for interpreting fossil sediment records from monsoon-dominated lacustrine systems and strengthen the reliability of palaeoenvironmental reconstructions from the CGP.
Implications for palaeoclimatic and palaeoecological reconstruction
The modern multiproxy dataset from Bakhira Lake provides a robust analogue framework for interpreting palaeoenvironmental records preserved in lacustrine archives of the CGP. Modern pollen assemblages capture the heterogeneous vegetation landscape surrounding the lake, including scrub woodland, grassland, cropland and wetland communities. Variations in the relative abundance of arboreal and non-arboreal pollen therefore provide valuable insights into changes in vegetation structure and land-use intensity (Birks and Birks, 1980; Prentice, 1985; Sugita, 2007a). The dominance of non-arboreal pollen, particularly Poaceae and Asteraceae, reflects the prevalence of open grassland and disturbed habitats in the modern landscape. Such assemblages are typically associated with open vegetation under seasonally dry conditions or human-modified landscapes (Dehghani et al., 2017; He et al., 2024; Trivedi et al., 2014, 2015; Trivedi and Chauhan, 2011; Zhao et al., 2024). In contrast, arboreal pollen taxa including Acacia sp., Madhuca indica, Aegle marmelos, Terminalia and Trewia nudiflora represent components of tropical deciduous woodland occurring in patches across the regional vegetation mosaic. Consequently, variations in the relative abundance of AP and NAP, therefore provide a useful proxy for reconstructing transitions between woodland-dominated and grassland-dominated ecosystems in palaeo-records (Birks and Birks, 1980; Sugita, 2007a). Aquatic and wetland taxa such as Typha, Potamogeton and Cyperaceae primarily represent vegetation growing within lake margins and shallow-water environments. Fluctuations in these taxa in sedimentary sequences therefore serve as sensitive indicators of local hydrological variability and lake-level fluctuations associated with monsoon variability. Similarly, NPPs, including algal remains and fungal spores, provide complementary information on environmental conditions that cannot be inferred from pollen alone. Algal remains reflect variations in lake productivity, nutrient availability and water-level fluctuations, whereas fungal spores, particularly coprophilous taxa such as Sordaria, Ascodesmis, Podospora and Delitschia, indicate grazing activity, cattle herbivores presence and catchment disturbance. Other soil-derived fungal remains may also reflect soil erosion and organic matter influx into the lake basin. Comparable interpretations have been widely reported, where algal remains are linked to lake-level and trophic changes, and coprophilous fungi and Glomus indicate grazing pressure and catchment erosion (Cugny et al., 2010; Pandey et al., 2026; Tang et al., 2013; van Geel, 1986; Zhao et al., 2013; Cheruiyot et al., 2020). Together, these NPPs provide important insights into moisture availability, nutrient dynamics and anthropogenic influence. Stable carbon isotope values further strengthen palaeoenvironmental interpretation by distinguishing between organic matter derived from C3 woody vegetation and C4 grasses. Low δ13C values generally indicate increased input from C3 plants typical of woodland environments under relatively humid conditions, whereas high δ13C values suggest greater contributions from C4 grasses associated with open landscapes and seasonal moisture stress (Agrawal et al., 2012; Basu et al., 2015; Cerling and Quade, 1989; Farquhar et al., 1989). When interpreted together with pollen assemblages, δ13C values provide an additional line of evidence for evaluating vegetation dynamics and distinguishing hydroclimatic variability from anthropogenic landscape modification.
The modern multiproxy relationships documented from Bakhira Lake are consistent with pollen-vegetation studies conducted across several lakes of the CGP. Surface pollen investigations from Jalesar Lake, Lashoda Tal, Chaudhary-ka-Tal and Kikar Tal demonstrate that modern pollen rain is typically dominated by non-arboreal taxa, particularly Poaceae and other herbaceous elements, whereas arboreal taxa such as Acacia, Madhuca indica and Holoptelea integrifolia occur in comparatively lower frequencies despite their presence in the surrounding vegetation (Saxena et al., 2017; Trivedi et al., 2013, 2014, 2015; Trivedi and Chauhan, 2011). These studies highlight the strong influence of pollen productivity, dispersal efficiency and preservation on pollen representation in tropical deciduous landscapes and emphasise the importance of modern pollen-vegetation calibration for interpreting fossil assemblages. The modern analogue datasets generated from these surface studies have subsequently been applied in lacustrine sediment records from the same lakes to reconstruct Holocene vegetation dynamics, monsoon variability and human-environment interactions across the CGP. In above said studies, fluctuations in AP and NAP proportions are interpreted as indicators of vegetation transitions between woodland and grassland in response to variation in monsoon intensity and anthropogenic disturbance. As vegetation distribution, hydrological variability and lake productivity in the CGP are closely linked to fluctuations in the Indian summer monsoon, multiproxy signals preserved in lacustrine sediments provide a sensitive archive of monsoon-driven environmental variability. By integrating modern vegetation, hydrological conditions and land-use patterns with multiproxy sediment signatures, this study strengthens the regional calibration dataset and provides a robust framework for interpreting fossil records and reconstructing Holocene vegetation dynamics, hydroclimatic variability and human-environment interactions in monsoon-dominated lacustrine systems of the Indian subcontinent.
Conclusions
Based on the integrated analysis of pollen, non-pollen palynomorphs, stable carbon isotopes and TOC/TN ratios from modern surface sediments of Bakhira Lake, the following conclusions are drawn:
Modern pollen assemblages reflect the vegetation mosaic surrounding Bakhira Lake. Non-arboreal pollen NAP dominates marginal and cropland sites, indicating open grassland and agricultural landscapes, whereas the lake centre preserves relatively higher arboreal pollen representing regional tropical deciduous woodland elements. The central and deepest part of the lake therefore provides a more suitable location for sediment coring, as relatively permanent waterlogging and reduced sediment disturbance favour better preservation of pollen and other palaeoenvironmental proxies under low-oxidation conditions.
Anthropogenic land use strongly influences modern pollen rain. The frequent occurrence of Cerealia, Cheno/Am, Alternanthera, and other anthropophytes taxa reflects intensive cultivation and human-modified landscapes within the lake catchment.
Aquatic and marsh taxa indicate local hydrological conditions. Taxa such as Cyperaceae, Typha, and Potamogeton represent vegetation growing along lake margins and shallow-water environments and therefore reflect local water-level fluctuations.
NPP assemblages provide additional ecological information on grazing activity and catchment disturbance. Coprophilous fungal spores (e.g. Sordaria, Podospora, Delitschia, and Ascodesmis) indicate grazing pressure, whereas algal remains and fungal indicators such as Glomus reflect aquatic productivity and soil erosion processes.
Stable carbon isotope values and TOC/TN ratios constrain organic matter sources and vegetation types. Low δ13C values indicate dominant C3 vegetation input in peripheral zones, while relatively enriched values in the lake centre suggest mixed C3–C4 vegetation contributions.
The integrated multiproxy dataset from Bakhira Lake provides a modern analogue framework for palaeoenvironmental interpretation in the Central Ganga Plain. The combined pollen, NPPs, δ13C, and TOC/TN data provide a modern reference for interpreting fossil lacustrine records and identifying vegetation changes, hydrological variability, grazing activity, and anthropogenic influence.
Supplemental Material
sj-docx-1-hol-10.1177_09596836261458257 – Supplemental material for Modern pollen-vegetation-stable carbon isotope analogues of Bakhira lake, central Ganga plain, India: implications for palaeoclimatic and palaeoecological reconstruction
Supplemental material, sj-docx-1-hol-10.1177_09596836261458257 for Modern pollen-vegetation-stable carbon isotope analogues of Bakhira lake, central Ganga plain, India: implications for palaeoclimatic and palaeoecological reconstruction by Mohd Ikram, Anjali Trivedi, Shailesh Agrawal, Anupam Sharma and Parminder Singh Ranhotra in The Holocene
Footnotes
Acknowledgements
The authors sincerely thanks Prof. Mahesh G. Thakkar, Director, Birbal Sahni Institute of Palaeosciences (BSIP), Lucknow, for his consistent support and encouragement. The authors are especially grateful to Dr. S. K. Basumatary for his expert guidance in the identification of non-pollen palynomorphs (NPPs). Mohd Ikram acknowledges the Council of Scientific and Industrial Research (CSIR), Government of India, for financial support through a Senior Research Fellowship (SRF, Sr. No. 1061920588). This study contributes to the Quaternary Lake Deep Coring Project (QLDP, Project 8), part of the collective research programme at BSIP.
Ethical considerations
Ethical approval was not required for this research as it did not involve human participants, animal subjects, or sensitive data.
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was conducted with institutional support from the Birbal Sahni Institute of Palaeosciences (BSIP), Lucknow, India. No external funding was received for this study.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
All data supporting the findings of this study are available within the article and its supplementary materials. Additional data can be obtained from the corresponding author upon reasonable request, using the email address provided*.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.