
Abstract
The small mammal assemblages from seven open-air archeological sites in the Argentinean Humid Chaco are analyzed from taphonomic, biogeographic and paleoenvironmental perspectives. The study employs standard taxonomic and taphonomic methodologies, complemented by Temporal Niche of Occurrence (NOO3D) analysis, diversity indices, and multivariate statistical techniques. Small mammal remains were accumulated at the archeological sites through both anthropogenic activities and natural in situ deaths. Despite good preservation due to rapid burial, the specimens exhibit evidence of trampling and root activity. With the specimens studied here, we expand the known Late-Holocene small mammal record in the Humid Chaco from four to seven taxa, reflecting the typical present day composition of this biome between 1,6 and 0,6 ka cal BP. The identified species are typical of flooded environments (Lutreolina crassicaudata, Holochilus brasiliensis, H. chacarius), open grasslands and sandy areas (Cavia aperea, Ctenomys sp.), hydrophytic vegetation (Scapteromys aquaticus), and gallery forests (Sooretamys angouya). The record of Sooretamys represents one of the few known fossil occurrences in South America, the second in Argentina, and the first in the Humid Chaco. Ecological niche models for this rodent support the persistence of the region’s fluvial system and gallery forests since the Pleistocene–Holocene boundary. Human groups inhabiting the Humid Chaco over the past 1600 years likely adapted their gathering, fishing, and hunting practices—including the exploitation of large, medium, and small vertebrates such as Cavia and Holochilus—along with their settlement strategies, to optimize resource use and reduce transportation costs in response to climatic cycles, including phases of increased rainfall.
Introduction
Chaco is the second largest region in South America—surpassed only by Amazonia—and encompasses southeastern Bolivia, the western fringe of Paraguay, and the central-northern region of Argentina (Maldonado and Höhne, 2006). This region is conventionally divided into the Dry Chaco in the west, characterized by xerophytic forests, and the Humid Chaco in the east (Figure 1(a)), which is formed by the distal areas of the alluvial fans of the Paraguay and Paraná rivers, along with their main tributaries, such as the Bermejo, Pilcomayo, and Oro rivers (Figure 1(b)). The gentle regional slope promotes water retention, leading to the formation of wetlands with permanent and seasonal marshes, estuaries, and a mosaic of woodlands, grasslands, and gallery forests, which support the high biological and cultural diversity characteristic of the Humid Chaco (Ginzburg and Adámoli, 2006; Iriondo, 1993; Lamenza, 2013, 2015; Oyarzabal et al., 2018). The rainy season coincides with the warm season, from October to April. Minimum rainfall occurs during the winter season, from June to August, when droughts take place (Ginzburg and Adámoli, 2006). The intensity of these precipitations is primarily regulated by the cycles of the El Niño-Southern Oscillation (ENSO), with its dry (La Niña) and wet (El Niño) events, triggering regular geomorphological changes through severe droughts and flooding (Iriondo, 1999; Meggers, 1994; Morello, 1995). Faunistic compositions in this area, especially small mammals, are sensitive to these fluctuations (Contreras and Berry, 1985).
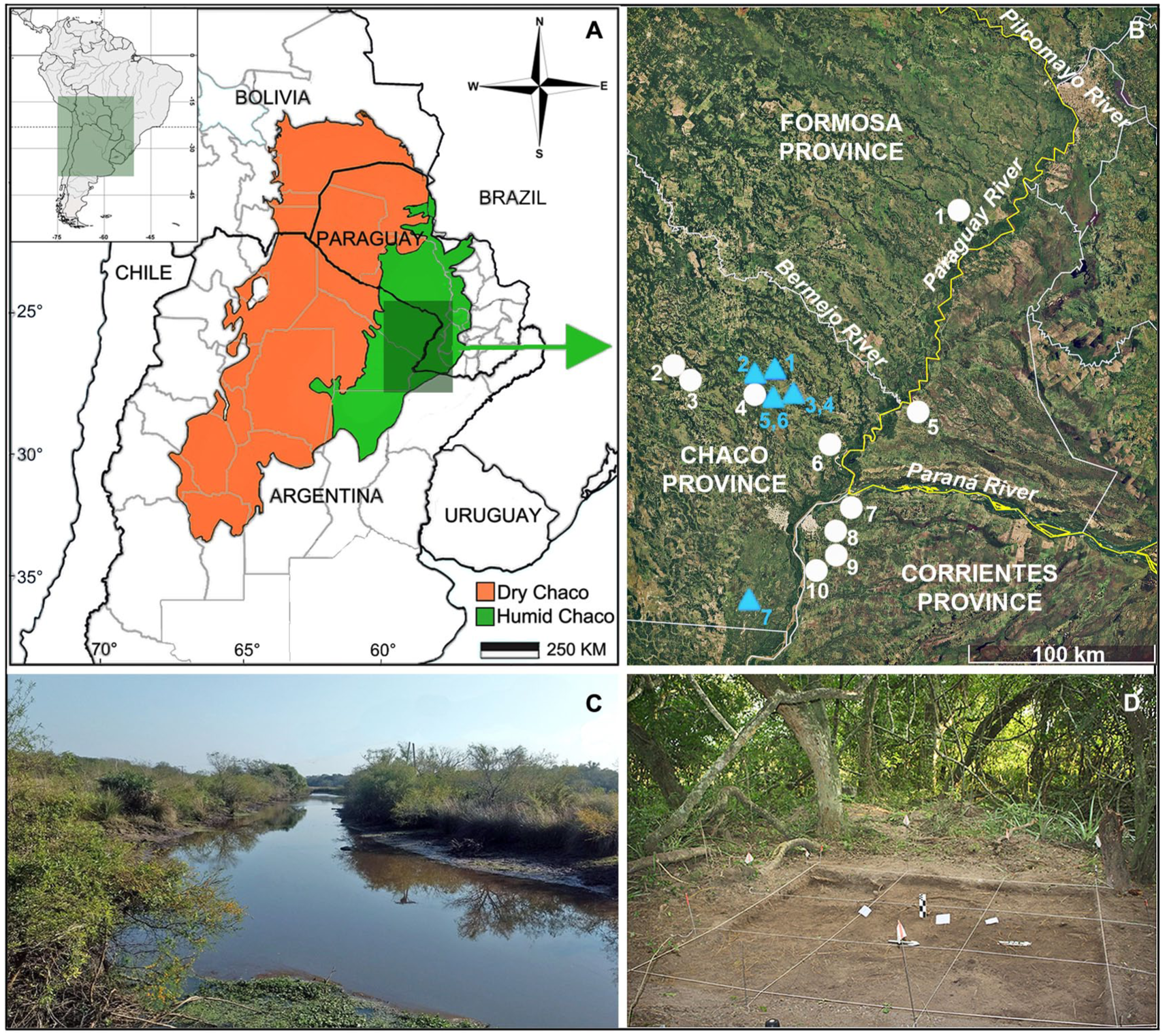
(a) Map of the Chaco region and its subdivisions: Dry Chaco and Humid Chaco; (b) Geographic location of the archeological sites (sky-blue triangles) and present day localities with small mammal data (white dots) considered in this study; (c) View of Quia Creek and the vicinity of El Cachapé Potrero V Anexo; (d) Photograph of the archeological excavation area at the same site. Archeological sites: (1) Sotelo I; (2) La Ilusión; (3) El Cachapé Potrero IVA; (4) El Cachapé Potrero IVB; (5) El Cachapé Potrero V; (6) El Cachapé Potrero V Anexo; (7) Puesto Fantín. Present day localities (#1–10) are detailed in Supplemental Table 1.
Specific paleoclimatic knowledge of the Humid Chaco remains limited, with most studies focusing on geomorphological and sedimentological evidence from various sectors of the Chaco-Pampean plain during the Late-Holocene, which are typically associated with the ‘Pampean pattern’ model (Iriondo, 1990, 1993, 1999, 2006).
In general, the climate of this region during the Late-Holocene was variable, shifting from a semi-arid phase between 3500 and 1400 years BP to a period of increased humidity during the Medieval Climate Anomaly (∼1200–700 years BP), before returning to more arid conditions during the Little Ice Age (∼700–150 years BP; Iriondo, 2006; Iriondo and Kröhling, 1995; Ljungqvist, 2010). Since the 20th century, the region has experienced a return to the humid conditions that characterize the present climate regime (Iriondo, 2006). Recent studies point to a progressive increase in temperature and humidity in the Humid Chaco since about 1600 years BP, inferred from remains of the endemic lungfish Lepidosiren paradoxa (Zarza et al., 2025). In Amazonia, cultural discontinuities at 1500, 1000, 700, and 400 years BP have been linked to major paleo-ENSO events (Meggers, 1994). Farther south, fossil pollen and small-mammal data from southern Brazil reveal a shift from drier, open landscapes with gallery forests in the Early-Holocene to a more closed, humid forest during the Late-Holocene (Fernández et al., 2019; Hadler et al., 2013, 2016, 2025; Stutz et al., 2017, 2020).
Knowledge of the terrestrial small mammals of the Chaco region remains sparse, primarily owing to its dense vegetation and swampy terrain, which severely hinder systematic fieldwork and fossil recovery (Pardiñas et al., 2005; Pardiñas and Teta, 2005; Teta et al., 2009). Following early 20th-century taxonomic studies (Gyldenstolpe, 1932; Shamel, 1931), the rich biodiversity of small mammals in the Humid Chaco drew the attention of mammalogists during the latter half of the century (e.g. Contreras and Berry, 1982, 1983, 1984, 1985; Hershkovitz, 1962, 1966; Kravetz et al., 1986; Massoia, 1970, 1971, 1973a, 1973b; Massoia and Fornes, 1964; Massoia et al., 1988, 1990, 1995, 1997, 1999). In this century, taxonomic and biogeographic knowledge of the terrestrial small mammals of the Humid Chaco has been enriched by a growing body of research (e.g. D’Elía et al., 2008; Heinonen Fortabat, 2001; Jayat et al., 2006; Pardiñas et al., 2005; Pardiñas and Teta, 2005; Teta et al., 2009; Teta and Pardiñas, 2010; Udrizar Sauthier and Carrera, 2013). Moreover, even the recent description of a new species of arboreal cricetid sigmodontine rodent, Oecomys franciscorum, apparently endemic to the Humid Chaco, underscores this progress (Pardiñas et al., 2016). However, fossil evidence for small mammals in the Humid Chaco remains exceptionally scarce, stemming from compounded logistical challenges, including the absence of caves and rock shelters in the study area, which are the environments where small-mammal remains most achieve optimal preservation (Andrews, 1990). Most of the small mammal fossil record derives from a handful of rodents and marsupials species recovered at open-air archeological sites, with dates corresponding to the latter half of the Late-Holocene (e.g. del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015). Notably, the discovery of Sooretamys angouya at one of these sites represents the first fossil record of this oryzomyine rodent in the Humid Chaco and one of the few known Quaternary occurrences in South America, providing key evidence for the long-term persistence of gallery forests in the region.
This paper aims to analyze the complete small mammal record from several Late-Holocene archeological sites in the Humid Chaco. We combine previously studied and novel samples to elucidate the provenance of these deposits and to characterize the composition and natural history of Late-Holocene small mammals from the eastern fringe of the Humid Chaco. Consequently, taphonomic, taxonomic, and biogeographic information—along with other biological and geological indicators recovered from the Humid Chaco and surrounding regions—will be used to reconstruct the paleoenvironmental conditions during the period when human groups adapted their fishing, hunting, gathering, and settlement strategies to the environment. Finally, we applied a temporal ecological-niche model (NOO3D) for the sigmodontine rodent Sooretamys angouya, integrating both fossil and contemporary occurrence records and projecting its climatic niche onto four paleoclimatic chronozones (Late-Pleistocene, Early-Holocene, Mid-Holocene and Late-Holocene) to evaluate niche stability through time. In this context, environmental stability refers to the persistence of the characteristic mosaic of wetlands, grasslands, and gallery forests that defines the Humid Chaco, rather than the absence of climatic fluctuations. This stability is evidenced by the continuous presence of indicator species and the maintenance of suitable ecological niches over time, even as precipitation regimes shifted in response to ENSO cycles.
Archeological sites
This study examines seven open-air archeological sites situated in the eastern sector of the Humid Chaco, Chaco Province, Argentina (Figure 1(b)): (1) Sotelo I (26°47'31.9′'S, 59°03'31.9′'W, 69 m asl); (2) La Ilusión (26°48'S, 59°08'W, and 70 m asl); (3) El Cachapé Potrero IVA (26°52'30′'S, 58°57'26′'W, 71 m asl); (4) El Cachapé Potrero IVB (26°52'27.5′'S, 58°57'25.6′W, 71 m asl); (5) El Cachapé Potrero V (26°53'47.2′'S, 59°00'49′'W, 70 m asl); (6) El Cachapé Potrero V Anexo (26°53'33′'S, 59°00'56′'W, 68 m asl); (7) Puesto Fantín (27°53'26.5′'S, 59°04'16.6′'W, 46 m asl).
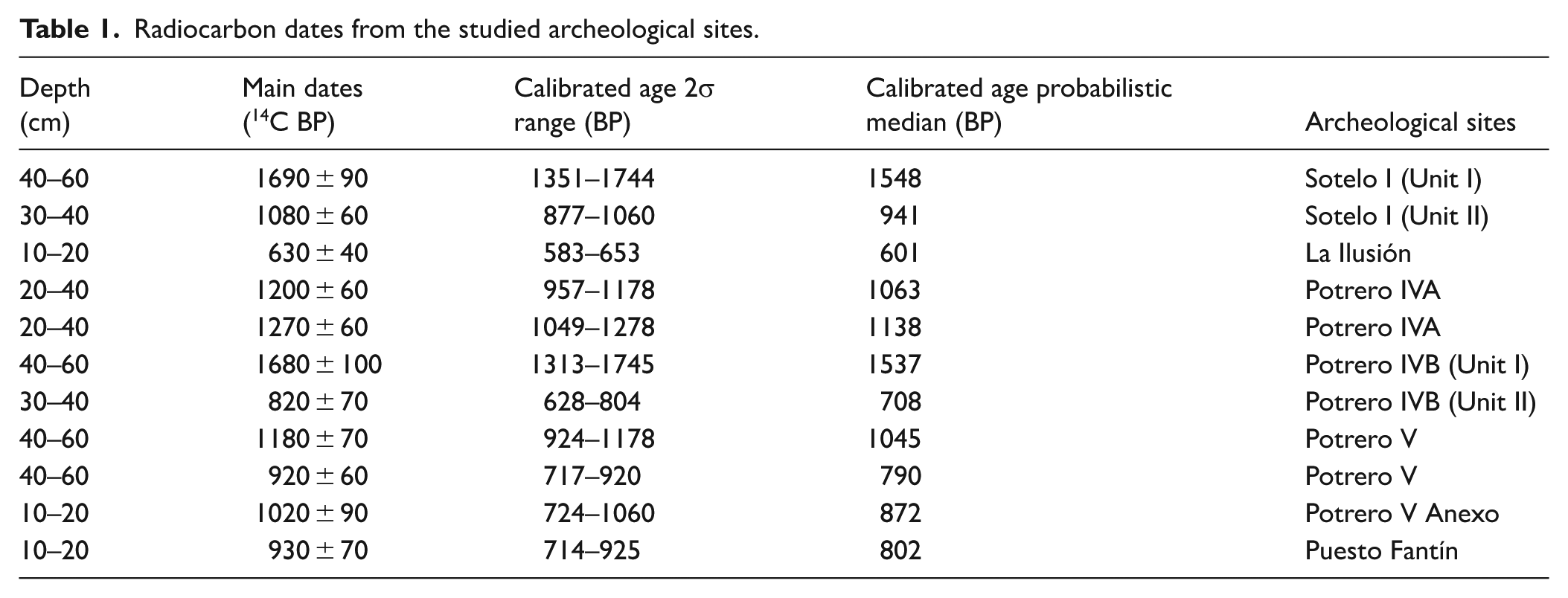
Almost all of the archeological deposits (El Cachapé Potrero IVA, IVB, V, V Anexo, and La Ilusión) are emplaced on fluvial levees characterized by the presence of riverine forests near creeks or other bodies of water (Figure 1(c) and (d)). They are surrounded by grasslands and temporary floodplains, and lie between 8 and 4 km from the Guaycurú and Oro rivers, respectively. Meanwhile, the southernmost site, Puesto Fantín, is situated on the meandering erosional bank of the Paranacito river (Santini, 2009). All of these archeological sites share a sedimentary matrix composed of shell-bearing, organic-rich black soil that contains abundant archaeofaunal remains and pottery fragments, but lacks lithic artifacts (e.g. del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015). Further details regarding the archeological excavations and recovered materials are available in the aforementioned works and other related studies (Zarza et al., 2019, 2025). Radiocarbon dating (Table 1) indicates that these sites fall within the terminal Late-Holocene (1500–600 years BP). The differing radiocarbon chronologies obtained across the strata of both Sotelo I and El Cachapé Potrero IVB justify subdividing each site into two distinct subunits (Table 1).
Radiocarbon dates from the studied archeological sites.
Hunter-gatherer-fisher societies established long-term base camps along riverbanks and followed seasonal mobility circuits governed by precipitation cycles and droughts, optimizing their access to water-dependent resources. This dual settlement strategy facilitated the efficient procurement of both small and large prey, thereby minimizing foraging time and transport costs. The human groups that occupied these sites were well adapted to riverine environments, where they exploited predictable resources (del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015; Zarza et al., 2019, 2025).
To date, 20 mammalian taxa have been identified from these archeological sites. These include two didelphids (Didelphis albiventris, Lutreolina crassicaudata), three cervids (Blastocerus dichotomus, Mazama sp., Ozotoceros bezoarticus), one tayasuid (Tayassu pecari), two felids (Leopardus pardalis, Puma concolor), two canids (Cerdocyon thous, Chrysocyon brachyurus), one procyonid (Procyon cancrivorus), one mustelid (Lontra longicaudis), one cingulate (Dasypus novemcinctus), one leporid (Sylvilagus brasiliensis), five caviomorphs (Cavia aperea, Ctenomys sp., Dolichotis salinicola, Hydrochoerus hydrochaeris, Myocastor coypus), and one sigmodontine—Holochilus brasiliensis—(del Papa and Lamenza, 2019; Santini, 2009; Santini and Lamenza, 2015). The mammalian record at these sites was primarily accumulated through human subsistence activities (del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015).
Materials and methods
Samples
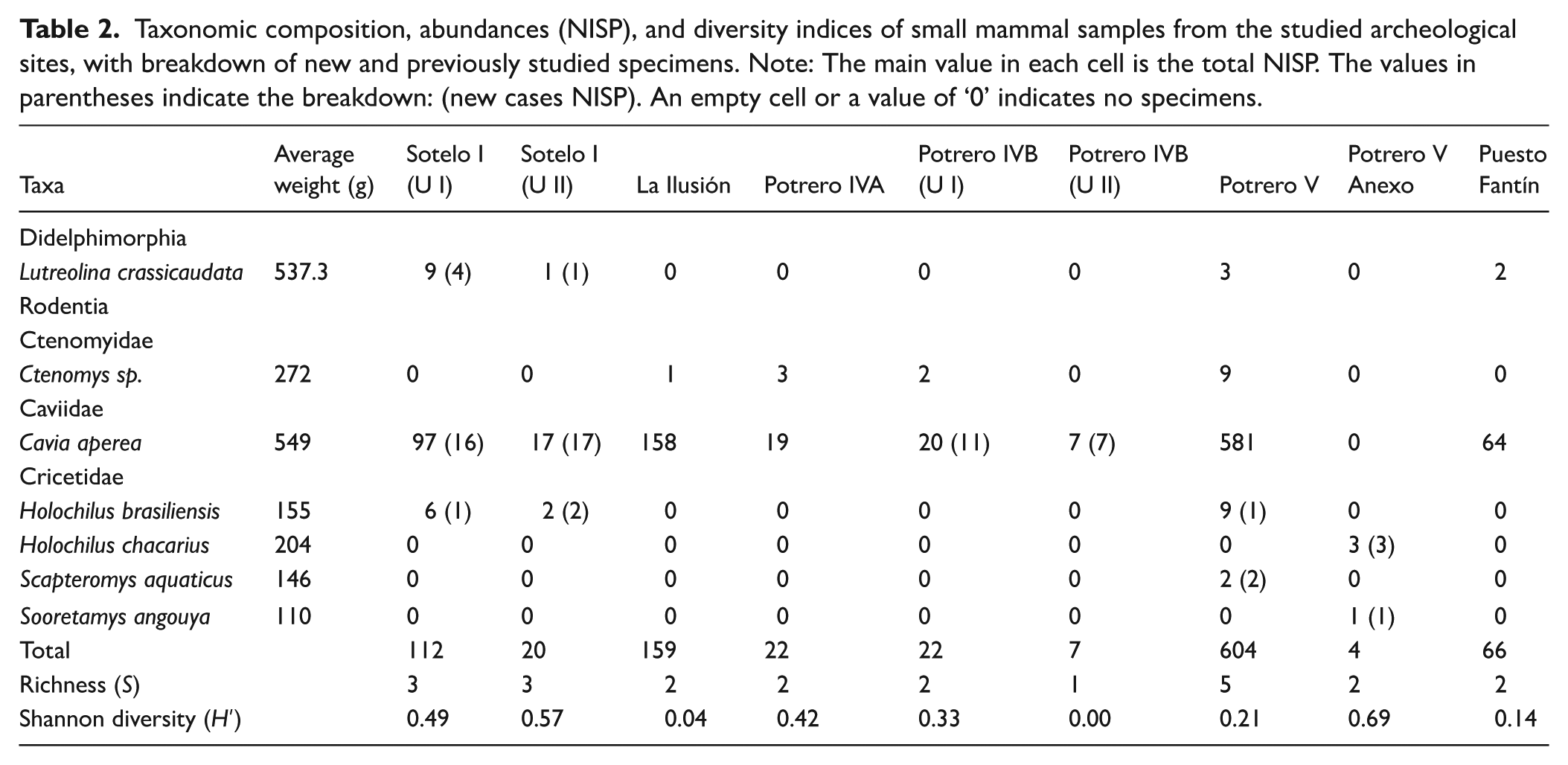
In this research, we analyzed 950 well-preserved small mammal Holocene specimens previously studied from the seven archeological sites (del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015; Zarza et al., 2019), together with 66 newly recovered specimens from Sotelo I (Units I and II), Potrero IVB (Units I and II), Potrero V, and Potrero V Anexo (Table 2). The inclusion of these new specimens adds three sigmodontine species to the small mammal record of the latter half of the Late-Holocene in the Humid Chaco region, thereby improving paleoenvironmental inferences.
Taxonomic composition, abundances (NISP), and diversity indices of small mammal samples from the studied archeological sites, with breakdown of new and previously studied specimens. Note: The main value in each cell is the total NISP. The values in parentheses indicate the breakdown: (new cases NISP). An empty cell or a value of ‘0’ indicates no specimens.
The present day small mammal communities in the surroundings of the archeological sites were characterized based on analyses of owl-pellet samples and on published trapping data (Supplemental Table 1; Kravetz et al., 1986; Massoia et al., 1988, 1990, 1995, 1997, 1999; Pardiñas et al., 2005; Teta et al., 2009; Udrizar Sauthier and Carrera, 2013).
Methods
Radiocarbon ages were calibrated using Calib 8.2 with the SHCal20 curve, expressed as 2σ cal BP, with the most probable time interval and probabilistic median (Hogg et al., 2020).
From a taphonomic perspective, we considered both, the action of predators (Montalvo and Fernández, 2019) and post-depositional processes—such as weathering, trampling, and manganese oxide impregnations—following Fernández-Jalvo and Andrews (2016, and references therein). The taxonomic identification and nomenclature of the specimens were established by comparing their anatomical features with fossil material described in the literature (e.g. Pardiñas and Teta, 2011; Patton et al., 2015, and references therein), as well as with reference collections housed at the Grupo de Estudios en Arqueometría, Facultad de Ingeniería (GEArq-FIUBA, Buenos Aires) and Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’ (MACN, Buenos Aires).
The number of identified specimens (NISP) was calculated following Lyman (1994). The Minimum Number of Individuals (MNI) was calculated based on the most frequent lateralized skeletal element for each taxon (Lyman, 1994). Paleoenvironmental analysis was carried out using the frequencies and presence/absence of some stenotopic species as indicators of environmental conditions (Andrews, 1990). Richness (S) and the Shannon diversity index (H′) were calculated based on MNI-derived relative abundances for each archeological site and on species counts for present day samples. Principal Component Analysis (PCA) was conducted using a correlation matrix based on the relative abundances of small mammal specimens from both present-day localities and archeological sites. The correlation matrix was selected to normalize the influence of dominant species and enhance the representation of rarer taxa. For this analysis, taxa were considered at the genus level to better visualize potential trends in the dataset. The statistical analyses were performed using the software PAST v. 5.02 (Hammer et al., 2001).
The Temporal Niche of Occurence (NOO3D) for Sooretamys angouya was calculated using both present and fossil localities compiled from the literature (Supplemental Tables 2 and 3). A total of 19 bioclimatic variables at a 5′ spatial resolution (approximately 9.3 × 9.3 km at the equator), representing both current and past conditions, were used as environmental predictors. Current environmental layers were downloaded from Climatologies at High Resolution for the Earth’s Land Surface Areas (CHELSA), average data from 1979 to 2013 (CHELSA, 2017; Karger et al., 2017). Paleoclimatic layers were obtained from the PaleoClim database created by Brown et al. (2018). Four chronozones were considered for predicting species distribution: the Late-Holocene (4.2 ka to present), the Mid-Holocene (8.2–4.2 ka), the Early-Holocene (11.7–8.2 ka), and the end of the Late-Pleistocene (13–11.6 ka). The NOO3D approach was applied to estimate the geographic extent of paleoclimatic areas whose environmental conditions are analogous to those currently occupied by Sooretamys angouya. Modeling was implemented using the freely available ModestR software (García-Roselló et al., 2019). NOO3D extends the NOO (Niche of Occurrence) method into a three-dimensional space defined by latitude, longitude, and time, overcoming three main limitations of other species distribution models (Lobo, 2016): (1) The absence of reliable data, often replaced by artificial background absences; (2) reliance on complex modeling techniques; (3) the selection of an arbitrary geographical extent for running models. The complete procedure is detailed in García-Roselló et al. (2019) with a step-by-step tutorial available at http://www.ipez.es/modestr/Manual_Tutorial.html. In summary, the methodology consists of four steps:
Defining the accessible area: The species’ environmental preferences are calculated based on river basins it currently occupies, ensuring connectivity among all considered occurrences (García-Roselló et al., 2019).
Selecting relevant predictors: Variables are chosen based on the Variation Inflation Factor (VIF), retaining only those with a VIF < 5 to minimize multicollinearity among predictors. This conservative threshold ensures that parameter estimates remain stable, robust, and interpretable (O’brien, 2007).
Screening predictors: Variables representing an accumulated percentage contribution of 80% in the variability of the selected predictors are retained (García-Roselló et al., 2019).
Calculating NOO3D: A Compound Environmental Layer (CEL) is created, integrating all relevant predictor data within the defined geographic extent. This CEL is used to determine the species’ current niche and project it onto past climatic scenarios. Default settings include a tolerance of 1% and a smoothing parameter of 3 (García-Roselló et al., 2019). The minimum density estimation in an occurrence cell serves as the threshold to delineate predicted suitable areas, generating binary maps for each scenario.
Results and discussion
The small mammal assemblages recovered from the archeological sites studied in the Humid Chaco are composed of seven taxa: a didelphid marsupial, two caviomorph rodents, and four sigmodontine rodents (Table 2). Except for the caviomorph Ctenomys sp., all the other small mammals that had previously been recorded (see the ‘Archeological sites’ section) were also identified in the new samples (Table 2). These included the marsupial Lutreolina crassicaudata (Figure 2(a)), the caviomorph Cavia aperea (Figure 2(b)), and the cricetid sigmodontine Holochilus brasiliensis (Figure 2(c)). The other three sigmodontine rodents listed in Table 2—Sooretamys angouya (Figure 2(d)), Scapteromys aquaticus (Figure 2(e)), and Holochilus chacarius (Figure 2(f))—were recorded here for the first time at the archeological sites studied. Among the four sigmodontine identified at the archeological sites, three are members of the tribe Oryzomyini, and one of the tribe Akodontini. Congruently, with 172 recognized species, Oryzomyini is the richest tribe of Sigmodontinae (497 species), followed by Akodontini (89 species), Thomasomyini (89 species), Phyllotini (65 species), and other poorly represented tribes (Brito and Pardiñas, 2025).
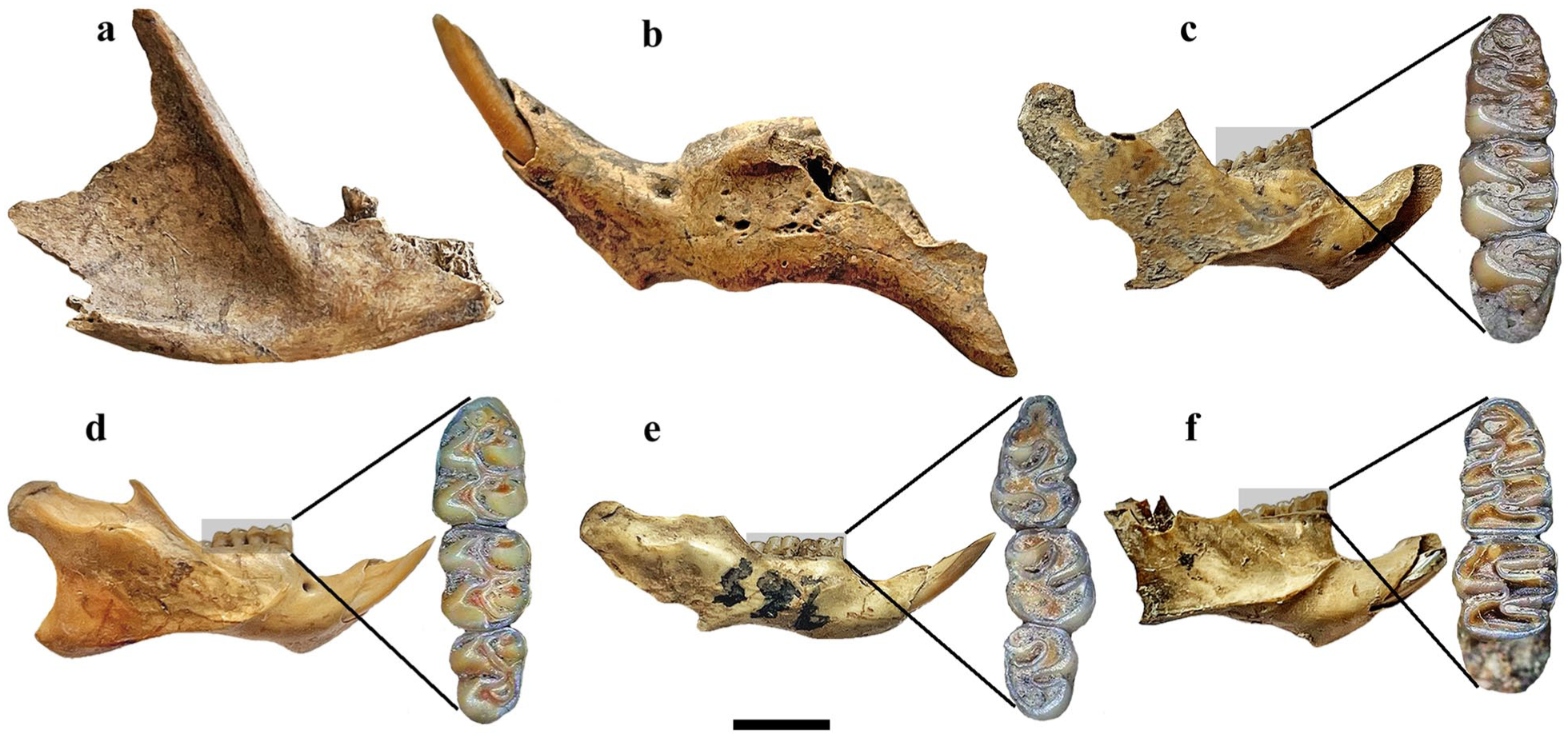
Taxonomic identification and taphonomy of the small mammals from the archeological sites of the Humid Chaco. (a) Lutreolina crassicaudata, fragment of right mandible with root marks (Sitio Sotelo I Unit I). (b) Cavia aperea, fragment of left mandible with root marks (Sotelo I Unit II). (c) Holochilus brasiliensis, fragment of left mandible (mirrored image) including a detail of its molar series in occlusal view (El Cachapé Potrero V). (d) Sooretamys angouya, complete right mandible with root marks, and its molar series (El Cachapé V Anexo). (e) Scapteromys aquaticus, fragment of right mandible with root marks, and its molar series (El Cachapé Potrero V). (f) Holochilus chacarius, fragment of left mandible (mirrored image) with manganese oxide rosette etching, and its corresponding molar series (El Cachapé V Anexo). Scale bar = 5 mm.
The predominance of Cavia aperea (NISP = 94.78%) likely reflects anthropic accumulation, as evidenced by the thermoalteration pattern documented in previous studies (del Papa and Lamenza, 2019; Santini, 2009; Santini and Lamenza, 2015; Zarza et al., 2019). None of these studies reported any taphonomic signatures associated with predation, such as digestive corrosion or bite and tooth marks, in marsupials (L. crassicaudata), ctenomyids (Ctenomys sp.), and sigmodontines (H. brasiliensis). Similarly, the 66 specimens evaluated here also showed no evidence of predator activity. Instead, postdepositional alterations—fragmentation in 65 specimens, root etching in 63, and a single case of manganese oxide impregnation—combined with a complete absence of weathering (Figure 2) point to trampling followed by rapid burial under humid, herbaceous conditions (Fernández-Jalvo and Andrews, 2016). Together with the ethological tendencies of species inhabiting forested levees, these taphonomic patterns support an in situ accumulation of the small-mammal remains (Fernández-Jalvo and Andrews, 2016).
The PCA results (Figure 3) support these observations, revealing a clear distinction between present day and archeological assemblages. The former are positioned along the positive values of PC1, which explains 24.81% of the total variance, whereas the latter are associated with negative values on PC2, accounting for 15.62% of the variance. Overall, present day assemblages accumulated by barn owls (Tyto furcata) exhibit higher taxonomic richness diversity compared to archeological assemblages (Table 2 and Supplemental Table 1). In contrast, the latter show a stronger correlation with the presence of Cavia, Ctenomys sp., and Lutreolina, with Cavia being the dominant taxon. Only one archeological site, El Cachapé Potrero V Anexo, is separated from the others, falling along the positive values of PC2. This indicates a closer affinity with present day assemblages and is also characterized by the presence of Sooretamys angouya.
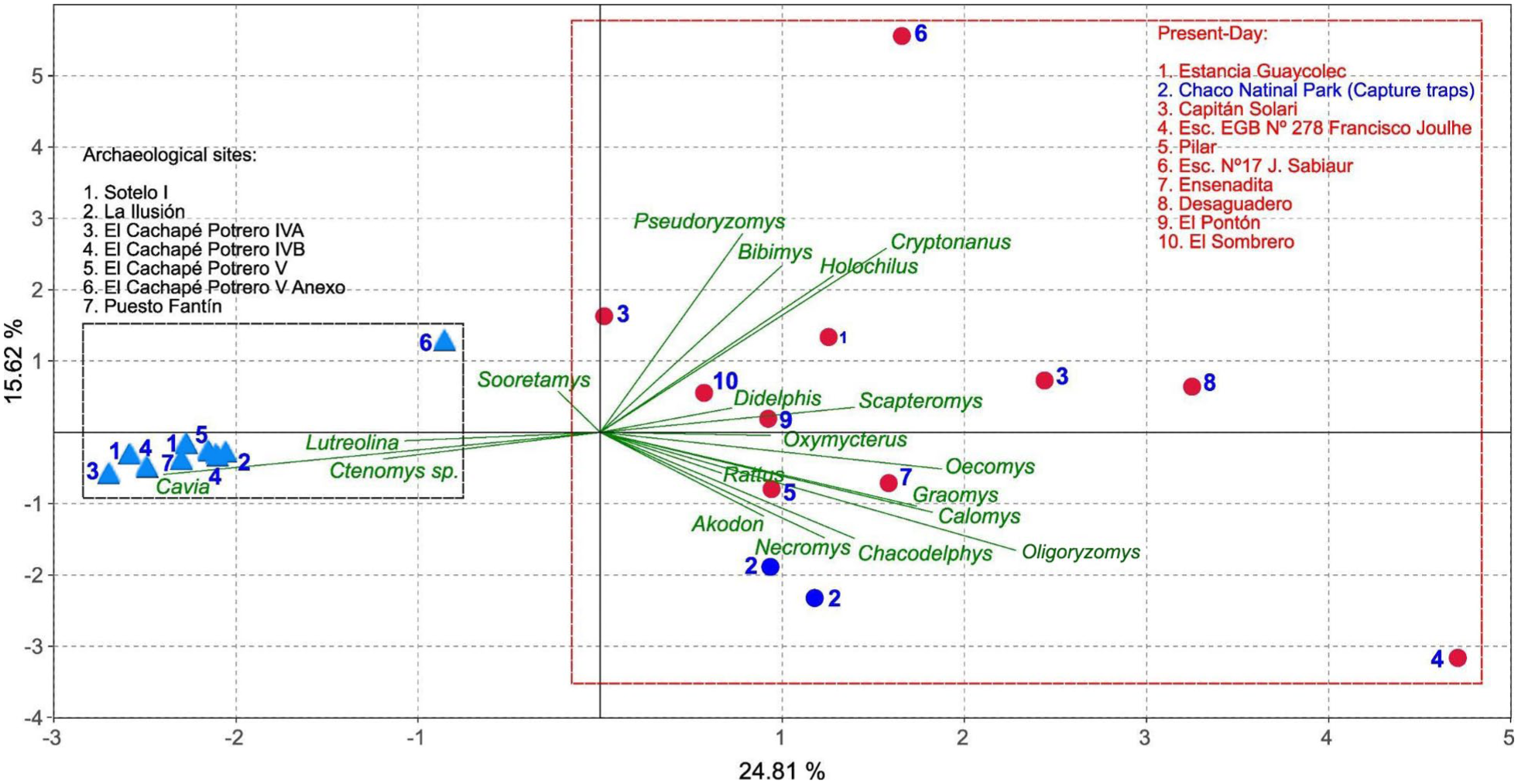
Principal Component Analysis (PCA) based on a correlation matrix, comparing the relative abundances of small mammal assemblages from present day localities (dots) and archeological sites (sky-blue triangles). Blue colors in present day localities are referred for trapped specimens and red colors to barn owl pellets.
Overall, the assemblages recovered from the archeological sites of the Humid Chaco indicate the presence of flooded environments (L. crassicaudata, H. brasiliensis, and H. chacarius), open grasslands and sandy areas (C. aperea and Ctenomys sp.), hydrophytic vegetation (S. aquaticus), and gallery forests (S. angouya; Hadler et al., 2025; Massoia, 1973a, 1973b; Massoia and Fornes, 1964; Massoia et al., 1999; Pardiñas et al., 2005; Pardiñas and Teta, 2005, 2011; Percequillo, 2015; Teta et al., 2009; Voglino et al., 2005). Most of these species (L. crassicaudata, H. brasiliensis, H. chacarius, S. aquaticus) are renowned for their swimming prowess and display morphological specializations for a semiaquatic habitat (Pardiñas and Cuellar Soto, 2023).
The finding of Sooretamys angouya at El Cachapé V Anexo, associated with a radiocarbon-calibrated date of approximately 0.9 ka BP (Table 1), is particularly significant, as it represents the westernmost limit of the species’ current distribution in Argentina (Figure 4(a)). To date, the fossil record of S. angouya has been limited to a few areas from the Quaternary of Brazil and Argentina (Figure 4 and Supplemental Table 3): (1) several caves in Lagoa Santa, Brazil—such as Lapa do Capão Secco, Lapa da Escrivania (caves 3, 5, and 11), Lapa da Quebra Chavelha, and Serra das Abelhas—excavated by Peter Lund (Pleistocene–Holocene; Boroni et al., 2018, Lund, 1836, Winge, 1887 [cited as Calomys rex, type locality]); (2) Gruta do Presépio in Santa Catarina, Brazil (4.6–1.3 ka cal BP and an undated historical unit; Hadler et al., 2024); (3) Garivaldino (10.7–8 ka cal BP and a Late-Holocene undated unit; Stutz et al., 2017), Pilger (7–3.1 ka cal BP; Hadler et al., 2016), and Sangão (5.2–4 ka cal BP; Stutz et al., 2020) in Rio Grande do Sul, Brazil; (4) Gruta Tres de Mayo in Misiones, Argentina (3.8–0.7 ka cal BP; Teta et al., 2009).
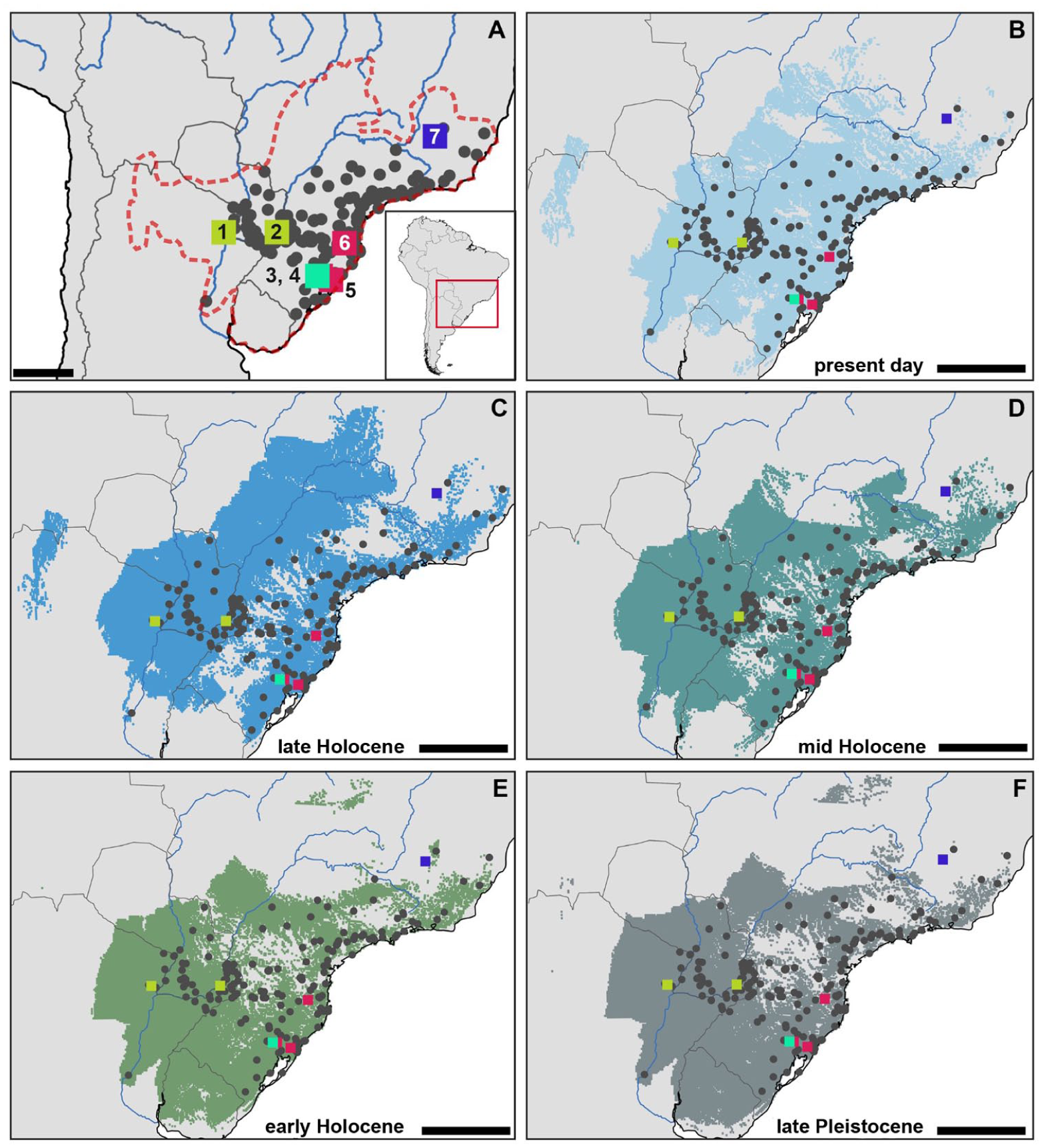
Predicted niche suitable distributions of Sooretamys angouya. (a) Area of the hydrobasins occupied (red dashed line) by the present day and fossil localities. (b) Predicted distribution under present day climatic conditions; (c) Distribution under Late-Holocene climatic conditions (4.2 ka BP-Present day). (d) Distribution under Mid-Holocene climatic conditions (8.2–4.2 ka BP). (e) Distribution under Early-Holocene climatic conditions (11.7–8.2 ka BP). (f) Predicted distribution during the end of the Late-Pleistocene (13–11.6 ka BP). Black dots: present day localities; color square: fossil localities (1. El Cachapé Potrero V Anexo; 2. Gruta Tres de Mayo; 3. Sangão; 4. Adelar Pilger; 5. Alfonso Garivaldino Rodriguez; 6. Gruta do Presepio; 7. Lagoa Santa). Paleoclimatic reconstructions (b–f) were generated using data from PaleoClim (Brown et al., 2018).
At present, this oryzomyine is widely distributed across eastern South America, from the Brazilian Atlantic rainforest to the humid forests of eastern Argentina and Paraguay. Additionally, S. angouya occurs in gallery forests of the Cerrado and in patches and remnants of semideciduous and gallery forests within the Humid Chaco (de Almeida Chiquito et al., 2014; Percequillo, 2015). Notably, neither extensive trapping in Chaco National Park nor analysis of owl-pellet assemblages from Capitán Solari—both localities situated roughly 60 km west of El Cachapé (Massoia et al., 1995; Teta et al., 2009) yielded any occurrences of S. angouya. These absences support the idea that S. angouya reaches the western limit of its distribution within the gallery forest zones of the Argentine Humid Chaco (Jayat et al., 2006; Teta et al., 2007; Udrizar Sauthier and Carrera, 2013).
Regarding the ecological niche modeling of S. angouya, 8 of the 19 bioclimatic variables were retained based on variance inflation factor (VIF) values (Table 3). Of these, five contributed cumulatively to 80% of the Instability Index, distinguishing environmental conditions at presence locations from those prevailing across the accessible area, defined by the hydrographic basin. These variables were: mean diurnal range, isothermality, precipitation of the coldest quarter, precipitation seasonality, and precipitation of the warmest quarter (Table 3). The distribution of S. angouya appears to be primarily explained by temperature-related variables (BIO 2 and 3) and seasonal precipitation patterns (BIO 19, 15, and 18; O’Donnell and Ignizio, 2012). All predicted chronozones overlapped with the niche derived from contemporaneous occurrences of S. angouya. Geographic variation in niche extent was minimal, though a northward expansion has been observed since the Early-Holocene, accompanied by a contraction in southern areas. Notably, although neither fossil nor contemporaneous records of S. angouya have been found in Uruguay, most of the country appears to have been occupied by the species during the Late-Pleistocene and Early-Holocene. Nonetheless, fossil records of S. angouya, regardless of chronozone, are consistently contained within the predicted niche range. These findings suggest that S. angouya has maintained stable environmental preferences over the time here analyzed, implying low phenotypic plasticity for the species (McGuire and Davis, 2013). In fact, as we mentioned, S. angouya has been recorded in fossil and present day sites across the extreme southern Brazil (Rio Grande do Sul) from the Early-Holocene to the present (Supplemental Tables 2 and 3), providing additional evidence of habitat suitability (rainforest) throughout the Holocene. Both pollen and small mammal data from these archeological sites indicate the continuous presence of forest in the extreme south of Brazil since the Early-Holocene. Although the Temporal Niche of Occurrence for S. angouya was estimated based on its climatic preferences, the results suggest that suitable climatic conditions have persisted throughout the analyzed period. This, in turn, also implies that vegetation, closely linked to climate, has remained relatively stable over time. The forest was initially present as gallery formations, followed by a significant expansion beginning around the Mid-to-Late-Holocene transition and continuing until historical times, when anthropic activities transformed the area into cropland and livestock zones (Fernández et al., 2019; Hadler et al., 2013, 2016, 2025; Stutz et al., 2017, 2020). Particularly, S. angouya was also found at these archeological sites alongside several other sigmodontine and caviomorph rodents, including forest taxa (Oecomys catherinae, Delomys sp., Juliomys sp., and Phyllomys sp.), as well as others associated with the Cerrado and Chaco biomes (Gyldenstolpia cf. G. planaltensis, Kunsia tomentosus, Pseudoryzomys simplex, Euryzygomatomys mordax†, Dicolpomys fossor†, and Clyomys riograndensis†). However, this latter species assemblage, more dependent on open habitats, disappears from the Late-Holocene fossil record. These findings, corroborated by palynological studies from Rio Grande do Sul, indicate a drier climate characterized by a mosaic of grassland-dominated regions interspersed with gallery and patch forests during the Early to Mid-Holocene. The subsequent increase in humidity during the Late-Holocene facilitated the expansion and consolidation of the Atlantic forest (e.g. Fernández et al., 2019; Hadler et al., 2013, 2016, 2025; Stutz et al., 2017, 2020). Finally, the absence of mismatches between niches derived from fossil and contemporary data suggests that anthropogenic processes have not significantly altered the current distribution of the species over the past centuries, which might otherwise have resulted in such discrepancies between the contemporary and fossil niches (Martínez-Meyer et al., 2004; McGuire and Davis, 2013).
Environmental variables selected based on their VIF values and their contribution to the Instability Index.

The asterisk (*) indicate the predictors that account for a cumulative 80% contribution to the Instability Index for the accessible area of the species.
The convergence of multiple paleoenvironmental proxies reinforces the hypothesis of environmental stability in the Humid Chaco during the Late-Holocene. The small mammal assemblages, dominated by taxa associated with flooded environments and gallery forests, indicate persistent humid conditions between 1600 and 600 years BP. This interpretation is supported by the enlarged dental plates of Lepidosiren paradoxa recovered from the same archeological sites, which suggest increased water availability and rising temperatures since approximately 1600 years BP (Zarza et al., 2025). Furthermore, the temporal niche modeling of Sooretamys angouya demonstrates that suitable climatic conditions for gallery forest species have persisted in the region since at least the Late-Pleistocene, with minimal geographic variation across chronozones (Figure 4). Fossil pollen and small-mammal data from southern Brazil corroborate a regional trend toward increased humidity during the Late-Holocene, marked by the expansion of Atlantic forest formations (Fernández et al., 2019; Hadler et al., 2013, 2016, 2025). Taken together, these independent lines of evidence suggest that the fluvial systems and associated gallery forests of the Humid Chaco have remained functionally stable throughout the period of human occupation examined here, providing a predictable resource base for hunter-gatherer-fisher societies.
Although the fossil record of amphibious rats Holochilus is abundant in the South American lowlands throughout the Quaternary (e.g. Massoia, 1971; Pardiñas and Teta, 2011), the presence of H. brasiliensis and H. chacarius at El Cachapé indicates that their current sympatric distribution in the Humid Chaco, nearby savannas, the Iberá–Ñeembucú system, and the Paraná Delta (D’Elía et al., 2008; Massoia, 1971; Pardiñas et al., 2005; Teta and Pardiñas, 2010; Voglino et al., 2005) was established during the Late-Holocene.
Except for Ctenomys sp., all other small mammal species found at the archeological site of Humid Chaco studied here were also recorded in the locality of Capitán Solari (Massoia et al., 1995; Teta et al., 2009). The absence of Ctenomys in the present-day pellet sample could be related to the fact that this genus, composed of strictly fossorial and primarily diurnal species, is not a typical prey for nocturnal or crepuscular raptors such as Tyto furcata (e.g. Bellocq, 2000). In this area, three species of Ctenomys are currently recognized: C. bonettoi, C. yolandae and C. argentinus, while other species, such as C. pilarensis, C. perrensis, and C. roigi, can also be found in nearby areas (Bidau, 2015). C. benettoi is known only in a few communities inhabiting sandy patches within periodically flooded areas surrounding the type locality, 7.5 km from Capitán Solari (Contreras and Berry, 1982). C. yolandae occurs in both sandy soils within open forests and in flooded or anthropized areas (Contreras and Berry, 1984). The Chacoan-endemic C. argentinus, which inhabits sandy and grassy soils, represents a biogeographic and evolutionary model linked to ENSO precipitation cycles, driving expansion and contact in droughts, and retraction and isolation in floods (Contreras and Berry, 1985). The notable absence in the Humid Chaco of other small mammals common such as Akodon montensis, Bibimys chacoensis, Calomys callosus, Chacodelphys formosa, Cryptonanus chacoensis, Graomys chacoensis, Necromys lasiurus, Oecomys franciscorum, Oligoryzomys fornesi, Oligoryzomys nigripes, and Pseudoryzomys simplex (e.g. Hershkovitz, 1962; Massoia et al., 1995, 1997; Pardiñas et al., 2005; Pardiñas and Teta, 2005; Shamel, 1931; Teta et al., 2009; Teta and Pardiñas, 2010; Udrizar Sauthier and Carrera, 2013) could be associated with taphonomic formation processes. As we demonstrated, most of the specimens belong to C. aperea, which were deposited by humans as food waste (del Papa and Lamenza, 2019; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015; Zarza et al., 2019). The lower richness and diversity observed in the archeological samples compared to the present day samples is associated with the markedly higher abundance of Cavia aperea in the former, to the detriment of smaller species that are more frequent in the owl’s diet (Table 2 and Supplemental Table 1). Non-predatory natural agents through a taphonomic process of in situ accumulation accumulated the remaining species evaluated in this study. As a result, marsupials and sigmodontines are proportionally underrepresented, and the assemblages may not fully reflect the local biodiversity.
Other paleoenvironmental proxies support the establishment of humid conditions in the Humid Chaco toward the end of the Late-Holocene (Iriondo, 2006; Lamenza, 2013; Zarza et al., 2019, 2025). Notably, temperature and humidity rose from 1600 years BP to the present—peaking around 1000 years BP and likely linked to a mega–El Niño event (Meggers, 1994)—as indicated by the enlarged dentary plates of the lungfish Lepidosiren paradoxa (Dipnoi) recovered from the studied archeological sites (Zarza et al., 2025). This paleoenvironmental evidence supports the idea that human groups in the Humid Chaco adapted their fishing, hunting, gathering, and settlement practices (including longer-term residences on levees) to optimize the use of riverine resources—both small and large prey—thereby reducing search time and transportation costs in response to climatic cycles and increased precipitation. Additionally, the flat topography of the Humid Chaco likely facilitated the development of a hydrographic network with lentic environments seasonally connected to lotic channels, all rich in food resources. These included local plants, mollusks (Asolene sp., Diplodon sp., Pomacea sp.), fishes (Lepidosiren paradoxa, Pimelodus sp., Pseudoplatystoma sp., Pterodoras sp., Synbranchus sp.), reptiles (Caiman sp., Eunectes notaeus, Hydrodynastes gigas), birds (Anas sp., Jabiru mycteria, Rhea sp.), and mammals adapted to a mosaic of aquatic habitats, forests, and grasslands. The mammals include large species (Blastoceros, Mazama, Ozotoceros), medium-sized species (Myocastor), and small-sized ones (Cavia and Holochilus; del Papa and Lamenza, 2019; Lamenza, 2013; Santini, 2009; Santini and De Santis, 2011; Santini and Lamenza, 2015; Zarza et al., 2019).
Conclusion
The multi-proxy analysis of small mammal assemblages from open-air archeological sites in the Argentinean Humid Chaco provides critical insights into the environmental and cultural dynamics of the Late-Holocene (ca. 1600–600 cal BP). Our results lead to the following key conclusions:
In summary, the small mammal record of the Humid Chaco reveals long-term persistence in biodiversity and habitat configuration. These findings emphasize the role of South American fluvial systems as stable environmental anchors, providing a baseline for understanding current biodiversity patterns and the historical resilience of human societies in the face of Holocene climate variability.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836261458259 – Supplemental material for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers
Supplemental material, sj-xlsx-1-hol-10.1177_09596836261458259 for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers by Fernando Julián Fernández, Sara García-Morato, María Macarena Zarza, Luis Manuel del Papa and Guillermo Nicolás Lamenza in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_09596836261458259 – Supplemental material for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers
Supplemental material, sj-xlsx-2-hol-10.1177_09596836261458259 for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers by Fernando Julián Fernández, Sara García-Morato, María Macarena Zarza, Luis Manuel del Papa and Guillermo Nicolás Lamenza in The Holocene
Supplemental Material
sj-xlsx-3-hol-10.1177_09596836261458259 – Supplemental material for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers
Supplemental material, sj-xlsx-3-hol-10.1177_09596836261458259 for Late-Holocene small mammals in the Humid Chaco of northern Argentina: Evidence of environmental stability in fluvial contexts inhabited by hunter-gatherer-fishers by Fernando Julián Fernández, Sara García-Morato, María Macarena Zarza, Luis Manuel del Papa and Guillermo Nicolás Lamenza in The Holocene
Footnotes
Acknowledgements
We are grateful to Susana Salceda† and Horacio Calandra†, former directors, who were key proponents of archeological research in the area. Their contributions laid the foundation for ongoing scholarly investigations. Mariano Santini also contributed to the initial sorting and analysis of the zooarchaeological remains. Pablo Teta, curator at MACN (Buenos Aires), provided the comparative specimens of Sooretamys angouya used for taxonomic identification. We appreciate the valuable feedback provided by the three reviewers, which has helped us to improve the clarity of this work.
ORCID iDs
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Fieldwork and laboratory analyses were funded by three grants: from the Ministerio de Ciencia, Innovación y Universidades of Spain (i-COOPB24012), Universidad de La Plata of Argentina (11/N983), and Consejo Nacional de Investigaciones Científicas y Técnicas (PIP-CONICET-0001). SGM holds a ‘Juan de la Cierva’ postdoctoral contract at the Instituto de Historia, funded by the Ministerio de Ciencia e Innovación of Spain (JDC2023-051162-I).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.