
Abstract
Introduction
Sulfur mustard (SM) as a potent alkylating chemical warfare agent, results in long-lasting health issues. This study aims to evaluate the associations among polymorphisms, the expression levels of DNA repair-related genes, oxidative stress, and the severity of sulfur mustard toxicity.
Methods
XPC (Xeroderma pigmentosum group C) rs2228001 and XRCC1 (X-ray repair cross-complementing protein 1) rs25487 were genotyped in three subgroups of 545 SM-exposed participants (asymptomatic, mild, and moderate-severe). Also, their transcriptional changes were evaluated.
Results
In the codominant model (AA reference), the GG genotype showed a negative association with the moderate-severe subgroup when compared to the mild subgroup. The AG genotype showed negative associations with the moderate-severe subgroup compared to both the asymptomatic and mild subgroups. In the recessive model (AA reference), the combined GG + AG genotypes only showed a negative association for the moderate-severe subgroup compared to the mild subgroup, suggesting that these genotypes may be associated with a lower risk of severe clinical signs. The XPC transcript decreased in the SM-exposed group and moderate-severe subgroup compared to the non-exposed group, while the XRCC1 transcript increased, especially in the mild and moderate-severe subgroups. This increase in XRCC1 transcript was only in the individuals with GG and AG genotypes. There was a negative correlation between XPC transcript and malondialdehyde (indicator of lipid peroxidation) levels.
Discussion
Variations in XRCC1 genotypes and expression of XRCC1/XPC gene are associated with responses to SM exposure and the risk of developing moderate-severe clinical signs, highlighting the complex interplay between oxidative stress and DNA repair.
Introduction
Sulfur mustard (SM) is a chemical warfare agent known for its harmful effects on human health. 1 The toxic mechanism of SM involves the formation of reactive metabolites that interact with cellular macromolecules, leading to the disruption of normal physiological processes. 2 Upon expossure, SM can induce acute clinical signs such as skin blistering, respiratory distress, and ocular damage, while long-term complications may include chronic respiratory diseases, skin and eye complications, and cancers.2–4 At the cellular level, exposure to SM triggers significant DNA damage, primarily through its alkylating effects and the generation of oxidative stress and inflammatory responses.5,6 As an alkylating agent, SM forms reactive metabolites that can covalently modify DNA, leading to the formation of DNA adducts, interstrand cross-links, and strand breaks. These modifications compromise genomic integrity and disrupt normal cellular processes. 7 The reactive species produced during the metabolism of SM not only damage DNA directly but also activate inflammatory pathways that can exacerbate cellular injury and promote further genomic instability.8,9 The accumulation of such DNA damage poses a serious threat to cellular function, potentially leading to mutations, apoptosis, and the development of long-term health complications. 10
To mitigate the effects of DNA damage, cells rely on a sophisticated network of DNA repair mechanisms essential for maintaining genomic stability and cellular health. Among the key players in this intricate repair system are the XRCC1 (X-ray repair cross-complementing protein 1) and XPC (Xeroderma pigmentosum group C) genes, which perform critical functions in different DNA repair pathways.11,12 XRCC1 is a vital component of the base excision repair (BER) pathway, responsible for repairing single-strand breaks and small base modifications that can occur as a result of alkylation, oxidative stress, and other forms of DNA damage. 13 XRCC1 acts as a scaffold protein, facilitating the recruitment of various DNA repair enzymes, including DNA polymerase and ligase, to the site of damage. The N-terminal domain of XRCC1 binds to DNA polymerase β, which is essential for filling gaps in DNA during repair, while its C-terminal domain interacts with DNA ligase IIIα, which seals the breaks in DNA strands. The central domain of XRCC1 contains a poly (ADP-ribose) binding motif, allowing it to interact with Poly [ADP-ribose] polymerase 1(PARP1), which is activated at sites of DNA damage. PARP1 is a first responder that detects DNA damage and then facilitates repair pathway choice. This coordinated action of XRCC1 ensures that damaged bases are excised and replaced, restoring the integrity of the DNA strand.14,15 The efficiency of XRCC1 in executing these repair processes is crucial, as any deficiencies may lead to the accumulation of mutations and increased susceptibility to genotoxic agents, including SM.16,17 On the other hand, XPC plays a pivotal role in the nucleotide excision repair (NER) pathway, a DNA repair mechanism that removes bulky, helix-distorting lesions such as those caused by UV radiation or chemical agents. 18 (XP)C is responsible for the initial detection of DNA damage, particularly helix-distorting lesions, and facilitates the recruitment of other repair factors to initiate the excision of the damaged DNA segment. 19 This pathway is particularly important for addressing the types of DNA damage that SM can induce, as the agent can create complex lesions that disrupt the normal structure of the DNA helix.20,21
Oxidative stress can significantly impact the expression and function of both XRCC1 and XPC. During inflammatory responses, the production of reactive oxygen species (ROS), highly reactive molecules containing oxygen, increases, leading to oxidative stress that can damage cellular components, including DNA. Elevated levels of ROS can directly affect the expression of DNA repair genes, including XRCC1 and XPC.22,23 Oxidative stress can induce the activation of transcription factors that may either upregulate or downregulate the expression of these genes, depending on the cellular context and the extent of damage. 24
Moreover, oxidative stress can impair the functional capacity of XRCC1 and XPC. For XRCC1, the presence of excessive oxidative damage may overwhelm its repair capabilities, leading to a backlog of unrepaired DNA lesions. This can result in a vicious cycle where the accumulation of DNA damage further exacerbates oxidative stress, ultimately compromising cellular integrity. 25 Similarly, XPC’s ability to recognize and repair bulky lesions may be hindered by oxidative modifications, reducing its efficiency in initiating the NER process. 26
Polymorphisms in the XRCC1 and XPC genes can significantly influence their expression levels and functional capabilities. Variations in these genes may lead to altered protein function, reduced repair efficiency, or changes in the cellular response to DNA damage. Such genetic variations can affect an individual’s capacity to effectively repair DNA damage, thereby modulating their susceptibility to the toxic effects of SM and other genotoxic agents.27–29 Certain polymorphisms may result in a less efficient repair process, increasing the likelihood of persistent DNA damage and subsequent cellular dysfunction. 30 While Next-Generation Sequencing (NGS) has revealed thousands of polymorphisms in the XRCC1 and XPC genes, 31 the selection of XRCC1 rs25487 and XPC rs2228001 for this study was based on their well-documented functional relevance and prior evidence of association with DNA repair efficiency and oxidative stress responses.32,33
This study aims to evaluate the expression and polymorphism of the XRCC1 and XPC genes and their association with the severity of SM toxicity and lipid peroxidation. We hypothesize that specific polymorphisms in XRCC1 and XPC genes may lead to differential expression levels and functional alterations, thereby influencing the severity of SM toxicity. Specifically, we expect that individuals with certain genotypes may exhibit different DNA repair capacity, which could either mitigate or exacerbate the effects of SM-induced oxidative stress and DNA damage.
Materials and methods
Study population and classifications
All participants provided written informed consent voluntarily as part of the “Isfahan and Sardasht-Iran cohort study of chemical warfare victims.” Detailed information about the study design and methodology, including the criteria for inclusion and exclusion, has been published previously. 34 Individuals with a history of systemic diseases before exposure (based on medical records), those suffering from acute infectious diseases at the time of sampling, or those who had long-term exposure to other chemical agents (e.g., industrial chemicals, radiation, or other hazardous substances) were excluded from the study.
A total of 701 participants were selected based on these criteria, comprising two groups of volunteers. The first group, the SM-exposed group, included 545 individuals who had been exposed to SM on a single occasion during the Iraq-Iran war in 1987. The second group, the non-exposed group, consisted of 156 healthy individuals with no history of SM exposure, matched for similar life conditions to the exposed group. Participants were categorized into three subgroups based on the severity of their SM exposure: The asymptomatic subgroup (n = 275) comprised participants who had experienced only minor acute effects (e.g., minor skin lesions or slight eye irritation) at the time of wartime exposure and who showed no residual clinical signs at the time of sampling; the mild subgroup included 130 individuals; and the moderate-severe subgroup comprised 140 individuals. The pulmonary function of participants was evaluated using spirometry (Chest 801 Spirometry) in accordance with American Thoracic Society criteria. Key parameters assessed included forced expiratory volume in one second (FEV1 % predicted), a measure of lung function that quantifies the amount of air a person can exhale forcefully in one second; forced vital capacity (FVC % predicted), a measure of lung function that quantifies the total amount of air a person can exhale after a deep inhalation; and the FEV1/FVC% ratio, along with pulmonary auscultation and clinical sign evaluation by pulmonary specialists. Patients with 65≤ FEV1 <80 % predicted or 65≤ FVC <80 % predicted were predicted to have mild, 40≤ FEV1 <65 % predicted or 40≤ FVC <65 % predicted were predicted to have moderate-severe lung lesions. Participants with FEV1 ≥ 80% predicted and FVC ≥ 80% predicted were classified as asymptomatic. Individuals with FEV1 < 40% predicted or FVC < 40% predicted were included in the moderate–severe subgroup.35–38
Genomic DNA extraction and rs2228001 (XPC) and rs25487 (XRCC1) genotyping
A whole blood sample (5 ml) was obtained from each SM-exposed participant and collected in vacuum blood collection tubes coated with EDTA. The whole blood samples were stored at 4°C. Genomic DNA was isolated from the whole blood using the salting-out method. The salting-out DNA extraction method was set up and optimized by the Immunoregulation Research Center of Shahed University. The concentration and purity (260/280 ratio) of the genomic DNA were evaluated by NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA).
The sequence and characteristics of primers and annealing conditions used for genotyping and gene expression analysis.
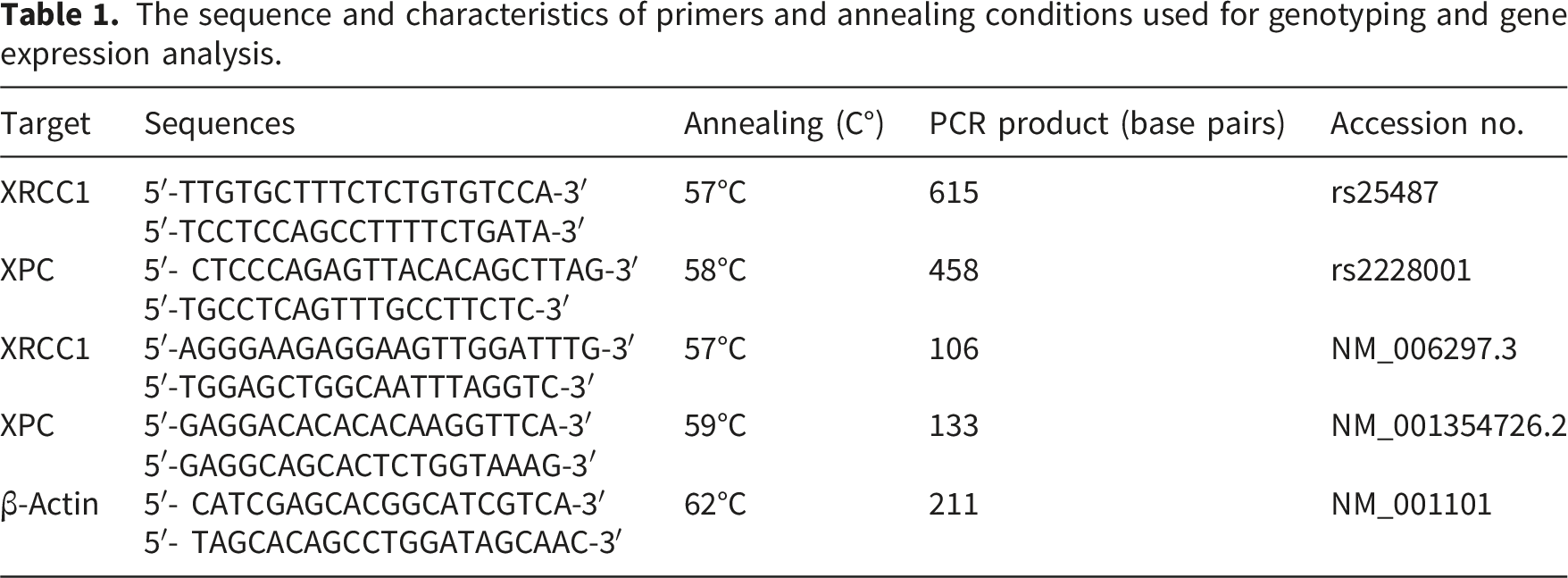
RNA extraction, cDNA synthesis, quantitative real-time PCR
After the separation of buffy coat from EDTA-treated blood samples, total RNA was extracted from a subset of participants (n = 138 from the SM-exposed group and n = 37 from the non-exposed group) using GeneAll® Hybrid-R™ kit (GeneAll Biotechnology Co., Ltd., Seoul, South Korea) according to the manufacturer’s instructions. This subset was selected consecutively based on the availability of high-quality RNA samples and was representative of the full cohort’s severity distribution: asymptomatic (n=63), mild (n=42), and moderate-severe (n=33). The quality and integrity of RNA samples were evaluated using the Nanodrop spectrophotometer, Only RNA samples representing an A260/A280 ratio of 1.8–2.2 were suitable for cDNA synthesis, a DNA copy synthesized from an RNA template using reverse transcriptase. Subsequently, 1µL of total RNA was reverse transcribed into complementary DNA (cDNA) utilizing a High-Capacity cDNA Reverse Transcription Kit (ABI, USA) in accordance with the manufacturer’s instructions. The synthesized cDNA was stored at −20°C until further analysis. Quantitative real-time PCR (qRT-PCR), a method used to amplify and quantify DNA or RNA in real time was performed in a total reaction volume of 20 µL, which included 10 µL of master mix (RealQ Plus 2x Master Mix Green), 2 µL of 1:10 diluted cDNA, 5 pM of each primer (XPC, XRCC1, and β-Actin) and 6 µL of RNase-free water. The specific primers are represented in Table 1. The optimal thermal cycling conditions were as follows: initial denaturation at 95°C for 16 minutes, followed by 40 cycles consisting of denaturation at 95°C for 30 seconds, annealing at 57 - 62°C for 60 seconds, and extension at 72°C for 30 seconds. All samples were analyzed in triplicate. β-Actin mRNA served as an internal control gene. No-template controls and no-reverse transcriptase controls were included in each PCR reaction to ensure specificity. Melt curve analysis was performed to confirm the specificity of the PCR products for each target gene. An aliquot from each reaction mixture was subjected to electrophoresis on a 2% agarose gel and stained with GelRed™ to visualize specific bands corresponding to each gene. Amplification efficiencies were quantified using the standard curve method for all target genes, with correlation coefficients (R2) exceeding 0.997. Relative gene expression levels (target/β-Actin) were calculated according to the Livak method. 39
Serum malondialdehyde (MDA) measurement
The serum MDA level, a marker of lipid peroxidation and oxidative stress, was assessed using the thiobarbituric acid reactive substances (TBARS) assay, as described previously. 40 The reliability of this method has been validated in earlier studies in SM-exposed individuals.41,42 In brief, 0.1 ml of fasting blood serum was mixed with 0.2 ml of 20% trichloroacetic acid and centrifuged at 2000g for 10 minutes. The resulting pellet was washed once with 0.5 ml of 50 mM sulfuric acid and then heated in a boiling water bath for 30 minutes in the presence of TBA reagent (0.2% in 2 M sodium sulfate) and 50 mM sulfuric acid. The lipid peroxide adducts formed were subsequently extracted using n-butanol, and the absorbance was measured at 530 nm. The concentration of MDA was calculated using a standard curve generated from MDA standard solutions (1,1,3,3-tetraethoxypropane in 50 mM H2SO4), with results reported in μmol/L.
Figure 1 represents the classification of participants and measured parameters. Classification of participants and measured parameters.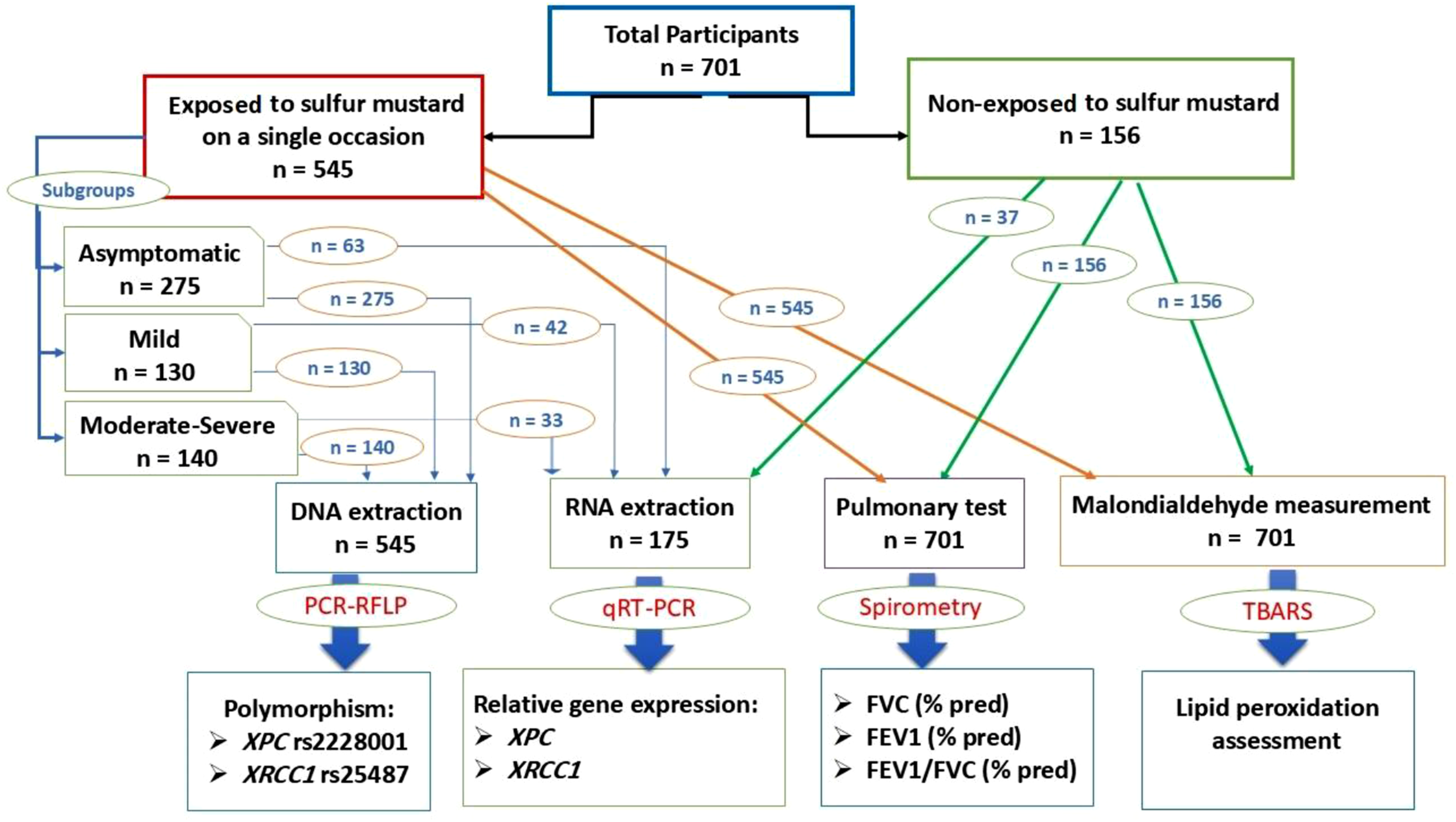
Statistical analyses
Data are presented as mean ± standard deviation (SD). The association between different genotypes and the severity of participants exposed to SM was assessed using odds ratios (OR) and 95% confidence intervals (CI) through logistic regression across various genetic models. An OR less than 1 indicates a negative association. Logistic regression models were performed to assess associations between XRCC1 and XPC genotypes and the severity of clinical signs in SM-exposed participants. Given the categorical nature of the severity subgroups (asymptomatic, mild, and moderate–severe), multinomial logistic regression was employed to evaluate genotype effects across all severity levels simultaneously, with the asymptomatic subgroup as the reference category. For pairwise comparisons, results were derived from the same multinomial model. All regression models were unadjusted for covariates such as age, smoking status, or comorbidities. This decision was based on preliminary analyses showing no significant differences in age or smoking status across severity subgroups, and because the primary aim was to explore genotype-severity associations without confounding by these factors in this hypothesis-generating study. Interaction terms were not included, and collinearity was not assessed given the limited set of predictors. Pearson linear regression was also performed to assess correlations of parameters. We utilized the online SNPStats software to evaluate deviations from Hardy-Weinberg equilibrium (HWE), a principle stating that allele and genotype frequencies in a population will remain constant from generation to generation in the absence of evolutionary influences. Genetic association analyses for XRCC1 and XPC polymorphisms were performed only in SM-exposed participants to assess genotype-severity relationships, while the non-exposed group served for baseline comparisons of gene expression, oxidative stress, and demographics. The expression levels of the XPC and XRCC1 genes, stratified by genotype or in comparison between subgroups, were compared using an independent t-test and one-way ANOVA (Analysis of Variance) after confirming data normality. Chi-square tests of independence were also used to assess the relationship between smoking status and SM exposure. Data analysis was conducted with SPSS software (version 26.0, SPSS Inc., Chicago, IL, USA). P < 0.05 was considered statistically significant.
Given the exploratory nature of the genetic association analyses across multiple inheritance models, no formal correction for multiple testing was applied; therefore, the findings should be considered hypothesis-generating and warrant validation in independent cohorts.
Results
Characteristics of participants
The study comprised two distinct analyses with overlapping participants: a polymorphism analysis on all eligible SM-exposed individuals, and a gene expression analysis on a consecutively selected subset with available high-quality RNA samples.
Characteristics of SM-exposed participants in polymorphism and gene expression analyses.
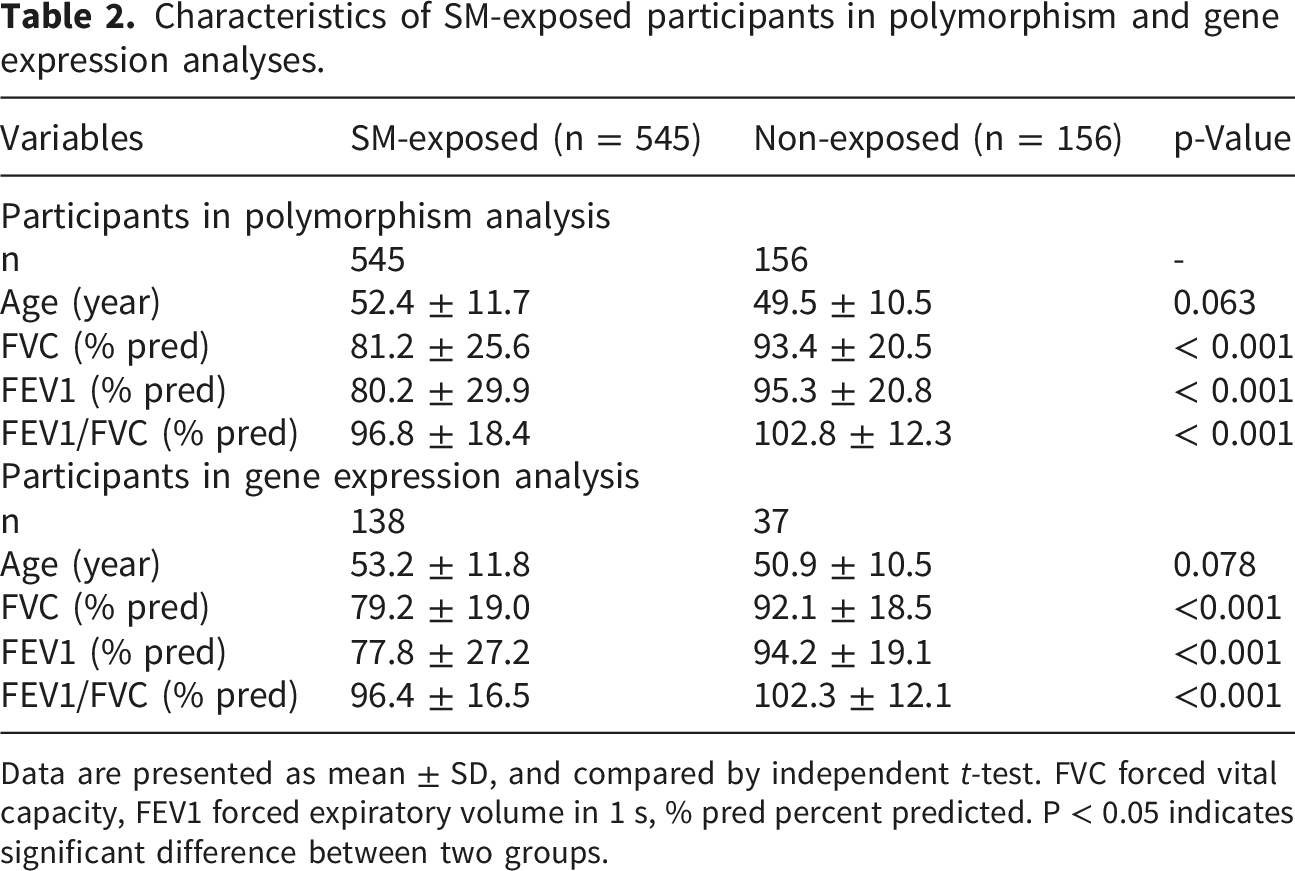
Data are presented as mean ± SD, and compared by independent t-test. FVC forced vital capacity, FEV1 forced expiratory volume in 1 s, % pred percent predicted. P < 0.05 indicates significant difference between two groups.
Characteristics of SM-exposed participants in polymorphism and gene expression analyses.
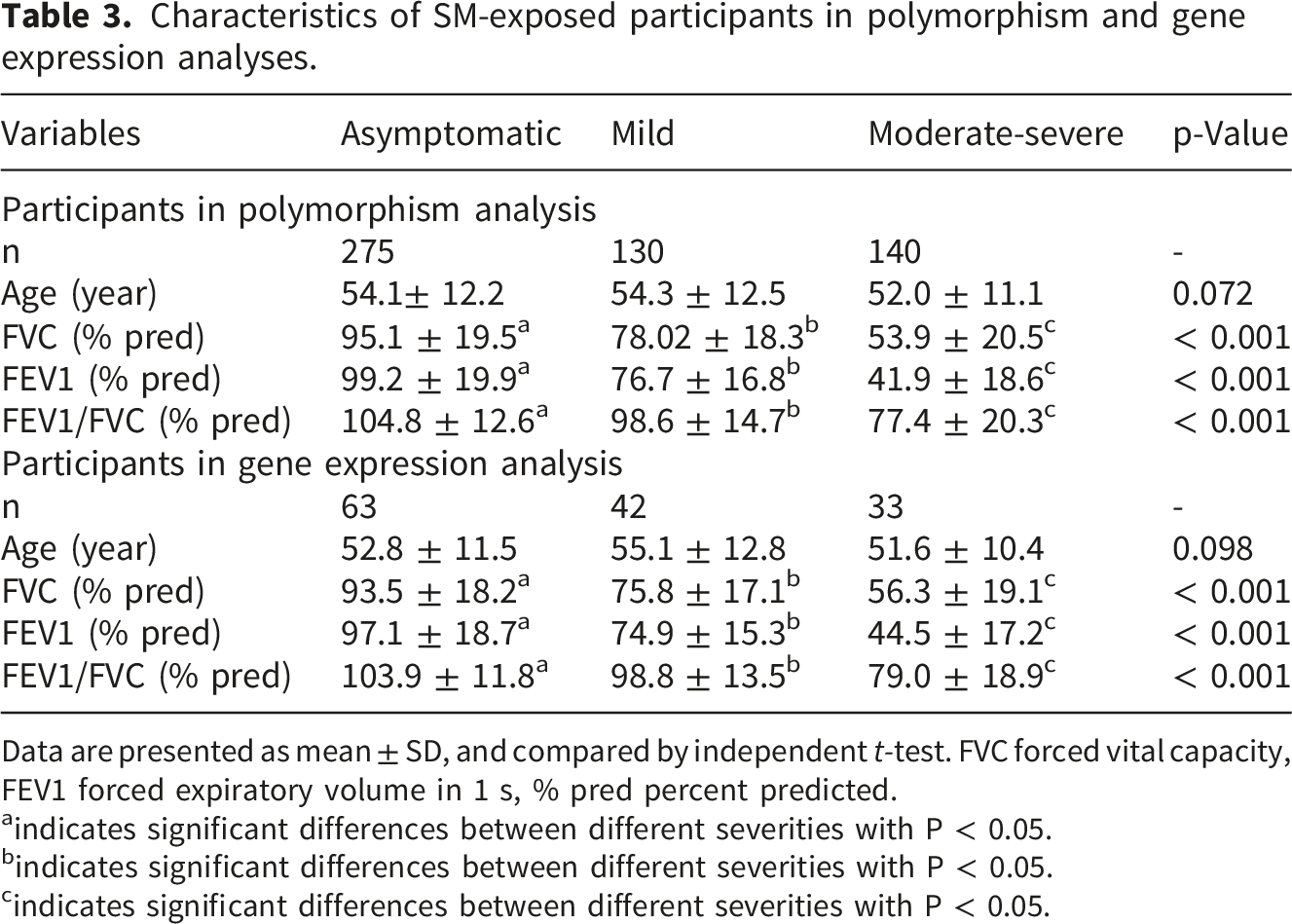
Data are presented as mean ± SD, and compared by independent t-test. FVC forced vital capacity, FEV1 forced expiratory volume in 1 s, % pred percent predicted.
aindicates significant differences between different severities with P < 0.05.
bindicates significant differences between different severities with P < 0.05.
cindicates significant differences between different severities with P < 0.05.
The analysis of frequency data revealed that smoking status (with 6 smokers and 150 non-smokers in the control group, and 8 smokers and 537 non-smokers in the SM-exposed group) was not significantly associated with SM exposure (X2 = 3.77, P > 0.05).
Evaluation of XRCC1 (rs25487, A > G) polymorphisms
According to online SNPStats software analysis, the frequency of this XRCC1 (rs25487) SNP was consistent with Hardy-Weinberg equilibrium (P > 0.05).
The association of XRCC1 rs25487 between SM-exposed subgroups with different severities under hereditary models’ analysis.
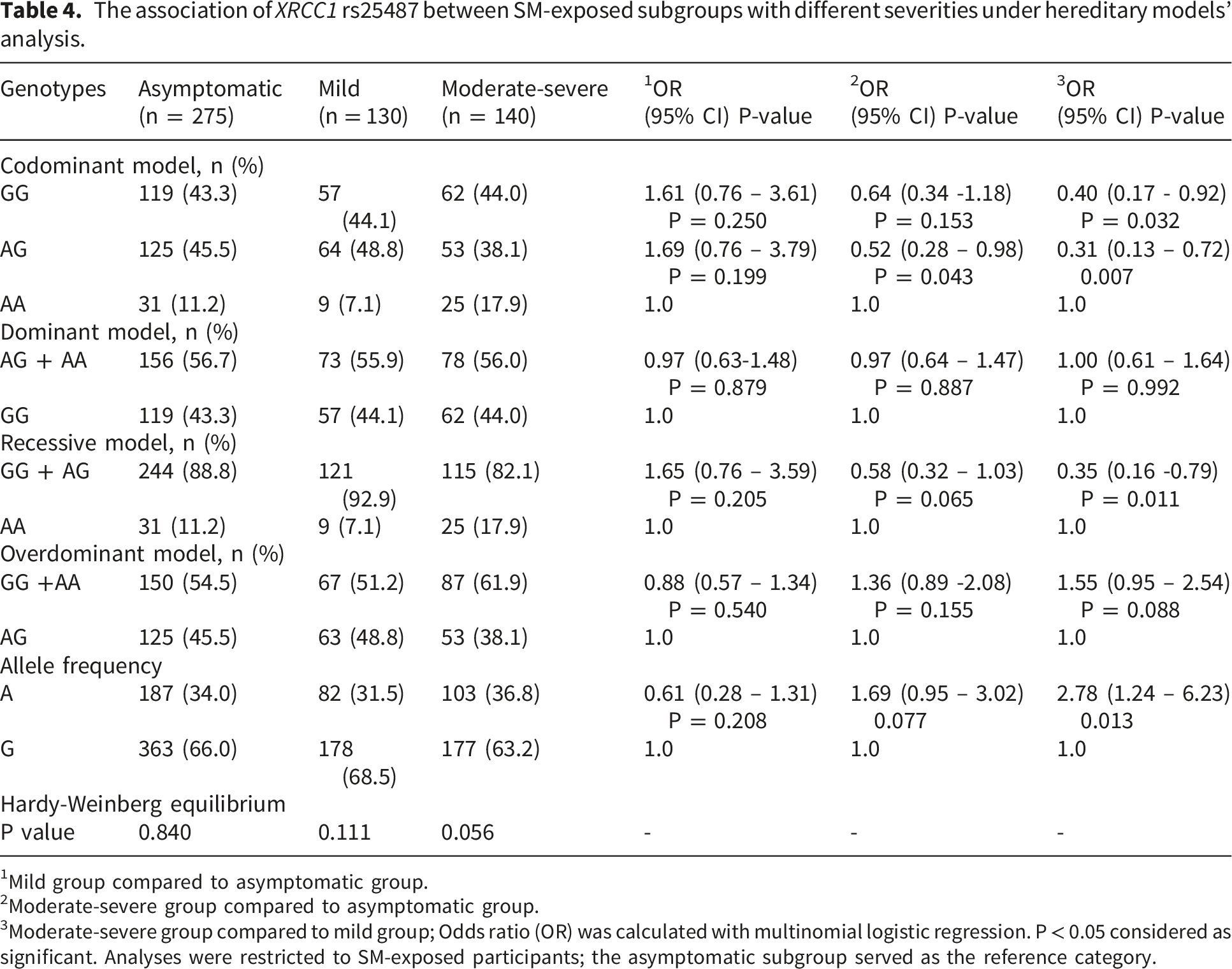
1Mild group compared to asymptomatic group.
2Moderate-severe group compared to asymptomatic group.
3Moderate-severe group compared to mild group; Odds ratio (OR) was calculated with multinomial logistic regression. P < 0.05 considered as significant. Analyses were restricted to SM-exposed participants; the asymptomatic subgroup served as the reference category.
The dominant model, including the AG + AA genotypes, did not reveal significant differences among the groups (P > 0.05), indicating that these genotypes do not appear to influence clinical sign severity meaningfully (Table 4).
In the recessive model, GG + AG genotypes were prevalent across all groups. A negative association was observed for the moderate-severe subgroup compared to the mild subgroup (P < 0.05), suggesting that individuals with this genotype may be less likely to be classified in the more severe subgroup (Table 4). The overdominant model showed that the GG + AA genotypes did not demonstrate significant associations when comparing the mild and asymptomatic subgroups (P > 0.05). However, a trend was noted for the moderate-severe subgroup compared to the mild subgroup, indicating a significant relationship (P < 0.05, Table 4).
Regarding allele frequency, the OR for the mild group compared to the asymptomatic group indicated no significant association (P > 0.05). For the moderate-severe group compared to the asymptomatic group, the OR suggested a trend toward increased risk but not reaching statistical significance (P > 0.05). The comparison of the moderate-severe group to the mild group yielded a significant OR (P < 0.05), indicating that individuals with the A allele in the moderate-severe group are significantly more likely to experience severe clinical signs compared to those in the mild group (Table 4).
Evaluation of XPC (rs2228001, C > A) polymorphisms
According to online SNPStats software analysis, the frequency of this XPC (rs2228001) SNP was consistent with Hardy-Weinberg equilibrium (P > 0.05).
The association of XPC rs2228001 between SM-exposed subgroups with different severities under hereditary models’ analysis.
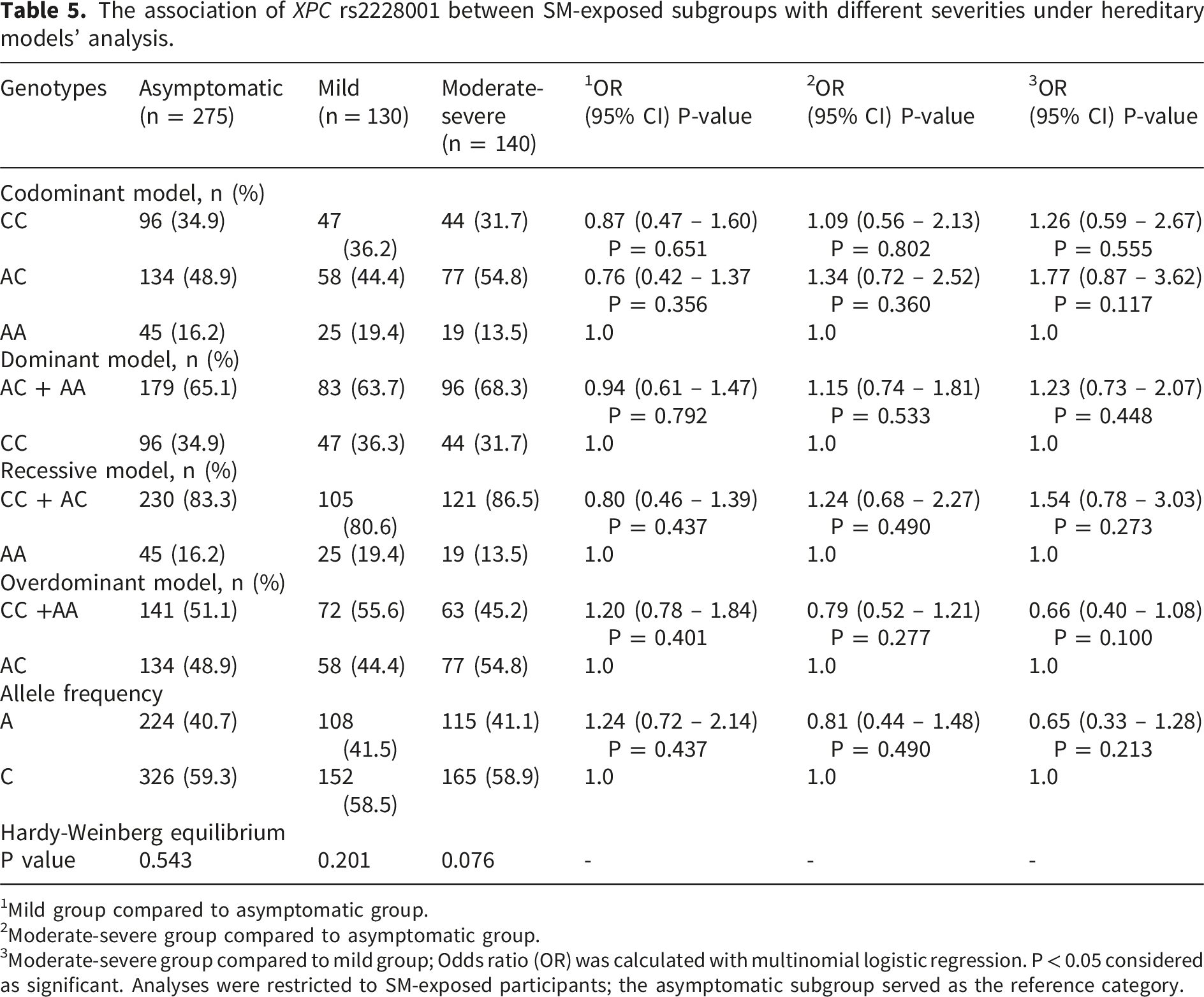
1Mild group compared to asymptomatic group.
2Moderate-severe group compared to asymptomatic group.
3Moderate-severe group compared to mild group; Odds ratio (OR) was calculated with multinomial logistic regression. P < 0.05 considered as significant. Analyses were restricted to SM-exposed participants; the asymptomatic subgroup served as the reference category.
Expression of XRCC1 and XPC genes
Figure 2 provided data on the mRNA levels of XPC and XRCC1 in a group exposed to SM compared to a non-exposed group, categorized by genotypes. The relative expression of XPC gene decreased in the SM-exposed group compared to the non-exposed group (P < 0.05) while the expression of XRCC1 gene increased (P < 0.05). In the classification of genotypes, the relative expression of XRCC1 in the SM-exposed group was significantly higher than in the non-exposed group for both the GG and AG genotypes (P < 0.05) while no significant change in XRCC1 transcript was detected in the AA genotype (P > 0.05). There were no changes in the relative expression of the XPC gene across its genotypes (P > 0.05). XPC and XRCC1 mRNA levels in SM-exposed group compared to the non-exposed group according to their genotypes. Data are presented as mean ± SD. Statistical significance was determined using independent t-tests; No correction for multiple comparisons was applied within genotype subgroups; these comparisons are exploratory. P < 0.05 is considered statistically significant. “Overall” means individuals with all genotypes.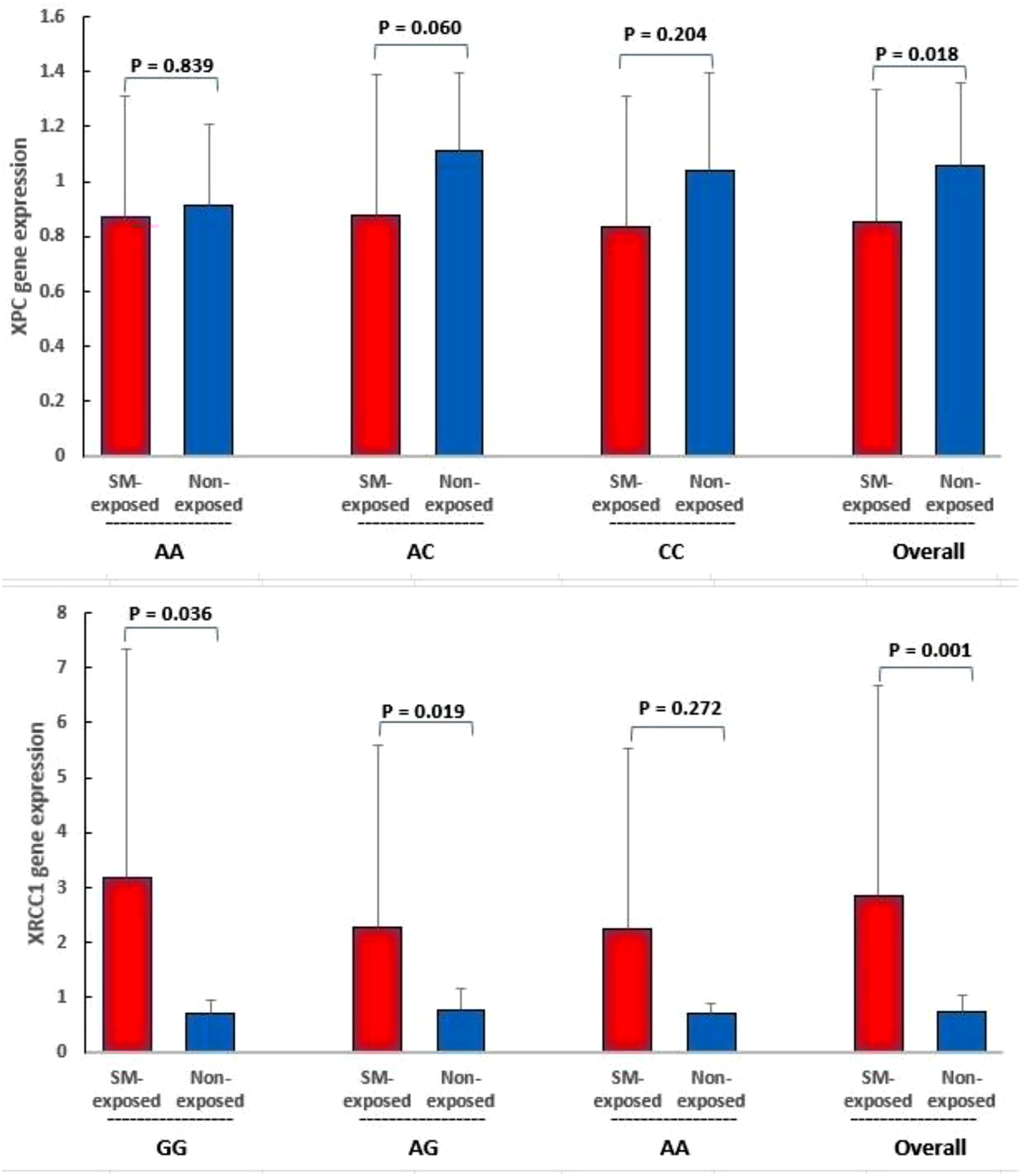
Figure 3 compares the XRCC1 and XPC mRNA levels between SM-exposed subgroups. The relative expression of XPC gene decreased in the moderate-severe subgroup compared to the non-exposed group and asymptomatic/mild subgroups (P < 0.05), while the changes between the non-exposed group and asymptomatic/mild subgroups were not significant (P > 0.05). The expression of XRCC1 gene increased in the moderate-severe and mild subgroups compared to the non-exposed group and asymptomatic subgroup (P < 0.05), while no significant change in XRCC1 transcript was detected between the non-exposed group and asymptomatic subgroup or between the moderate-severe and mild subgroups (P > 0.05). XPC and XRCC1 mRNA levels in SM-exposed subgroups compared to the non-exposed group. Data are presented as mean ± SD. Comparisons were made using one-way ANOVA followed by Tukey’s post-hoc test. (a) (b) Groups not sharing the same letter differ significantly (P < 0.05).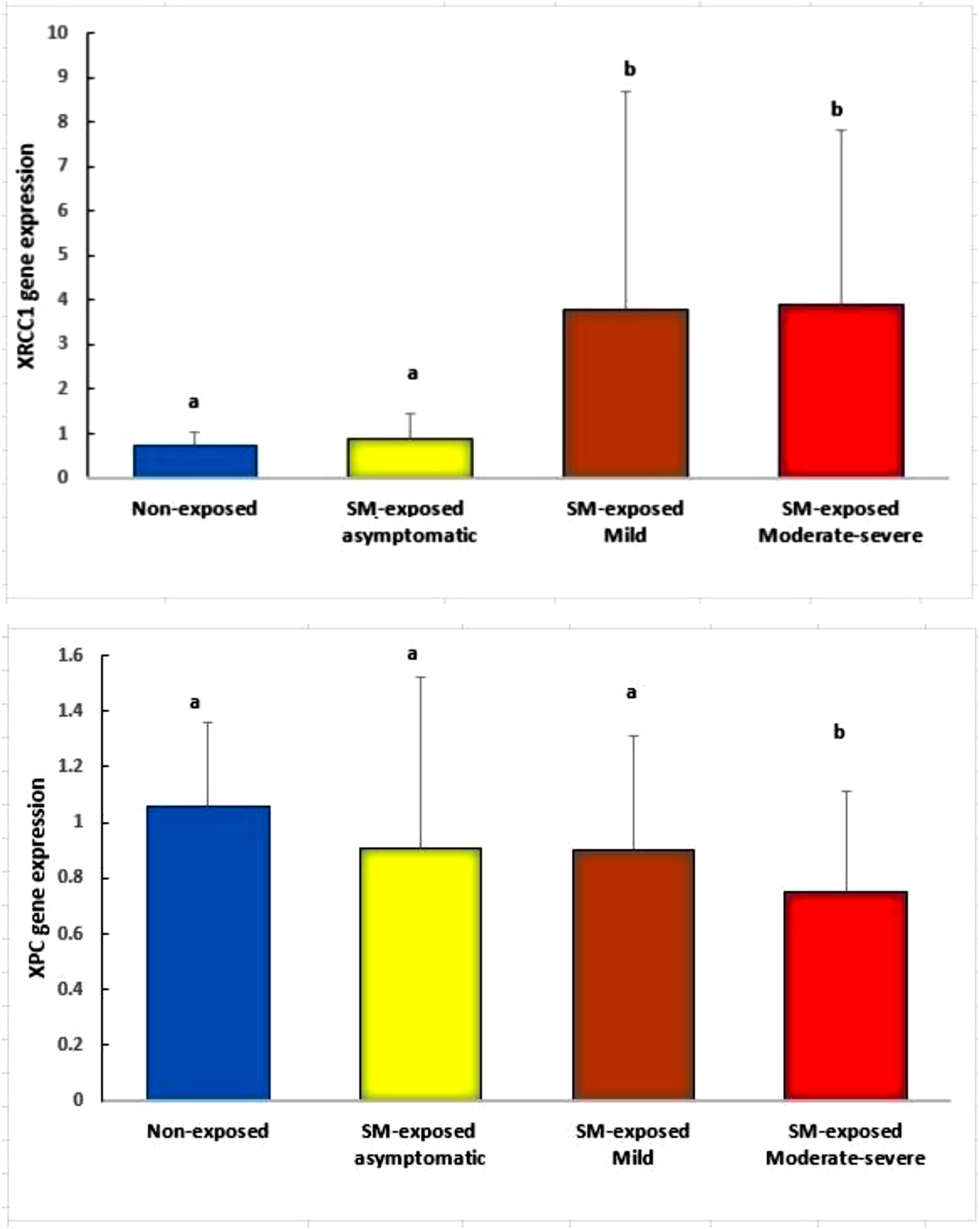
In the measurement of serum MDA levels, the MDA in the SM-exposed group (27.4 ± 20.51) was higher than in the non-exposed group (19.2 ± 12.53) (P < 0.05).
Figure 4 displayed the linear regression between MDA and XPC/XRCC1 mRNA levels in SM-exposed participants. A significant negative correlation was observed between MDA and the XPC transcript (R = - 0.18, P < 0.05), whereas the regression analysis between MDA and the XRCC1 transcript did not yield significant results (R = 0.039, P > 0.05). Pearson linear regression between malondialdehyde (MDA, μmol/L) and XPC/XRCC1 mRNA levels (relative expression) in SM-exposed participants. Each data point represents an individual measurement, and the shaded area around the regression line represents the 95% confidence interval. Pearson correlation coefficient (R) and P-value are shown in the figure; P < 0.05 was considered statistically significant.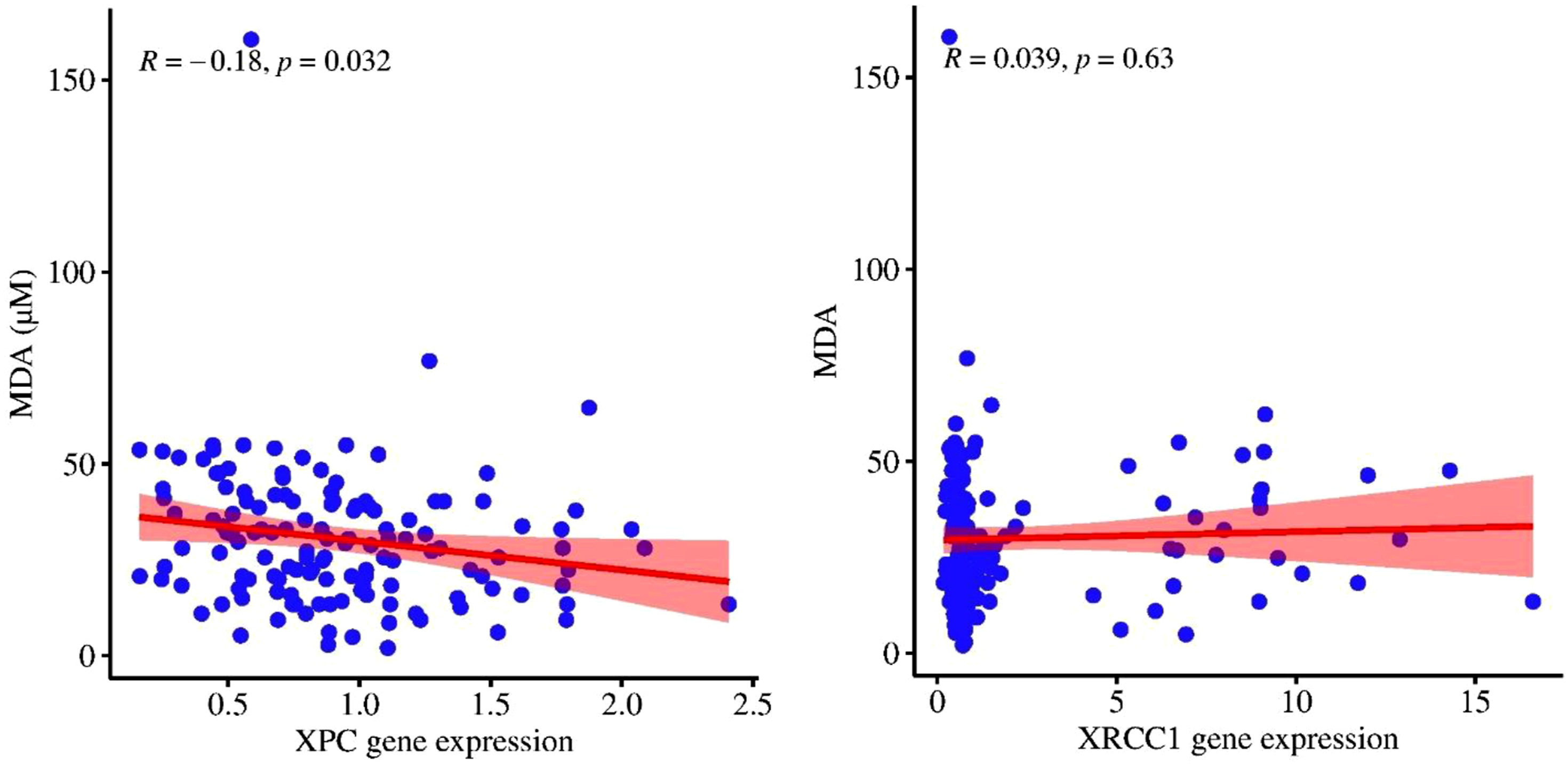
Discussion
This study aimed to evaluate the expression and polymorphism of the XRCC1 and XPC genes and their association with the severity of SM toxicity and lipid peroxidation. Our findings provide important insights into the genetic factors that may influence individual susceptibility to the harmful effects of SM exposure.
The significant reductions in pulmonary function parameters among SM-exposed participants highlight the chronic nature of the damage inflicted by this chemical agent. The progressive decline in FVC, FEV1, and FEV1/FVC ratio with increasing severity of clinical signs suggests that the extent of initial exposure and the resulting tissue damage correlate with more pronounced respiratory impairment. While the exact duration of exposure was not quantified in this study, the severity of clinical signs is likely influenced by the intensity of exposure and the individual’s physiological response to SM. This aligns with findings from other studies that have reported similar trends in lung function deterioration among chemical warfare victims, where the degree of respiratory impairment often reflects the severity of initial exposure and subsequent chronic damage.43–45
The analysis of polymorphisms in the XRCC1 gene provided valuable insights into the relationship between its genotypes and the severity of signs in individuals exposed to SM. By categorizing participants into asymptomatic, mild, and moderate-severe groups based on clinical severity, the analysis reveals distinct patterns of association for each genotype, which may have implications for understanding genetic susceptibility to SM toxicity. In the codominant model of XRCC1 genotypes, the GG genotype was prevalent across all severity groups, and individuals with this genotype did not have an increased risk of developing mild signs relative to those without clinical signs. Also, these individuals may be less likely to experience severe signs. This association suggests that the GG genotype may be linked to a more effective response to DNA damage caused by SM exposure, thereby correlating with lower severity of clinical signs. In this model, individuals with the AG genotype also showed an association with a reduced risk of severe clinical signs. The even stronger association observed when comparing the moderate-severe group to the mild group further emphasizes the potential role of the AG genotype in modulating clinical sign severity. This finding implies an association in which the AG genotype may be associated with differences in the clinical response to SM, possibly through improved DNA repair or other protective biological mechanisms. The AA genotype did not show a significant association with the severity of clinical signs in SM-exposed individuals. This lack of association may indicate that the AA genotype does not play a significant role in the biological pathways affected by SM exposure. In the recessive model, individuals with the GG + AG genotypes were associated with a lower likelihood of progressing from mild to moderate-severe signs. In the dominant and overdominant models of XRCC1, individuals with the AG + AA and GG + AA genotypes did not show a significant association with a higher or lower risk of developing mild and moderate-severe signs. However, the XRCC1 gene revealed significant associations with clinical sign severity, highlighting the potential role of genetic variability in modulating the effects of SM exposure. These findings are partially consistent with previous research indicating that individuals with certain XRCC1 polymorphisms show an association with higher DNA repair capacity, which has been associated with resilience against environmental toxins and a lower risk of developing chronic respiratory complications (e.g., chronic obstructive pulmonary disease, COPD).33,46
For the XPC gene, no significant associations were found across the severity subgroups, suggesting that the allele frequency and polymorphisms in XPC may not play a critical role in modulating the severity of clinical signs related to SM exposure. A previous study reported that the rs1805329 SNP located in the Rad23B gene, which is part of the NER pathway, is not associated with the severity of lung complications in patients exposed to SM. Additionally, no statistically significant differences were observed in genotypic frequencies across co-dominant, dominant, recessive, and over-dominant models. 44 These findings are consistent with our data regarding XPC genotypes. Conversely, a previous study determined that the XPC genotypes are associated with pulmonary function in COPD. 33 In that study, specific XPC variants were linked to poorer lung function outcomes, suggesting a potential role for XPC in respiratory disease susceptibility. This discrepancy may reflect differences in the underlying disease mechanisms or the influence of long-term adaptive responses in SM-exposed survivors. 47
The observed reduction in XPC mRNA levels among veterans exposed to SM (especially in the moderate-severe subgroup) is noteworthy. This decrease appears to be independent of specific genotypes, suggesting that the downregulation of XPC is a direct consequence of oxidative stress induced by SM exposure rather than genetic predisposition. This aligns with previous studies indicating that oxidative stress can lead to alterations in gene expression, potentially increasing cancer susceptibility due to impaired DNA repair capabilities.26,48 The negative correlation between XPC transcript levels and elevated MDA levels further supports the hypothesis that oxidative stress plays a pivotal role in downregulating XPC mRNA levels. As mentioned, MDA is a marker of lipid peroxidation and is often elevated in conditions characterized by oxidative stress, reinforcing the notion that SM exposure leads to significant cellular damage.5,49 It should be noted, however, that the TBARS assay used to measure MDA has recognized limitations in specificity, as it can also detect other aldehydes and is subject to interference. Therefore, MDA levels reported here should be interpreted as indicative of lipid peroxidation rather than as definitive or exclusive measures of oxidative damage. The findings suggest that the XPC gene, which is crucial for nucleotide excision repair, may become less effective in repairing oxidative DNA lesions due to its reduced expression in SM-exposed individuals.
In contrast, the significant elevation of XRCC1 expression among SM-exposed individuals, particularly in the mild and moderate-severe subgroups and those with GG and AG genotypes, is consistent with a possible compensatory response to heightened DNA damage. XRCC1 plays a critical role in base excision repair, which is essential for correcting oxidative DNA damage. The lack of correlation between XRCC1 expression and MDA levels suggests that its upregulation may not be directly tied to oxidative stress but rather may be consistent with an adaptive response to increased DNA damage. However, it has been established that XRCC1 polymorphisms serve as indicators of individual susceptibility to DNA damage induced by oxidative stress. 50 On the other hand, while oxidative stress can lead to DNA damage, the expression levels of XRCC1 remain stable, highlighting its essential role in sustaining DNA repair processes without significant variations. 51 This divergence in the behavior of XPC and XRCC1 highlights the complex interplay between different DNA repair pathways under oxidative stress conditions.
Overall, our findings underscore the importance of genetic factors in determining individual responses to SM exposure. The differential expression of DNA repair genes and the presence of specific polymorphisms may contribute to variations in susceptibility and severity of clinical signs among affected individuals.
The study population was limited to individuals exposed to sulfur mustard during the Iraq-Iran war, which may restrict generalizability to other populations or exposure scenarios. The cross-sectional design prevents establishing causal relationships; longitudinal studies are needed to explore temporal dynamics. Furthermore, genotype frequencies and transcript levels were measured decades after initial exposure. Therefore, the observed associations cannot establish mechanistic involvement in acute injury or disease progression; they may reflect survivor effects, long-term adaptive changes, or other confounding factors inherent in the cross-sectional design. Importantly, the absence of baseline (pre-exposure) biological samples limits our ability to assess individual-level changes in gene expression or oxidative stress markers directly attributable to SM. Additionally, we were unable to control for cumulative environmental exposures over the intervening decades, which may independently influence DNA repair capacity and oxidative stress. The logistic regression models were unadjusted for covariates. Although no significant differences in age or smoking status were observed across severity subgroups, non-significance does not preclude confounding. Moreover, unmeasured comorbidities (e.g., hypertension, diabetes, cardiovascular disease, or other chronic conditions) were not accounted for in the analyses and may independently influence both DNA repair capacity and the severity of clinical signs following sulfur mustard exposure. Therefore, the unadjusted odds ratios may be subject to residual confounding from age, smoking status, and comorbidities and should be interpreted cautiously. Adjusted analyses in future cohorts are required before drawing firm conclusions about genotype–severity relationships. While the sample size was substantial, it may have been insufficient to detect smaller genetic effects or rare variants. Moreover, the potential inflation of false-positive findings due to multiple testing across genetic models should be noted. Although the analyses were hypothesis-driven and focused on biologically relevant DNA repair genes, the results should be interpreted cautiously and require confirmation in future studies with independent populations and appropriate correction strategies.
Conclusion
The study highlights significant associations between XRCC1 and XPC gene polymorphisms, their expression levels, and the severity of sulfur mustard toxicity. The findings indicate that exposure to SM is associated with notable DNA damage, which appears to be linked to cellular DNA repair mechanisms involving XRCC1 and XPC. Specifically, polymorphisms in these genes are associated with individual susceptibility to SM-induced toxicity, as observed in the varying expression levels among participants with different genotypes. The results suggest that individuals with specific XRCC1 genotypes, particularly GG and AG, may show differing associations with SM exposure outcomes, potentially correlating with the risk of developing severe clinical signs. Furthermore, the interplay between oxidative stress and DNA repair gene expression underscores the complexity of cellular responses to sulfur mustard exposure. It is important to note that this cross-sectional study cannot establish causality; the observed associations may reflect long-term adaptive changes or survivor effects rather than direct mechanistic roles in acute injury or disease progression.
Supplemental material
Supplemental material - DNA repair gene polymorphisms and expression in sulfur mustard-exposed veterans: associations with toxicity severity
Supplemental material for DNA repair gene polymorphisms and expression in sulfur mustard-exposed veterans: associations with toxicity severity by Marzieh Mojtahed, Tooba Ghazanfari, Soyar Sari and Mehrdad Hashemi in Human & Experimental Toxicology.
Footnotes
Acknowledgements
We thank the Immunoregulation Research Center of Shahed University, which has carried out this study.
Ethical considerations
The study was conducted in accordance with the ethical principles of the Declaration of Helsinki 52 and was approved by the Shahed University Research Ethics Committee (approval codes: IR. SHAHED.REC.1399.151 and IR. SHAHED.REC.1403.013). Written informed consent was obtained from all participants prior to their inclusion in the study.
Author contributions
Conceptualization, M.M., T.G., S.S., and M.H.; Methodology, M.M., T.G., S.S., and M.H.; Formal Analysis, M.M., and T.G.; Investigation, M.M., T.G., S.S., and M.H.; Writing– Original Draft Preparation, M.M., and T.G.; Writing– Review and Editing, M.M., and T.G.; Supervision, T.G.; Project Administration, T.G., S.S., and M.H.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Data are available from the corresponding author upon request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.