
Abstract
The clearance of apoptotic cells has an important role in the maintenance of tissue homeostasis and in the protection of tissues from the inflammatory and immunogenic contents of dying cells. A defect in the recognition and phagocytosis of apoptotic cells contributes to the development of chronic inflammation and autoimmune disorders. We have observed that compared with healthy donors, differentiated macrophages from patients with untreated systemic lupus erythematosus (SLE) showed decreased phagocytosis of apoptotic neutrophils. A TaqMan Low Density Array was designed to determine the mRNA expression levels of 95 apopto-phagocytic genes in differentiated non-phagocytosing and phagocytosing macrophages. In the macrophages of clinically and immunoserologically active SLE patients, 39 genes were expressed at lower levels than in the control macrophages. When inactive patients were compared with those with minor immunoserological abnormalities or patients in an immunoserologically active state, a relationship was observed between the altered gene expression profile and the disease state. In the macrophages of patients with engulfing apoptotic cells, an upregulation of genes involved in inflammation, autophagy, and signaling was observed. These results indicate that novel immune-pathological pathways are involved in SLE and suggest targets for potential therapeutic modulation.
Introduction
Apoptosis, or programmed cell death, occurs daily at a high rate in the body. Apoptotic cells are rapidly removed by professional and non-professional phagocytes, which maintain tissue homeostasis even during inflammation. For efficient clearance, cells undergoing apoptosis secrete “find-me” and “eat-me” signals into their environments. These signals act as a trigger for rapid phagocytic engulfment, which employs a complex molecular mechanism with redundant receptor and signaling elements. 1 A deficiency in the phagocytosis of apoptotic cells will result in the accumulation of secondary necrotic cells with concomitant leakage of toxic contents, resulting in tissue injury, chronic inflammation and autoimmune disease.
Systemic lupus erythematosus (SLE) is a multifactorial and multi-organ autoimmune disease which, based on the clinical manifestations, can be divided into phenotypes involving patients with lupus nephritis, those with mucocutaneous or musculoskeletal disease, a neuropsychiatric group, and those with diseases involving more systems.2,3 The disease is characterized by a chronic and antigen-driven autoimmune response against nuclear, cytosolic and membranous materials, including microparticles and neutrophil extracellular traps.4–7 Studies have led to the identification of a series of genes that can determine the susceptibility or severity of the disease. 8 Reduced DNASE I activity in serum, 9 complement deficiency10,11 and C reactive protein deficiency 12 were described as molecular elements that play roles in the pathogenesis of SLE. Recent studies reported that a decrease in protein S levels might contribute to SLE pathogenesis. GAS6, a TAM (tyro3, axl, mer) kinase ligand, was increased in a small number of patients with a history of neurological disorder and acute activity in the BILAG hematology system. 13 The acceleration of apoptosis and the impaired clearance of the apoptotic cells were described in a study involving skin biopsies from patients with cutaneous lupus erythematosus. 14 C1q, which is linked to the recognition and removal of apoptotic cells, is decreased in patients with SLE and contributes to the reduced phagocytosis of apoptotic material. 15 The upregulation of two interferon-regulated genes, IL6R and PKRCG (protein kinase C gamma), was also described in peripheral blood mononuclear cells and bone marrow mononuclear cells. In patients with active disease, β2 integrin and genes involved in the integrin signaling pathway such as MAP3K11 (mitogen-activated protein kinase), ACTB (actin beta gene) ACTG1 (actin, gamma 1), ARPC3 (actin-related protein 2/3 complex), MRLC2 (swine myosin regulatory light chain 2) and MRLC3 (myosin regulatory light chain) were upregulated. 16 The toll-like receptors (TLRs) also appear to be involved in the pathophysiology of lupus. 17
In this study, a high-throughput, quantitative RT-PCR assay (TaqMan Low Density Array (TLDA)) containing 95 genes involved in the clearance of apoptotic cells was used to determine whether the expression of any of these genes might impair phagocytosis of dying cells in patients with lupus erythematosus. The main criterion for patient selection was that patients had not received immunosuppressive or immune-modulator therapy. It is reported that expression of phagocytosis receptors, bridging molecules, signal generators, effectors, cytokines, nuclear receptors, engulfment genes, autophagy genes, and interferon regulatory family genes were downregulated or differentially regulated in the monocyte-differentiated macrophages of lupus patients.
Material and methods
Patients and controls
Peripheral blood samples were obtained from patients with SLE. The diagnosis of SLE was established according to ACR criteria. 18 The 10 patients with SLE were classified as having active or inactive disease. The average age of the patients was 42 ± 18.76 (one male and nine female). Using clinical and serological characteristics, two further subgroups were formed. The patient subgroups were as follows: clinically and immunoserologically active (three patients); immunoserologically active without clinical symptoms (three patients); expressing minor immunoserological abnormalities (two patients); and not expressing serological changes or clinical signs at the time of enrollment, termed inactive patients (two patients). The control group contained three healthy individuals. None of the patients used in this study were being treated with glucocorticoids or other immunosuppressant therapies. The immunoserological characteristics of the patients are presented in Table S1 in the Appendix.
Cell culture procedures
Peripheral human blood mononuclear cells were isolated using density gradient centrifugation with Ficoll–Paque Plus from the fresh blood of donors. CD14+ cells were separated using magnetic sorting with MACS, followed by washing with PBS containing 0.5% BSA and 2 mM EDTA. Neutrophils were isolated from freshly drawn blood with density gradient centrifugation using Histopaque1119 and Histopaque1077. Freshly isolated monocytes were cultured for a period of 5 days in IMDM supplemented with 10% AB serum and 5 ng/ml M-CSF. Neutrophils were incubated for 24 h in IMDM with 1% AB serum to allow 50–80% to undergo apoptosis. 19 Neutrophil apoptotic levels were tested with an Annexin-PI detection kit (MBL International Corporation, Woburn, MA).
Macrophage phagocytosis assays
Before exposure to apoptotic stimuli, neutrophils were labeled with 5(6)-CFDA-SE (15 µM, overnight), the conditioned media was washed away, and the cells were re-suspended in PBS prior to adding the cells to a prewashed macrophage monolayer labeled with Cell Tracker™ Orange CMTMR (3.75 µM, overnight). Macrophages and apoptotic neutrophils were mixed at a ratio of 1:5 and incubated for 1 h at 37℃ in 5% CO2. Following incubation, non-ingested neutrophils were removed using trypsin for flow cytometry analysis. For TLDA, the apoptotic neutrophils were removed from the macrophage monolayer by washing with PBS.
RNA isolation and quantification
Total RNA was isolated from cells using Trizol. For TLDA (Applied Biosystems, Assay ID: 4342379-18 S), a pre-designed TaqMan probe and primer sets for target genes were chosen from an online catalog. The sets were factory-loaded into the 384 wells of the TLDA. The array format was customized with two replicates per target gene, and two parallel analyses were carried out for each biological sample. TLDA was used in a two-step RT-PCR process. First-strand DNA was synthesized from 1 µg of total RNA using the High-Capacity cDNA Archive Kit (Applied Biosystems). Q-PCR reactions and analyses were then performed with TLDAs using TaqMan Universal Master Mix and the ABI PRISM 7900 HT Sequence Detection System (Applied Biosystems). A sample volume of 100 μl per port (4 ports/sample) was used at 1 ng/μl concentrations. The expression level of target genes was normalized to 18 S rRNA as an endogenous control. Gene expression values were calculated based on the ΔΔCt method, in which one sample was designated as the calibrator, against which all other samples were analyzed. ΔCt represents the threshold cycle (Ct) of the target minus that of 18 S rRNA and ΔΔCt represents the ΔCt of each target minus that of the calibrator. Relative quantities (RQ or fold changes) were determined using an equation in which relative quantity equals 2−ΔΔCt.
The fold change represents the ratio of relative gene expression comparing differentiated SLE and control macrophages or the ratio of relative gene expression between phagocytosing SLE macrophages and non-phagocytosing SLE macrophages. The data presented are given as the mean of the fold changes ± standard error mean (SEM) of SLE donors used.
Statistical analysis
Data were analyzed statistically using unpaired and paired Student’s t-tests.
Results
Impaired phagocytosis of apoptotic neutrophils by macrophages from patients with systemic lupus erythematosus
Monocytes from patients with SLE who did not receive glucocorticoid or other immunosuppressant therapy were differentiated to macrophages. The phagocytosis of apoptotic neutrophils by SLE macrophages was investigated with flow cytometry using a two-dye system. Our results clearly show that monocyte-derived macrophages from SLE patients phagocytosed fewer apoptotic neutrophils compared with macrophages from healthy individuals (Figure 1).
(a) The percentage of apoptotic neutrophils cultered for 24 hours in IMDM and 10% AB serum and stained with Annexin-PI. The presented flow cytometric data are from one representative experiment. (b) The phagocytic activity of control (black) and SLE patient-derived (grey) macrophages measured by flow cytometry utilizing apoptotic neutrophils as the target. (c) The phagocytic capacity of one representative control and SLE patient-derived macrophages, as determined by flow cytometry. M1 represents gating for CFDA-SE-labeled neutrophil granulocytes engulfing cells.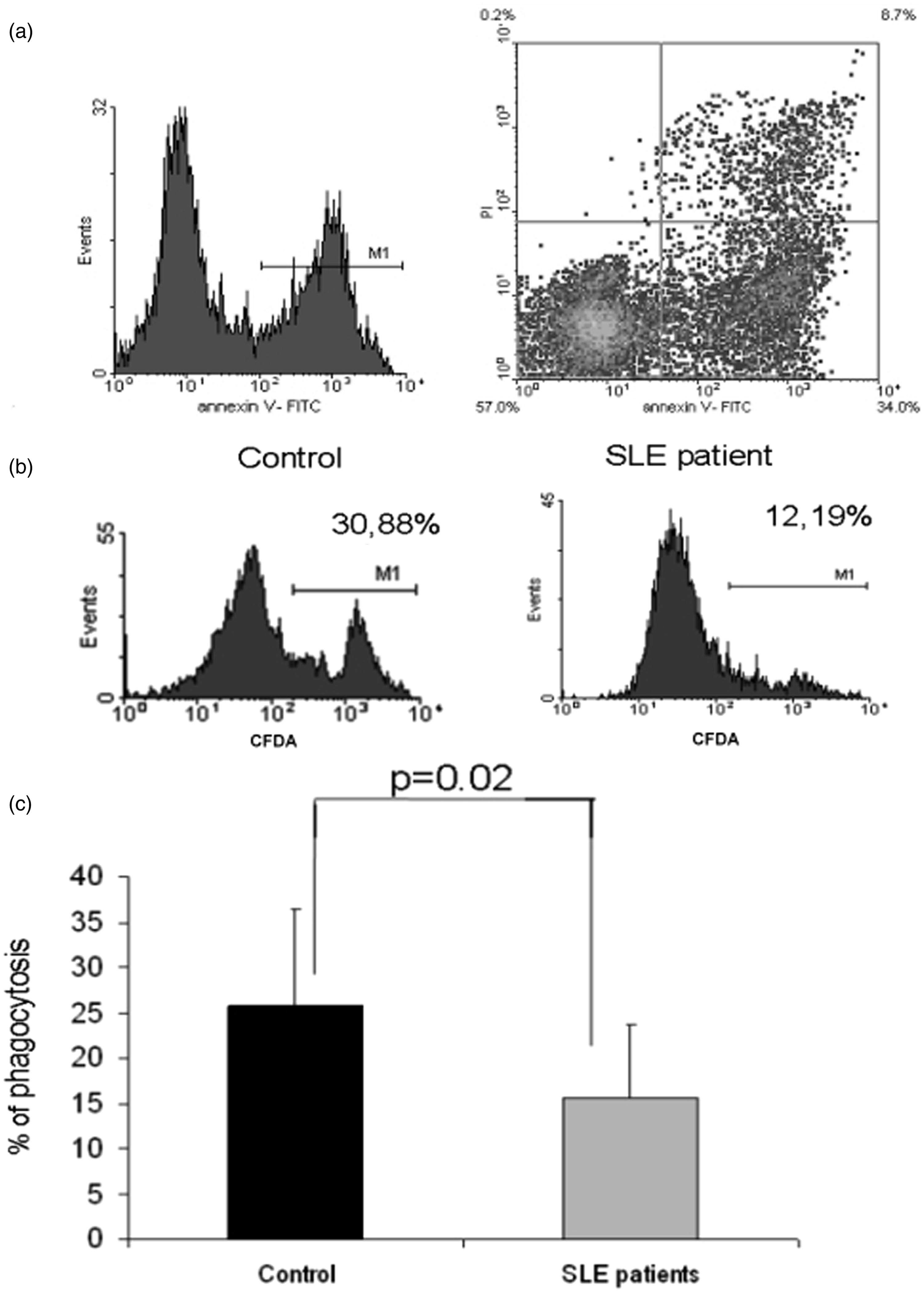
Expression pattern profile of apopto-phagocytic genes in patient-derived macrophages
In macrophages from 10 patients with SLE showing deficient phagocytosis of apoptotic cells, a TLDA containing 95 genes (Table S2, Appendix) was used to determine the expression pattern of apopto-phagocytic genes. As described in the materials and methods, patients were classified with either clinically active or inactive disease. Using the clinical and serological characteristics of the patients, an additional two subgroups were included: patients with minor immunoserological abnormalities and patients with immunoserological activity in the absence of clinical signs. In the gene array analysis for every donor, we selected only genes that exhibited at least a twofold decrease or increase. The data were analyzed in relation to the activity of the disease and the immunoserological status.
In all of the macrophages from clinically and immunoserologically active SLE patients, 39 genes were downregulated (Figure 2, Table 1) and six were upregulated (Table 2) compared with control macrophages. In patients with immunoserological activity, 31 genes were downregulated; in patients with minor immunoserological abnormalities, four genes were downregulated; and in patients with inactive disease, five genes were downregulated, suggesting a relationship between the gene expression profile and the activity of the disease (Figure 2, Table 1). The data presented are presented as the mean fold change ± SEM of the donors. The downregulated genes included receptors, autophagy genes, bridging molecules, signal generators and effector molecules. Comparing the four subgroups of macrophages from SLE patients, two downregulated genes were observed in all groups (Figure 2, Table 1), namely the bridging molecule MFGE-820 and ITGB5.
Downregulated genes in monocyte-differentiated macrophages from SLE patients. Downregulated genes in monocyte-differentiated macrophages in SLE patients with clinically and immunoserologically active disease downregulated genes in immunoserological active patients downregulated genes in patients with minor immunoserological changes downregulated genes in inactive patients p ≤ 0.05 Upregulated genes in monocyte-differentiated macrophages in SLE patients with clinically and immunoserologically active disease p ≤ 0.05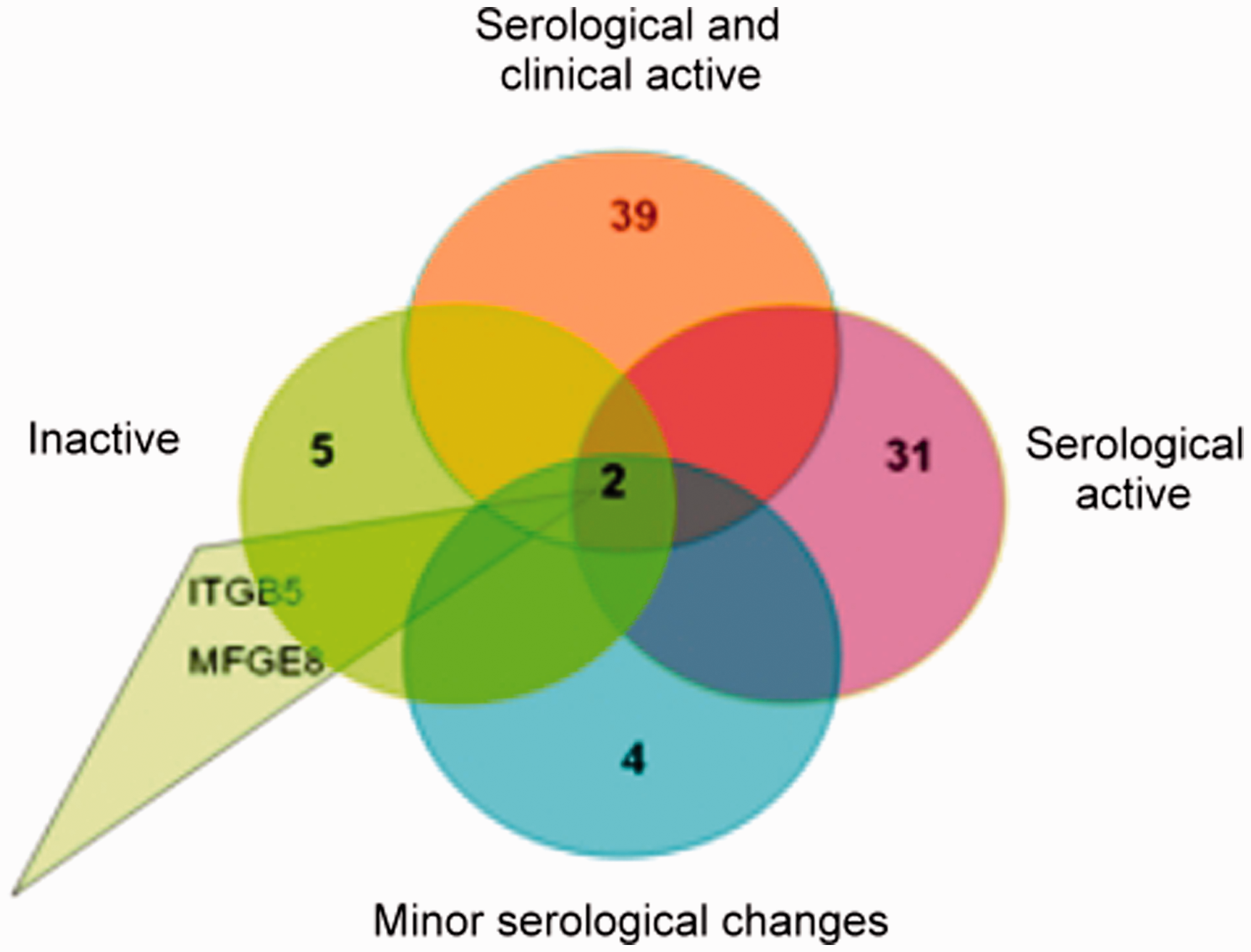
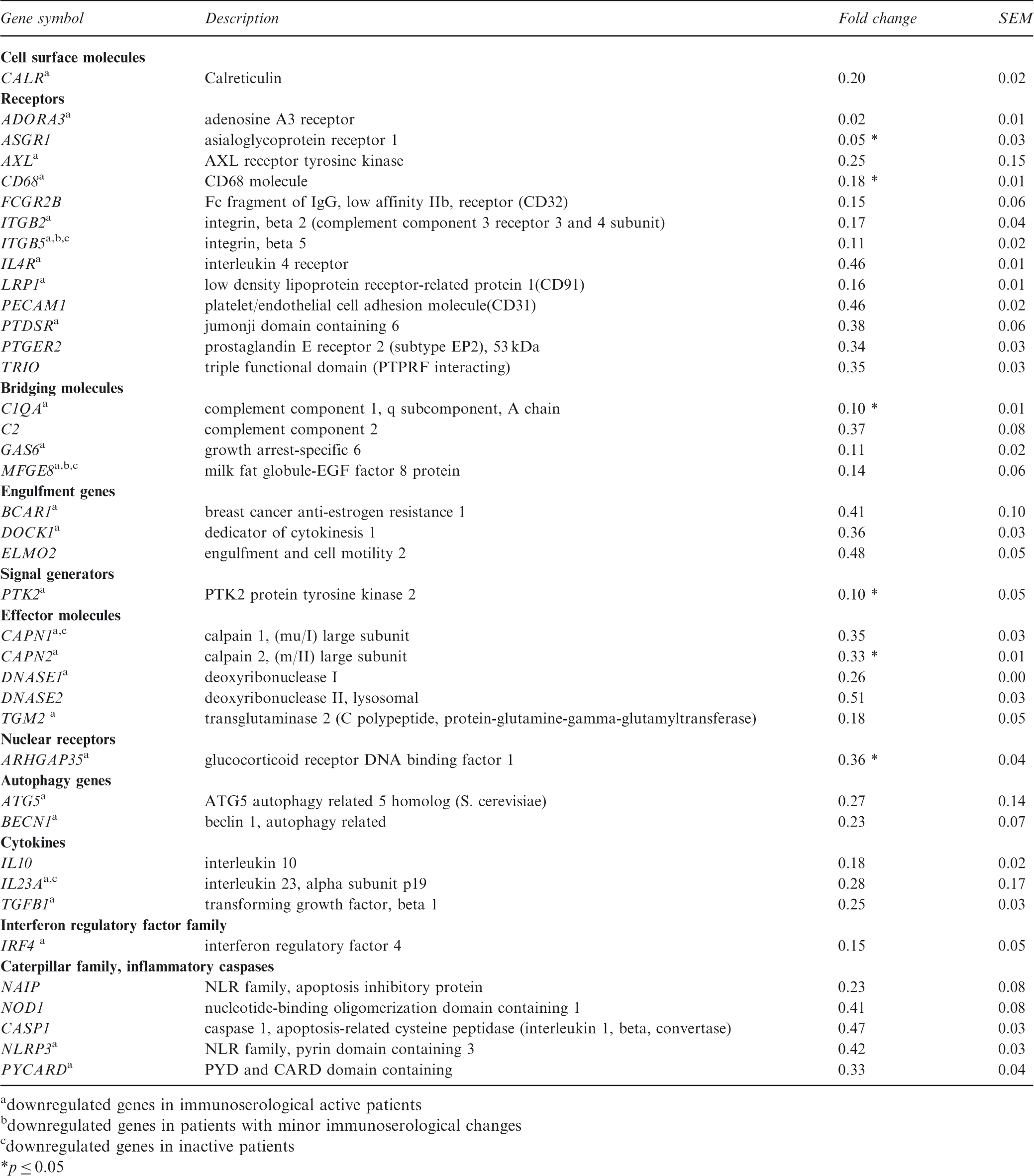

Here, we show that the expression of the receptors CD68 and ASGR1, the effector molecule CAPN2, and PTK2, which is involved in integrin-mediated phagocytosis, are significantly downregulated in SLE patients with clinically and immunoserologically active disease (Table 1).
The expression of the receptors ADORA3, PECAM1; the bridging molecules GAS6 and C2; the engulfment genes DOCK1, ELMO2 and the enzyme JMJD6 are also impaired in SLE patients with clinically and immunoserologically active disease (Table 1). As expected, the effector molecules DNASE I, DNASE II, and TG2 (Table 1), which are involved in the pathogenesis of glomerulonephritis with rheumatoid factor type auto antibodies,21,22 were also expressed at lower levels. In SLE patients with clinically and immunoserologically active disease, the receptor FPR2, the bridging molecule PROS1, C3, and the inflammatory cytokines IL6 and IL12B were highly expressed (Table 2). There was also an increase in the expression of OLR1, which is regulated by stimuli related to atherosclerosis. Figure 3 represents the comparison of the normalized expression of the 95 apopto-phagocytic genes in one control and one representative SLE patient with clinically and immunoserologically active disease.
Comparison of normalized expression of the 95 apopto-phagocytic genes in one control and one representative SLE patient with clinically and immunoserologically active disease.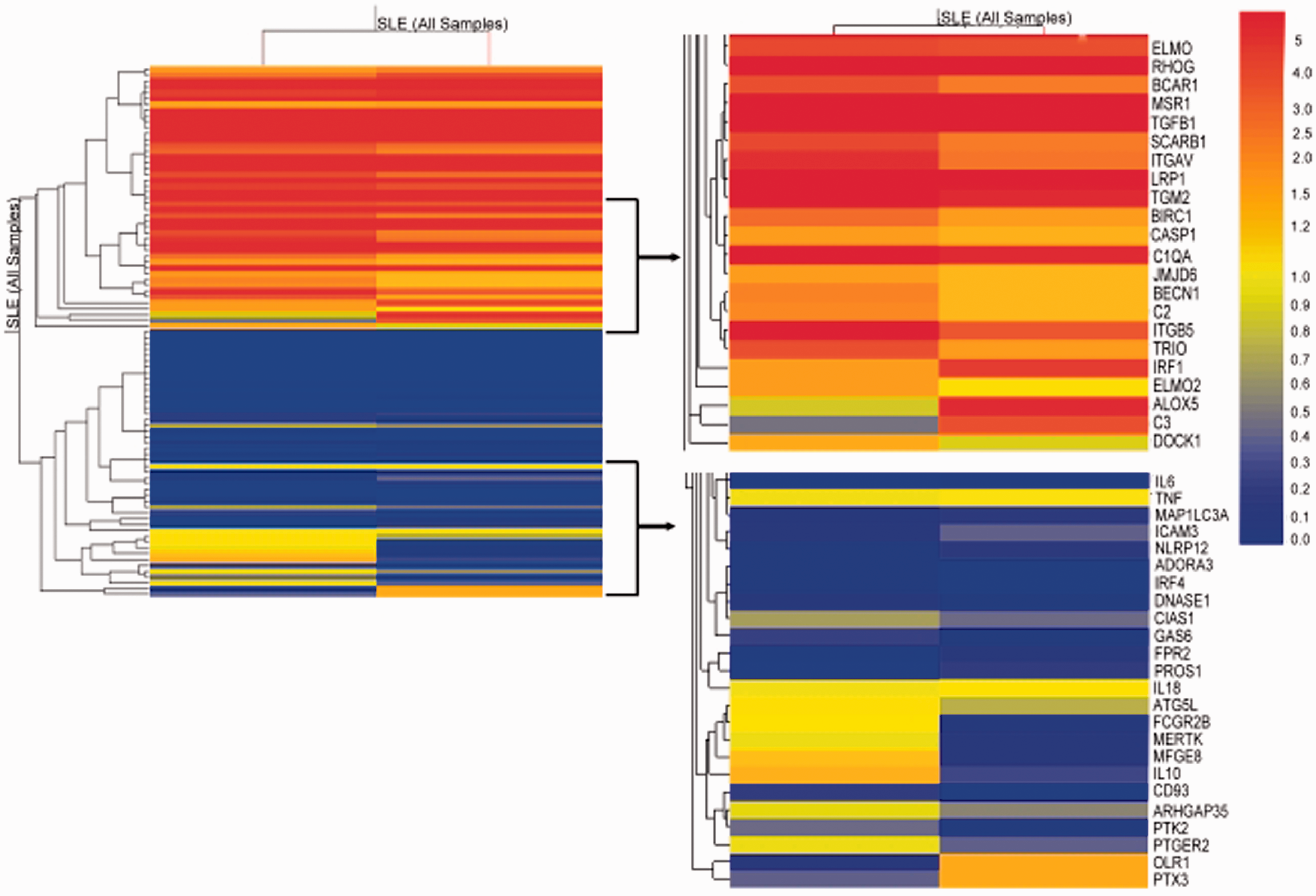
Changes in macrophage expression pattern during the phagocytosis of apoptotic neutrophils
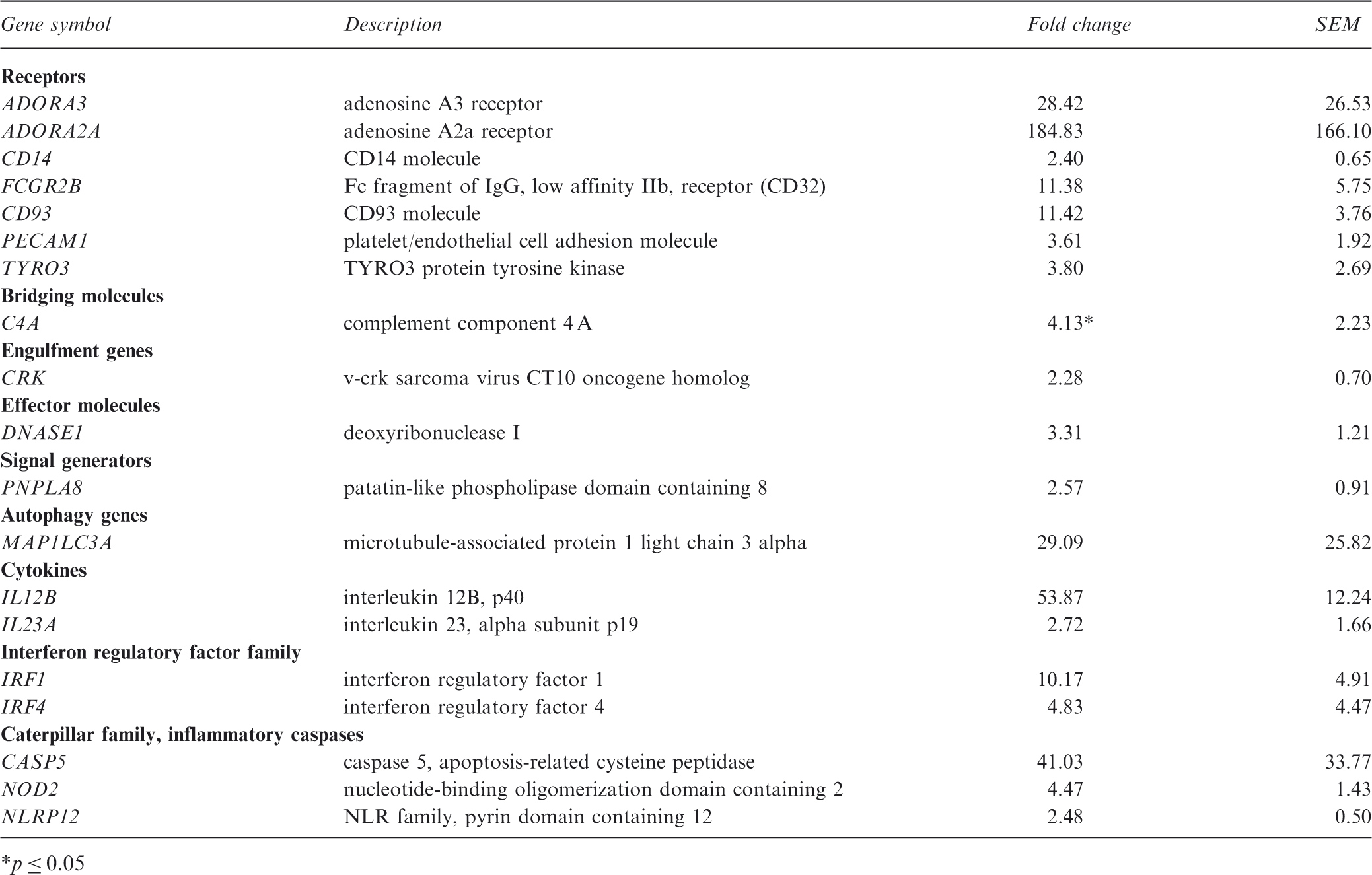
To learn more about the macrophages differentiated from the monocytes of lupus patients, the expression of apopto-phagocytic genes in response to engulfed apoptotic neutrophils was also investigated. Using the same analysis methods described above, we found that in control macrophages eight genes (ADORA1, ADORA2A, AXL, FPR2, ICAM3, THBS1, TNF-α, and PTGE2R) were upregulated. Comparing the gene expression in control cells and SLE macrophages, we found that of the genes listed above, only five receptors (ADORA2A, FPR2, ICAM3, THBS1, and TNF-α) were upregulated during the engulfment process (Figure 4). During phagocytosis, AXL was not increased in six patients, ADORA1 was not increased in 8 patients, and PTGE2R was not increased in four patients (Figure 4). The AXL receptor mRNA was expressed at a twofold lower level compared with control phagocytosing cells in one inactive patient. AXL downregulation in the other SLE patient subgroups was not evaluated because the expression level was not decreased by twofold compared with controls. In addition, during phagocytosis in patients with clinically and immunoserologically active disease, 18 genes were upregulated; in patients with immunoserological activity, seven genes were upregulated; in patients with minor immunoserological abnormalities, four genes were upregulated; and in the inactive group, seven genes were upregulated (Figure 4). These genes are mostly involved in inflammation, autophagy, and signaling (Table 3). Comparing the subgroups of patients, two genes were highly expressed in all cases: the inflammatory cytokine IL12B and the autophagy gene MAP1LC3. C4a, which is a short-lived fragment of C4 generated during activation of the classical or lectin pathway, was significantly elevated during phagocytosis of apoptotic neutrophils in patients with active disease. In the same patients, NOD2, a member of the NLR family and a regulator of autophagy, was also increased during phagocytosis. In all of the SLE patients with clinical and immunoserological changes, ADORA3 and ADORA2A were strongly upregulated compared with the control cells during phagocytosis (Table 3). The expression of IL-23, which was shown to contribute to the development of nephritis in lupus-prone mice,
23
was increased in patients with clinically active disease and in one inactive patient with glomerulonephritis in their history. PECAM1, CD93 and the interferon regulatory factors 1 and 4 were increased during phagocytosis in most of the patients.
Upregulated genes in monocyte-differentiated macrophages from SLE patients. Upregulated genes during phagocytosis in SLE patients with clinically and immunoserologically active disease p ≤ 0.05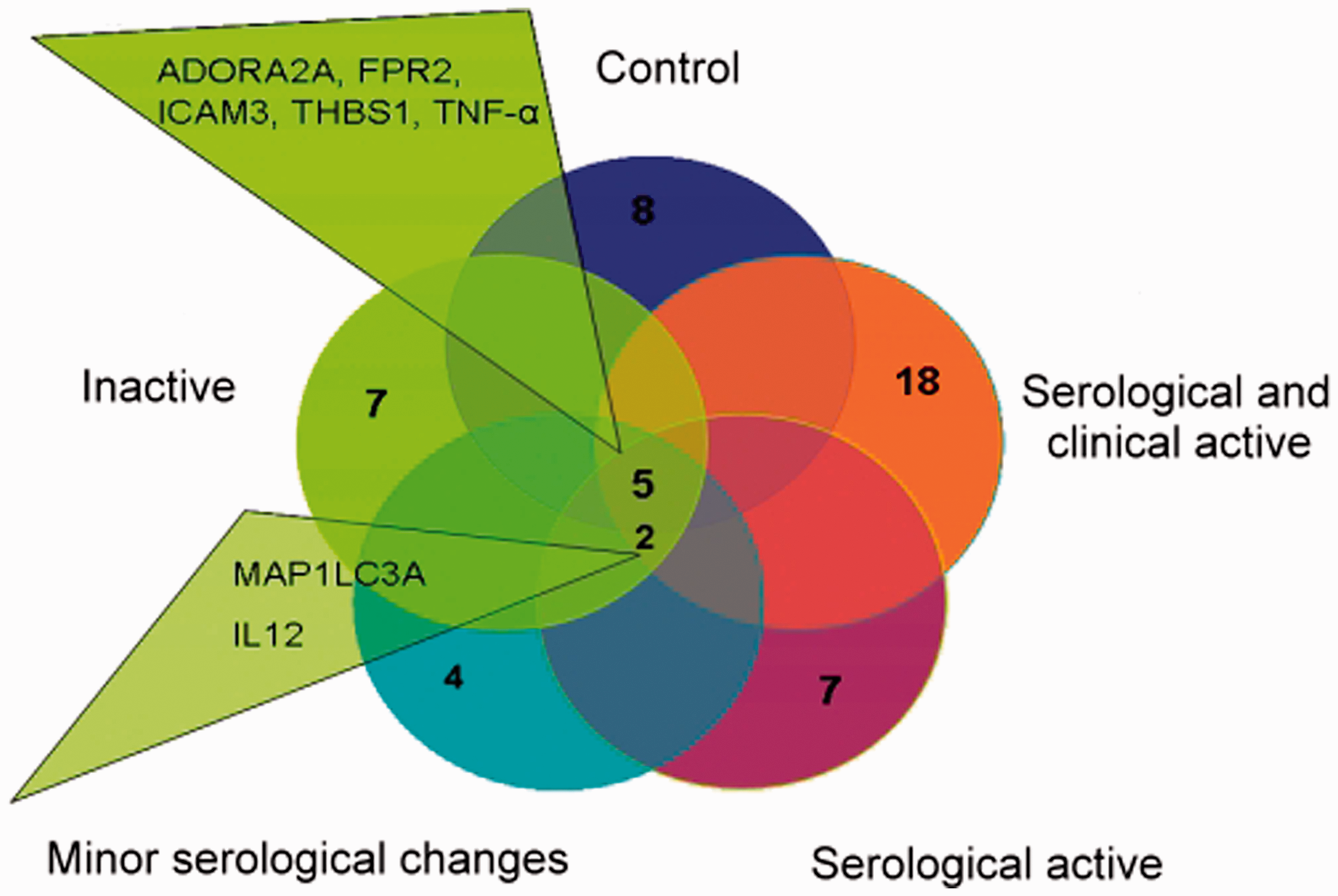
Discussion
Lupus erythematosus is a multifactorial disease with an etiology affected by genetic predisposition, environmental factors, infections and UV exposure. The defective clearance of apoptotic cells was shown to be involved in the pathogenesis of SLE. 24 The generation of nucleosomes in cells undergoing apoptosis is a potential source of autoantigens. These autoantigens are clustered and concentrated on the surface of apoptotic blebs 25 and when present in a complex with HMGB1 they could lead to an autoimmune response. 26
In the present study using a TLDA, our aim was to identify misregulated genes involved in the defective clearance of apoptotic neutrophils in patients with SLE. The main strength of this study was the ability to select patients that had not received immunosuppressant or immunomodulatory therapy, which would influence the gene expression pattern observed. Despite this strength, a very high variability of expression levels were observed among healthy and SLE macrophage donors. Although there were genes with low variability, as measured by the SEM, the expression level of other genes varied 2–10 orders of magnitude, impacting the results of the statistical analysis.
Our data show that the apopto-phagocytic system is downregulated in lupus, which is detectable even after the differentiation of freshly isolated monocytes to macrophages, and results in the decreased phagocytosis of apoptotic cells shown both here and by previous investigators.14,16 The downregulated genes were most numerous in SLE patients with clinically and immunoserologically active disease.
Among the 39 downregulated genes in clinically and immunoserologically active SLE patients macrophages, only a few such as C1Q, calreticulin, MFGE8, and FC2RG10,20,27–29 have been previously reported to be misregulated. Interestingly, in patients with SLE, the phagocytic machinery is already affected at the level of apoptotic cell recognition. Molecules involved in the recognition such as MFGE8, ITGB5, and GAS6 were downregulated. ASGR1, which recognizes the exposed β-galactose residues on apoptotic cells, was expressed at a significantly lower level in SLE macrophages, perturbing the prompt recognition of apoptotic cells. Signaling pathway genes involved in engulfment also exhibited reduced mRNA expression levels; genes encoding actin polymerization proteins (DOCK1, ELMO2, and PTK2), which also affect the engulfment and migration process, had less support from expression analysis. Furthermore, decreased levels of DNASE I and DNASE II in macrophages might cause the accumulation of fragmented DNA with concomitant activation of these cells and the production of inflammatory cytokines. DNASE II deficiency has been linked to the production of inflammatory cytokines and the development of chronic polyarthritis. 22 The role of the downregulated effector molecule TG2 in the pathogenesis of glomerulonephritis with rheumatoid factor type autoantibodies was published earlier by Szondy et al. 21 In addition to the above-mentioned downregulated genes, molecules involved in inflammation were upregulated. The bridging molecule PROS1, which is considered a clinical biomarker for disease activity, 30 was highly expressed in patients with clinically and immunoserologically active disease. IL12B (Table 3), which is involved in IFNγ production by NK cells, is also upregulated in the macrophages of patients with clinically active disease.
The patient-derived macrophages were also investigated during the engulfment of apoptotic cells, and it was revealed that the response to dying cells was different from the control macrophages. During phagocytosis, of the eight genes elevated in control cells, three receptors (AXL, PTGE2R, and ADORA1) were not upregulated in SLE macrophages. AXL, a member of the tyrosine kinase receptor family, is known as a receptor for GAS6 and PROS. The TAM/GAS6 system was proposed to play a role in the phagocytic process in the testis and retina.31,32 TAM receptors negatively regulate immune reactions, and a lack of TAM receptors was demonstrated to produce high levels of IL6 and TNF in dendritic cells, which might be responsible for the SLE-like autoimmunity observed in mice.33,34 PGE2 analogs were shown to inhibit IFN-α production by TLR-activated plasmocytoid dendritic cells, an effect which is mediated by the E-prostanoid 2 and E-prostanoid 4 receptors. Interestingly, PGE2 has a negative effect on these receptors. The ligation of FCγRII using immune complexes in SLE patients induced a massive production of PGE2, 35 which has a protective role via direct inhibition of the Th1 response by inducing Foxp3+ T reg differentiation and suppressing B-cell activation and Ig production.36–40 The complement system plays an important role in the pathophysiology of SLE. The activation of the complement cascade is initiated by the high titer of autoantibodies and immune complexes, and this activation results in the generation of several effector molecules, such as anaphylatoxins. The level of the anaphylatoxin C4a was found to correlate with the activity of the disease and the anti-DNA antibody level. 41 In addition, we show that C4a is also significantly increased during the phagocytic process in SLE patients with immunoserologically and clinically active disease.
PECAM1 (CD31) was shown to prevent activation-induced cell death in T cells. 42 Considering the high PECAM expression in macrophages from patients with SLE in our study and the data from the literature that suggests lupus correlates with an apoptotic defect in T cells, 43 we speculate that PECAM affects the survival of activated T cells and the associated molecules, influencing tolerance during the course of the disease.
Interestingly, during phagocytosis in all of the SLE patients with clinical and immunoserological changes, ADORA3 and ADORA2A were upregulated. It is known that macrophages engulfing apoptotic cells produce adenosine in sufficient amounts to trigger ADORA2, which contributes to the production of neutrophil migration factors and the inhibition of inflammation. 44 It was published that in patients with rheumatoid arthritis, the plasma adenosine level is increased, suggesting activation of the overexpressed A2A and A3 adenosine receptors. The upregulation of these two receptors was explained as a possible compensatory response to maintain the anti-inflammatory effect resulting from stimulation of these receptors. 45 In addition, an A3 adenosine agonist prevents collagen- and adjuvant-induced arthritis in experimental models by inhibiting MIP-1α and TNF-α. 46 In lieu of these findings, our results suggest possible therapies based on the manipulation of the inflammatory state by modulating these two receptors.
Another receptor upregulated during phagocytosis in patients with SLE was CD93. This receptor was shown to increase macrophage sensitivity to TLR stimulation and the concomitant TNF-α production upon lipopolysaccharide (LPS) stimulation. In patients with rheumatoid arthritis, soluble CD93 was elevated, suggesting a role in the pathogenesis of the disease. 47 Interestingly, this receptor exhibited high expression levels in SLE patients with clinically and immunoserologically active disease.
Previous work confirmed that cytokine-mediated immunity plays a crucial role in the pathogenesis of SLE. IL23/IL17 axis activation was shown to contribute to the development of nephritis in lupus-prone mice. 23 In our system, IL23A was increased in patients with clinically active disease and in one inactive patient with glomerulonephritis in their history. IL12B expression was upregulated in all of the patients. Our results are in accordance with data from the literature, suggesting that a specific blockade of the IL12 and IL23 immune pathway might be an effective therapy for SLE.
Autophagy is an evolutionarily conserved, self-cannibalistic, catabolic lysosomal degradation pathway that occurs in all cells as a homeostatic function in the process of protein and organelle turnover. It is substantially upregulated when cells need to generate intracellular nutrients and energy, and also when structural remodeling is required. 48 NOD2, an initiator of autophagy, was shown to play a role in the pathogenesis of chronic inflammatory disorders such as Crohn’s disease. 49 Genome-wide association studies have linked several single nucleotide polymorphisms in the autophagy protein ATG5 to SLE susceptibility.50–52 Here, we show that NOD2 is highly upregulated in SLE patients with active disease, and for the first time, we show that expression of the autophagy gene MAP1LC3 is increased during phagocytosis in all of the patients involved in this study.
The presented data suggest that in clinical practice, at least the two common downregulated genes, MFGE8 and ITGB5, should be tested in the differentiated macrophages of patients with non-differentiated collagenosis. The downregulation of these two genes may have a predictive role for SLE. In parallel, the level of the IL12B gene and soluble IL12B cytokine measurements would provide information about the activation of the inflammatory cascade. These data might help in the early diagnosis of SLE and contribute to a better understanding of the role of apopto-phagocytic genes in the pathogenesis and clinical presentation of the disease.
Funding
This study was supported by grants from the Hungarian National Research Fund (OTKA NI 67877 and NK-72937), the Hungarian Ministry of Health and European Union (MRTN-CT-2006-036032, MRTN-CT 2006-035624, LSHB-CT-2007-037730), and the TÁMOP 4.2.1./B-09/1/KONV-2010-0007 project, implemented through the New Hungary Development Plan and co-financed by the European Social Fund.
The study was approved by the local ethics committee (approval number DEOEC RKEB/IKEB3294-2010) and carried out according to the principles of the Declaration of Helsinki.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Footnotes
Acknowledgements
The authors would like to thank Jennifer Nagy for technical assistance in laboratory work, Ibolya Fürtös for technical assistance and advice on RNA isolation and István Szatmári for his criticism during the writing of the manuscript.