
Abstract
B-cell activating factor of the tumour necrosis factor family (BAFF) is a cytokine, mainly produced by hematopoietic cells (e.g. monocytes/macrophages, dendritic cells), indispensable for B-cell maturation. The BLISS studies have demonstrated that blocking BAFF by the human monoclonal antibody belimumab is a valuable therapeutic approach in patients with clinically and serologically active systemic lupus erythematosus (SLE). However, the defined sources of BAFF, which contributes to SLE, are still unclear. Recent findings show that BAFF expression is not restricted to myeloid cells. Since lupus nephritis is the main cause of morbidity and mortality for SLE patients, the aim of this study was to investigate whether renal tubular epithelial cells (TEC) are an important source of BAFF and thus may contribute to the pathogenesis and progression of SLE. We found BAFF expression both in cultured murine and human TEC. These results could be verified with in situ data from the kidney. Moreover, BAFF expression in the kidneys of lupus-prone MRL-Faslpr mice correlated with disease activity, and BAFF expression on TEC in biopsies of patients with diffuse proliferative lupus nephritis showed a correlation with the histopathological activity index. In vitro functional assays revealed an autocrine loop of BAFF with its binding receptors on TEC, resulting in a strong induction of colony stimulating factor-1. Finally, we identified divergent effects of BAFF on TEC depending on the surrounding milieu (‘inflammatory versus non-inflammatory’).
Taken together, our findings indicate that renal-derived BAFF may play an important role in the pathophysiology of the systemic autoimmune disease SLE.
Introduction
The hallmark of systemic lupus erythematosus (SLE) includes circulating pathogenic autoantibodies and inflammation in multiple organs. Lupus nephritis (LN) is one of the most common and serious organ manifestations of SLE. It is mediated by the infiltration of macrophages and lymphocytes and the deposition of autoantibodies in capillary walls. Advancing tubulointerstitial pathology and an increase in infiltrating leucocytes are unfavorable prognostic indicators. MRL-Faslpr mice spontaneously develop a severe autoimmune syndrome that closely resembles human SLE, characterized by glomerulonephritis (GN), lymphadenopathy, circulating autoantibodies (Abs) (anti-DNA, anti-RNP, etc.), arthritis, and vasculitis.1–4 Kidney disease in MRL-Faslpr mice is characterized by accumulation of T cells5–11 and macrophages (Mø) 12 that are required for disease. Based on the importance of T-cell and Mø function in the pathogenesis of SLE, several cytokines, including interferon gamma (IFN-γ) and interleukin (IL)-23, IL-6, IL-18 and IL-12, have been implicated in the pathophysiology of autoimmune disease.13,14 Moreover, autoreactive B cells and long-lived plasma cells have been implicated in the pathophysiology of SLE. It has been shown that long-lived plasma cells can be found in the medulla of the kidney with autochthone autoantibody secretion. 15 In preliminary studies, blockade of long-lived plasma cells with bortezomib dampens the course of the disease in experimental models and patients.16–18
The B-cell activating factor from the tumour necrosis factor (TNF) family (BAFF, B lymphocyte stimulator (Blys), TNFSF13B) was originally discovered in 1999 as a B-cell growth factor.19,20 This cytokine can be released as a soluble form after proteolytic cleavage of the homotrimeric transmembrane protein variant. Meanwhile, the crucial role of BAFF in autoimmunity is well accepted. Elevated levels of BAFF correlate with autoimmune disease in humans, mice and dogs. 21 Transgenic overexpression of BAFF led to an accelerated lymphoproliferative disorder in mice, whereas BAFF-deficient mice were protected from SLE. The efficacy of anti-BAFF treatment with belimumab has been reported in two phase III trials resulting in the approval of belimumab as the first and sole biologic therapy for SLE.22,23 In recent years novel functions of BAFF in different tissues other than B cells emerged suggesting a role of BAFF in cell survival in cancer and infection. 24
In the present study we analysed the role of renal BAFF expression in LN by different approaches: (1) kinetics of BAFF expression in the course of LN in MRL-Faslpr mice, (2) functional effects of BAFF in the cytokine milieu of LN and (3) expression pattern of BAFF in patients with LN. We now report for the first time that BAFF and its receptors (TNFRSF13B/TACI, TNFRSF17/BCMA, and TNFRSF13C/BAFFR) are expressed by tubular epithelial cells (TEC) in the kidney. In addition, BAFF is upregulated at a very early stage in the course of the disease, thus, intrarenal BAFF is closely associated with the inflammatory cascade in the lupus kidney. Furthermore, BAFF is both a potent inducer of renal parenchymal cell-derived pro-inflammatory cytokines and a good biomarker for the histopathological activity of diffuse proliferative LN III/IV but not membranous LN V. Therefore, targeting intrarenal BAFF may have the potential to downregulate LN at an essential pathophysiological step in the course of SLE disease.
Material and methods
Mouse strains
MRL/MpJ-Faslpr/Faslpr (MRL-Faslpr) and C57Bl/6 (B6) mice were purchased from The Charles River Laboratories (Boston, MA, USA). Mice were bred and housed in our pathogen-free facilities at Johannes-Gutenberg-University in Mainz, Germany. Only female mice were used for analyses. The use of mice in this study was reviewed and approved by the Standing Committee on Animals at the University of Mainz.
Isolation of renal TEC
We isolated and cultured TEC derived from MRL-Faslpr mice as previously reported. 25
Serum and renal biopsy specimens
To diagnose LN (comporting with the International Society of Nephrology/Renal Pathology Society (ISN/RNP) 2004 classification) 26 and assess histopathology activity and chronicity indices, 27 human kidney sections from renal biopsy specimens were provided by the Department of Pathology, Friedrich-Alexander University Erlangen-Nuremberg, Germany. Renal pathologists, without access to the patient’s clinical data, evaluated these biopsy specimens. We analysed serum samples that were collected in Mainz, Germany. Specimens were taken from patients who fulfilled at least four of the American College of Rheumatology criteria for the classification of SLE, after informed consent. Volunteers (age range, 18–70 years) were screened for health by exclusion of any prior kidney diseases, diabetes, hypertension, and autoimmune diseases. Serum was collected at a single visit. Freshly drawn blood samples were collected, centrifuged, divided into aliquots, and stored at −80℃ before analysis. The use of these specimens was reviewed and approved by the Standing Committee for Clinical Studies of the Johannes-Gutenberg University in adherence to the Declaration of Helsinki. All samples were analysed retrospectively in Mainz.
Disease activity
Disease activity was evaluated by standard clinical serologic activity measures (C3c, C4, antinuclear antibodies (ANA), anti-double-stranded DNA (anti-dsDNA) antibodies, creatinine, C-reactive protein (CRP), erythrocyte sedimentation rate (ESR) (after one hour/two hours)) and urine measures (proteinuria (24-hour collection) and active sediment) as previously described. 28 The following standard values of serologic activity markers were determined: C3 (0.9–1.8 g/l), dsDNA (200–1000 IU/ml) by enzyme-linked immunosorbent assay (ELISA), proteinuria (<150 mg/24 hours) by immunoturbidimetric assay.
Immunostaining
Kidneys were fixed in 10% neutral buffered formalin, embedded in paraffin, sectioned (4 µm) by the tissue bank of the University Medical Center Mainz, and stained for the presence of BAFF expression and its receptors as previously detailed. 29 Mouse kidney sections and kidney biopsy sections were incubated with primary Ab, rabbit anti-human BCMA Ab (Cat. No. 5972, abcam), rabbit anti-human TACI Ab (Cat. No. 79023, abcam), mouse anti-human BAFF-R Ab (Cat. No. 16232, abcam) and rabbit anti-human/mouse BAFF (Cat. No. Bs-2431R, BIOSS). The primary Ab was detected by incubating with biotinylated goat anti-rabbit Ab and goat anti-mouse Ab, followed by development with 3-3-diaminobenzidine (Vector Laboratories). Nonspecific binding was determined using rabbit anti-human immunoglobulin (Ig)G or mouse anti-human IgG (eBioscience). We determined the percent-positive TEC in 10 randomly selected high-power fields (HPFs).
Immunofluorescence staining
We stained cultured primary mouse/human TEC or human proximal TEC cell line (HK2) for the presence of BAFF using rabbit anti-human/mouse BAFF (Cat. No. Bs-2431R, BIOSS). Cells were fixed with ice-cold methanol and incubated with the rabbit anti-BAFF Ab followed by incubation with FITC-conjugated rat anti-rabbit Ab (Invitrogen).
Cyto-ELISA
We stained on gelatin-coated 96-well plates cultured primary mouse/human TEC or human proximal TEC cell line (HK2) for the presence of BAFF using rabbit anti-human/mouse BAFF (Cat. No. Bs-2431R, BIOSS). Cells were fixed with ice-cold methanol and incubated with the rabbit anti-BAFF Ab for 40 minutes followed by incubation with a horseradish-peroxidase-conjugated rat anti-rabbit Ab (eBioscience) for 40 minutes. Following washing the substrate solution tetramethylbenzidine was added to each well, and extinction was measured after stopping the color reaction with 1 N H2SO4 within 15 minutes at 450 nm.
ELISA
To quantify mouse and human BAFF levels in the circulation and colony stimulating factor-1 (CSF-1) in TEC, serum samples and supernatants were analysed using a mouse BAFF ELISA (Duoset ELISA, Cat. No. DY2106, R&D Systems) or human BAFF ELISA (Duoset ELISA, Cat. No. DY124, R&D Systems) and a mouse and human CSF-1 ELISA (R&D Systems) according to the manufacturer’s instructions.
Isolation and reverse transcription (RT) of messenger RNA (mRNA) from cultured cells
The mRNA from the cultured cells was isolated with the RNeasy Mini Kit (Qiagen GmbH) according to the manufacturer’s instructions. The concentration of the isolated RNA was measured and 1 µg RNA used for RT.
Detection of BAFF-expression by conventional and quantitative RT-polymerase chain reaction (qPCR)
For the detection of BAFF expression by TEC or renal tissue, conventional PCR of the generated complementary DNA (cDNA) was performed with the Fast Start Taq Polymerase Kit (Roche) using the following program: 10 minutes at 95℃ for initial activation of DNA polymerase, 30 cycles of 45 seconds at 95℃, 30 seconds at 57℃ and 1 minute at 72℃. The following primers were used for amplification: h-BAFF1 forward: 5‘-CGC CAT GGG ACA TCT AAT TC-3‘; reverse: 5’-ACA GCA GTT TCA ATG CAC CA-3’; h-BAFF2 forward: 5’-AAT TCA GAG GAA GAA GGT CC-3’; reverse: 5’-ATG TGA CAT CTC CAT CCA GT-3’. The PCR products were separated on 1.5% agarose gels and visualized with F-BLUE (Biomol).
To measure differential mRNA-expression quantitative RT-PCR was performed using the Quantitect SYBR Green PCR Kit (Qiagen) in combination with the Quantitect Primer Assays (human: Hs_TNFSF13B_1_SG, Cat no: QT00094759; Hs_TNFRSF13C_1_SG, Cat no: QT00217140, Hs_TNFRSF17_1_SG, Cat no: QT00200018; Hs_TNFRSF13C_1_SG, Cat. no: QT00028427; murine: Mm_Tnfsf13b_1_SG, Cat no: QT00117215; Mm_Tnfrsf13b_1_SG, Cat no: QT00149891, Mm_Tnfrsf13b_1_SG, Cat. no: QT00143178; Mm_Tnfrsf17_1_SG, Cat no: QT00116606). Beta actin was used as a housekeeping gene (human: Hs_ACTB_2SG, Cat no.: QT016804; murin: Mm_Actb_1_SG, Cat. No.: QT00095242). For this purpose, the following real-time PCR protocol was used: 15 minutes at 95℃ for initial activation of DNA polymerase, 45 cycles of 15 seconds at 95℃, 30 seconds at 57℃, 30 seconds at 72℃ and 30 seconds at 40℃ for cooling in Roche LightCycler. The data were analysed by the ΔΔ–CT method.
Luciferase reporter assay
The murine BAFF promoter was amplified with the following primers: for: 5′-TTTT
Western blot
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (10% acryl-amide) and Western blotting of kidney/spleen tissue, primary isolated and cultured TEC and freshly isolated peripheral blood leucocytes (PBLs) with goat anti-mouse BAFF (Cat. No. AF2106, R&D Systems) were carried out as previously described. 25
Proliferation and apoptosis of TEC
We stimulated cultured TEC for 24 hours with different cytokines (TNF-α (human, Cat. No. 210-TA-005/mouse, Cat. No. 410-MT-010; 25 ng/ml); IL-6 (human, Cat. No. 206-IL-010/mouse, Cat. No. 406-ML-005, 20 ng/ml); CSF-1 (human, Cat. No. 216-MC-005/mouse, Cat. No. 416-ML-010; 200 ng/ml), BAFF (human, Cat. No. 2149/mouse, Cat. No. 2106; 200 ng/ml) from R&D Systems, IFN-γ (Cat. No. 300-02, Preprotech, 20 ng/ml) and lipopolysaccharide (LPS) (Cat. No. alx-581-008-l002; 200 ngml) from Alexis. After incubating for 24 hours, we analysed TEC proliferation with the Cell Proliferation Reagent WST-1 (Cat. No. 1164480700, Roche) and the cytotoxicity using the ToxiLight BioAssay Kit (Cat. No. LT07-217, Lonza) following the manufacturer’s instructions.
Statistical analysis
We used the nonparametric Mann-Whitney U test for comparison between two groups and the Kruskal-Wallis test for comparisons between three or more groups. For correlation analysis, we used the Spearman correlation coefficient.
The data represent the mean ± SEM and were prepared using GraphPad Prism Version 7.0 (Graph Pad, San Diego, CA, USA).
Results
BAFF is expressed in the kidney of lupus-prone MRL-Faslpr mice and its expression level correlates with disease activity
To evaluate the role of BAFF in the kidney we analysed the expression of BAFF at transcript and protein level in normal B6 and lupus-prone MRL-Faslpr mice. BAFF mRNA could be detected both in normal and autoimmune mouse kidneys by qPCR. While there was no difference in relative expression between the strains at 0.75, 1.5 and 3 months of age, BAFF mRNA expression increased by 10- to 15-fold in the MRL-Faslpr mice at 5 months of age as compared to healthy B6 mice (Figure 1(a), left panel). In addition, we noticed BAFF protein in the spleen and kidneys of both strains, but only in MRL-Faslpr mice we detected a marked increase in BAFF expression at 5 months of age in the kidney (Figure 1(a) right panel). By immunostaining we could further demonstrate that TEC were the major intrarenal site for BAFF protein expression (Figure 1(b)). In contrast to the renal BAFF expression, circulating BAFF levels were significantly higher in lupus mice compared to normal even at very young ages (Figure 1(c)). Up to 3 months of age no overt signs of autoimmune or inflammatory activity could be detected in the kidneys of these MRL-Faslpr mice suggesting extra-renal upregulation of BAFF levels (data not shown).
BAFF is expressed in the kidneys of lupus-prone MRL-Faslpr mice and its expression level correlates with disease activity. (a) Left panel: BAFF mRNA expression (qPCR) in kidney and spleen tissue of MRL-Faslpr and B6 mice at different time points (0.75, 1.5, 3.0 and 5.0 months of age); Right panel: BAFF protein expression detected by Western blot analysis in kidney and spleen tissue in MRL-Faslpr and B6 mice at 0.75 and 5 months age. Blots were performed three times with different samples. WEHI cell line used as control (peripheral myelomonocytic leukaemia leucocytes from Balb/c mice). (b) BAFF expression in kidneys of MRL-Faslpr mice in comparison to B6 mice at 5 months of age. Representative microphotographs are shown (n = 5–8 per group) (c) Serum BAFF levels with increasing age in MRL-Faslpr mice in comparison to B6 mice. Values are means ± SEM. Analysis was performed using the Mann-Whitney U test. BAFF: B-cell activating factor; mRNA: messenger RNA; qPCR: quantitative reverse-transcription polymerase chain reaction.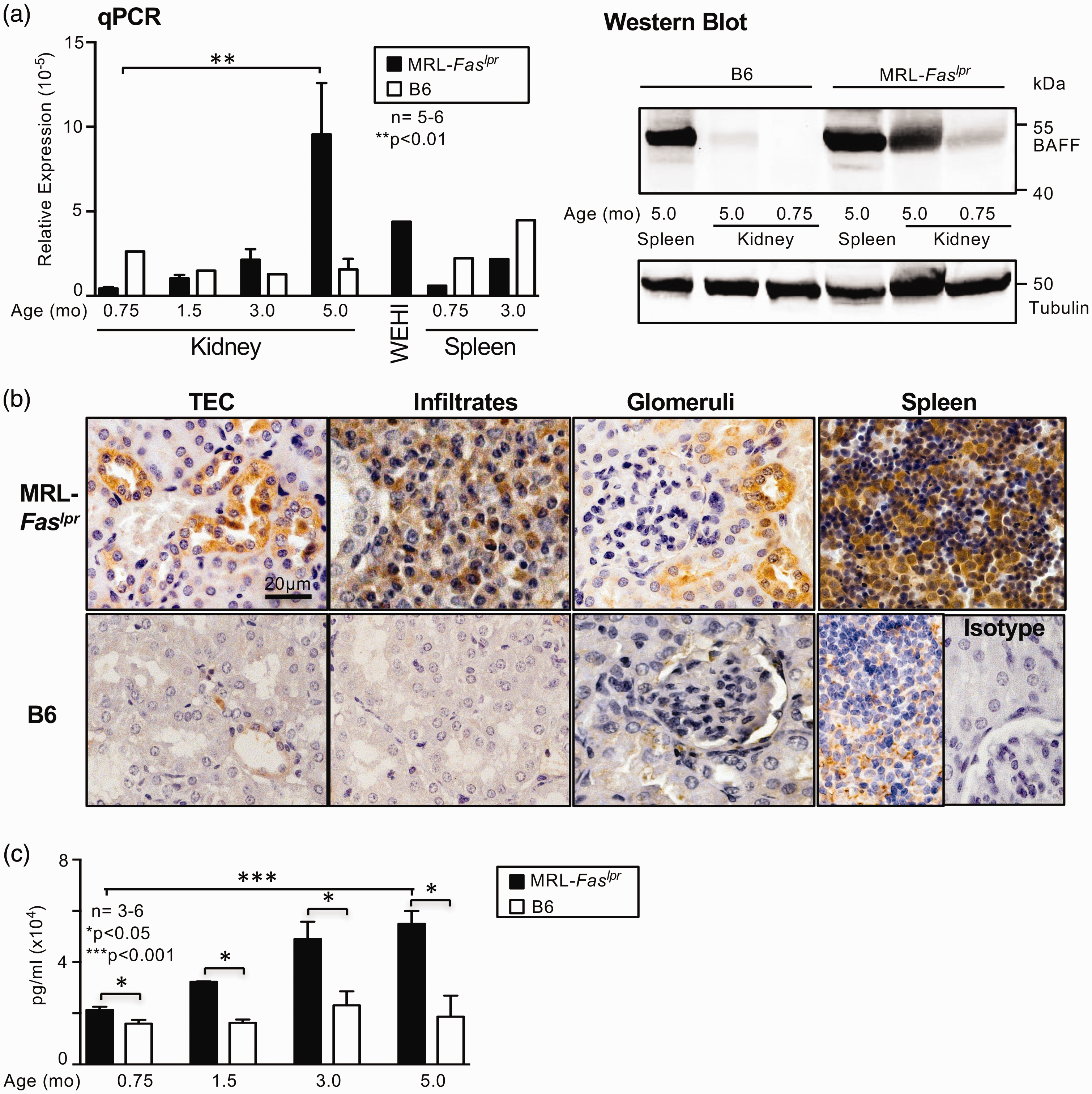
Renal TEC express BAFF
Based on the expression pattern of BAFF in the kidney, we further evaluated whether TEC are capable of de novo BAFF expression. BAFF mRNA could be detected in primary cultured human TEC and a human renal epithelial cell line HK-2, whereas other cell lines such as MCF-7, HeLa or HEK did not express BAFF mRNA (Figure 2(a)). Interestingly, we found BAFF mRNA not only in spleen and bone marrow as expected but also in heart tissue (Figure 2(a)). Furthermore, BAFF protein could be detected in primary cultured human and mouse TEC by Western blot technique, immunofluorescence and cyto-ELISA (Figure 2(b), (c)). Moreover, by evaluating the BAFF promotor activity we could show that the BAFF promotor is active in primary isolated mouse and human TEC (Figure 2(d)). Circulating factors induce promotor activity in TEC even in very young lupus mice without clinical signs of disease (1 month of age, Figure 2(d)). Thus, renal TEC are a source of BAFF mRNA and protein suggesting that the kidney itself produces a potent mediator for both normal immune response and autoimmune deviation.
Human and mouse TEC express BAFF. (a) BAFF mRNA expression in primary isolated human TEC and TEC cell line (HK2). (b) BAFF protein expression by Western blot analysis in primary isolated human TEC, in HK2 cells and primary isolated mouse TEC. Freshly isolated human and mouse peripheral blood leucocytes served as positive controls. Blots were performed three times with different samples. (c) Left panel: BAFF protein expression of cultured primary TEC and HK2 cells detected by immunofluorescence, representative microphotographs are shown; Right panel: BAFF expression detected by cyto-ELISA in human and mouse TEC. (d) Left panel: BAFF promotor activity evaluated in primary mouse TEC, WEHI cells (peripheral myelomonocytic leukaemia leucocytes from Balb/c mice) served as positive control. Right panel: Detection of BAFF promotor activity following incubation of cultured primary mouse TEC with serum of MRL-Faslpr (1 and 5 months of age) and B6 (5 months of age) mice. Values are means ± SEM. Analysis was performed using the Mann-Whitney U test. TEC: tubular epithelial cells; BAFF: B-cell activating factor; mRNA: messenger RNA; ELISA: enzyme-linked immunosorbent assay.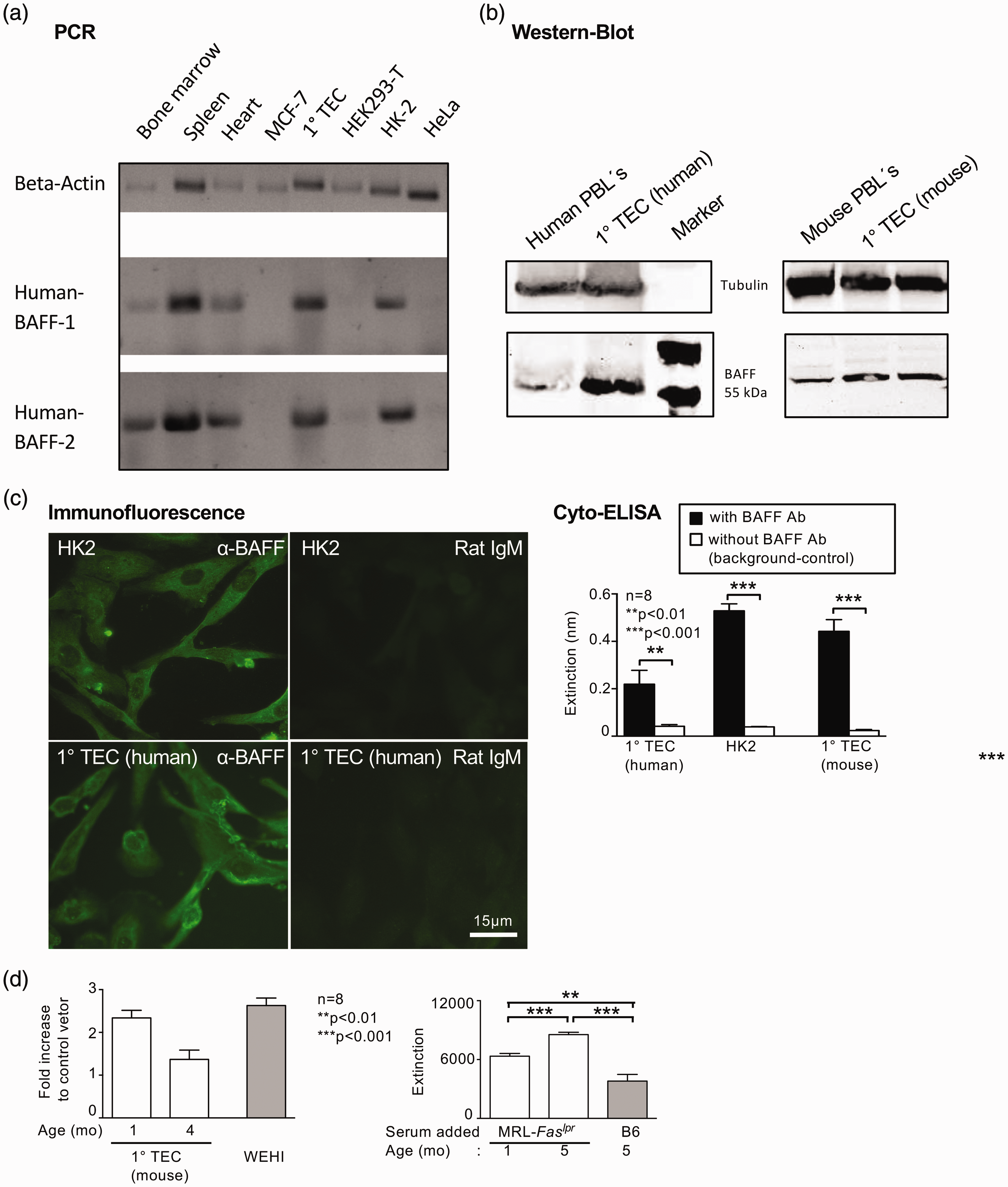
BAFF is a strong inducer of CSF-1 expression by TEC and vice versa
Since we have recently shown that CSF-1 plays a crucial role in the pathophysiology of LN,29,30 we assessed a putative link between BAFF and CSF-1 production by TEC in vitro. In fact, BAFF was a strong inducer of CSF-1 production by human TEC, even more potent than TNF-α or LPS (Figure 3(a), left panel). Since CSF-1 stimulation of TEC in turn leads to an increase in BAFF expression, both cytokines appear to form an ‘autocrine loop’ in TEC (Figure 3(a), right panel (data are representative for mouse and human TEC, data for mouse TEC not shown). Looking at direct effects of BAFF on TEC (proliferation/cytotoxicity) we found a more complex picture: Within a ‘non-inflammatory milieu’ BAFF increased proliferation and decreased cytotoxicity of TEC in a dose-dependent manner (Figure 3(b1)). By contrast, in an “inflammatory milieu” BAFF downregulated the effects mediated by pro-inflammatory cytokines resulting in reduced proliferation and increased cytotoxicity of TEC (Figure 3(b2)). Data represent the results of human TEC; experiments with mouse TEC showed comparable results (data not shown).
BAFF is a strong inducer of CSF-1 expression and CSF-1 of BAFF by TEC resulting in reduced proliferation and increased cytotoxicity of TEC in an inflammatory milieu. (a) CSF-1 and BAFF expression following stimulation with BAFF, CSF-1, TNF-α, and LPS by human TEC detected by ELISA. (b1) Evaluation of proliferating TEC and cytotoxicity of TEC following stimulation with BAFF (b2) Evaluation of proliferating TEC and cytotoxicity of TEC after prestimulation (inflammatory milieu) with TNF-α, LPS, IL-6 and CSF-1 followed by stimulation with BAFF. All experiments were repeated four times; representative data are shown. Values are means ± SEM. Analysis was performed using the Mann-Whitney U test. BAFF: B-cell activating factor; CSF-1: colony stimulating factor 1; TNF-α: tumour necrosis factor alpha; LPS: lipopolysaccharide; TEC: tubular epithelial cells; ELISA: enzyme-linked immunosorbent assay; IL-6: interleukin 6.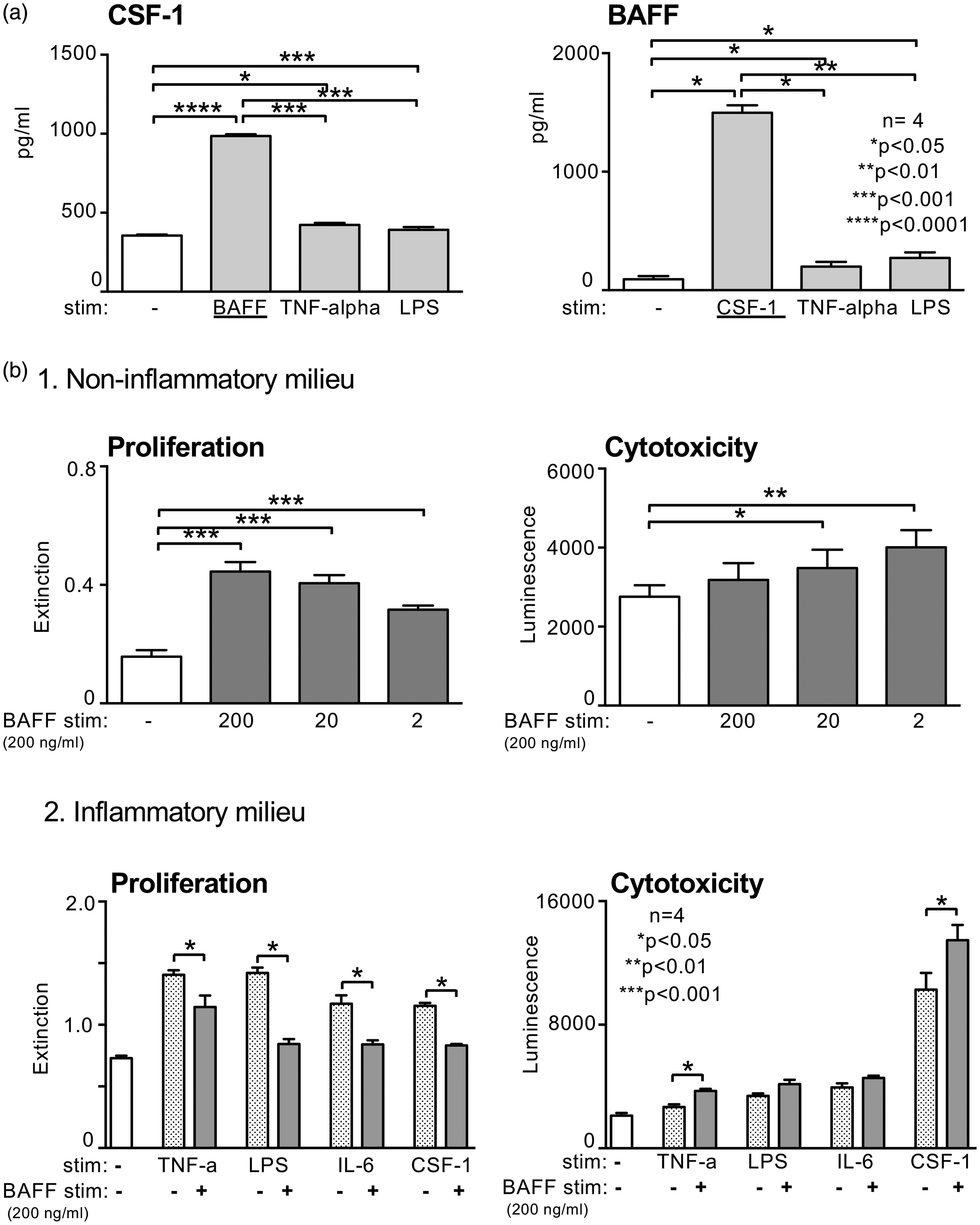
Different expression pattern of the BAFF binding receptors BAFF-R, BCMA and TACI in MRL-Faslpr mice
Based on our in vitro functional data suggesting the presence of an ‘autocrine loop’, we hypothesized the expression of BAFF binding receptors in the kidneys of lupus-prone mice. We analysed the relative receptor mRNA expression of BAFF-R, BCMA and TACI in the spleen and kidneys of MRL-Faslpr mice at different stages of the disease (Figure 4). We found a significant increase in the expression of all receptor mRNA in the kidneys at 5 months of age compared to younger lupus mice with yet non-overt kidney pathology. By comparison, the expression of BAFF-R in the spleen was several-fold higher than in the kidney (Figure 4(a)). To distinguish between receptor expression on infiltrating cells or renal parenchymal cells, we analysed the proportion of receptor-positive TEC at different ages. Interestingly, the expression of the BAFF-R was significantly upregulated during the course of the disease, while BCMA expression was stable throughout all time points. By contrast, almost no TACI expression could be visualized on TEC by immunostaining (Figure 4(a), right panel). Similarly, only a faint signal of TACI was obtained when analysing the mRNA expression of the receptors in cultured unstimulated TEC while BAFF-R and BCMA showed relevant expression (Figure 4(a), middle panel). However, analysing the receptor expression following stimulation with BAFF, CSF-1 or LPS showed an upregulation of all three receptors on TEC (Figure 4(b)). Thus, there is a distinct expression pattern of BAFF binding receptors on TEC under non-inflammatory and inflammatory conditions and in the kidneys of lupus-prone mice suggesting a more complex transmission of BAFF signals within the kidney.
Different expression pattern of the BAFF binding receptors BAFF-R, BCMA and TACI in MRL-Faslpr mice. (a) BAFF-R, BCMA and TACI mRNA expression in kidney and spleen tissue in MRL-Faslpr mice at different ages (0.75, 3.0 and 5.0 months of age) (left panel), in unstimulated primary cultured mouse TEC (middle panel) and protein expression in TEC evaluated by immunostaining in MRL-Faslpr at 0.75, 3.0, 5.0 months of age (right panel). (b) BAFF-R, BCMA and TACI mRNA expression of primary cultured TEC following stimulation with BAFF, CSF-1 or LPS. Values are means ± SEM. Analysis was performed using Mann-Whitney U test. BAFF: B-cell activating factor; mRNA: messenger RNA; TEC: tubular epithelial cells; CSF-1: colony stimulating factor 1; LPS: lipopolysaccharide.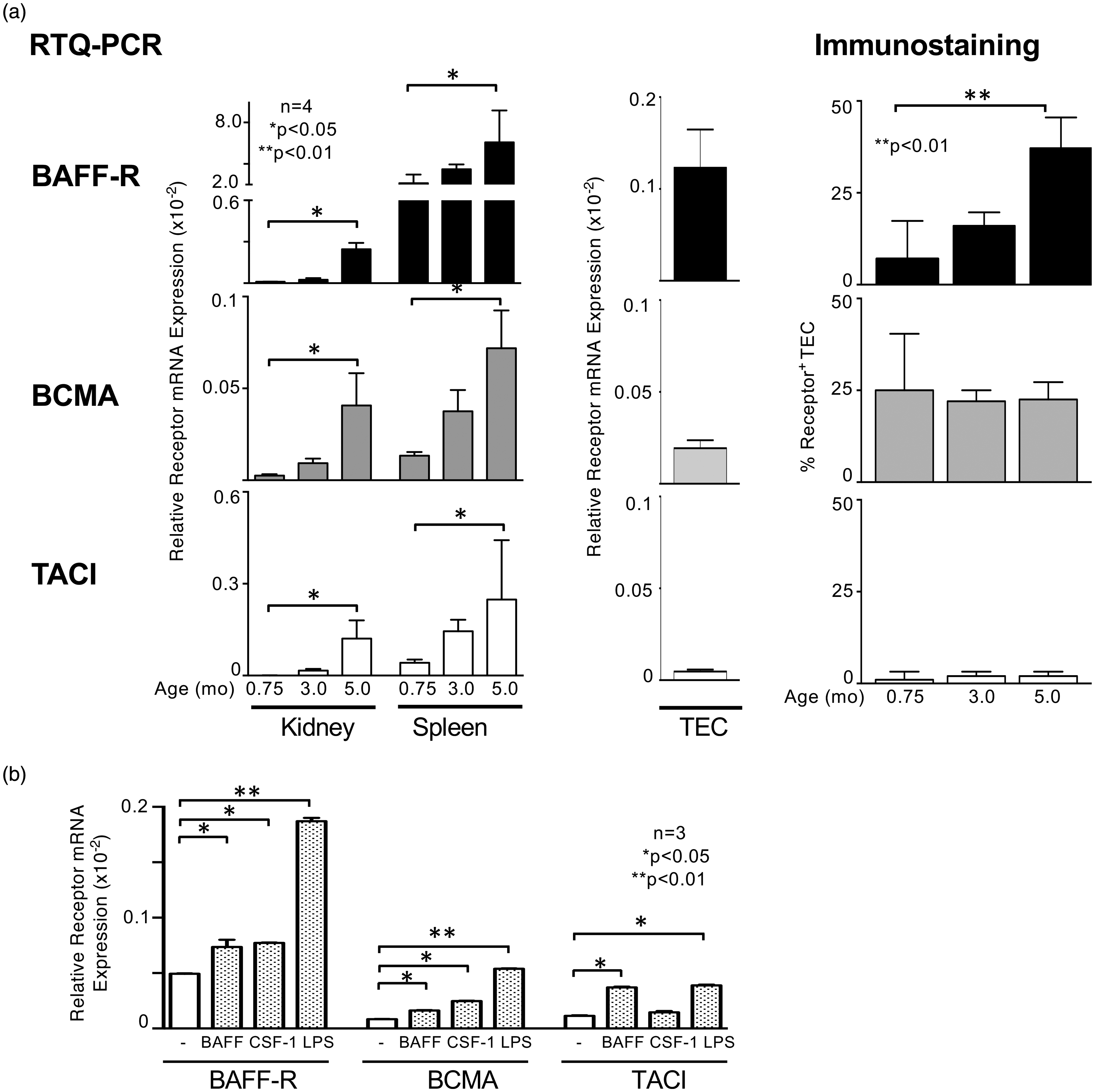
BAFF expression in kidney biopsies of patients correlates with LN type and activity
Based on our results of the experimental mouse model we sought to investigate the role of BAFF expression in lupus patients with nephritis. As depicted in Figure 5(a), BAFF is over-expressed in TEC of patients with LN compared to kidney biopsies of patients with thin-glomerular-basement-membrane (GBM) or haematuria without renal pathology. Moreover, the number of BAFF-expressing TEC correlates with the histopathological activity index but not with the chronicity index (Figure 5(b)). In addition, we detected significantly more BAFF-positive TEC in diffuse proliferative LN type III/IV (ISN/RPS classification) compared to the membranous LN type V (Figure 5(c)). While circulating levels of BAFF were highly upregulated in lupus patients compared to healthy controls (Figure 5(d)), we did not find a good correlation of serum-BAFF levels with serological activity markers or proteinuria up to 12 months following diagnosis of LN (types I–VI) (Figure 5(e), left panel). However, dividing the histological type of LN in subgroups reveals that serum BAFF levels correlate with serological activity markers (complement C3c, dsDNA) of LN and proteinuria in patients with type III or IV LN. Thus, these data support a complex pathophysiological of LN involving BAFF signalling.
BAFF expression in kidney biopsies of patients correlates with type of LN and activity. (a) BAFF expression in LN biopsies (n = 20) and biopsies with thin glomerular basal membrane or haematuria without kidney pathology (n = 8) as control tissue evaluated by immunostaining. Representative microphotographs are shown, magnification × 40. (b) Correlation of BAFF expression in kidney with histopathology activity and chronicity indices in LN. (c) BAFF expression by TEC stratified according to ISN/RPS. (d) Serum BAFF expression in patients with LN compared to healthy controls quantified using ELISA. Analysis was performed using the Mann-Whitney U test. (e) Longitudinally monitoring BAFF in serum positively correlates with disease activity only in patients with type III and IV LN. Serum BAFF levels at diagnosis of biopsy-proven LN and during therapy in comparison to conventional clinical disease activity measures. BAFF was quantified using ELISA. Values are means ± SEM. Analysis was performed using the Kruskal-Wallis test for multiple comparisons. BAFF: B-cell activating factor; LN: lupus nephritis; TEC: tubular epithelial cells; ISN/RPS: International Society of Nephrology/Renal Pathology Society; ELISA: enzyme-linked immunosorbent assay.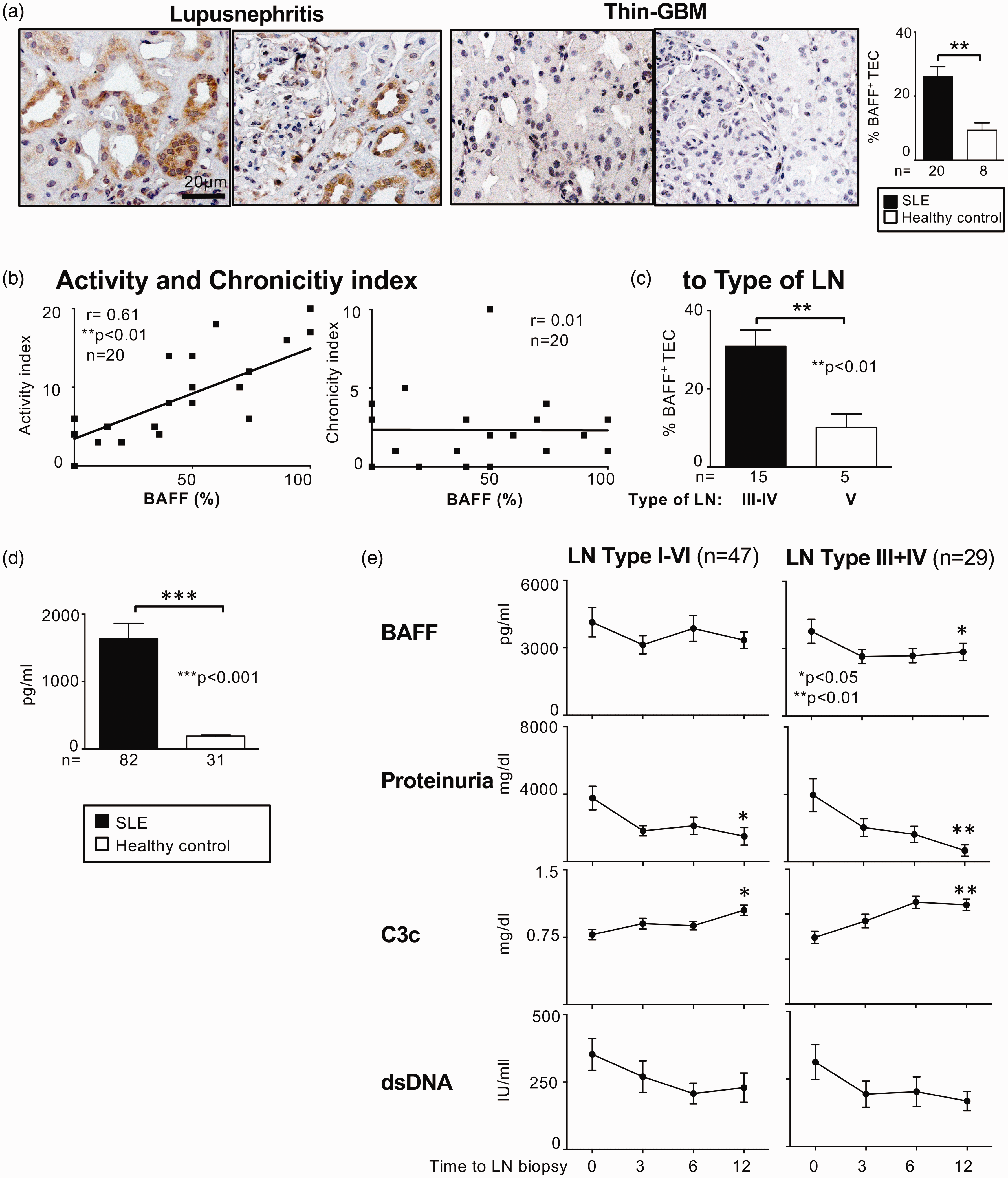
Expression pattern of BAFF binding receptors in biopsies of patients with LN
In vitro cultured human TEC and a renal tubular epithelial cell line (HK2) showed predominantly BAFF-R expression followed by BCMA transcripts while TACI mRNA could not be detected (Figure 6(a)). However, all three BAFF binding receptors are upregulated following stimulation with BAFF, CSF-1 or LPS, most prominent the BAFF-R comparable to the results seen in mouse TEC (Figures 6(c) and 4).
Expression pattern of BAFF binding receptors in biopsies of patients with LN. (a) BAFF-R, BCMA and TACI mRNA expression in human unstimulated primary TEC and HK2 cells. (b) BAFF-R, BCMA and TACI protein expression evaluated by immunostaining in LN biopsies (n = 15). Representative microphotographs are shown, magnification ×40. (c) BAFF-R, BCMA and TACI mRNA expression of primary cultured human TEC following stimulation with BAFF, CSF-1 or LPS; used concentration for stimulation 200 ng/ml. Values are means ± SEM. Analysis was performed using the Mann-Whitney U test. BAFF: B-cell activating factor; mRNA: messenger RNA; TEC: tubular epithelial cells; LN: lupus nephritis; CSF-1: colony stimulating factor 1; LPS: lipopolysaccharide.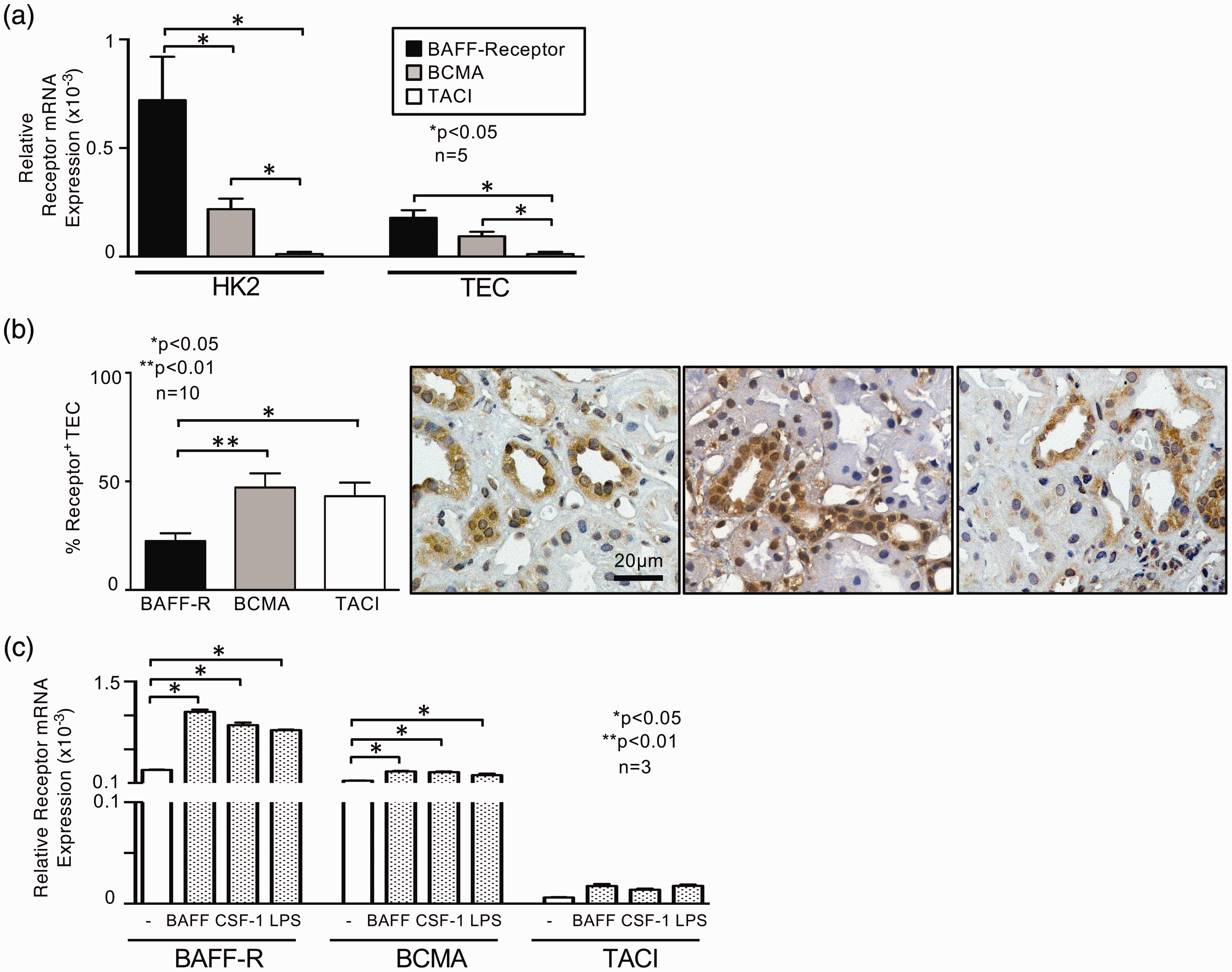
However, in contrast to the situation in the lupus-prone MRL-Faslpr mice (Figure 4), the expression pattern of the receptors in biopsies of patients with LN displayed a great difference (Figure 6(b)): BCMA and TACI receptor were predominantly expressed (ca. 50% positive TEC) whereas only 25% of TEC were BAFF-R positive suggesting a different (patho)physiological role of the BAFF binding receptors in men and mice.
Discussion
SLE is a heterogenic autoimmune disease characterized by circulating autoantibodies and different organ manifestations. It is well accepted that in patients with renal involvement prognosis and survival is greatly reduced. Korbet and colleagues showed that the 10-year survival rate of patients with SLE depends on whether the LN is in remission. 31 One explanation for this observation is the huge amount of cytokines and chemokines secreted by the kidney itself. We have recently shown that CSF-1 produced by TEC, for example, is a crucial mediator of autoimmune kidney disease. 28 Now, we report that the kidney is even capable of expressing BAFF, one of the most potent B-cell growth factors. BAFF is expressed by TEC and the expression correlates with the histopathological activity index in kidney biopsies. Moreover, BAFF not only activates B cells but further acts in an autocrine loop to induce CSF-1 in TEC, which in turn stimulates the cascade of autoimmune inflammation and macrophage influx. 28 Furthermore, several groups reported the presence of long-lived plasma cells within the ‘kidney niche’ in mouse models and patients with LN,32–34 wondering how these cells can survive in this ‘extramedullary niche’. Since TEC-derived BAFF is almost constitutively expressed in the kidney of lupus-prone MRL-Faslpr mice, we speculate that intrarenal BAFF expression is the local growth cytokine required for the survival of long-lived plasma cells. Thus, the kidney has the capacity to activate both the innate and adaptive immune response, suggesting that it is not just an innocent bystander in SLE, but even fosters the autoimmune process. As lupus-prone mice deficient in BAFF or treated with BAFF blockade suffer from GN and produce autoantibodies but do not develop kidney failure,35–37 the intrarenal BAFF expression might be a “conditio sine qua non” in murine lupus to get this severe disease. 38 The correlation of BAFF expression with the histopathological activity index may reflect a direct effect of BAFF on TEC in proliferative LN. But a secondary association of BAFF expression with proliferative forms of LN cannot be excluded and further in vivo experiments will elucidate the distinct role of BAFF in LN.
Originally, BAFF was identified as a critical B-cell maturation and survival factor19,20 leading to its exceptional role in autoimmune diseases and the development of the first Food and Drug Administration (FDA)-approved successful biologic treatment of lupus with belimumab.22,23 Meanwhile, the spectrum of the disease entities in which BAFF is involved extends far beyond autoimmunity: e.g. cancer, infection, asthma and transplantation. 24 It could be shown that the physiological role of BAFF is not restricted to B cells, as monocytes and activated T cells express BAFF receptor. On the one hand BAFF sustains a Th1-based immune response and promotes the expansion of regulatory T cells (Tregs)39,40 and on the other hand it also promotes monocyte survival and differentiation to macrophages. 41
Moreover, BAFF expression is no longer restricted to neutrophils, monocytes, macrophages or dendritic cells but can also be found in a growing number of different cells (astrocytes, skin keratinocytes and adipocytes etc.). 24 Like many ligands in the TNF superfamily BAFF exerts pleiotropic effects, both beneficial and detrimental.
In our in vitro studies we could unravel the surrounding milieu as one possible contributing factor. In a non-inflammatory milieu BAFF induces dose-dependently TEC proliferation, whereas in an inflammatory milieu with different pro-inflammatory cytokines BAFF downregulates the proliferative response. A similar effect was reported by Gao et al. for TNF-like weak inducer of apoptosis (TWEAK). They could show that TWEAK-induced renal TEC death requires a concomitant inflammatory microenvironment. 42 Therefore, constitutive BAFF expression could be necessary under non-inflammatory conditions to protect the organism by activating proliferation and activation of B cells and monocytes. On the other hand under already preexisting inflammatory conditions the intrarenal BAFF expression is overwhelming resulting in decreased proliferation and increased cytotoxicity of TEC.
However, the pathomechanisms are still unclear. Another factor that may influence the physiological effects of BAFF is the expression pattern of the three BAFF binding receptors BAFF-R, BCMA and TACI. We have shown that BCMA-expression by TEC was stable throughout the development of LN in MRL-Faslpr mice, whereas the BAFF-R expression was greatly upregulated during the course of the disease. Interestingly, it has been shown that soluble BCMA isoforms might function as natural decoys produced to regulate levels of free BAFF, which could also explain our steady expression level in TEC. 43
Moreover, we have shown a different expression pattern of the BAFF binding receptors BAFF-R, BCMA, and TACI in mice and humans. Since BAFF and a proliferation-inducing ligand (APRIL) bind with different affinities to BCMA, TACI and BAFF-R, this receptor expression pattern could represent different types and severity of lupus disease resulting in different therapeutic options (BAFF versus APRIL blockade). And the distinct expression pattern of the BAFF receptors in humans and mice may reflect the importance of BAFF in the mouse-model while in humans the significance of BAFF versus APRIL has to be further elucidated.
We have shown that BAFF is constitutively expressed in renal TEC and that its level is increased under inflammatory conditions. Renal epithelial cells are positioned at the interface between the internal milieu and external environment. Therefore, it comes as no surprise that they serve as primary guardians of the kidney and the whole organism. 44 Uropathogens, which pass the front-line defenses of the kidney and attach to the renal tubular epithelium, activate the Toll-like receptor signaling. 45 These infected or injured TEC release various pathogen-associated molecular pattern molecules (PAMPs) or damage-associated molecular patterns (DAMPs) that activate receptors on renal cells. Furthermore, a number of pattern recognition receptors including Toll-like receptors and nucleotide oligomerization domain (NOD)-like receptors are constitutively expressed in many parts of the kidney. 46 In this context, the constitutive expression level of membrane-bound BAFF in kidneys from normal mice may form an innate defense mechanism, ready to be released as soluble BAFF in case of an infection or danger signal to stimulate growth and activity of B cells and monocytes.
Clearly, more studies are needed to elucidate the complex network of BAFF and its binding receptors in renal epithelial cells.
Footnotes
Acknowledgements
We thank Professor Kerstin Amann for providing kidney biopsy tissue, Dr Knut Müller, Medical Department, Immunology and Rare Diseases Business Unit, GlaxoSmithKline, München, for fruitful discussions, and Mitra Golbas and Sabine Schmitt for technical support.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by GlaxoSmithKline.