
Abstract
Background
Systemic lupus erythematosus (SLE) is a complex autoimmune disorder characterized by a malfunction of the body’s immune defense system.
Objective
The objective of the present investigation was to examine the expression and diagnostic significance of NRIR in SLE and to prove whether it is involved in the progression of SLE.
Methods
The study involved 110 participants, including 55 healthy individuals and 55 SLE patients. The expression levels of NRIR, miR-31-5p, and ICAM-1 were measured using qRT-PCR. The ROC curve was performed to assess the diagnostic significance of NRIR in SLE patients. Pearson correlation analysis was utilized to explore the relationship between NRIR and other indicators. Cytokines including IL-4, IL-6, and IL-21, along with IgG levels, were assessed using ELISA. The interaction between NRIR and miR-31-5p was validated using a dual-luciferase reporter assay.
Result
Upregulated expression of NRIR was observed in individuals with SLE, serving a diagnostic function for SLE. Additionally, abnormal expression of NRIR impacted the viability of CD4+ T cells within SLE patients. NRIR could negatively modulate the expression of miR-31-5p.
Conclusion
LncRNA NRIR may be a potential biomarker for SLE and is likely involved in the progression of SLE.
Introduction
Systemic lupus erythematosus (SLE) is a multifaceted autoimmune disease in which the body’s immune system erroneously targets its own healthy cells and tissues. 1 The precise etiology and pathophysiological mechanisms that precipitate the autoimmune response in SLE, similar to many other autoimmune disorders, remain elusive. 2 Although the definitive cure for SLE is not yet available, the condition can be effectively managed with appropriate medications. Nonetheless, the mortality rate for individuals with SLE remains significantly elevated, approximately two to three times higher than that of the general population. This increased risk is particularly pronounced in females and individuals aged 65 years or older.3,4 While pharmacological treatments and multidisciplinary approaches primarily aim to alleviate symptoms and may potentially slow disease progression, they do not offer a complete healing solution. 1 Consequently, it is importance to enhance the capacity for early diagnosis of SLE to ensure effective treatment.
The disease presents a multifaceted challenge, affecting various organs with substantial variability in clinical manifestation and severity, complicating accurate identification and management. 5 Recent research highlights that numerous long non-coding RNAs (lncRNAs) are aberrantly expressed and may be crucial in the pathogenesis of SLE. 6 Yujie Bu et al observed distinct differences in lncRNA expression between SLE patients and healthy controls. This study identified 240 lncRNAs with altered expression levels, including 134 with elevated expression and 106 with reduced expression in the patient group. 7 LncRNAs are involved in the determination of immune cell lineages and developmental processes, influencing both immune response modulation and the maturation process of immune cells. 8
Studies have demonstrated that NRIR serves as a promising biomarker for a range of diseases, including colorectal cancer, 9 COVID-19, 10 and juvenile myelomonocytic leukemia. 11 A study spearheaded by Mengjia Shen and her team revealed that NRIR expression is elevated in the peripheral blood mononuclear cells (PBMCs) of patients with SLE. 12 miR-31, which is expressed by CD4 Treg (T) cells and plays a pivotal role in the pathogenesis of autoimmune conditions. 13 Notably, the upregulation of miR-31 expression has been observed in T cells from mice with experimental autoimmune encephalomyelitis and in keratinocytes from individuals with psoriasis.13,14 miR-31-5p regulates T cells through several mechanisms. It was found that miR-31-5p is strongly involved in the autoimmune process in chronic inflammatory demyelinating polyneuropathy (CIDP). 15 In addition, the expression level of miR-31-5p is decreased in PBMCs from patients with SLE. 16 Intercellular adhesion molecule-1 (ICAM-1) is a membrane-bound glycoprotein that functions as a cell adhesion molecule. 17 ICAM-1 is essential for white blood cells to adhere to endothelial cells and migrate from blood vessels and is therefore involved in many autoimmune diseases. 18
The aim of this study was to explore the clinical implications of the NRIR in patients with SLE, and to examine its role in the progression of the disease.
Materials and methods
Study subjects
The study enrolled the 55 individuals diagnosed with SLE at The People’s Hospital of Feicheng. alongside an equal number of healthy controls who were undergoing routine health check-ups at the same facility. The inclusion criteria for patients with SLE adhered to the 2020 Chinese guidelines for the diagnosis and treatment of SLE. 19 Each participant was meticulously screened to ensure the absence of other autoimmune conditions, significant traumas, malignancies, psychiatric disorders, or symptoms associated with conditions other than SLE. Peripheral blood was collected from the participants and subjected to centrifugation at 4°C, 3000 r/min for a duration of 15 min. Following this, the serum was retrieved and stored at −80°C until it was needed for subsequent experiments.
This study was conducted in accordance with the principles of the Declaration of Helsinki. Approval was obtained from the Ethics Committee of The People’s Hospital of Feicheng. All participants included in this study provided their written consent.
CD4+ T cell isolation, culturing, and activation
A volume of 10 mL of peripheral blood was collected from the participants. Subsequently, PBMCs were isolated from these samples using the Ficoll-Hypaque density gradient centrifugation method, 20 followed by two rounds of washing with phosphate-buffered saline (PBS). Subsequently, 200 μL of anti-CD4 immunomagnetic bead reagent (Miltenyi Biotec, Bergisch Gladbach, Germany) was added to the PBMCs and thoroughly mixed. The mixture was then incubated at 4°C for 30 min. Following this incubation, the sample was washed twice to remove any unbound components. The cells were then resuspended in 500 μL of PBS and passed through a separation column. The isolated cells were washed twice using PBS and subsequently quantified in preparation for further analysis. Ultimately, the purity of the CD4+ T cell subset was assessed using flow cytometry. For cell culture, the cells were maintained in a human T cell growth medium (Amaxa, Shanghai, China) supplemented with 15% fetal bovine serum (FBS) and antibiotics at a concentration of 100 μg/mL (penicillin G and streptomycin). To activate the purified normal CD4+ T cells, beads coated with anti-CD3 and anti-CD28 antibodies (Dynabeads™ Human T-Activator CD3/CD28, Invitrogen, USA) were employed.
CD4+ T cell/B cell costimulation and ELISA
B cells were isolated using CD19 magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany) in accordance with the manufacturer’s recommended procedures. The transfected CD4+ T cells were co-incubated with their respective autologous B cells in 24-well round-bottom culture plates for a duration of 8 days. The concentrations of IgG present in the culture supernatant from the T cell and B cell co-cultures were quantified using ELISA kits, according to the manufacturer’s instructions (R&D Systems, USA). Additionally, levels of interleukin-4 (IL-4), interleukin-10 (IL-10), and interleukin-21 (IL-21) in the serum were quantified with ELISA kits (Rocho, Norcross, GA, USA).
Cell transfection
The shRNA-NRIR (sh-NRIR) and its negative control (sh-NC) were sourced from Transheep Bio (Shanghai, China). The short hairpin RNA-NRIR (sh-NRIR) and the corresponding negative control (sh-NC) were obtained from BeNa Bio (Beijing, China). Furthermore, the pcDNA3.1-NRIR (pcDNA-NRIR), an empty pcDNA3.1 vector used for overexpression, as well as the miR-31-5p mimics/mimics noncoding sequences and miR-31-5p inhibitor/inhibitor noncoding sequences, were purchased from BeNa Bio (Beijing, China). CD4+ T cells were transfected with the aforementioned agents via Lipofectamine 3000-mediated transfection (Thermo Fisher Scientifc, Waltham, MA, USA).
qRT-PCR
Total RNA was extracted from cells and serum using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA). The quality of the extracted RNA samples was deemed satisfactory, with A260/A280 ratios ranging from 1.8 to 2.1. Reverse transcription and subsequent quantitative analysis were performed in adherence to the manufacturer’s instructions provided with the PrimerScript RT Reagent Kit (TaKaRa, Otsu, Shiga, Japan). The expression levels were assessed utilizing the 2−ΔΔCt method. The specific primers utilized for qRT-PCR were as follows: NRIR forward: 5′-AGCGATTATCCTGCCTCAGC-3’. NRIR reverse: 5′-TCCAGAGGCAGATGATGGGA-3’. miR-31-5p forward: 5′-TCGGCAGGAGGCAAGAUGCUG-3’. miR-31-5p reverse: 5′-CTCAACTGGTGTCGTGGA-3’. ICAM-1 forward: 5′-TCTTCCTCGGCCTTCCCATA-3’. ICAM-1 reverse: 5′-ATTAGGACTGGGAGCTCCGT-3’.
CCK-8 assay
The transfected CD4+ T cells were treated with a solution of CCK-8 (Procell Life Science & Technology, Wuhan, China) at a volume of 10 μL per well. Following this, the cells were then incubated at a temperature of 37°C for a duration of 2 h. Subsequently, the absorbance at a wavelength of 450 nm was determined using a microplate spectrophotometer (Thermo Fisher Scientifc, Waltham, MA, USA).
Dual-luciferase reporter gene assay
Subsequent to the insertion of the anticipated binding site and mutated fragments of NRIR, along with miR-31-5p into dual-luciferase reporter vectors, these constructs were designated as PGLO-NRIR-wild type (WT) and PGLO-NRIR-mutant type (MUT). Subsequently, these plasmids were co-transfected into CD4+ T cells using either miR-31-5p mimic or miR-31-5p inhibitor. After a 24-h transfection period, the cells were centrifugated at 12,000 r/min. Luciferase activities were quantified using the Dual-Luciferase Reporter System (Dual-Luciferase® Reporter Assay System, Promega, Madison, WI, USA).
Statistical analysis
In this study, GraphPad Prism nine was employed for data handling and statistical analysis. The data were presented as mean ± standard deviation (SD) and the differences between groups were determined by independent sample t test or one-way ANOVA. ROC curve was performed to assess the diagnostic potential of NRIR in SLE. Correlation assessments were conducted utilizing the Pearson correlation coefficient. A significant disparity was deemed present if the p-value was less than 0.05.
Result
Basic patient information
Baseline characteristics of patients with SLE and healthy controls.
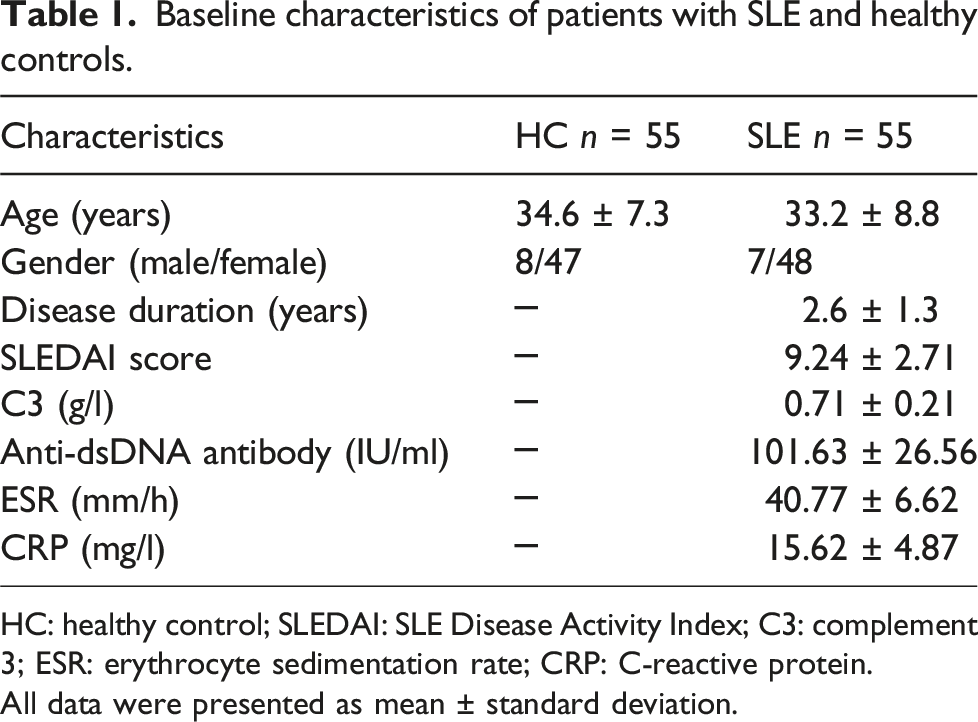
HC: healthy control; SLEDAI: SLE Disease Activity Index; C3: complement 3; ESR: erythrocyte sedimentation rate; CRP: C-reactive protein.
All data were presented as mean ± standard deviation.
Expression and diagnostic value of NRIR in SLE
In comparison to the healthy individuals, the level of NRIR in the serum was markedly elevated in patients with SLE (Figure 1(a)). The ROC curve analysis indicated that the ROC curve of NRIR in diagnosing SLE yielded an AUC of 0.887, the sensitivity of 83.6% and the specificity of 92.7% (Figure 1(b)). Expression and diagnostic role of NRIR in serum of SLE patients. (a) The expression of NRIR in serum of SLE patients was detected by qRT-PCR. (B) ROC curve b NRIR for the diagnosis of SLE. ***p < 0.001.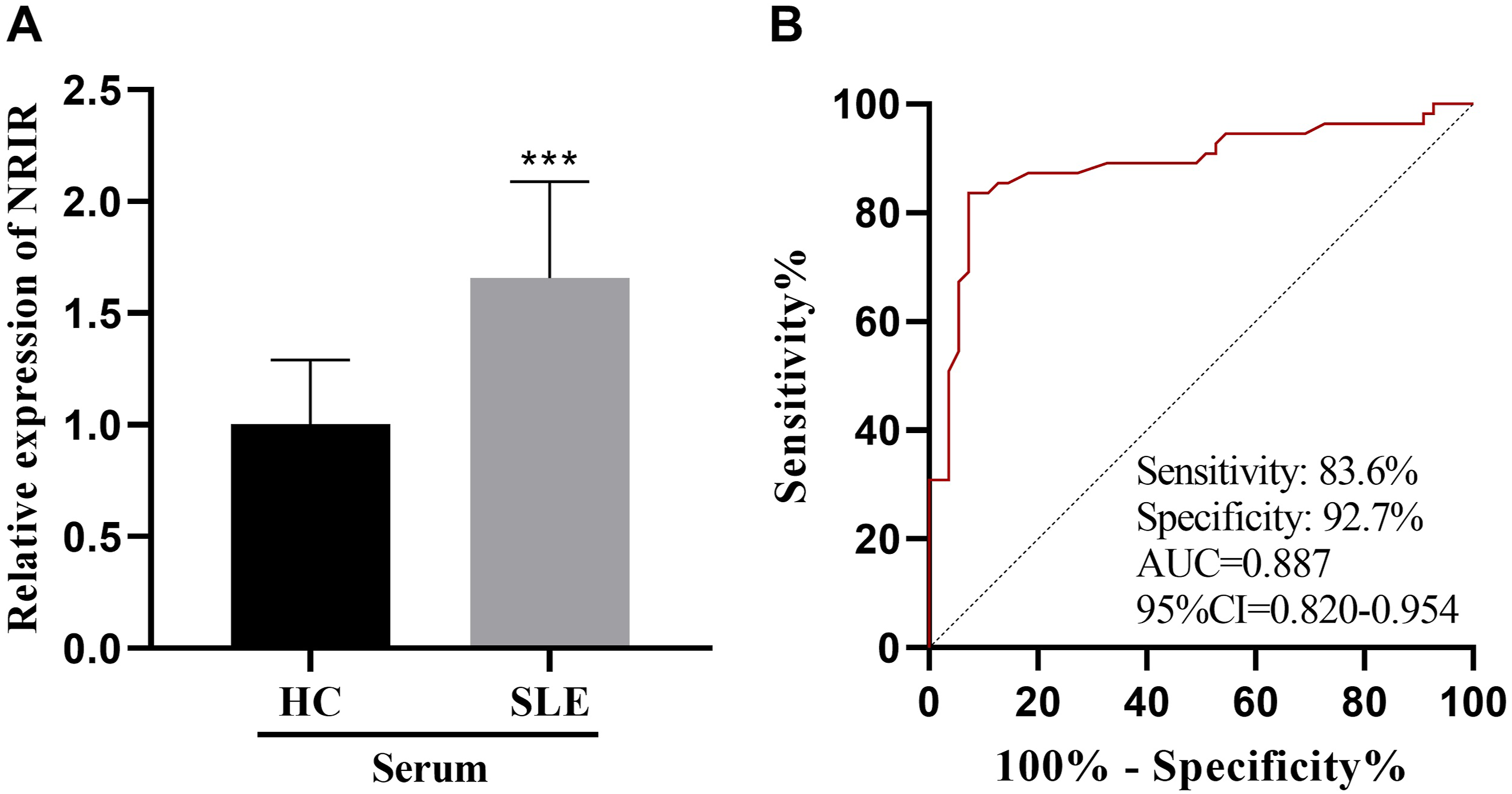
Correlation between serum NRIR expression and clinical indicators in SLE patients
The serum level of NRIR in the patients with SLE exhibited a significant positive correlation with the SLEDAI score (r = 0.7302, p < 0.001), erythrocyte sedimentation rate (ESR) (r = 0.5513, p < 0.001), and anti-dsDNA antibodies (r = 0.6761, p < 0.001) (Figure 2(a)–2(c)). Conversely, NRIR levels demonstrated a negative correlation with complement C3 levels (r = −0.6552, p < 0.001) (Figure 2(d)). Furthermore, no significant correlation with the levels was observed between NRIR expression and C-reactive protein (CRP) levels(p > 0.05) (Figure 2(e)). Correlation between NRIR and clinical indicators in SLE patients. The association of NRIR with (a) SLEDAI score, (b) ESR, (c) anti-dsDNA antibody, (d) complement C3, and (e) CRP in SLE patients was analyzed by Pearson analysis.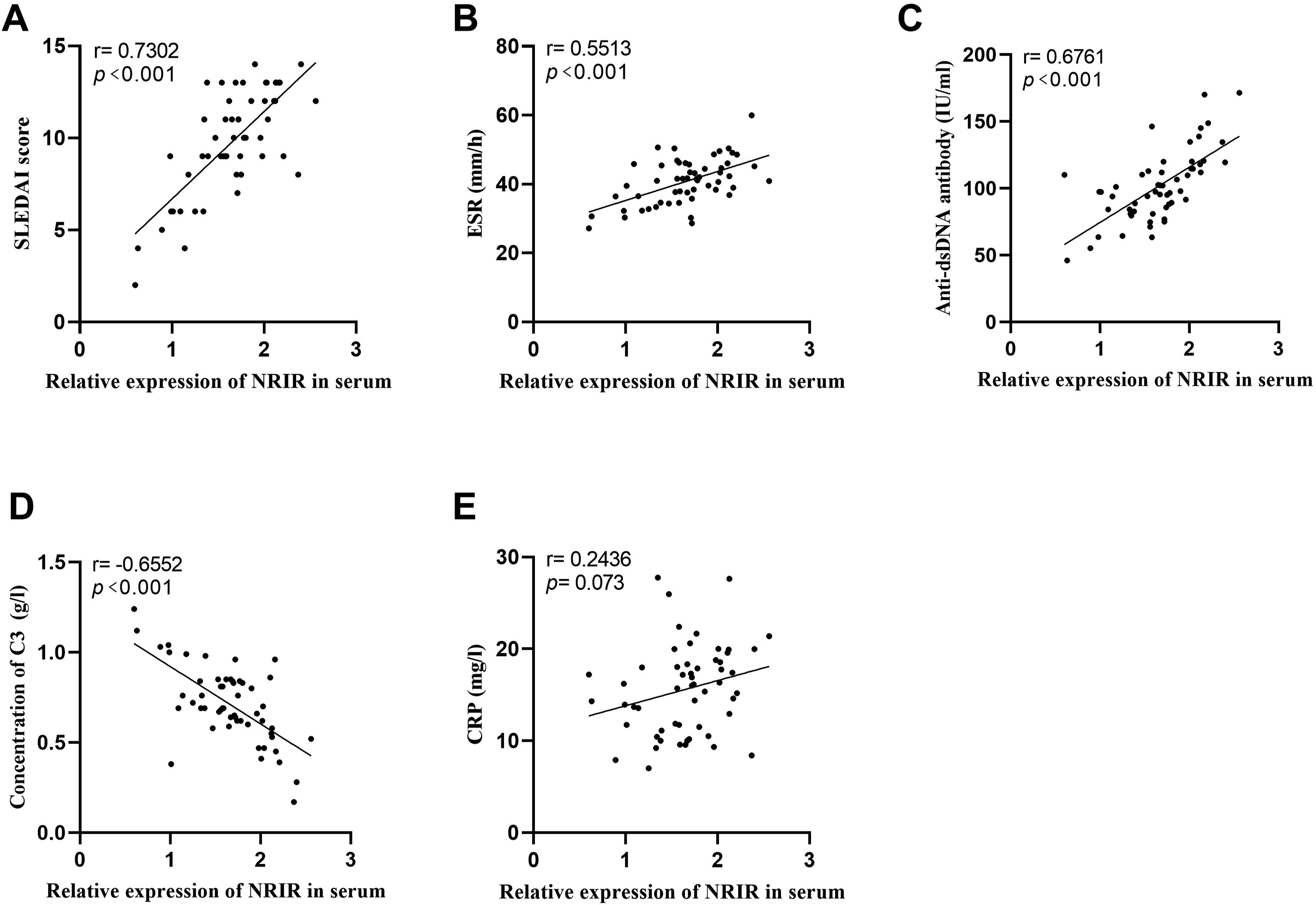
Serum secretion of inflammatory cytokines and their correlation with NRIR in SLE patients
The levels of IL-4, IL-6, and IL-21 in the serum of patients were assessed using ELISA kits. The findings revealed that the cytokine levels were significantly higher in the SLE patient compared to the healthy controls (Figure 3(a)–3(c)). Additionally, the elevations in these cytokine levels displayed a positive correlation with the expression of NRIR (Figure 3(d)–3(f)). Levels of IL-4, IL-6, IL-21, and their correlation with NRIR. (a-c) Serum levels of IL-4, IL-6 and IL-21 were detected by ELISA. (d-f) IL-4, IL-6 and IL-21 were positively correlated with the level of NRIR. ***p < 0.001.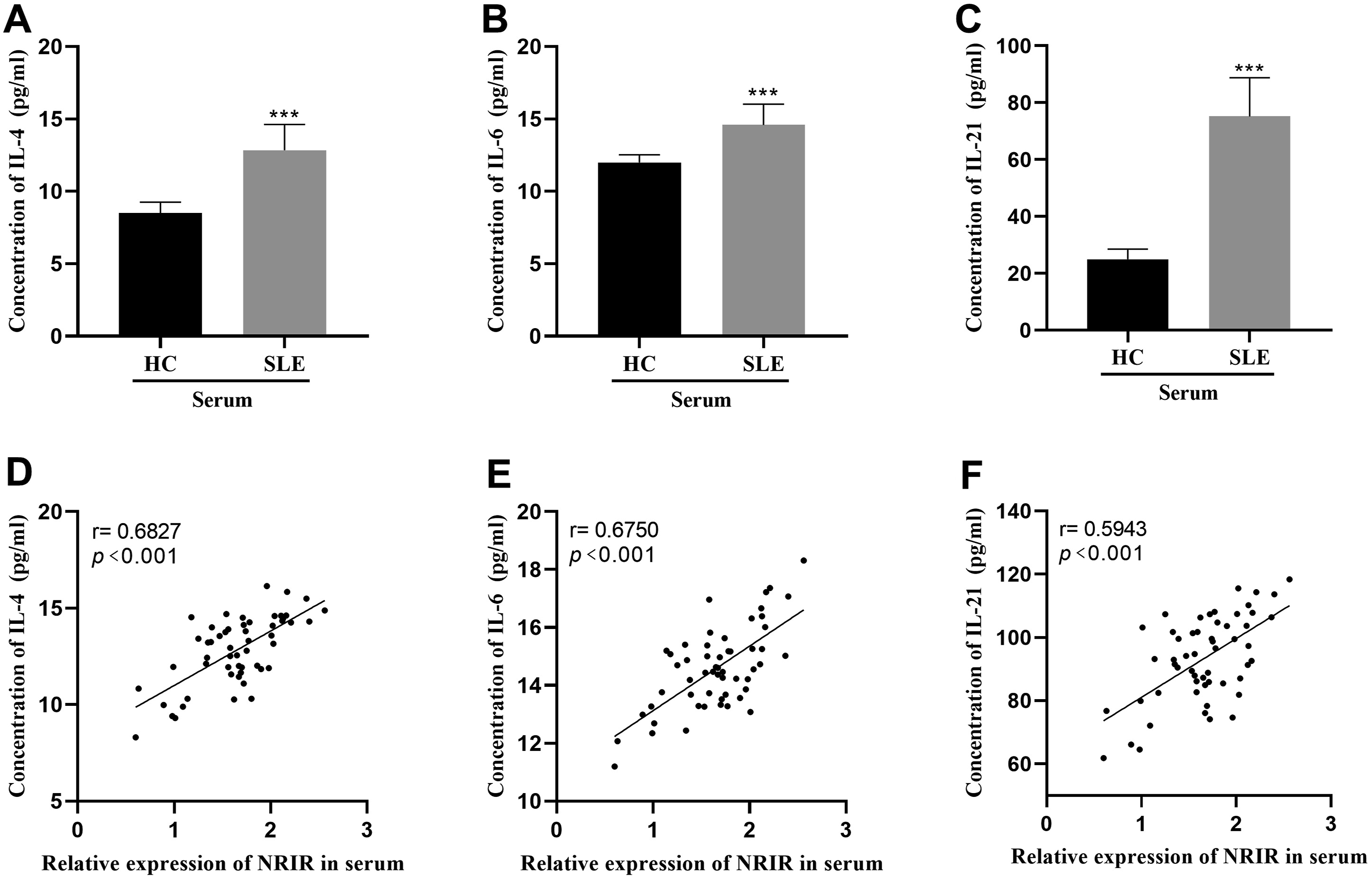
Effect of abnormal expression of NRIR on autoimmune response
The result showed that NRIR was highly expressed in CD4+ T cells of SLE patients compared with healthy controls (Figure 4(a)). To explore the impact of this abnormal NRIR expression on autoimmune responses, we manipulated NRIR levels in CD4 + T cells from SLE patients by either upregulating or downregulating NRIR levels of CD4+ T cells of SLE patients (Figure 4(b)). We evaluated cell viability and found that overexpression of NRIR resulted in increased viability of CD4 + T cells, while the inhibition of NRIR expression led to reduced cell viability (Figure 4(c)). Furthermore, NRIR overexpression was associated with elevated levels of IL-4, IL-6, and IL-21 secreted by CD4+ T cells, whereas the suppression of NRIR expression led to a reduction in the levels of these cytokines (Figure 4(d)–4(f)). Increased NRIR expression was linked to higher ICAM-1 expression and enhanced IgG production of IgG by B cells. Conversely, silencing of NRIR resulted in reduced ICAM-1 expression and diminished IgG production by B cells (Figure 4(g) and 4(h)). Effect of abnormally expressed NRIR on CD4+ T cells. (a) The level of NRIR in CD4+ T cells was detected by qRT-PCR. (b) The transfection efficiency of CD4+ T cells was detected by qRT-PCR. (c) The viability of CD4+ T cells was detected by CCK-8 assay. (d-f) The levels of IL-4, IL-6 and IL-21 in CD4+ T cells of SLE patients were detected by ELISA. (g) ICAM-1 levels in CD4+ T cells of SLE patients were detected by qRT-PCR. (h) The IgG concentration was detected by ELISA. ***p < 0.001, **p < 0.01, *p < 0.05.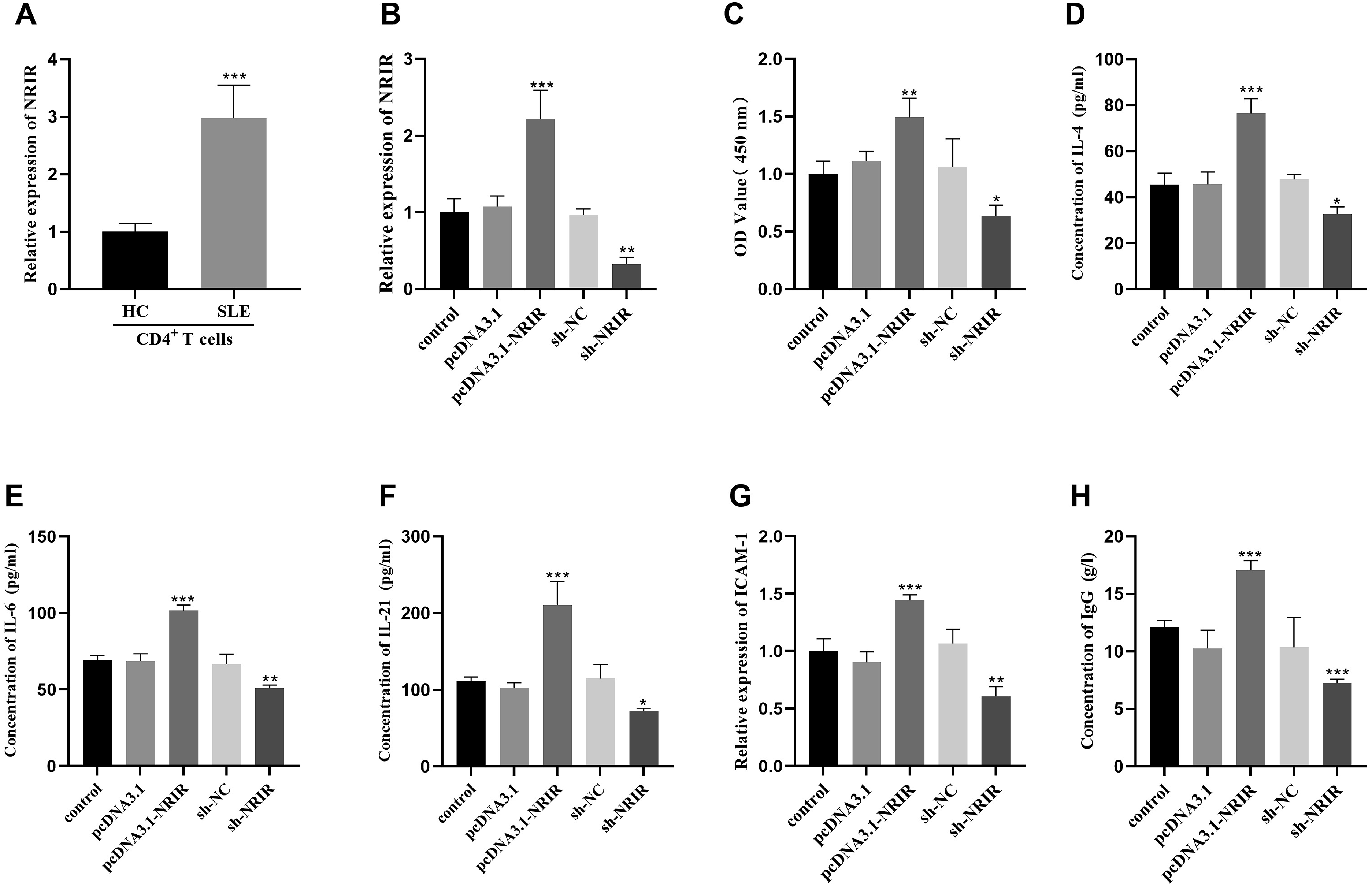
NRIR targets miR-31-5p and negatively regulates miR-31-5p
Reduced expression of miR-31-5p was observed in the serum of individuals with SLE, and its levels showed an inverse relationship with the expression of NRIR (Figure 5(a) and 5(b)). According to the bioinformatics prediction platform lncRNASNP2 (https://guolab.wchscu.cn/lncRNASNP/#!/), a potential interaction was predicted between NRIR and miR-31-5p, as illustrated in Figure 5(c). The dual-luciferase reporter gene assay revealed a significant increase in luciferase activity in CD4+ T cells co-transfected with NRIR-WT and miR-31-5p inhibitors. whereas a substantial decrease in luciferase activity was observed in cells treated with NRIR-WT and miR-31-5p mimics, (Figure 5(d)). After overexpression of NRIR, miR-31-5p levels within the cells were notably reduced. In contrast, NRIR suppression led to an increase in the miR-31-5p expression (Figure 5(e)). NRIR negatively regulated miR-31-5p. (a) The expression of miR-31-5p in serum of SLE patients was detected by qRT-PCR. (b) NRIR was negatively correlated with the expression of miR-31-5p. (c) The binding site sequence between NRIR and miR-31-5p was forecasted utilizing the lncRNASNP2 database. (d) The interaction between NRIR and miR-31-5p was verified by dual luciferase reporter gene assay. (e) After overexpression or silence of NRIR, the expression level of miR-31-5p was detected by qRT-PCR. ***p < 0.001, **p < 0.01.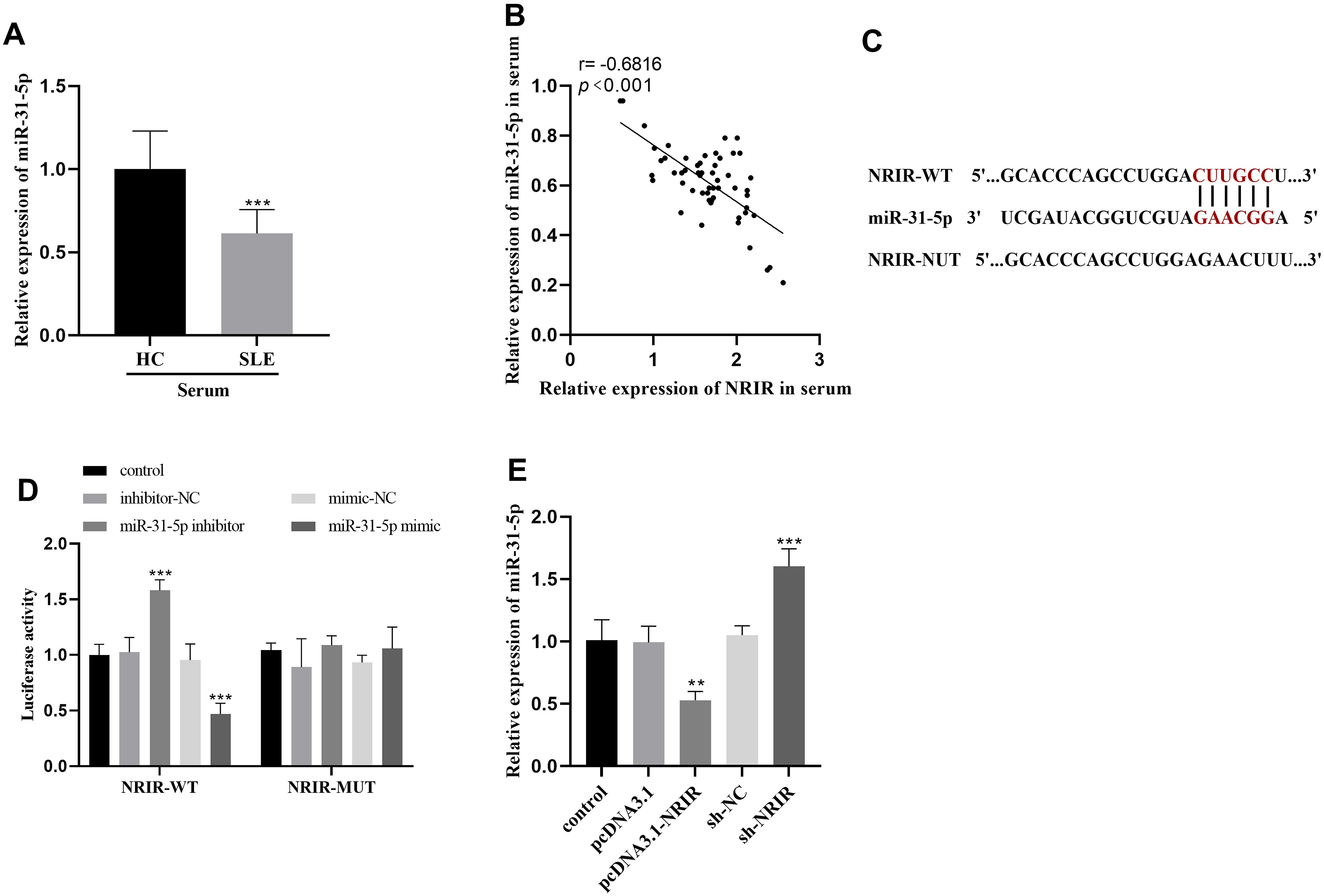
Discussion
In SLE, the immune system’s response is characterized by an excessive activation of both B and T cells, accompanied by a breakdown in the body’s natural self-tolerance. This dysregulation leads to the production of aberrant antibodies, the circulation and accumulation of immune complexes within tissues, and the activation of the complement system and cytokines, altogether contributing to the varied clinical presentations, which may span from trivial symptoms like tiredness and muscle discomfort to severe organ damage posing a significant threat to life. 1
The study conducted by Han Wu and colleagues demonstrated that through transcriptome sequencing, a significant number of novel lncRNAs were identified in PBMCs, serum, and exosomes derived from patients and animal models suffering from SLE. 21 LncRNAs display remarkable stability within serum and plasma matrices, suggesting their innovative, non-invasive biomarkers for the diagnosis and ongoing monitoring of SLE. 22
This investigation revealed that NRIR is significantly overexpressed in patients with SLE, aligning with prior research. 12 In addition, NRIR served as a discriminative marker between healthy individuals and those with SLE. Furthermore, NRIR expression displayed a notable association with several clinical parameters in SLE patients, including the SLEDAI score, ESR, Anti-dsDNA antibody levels, and complement C3 levels. This suggested that NRIR may be closely related to the disease status of SLE. In addition to its abnormal expression in SLE, 12 NRIR has been shown to play a new role in other autoimmune diseases. For example, Systemic sclerosis (SSc), also known as scleroderma, is a long-lasting autoimmune disorder. In SSc patients, NRIR is thought to regulate the interferon (IFN) response in monocytes. 23 NRIR is reported to be a key regulator of the IFN-mediated process and is differentially expressed in lupus nephritis. 24
IL-4 is produced by both Th2 cells and natural killer cells, playing a crucial role in driving inflammatory processes. 25 Elevated levels of IL-21 are commonly observed in various chronic inflammatory diseases, including SLE. 26 IL-21 is primarily secreted by activated CD4+ T lymphocytes and regulates the maturation and functional responses of effector T helper lineages, mitigates the inhibitory influence of regulatory T cells, and enhances the production of pro-inflammatory factors by non-immune cells. 27 This study further elucidated that the serum concentrations of IL-4, IL-6, and IL-21 in SLE patients are elevated in patients with SLE compared to healthy individuals. Moreover, these elevated levels were positively correlated with the expression of NRIR, this may be due to the fact that NRIR promoted the secretion of inflammatory factors and induced an inflammatory response.
Numerous investigations have revealed significant elevated levels of ICAM-1 in patients suffering from SLE.28,29 Previous investigations have demonstrated that increased ICAM-1 levels in hematuria serve as a biomarker for SLE. 18 Furthermore, the measurement of soluble ICAM-1 (sICAM-1) concentrations could facilitate the identification of patients who are at risk of developing lupus nephritis. 29 In this study, after the overexpression of NRIR, the activity of CD4+ T cells was enhanced. When these T cells were co-cultured with B cells, there was a notable increase in ICAM-1 expression, which stimulated B cells to produce a substantial quantity of antibodies. Additionally, the secretion of inflammatory factors was increased, which accelerated the progression of SLE. On the contrary, the inhibition of NRIR could inhibit the occurrence of autoimmunity. These findings suggested that the abnormal expression of NRIR in CD4+ T cells among SLE patients may be an important molecular mechanism leading to the abnormal activation of CD4+ T cells and autoimmune response in SLE.
miR-31-5p is abnormally expressed not only in SLE, but also in rheumatoid arthritis. 30 Moreover, the reduction of miR-31-5p in T cells from patients suffering from Sjögren’s syndrome (SjS) may contribute to the activation of autoimmune T-cell responses under chronic type I IFN exposure. 31 Our study observed a down-regulated expression of miR-31-5p in SLE patients, which was highly consistent with previous studies. 16 Notably, we found that the expression of miR-31-5p was significantly negatively correlated with NRIR levels. The modulated relations between these molecules was confirmed through a dual-luciferase reporter assay. Overexpression of NRIR corresponded with a decrease in miR-31-5p levels, and conversely, reduced NRIR expression was associated with increased miR-31-5p levels, thereby reinforcing the antagonistic relationship between them. This may indicate that NRIR mediated the progression of SLE by targeting and regulating the expression of miR-31-5p. Of course, more experimental data were needed to confirm this conclusion.
Overall, our findings revealed that NRIR expression was elevated in patients suffering from SLE, demonstrating considerable potential as a diagnostic marker for distinguishing SLE cases. In addition, NRIR may exert a negative regulatory effect on miR-31-5p expression, although the precise mechanism requires further investigation.
Conclusion
NRIR emerges as a potential biomarker for SLE and is implicated in the progression of SLE.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.