
Abstract
Background
Systemic lupus erythematosus (SLE) is a chronic autoimmune disorder characterized by the production of autoantibodies that target most of the organ systems and lead to their dysfunction. The exact etiology of SLE remains unclear; however, genetic and environmental factors are believed to play significant roles. Viral infections, particularly Epstein-Barr virus (EBV), have been implicated as environmental triggers in SLE pathogenesis however, the observations remained inconsistent among studies and populations. The present study uses a meta-analysis approach to explore the prevalence of EBV infection in the general population and their role in the pathogenesis of SLE.
Materials and Methods
Various databases such as PubMed, Scopus, and ScienceDirect were searched to obtain eligible studies based on predetermined inclusion and exclusion criteria. The Newcastle-Ottawa Scale (NOS) was used for quality assessment of the eligible studies, and Comprehensive Meta-Analysis (CMA) v4 software was used for the analysis. Publication bias was assessed with funnel plots and Egger’s regression, while heterogeneity was evaluated with Cochrane Q and I2 statistics.
Results
In the present investigation, a total of 28 studies comprising of 3926 healthy controls and 2968 SLE patients were included. EBV infections were prevalent in the healthy controls. While comparing the frequency of EBV DNA or antibodies positivity, the SLE patients had a higher positivity rate than the healthy controls, indicating that EBV infection is a risk factor for developing SLE. Furthermore, the sensitivity analysis also revealed that the meta-analysis was robust.
Conclusion
The majority of healthy subjects were previously exposed to EBV, and the infection could be a potential risk factor in SLE pathogenesis. However, future research is required to elucidate the possible mechanisms of EBV reactivation in SLE patients and examine potential preventive measures, such as antiviral therapies, in mitigating SLE risk.
Introduction
Systemic lupus erythematosus (SLE) is a chronic autoimmune disease characterized by dysfunctional B cell activation, altered T cell function, and elevated autoantibodies, which can affect a wide range of organs and tissue systems in the body. The exact cause of the SLE pathogenesis is still unknown. 1 However, the complex interaction between human genetic factors and environmental influences has been attributed to the development of SLE. Among the environmental factors, infections by viruses and bacteria have been linked with the onset of the disease.
Viral infections are documented as significant environmental triggers for SLE’s priming, pathogenesis, and clinical severity. Viruses such as Epstein-Barr virus (EBV), cytomegalovirus (CMV), and parvovirus B19 are frequently connected with the pathogenesis of SLE. Although the exact mechanism by which these viral infections lead to the development of SLE in a subset of infected individuals is not fully understood, it is believed that viral infections tempt immune dysregulation through molecular mimicry, bystander activation, and epitope spreading. 2 EBV infects B cells and remains inside the host cells throughout the entire lifetime of humans. 3 The EBV nuclear antigen-1 (EBNA-1) shares similarities with proteins responsible for triggering autoimmune responses and producing autoantibodies in SLE patients.4,5 Additionally, 6 documented in silico that nearly half of the SLE risk loci are linked with EBNA-2, further supporting the potential involvement of EBV in the pathogenesis of SLE.
A possible link between EBV infections and SLE pathogenesis was reported about 50 years ago. 7 However, the observations remained inconsistent among the studies and populations. Systematic reviews8,9 comprising earlier published articles documented that SLE patients had a higher prevalence of viral capsid antigen (VCA) IgG but not EBNA-1 when compared with those of control groups. In the general population, the prevalence of EBV remains widespread; thus, investigating the importance of EBV infections on the pathogenesis of SLE remained underpowered to draw a concrete conclusion. 10 Thus, there is a need for a well-designed investigation with a larger sample size in different populations, exploring both serological and DNA-based evidence to better understand this connection. Meta-analysis is a robust statistical analysis that combines similar types of investigations and draws a definitive conclusion. In the present systematic review and meta-analysis, we aim to explore the association between EBV infection and SLE pathogenesis by pooling the earlier published evidence using a meta-analysis approach.
Materials and methods
Literature search for meta-analysis
The present meta-analysis followed the PRISMA guidelines. A comprehensive literature search was conducted by two independent authors, SR and AKP, across various databases, including Scopus, PubMed, and ScienceDirect, to compile relevant published studies. The search employed a combination of keywords, including “Epstein-Barr Virus,” “EBV,” “systemic lupus erythematosus,” and “SLE,” utilizing Boolean operators for optimized retrieval. The last database search was performed on October 1, 2024.
Inclusion and exclusion criteria for the current meta-analysis. To ensure methodological rigor, predefined inclusion and exclusion criteria were established prior to the meta-analysis. The inclusion criteria were as follows: (1) Studies must include patients diagnosed with SLE according to the American College of Rheumatology (ACR) criteria; (2) Patients could be of any age group (excluding paediatric populations); (3) Studies must assess the EBV DNA genome using peripheral blood mononuclear cells (PBMCs) and/or serum, or detect antibodies against at least one of the following EBV antigens: VCA, EA E/D, EBNA-1, or EBNA-2; (4) the number of SLE patients and healthy controls must be explicitly stated. The exclusion criteria included the following considerations: (1) reviews and case reports.; (2) studies lacking data on either SLE patients or healthy controls; (3) studies with insufficient data for meta-analysis; (4) non-human (animal) studies; (5) reports on SLE patients with pregnancy, infection, juvenile SLE, or other autoimmune diseases; (6) studies reporting only serum antibodies without specifying the antigen or only an antigen without specifying the antibody type.
Extraction of data
Two authors (SR and AKP) independently extracted data from all eligible studies utilizing a pre-designed, standardized data extraction Microsoft Excel form. The form, developed in prior to the review, encompassed fields for the first author’s name, publication year, country of study, sample sizes for both SLE patients and healthy controls, age distributions, detection methods, sample types, matching criteria, ACR classification criteria, and EBV prevalence rates in both groups. To ensure consistency and minimize bias, both reviewers completed the extraction process independently, and any discrepancies were resolved through discussion with all authors until consensus was reached. This systematic and standardized approach ensured the accuracy and reliability of the extracted data for subsequent analysis.
Quality assessment
The methodological quality of the included studies was assessed using the Newcastle-Ottawa Scale (NOS). 11 Two authors independently evaluated each study and assigned scores based on NOS criteria, which assess three main domains: Selection (maximum four stars), Comparability (maximum two stars), and Exposure/Outcome (maximum three stars), with a total possible score of nine stars. Studies scoring 7–9 were considered high quality, 5–6 as moderate quality, and 0–4 as low quality, although no official threshold for quality classification exists. Any disagreements were resolved by consensus.
Statistical analysis
Meta-analysis was conducted using Comprehensive Meta-Analysis (CMA) v4 software. Publication bias was assessed through funnel plot analysis and Egger’s regression test. 12 Heterogeneity among the included studies was evaluated using Cochrane’s Q test, probability heterogeneity (P-value), and I2 statistics. 13 A P-value <.05 and an I2 value >50% indicated substantial heterogeneity, warranting the use of a random-effects model for meta-analysis. Given that the articles included in this meta-analysis examined diverse populations, a random effects model was employed to investigate both prevalence and association. 14 The prevalence of EBV infection among healthy controls across various populations was assessed by calculating pooled event rates and 95% confidence intervals. To evaluate the association between EBV infection and susceptibility to the development of SLE, pooled odds ratios, 95% confidence intervals, z-values, and p-values were determined. A p-value of less than 0.05 was considered statistically significant. Sensitivity analysis was performed by systematically excluding individual studies to assess the robustness of the meta-analysis results.
Results
Literature search
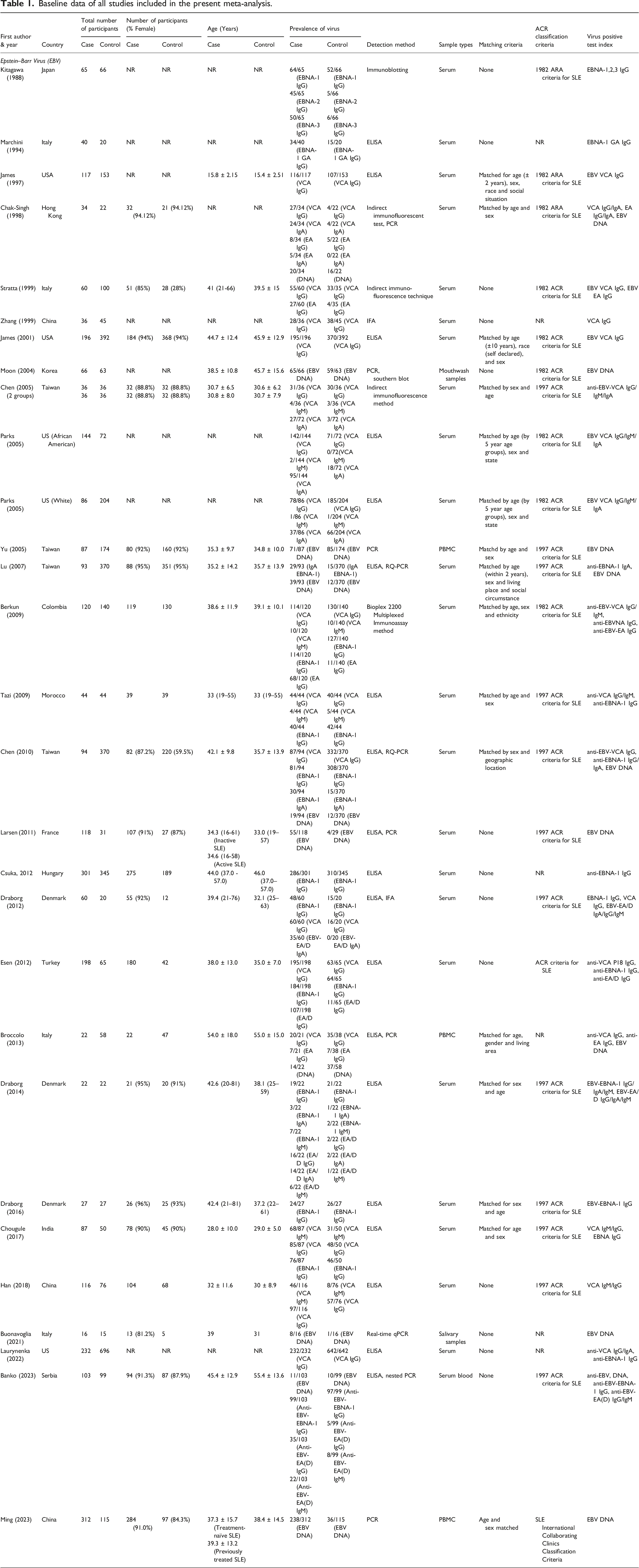
For this meta-analysis examining the association between EBV and SLE, we conducted a comprehensive search of three distinct online databases. In accordance with the PRISMA 2020 guidelines, we present a flowchart (Figure 1) delineating our literature search and selection process for eligible studies. Out of the 2551 studies initially identified from three distinct databases, 247 were excluded due to duplication, and 2304 were considered for screening (details are provided in the supplementary file). Ultimately, only 28 studies satisfied our rigorous inclusion and exclusion criteria.4,15–41 All included studies attained five or more stars in NOS analysis (Supplemental Figure 1). The baseline data of all studies incorporated in the present meta-analysis are presented in Table 1. PRISMA 2020 study flow chart showing the number of included and excluded articles for the present meta-analysis. Baseline data of all studies included in the present meta-analysis.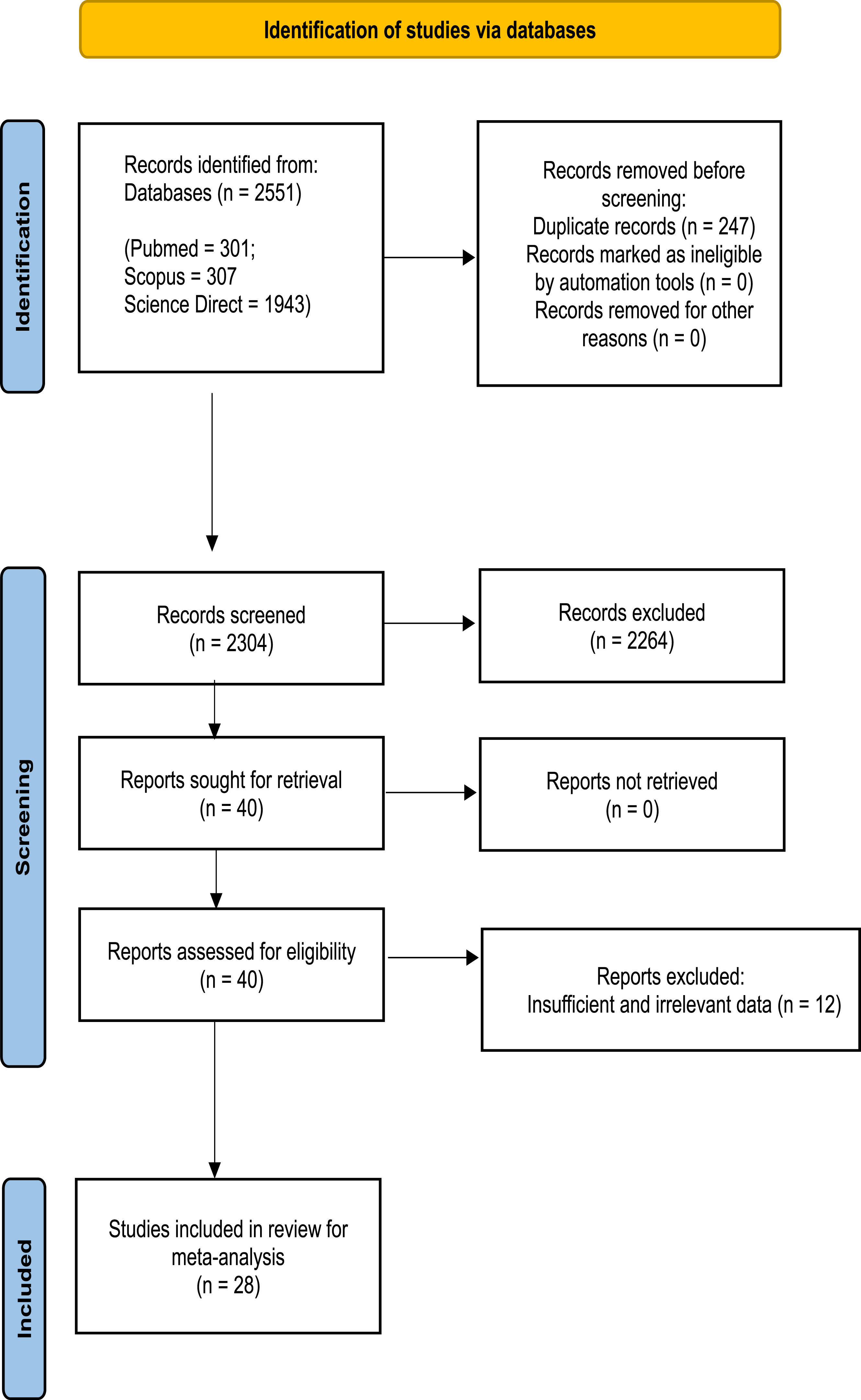
Study characteristics
The meta-analysis included 28 case–control studies published between 1988 and 2023, spanning various countries, including Japan, Italy, USA, China, South Korea, Taiwan, Morocco, France, Denmark, Turkey, India, Philippines, and Serbia. These studies investigated the association between EBV infections and SLE using diverse methods to detect EBV infection or immune responses to EBV antigens. Detection methods included antibody assays against EBV nuclear antigens, capsid antigens, and early antigens; measurement of EBV DNA in peripheral blood mononuclear cells; and assessment of T-cell and cytokine responses to EBV antigens. Sample sizes varied across studies, and the reports highlighted associations between EBV and SLE, with findings ranging from increased prevalence of EBV infection in SLE patients to abnormally elevated EBV loads in blood, impaired immune responses to EBV antigens, and potential correlations with disease activity and clinical presentation.
Publication bias
Publication bias and heterogeneity analysis of serological prevalence of EBV and its association with SLE.
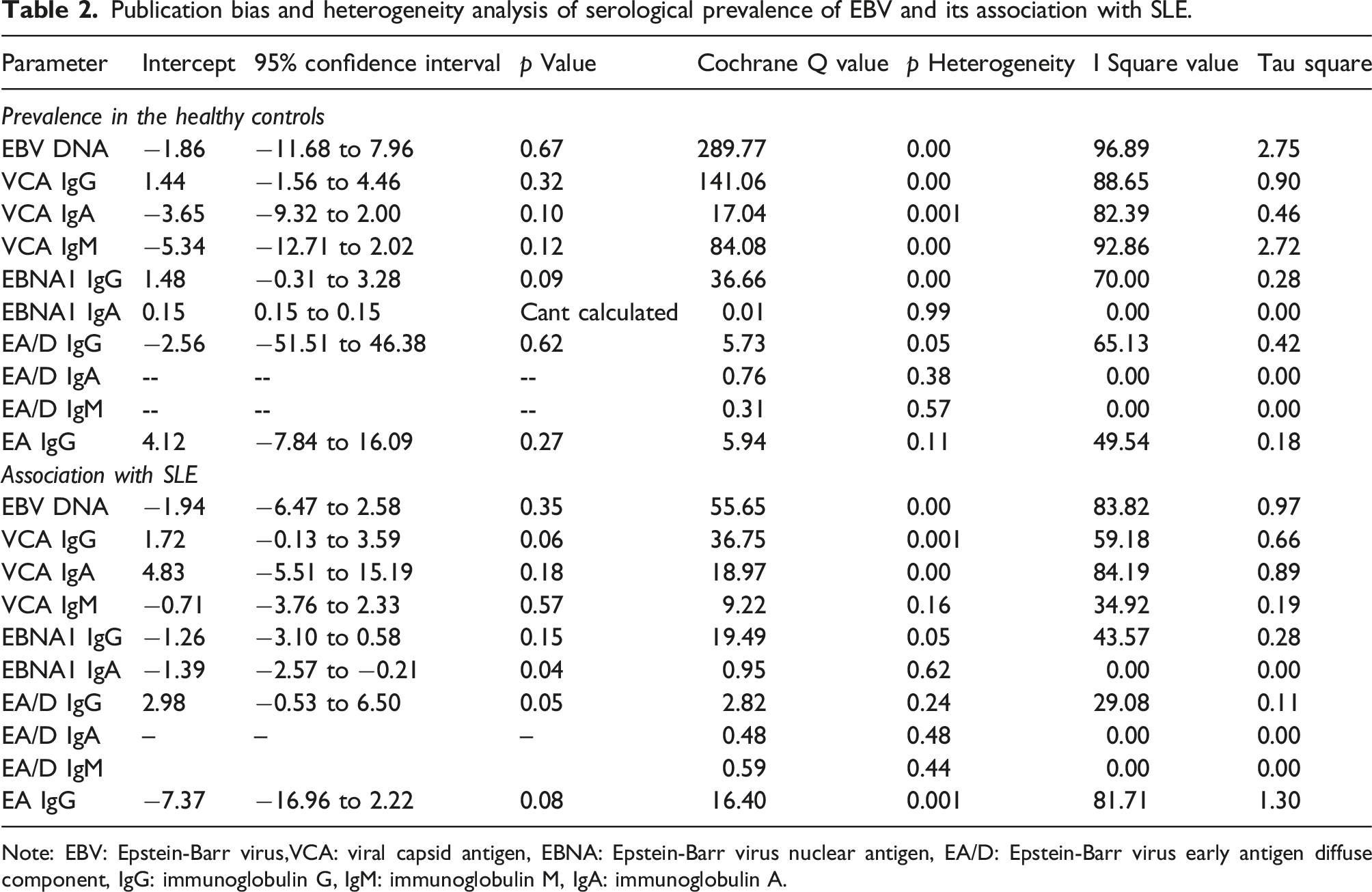
Note: EBV: Epstein-Barr virus,VCA: viral capsid antigen, EBNA: Epstein-Barr virus nuclear antigen, EA/D: Epstein-Barr virus early antigen diffuse component, IgG: immunoglobulin G, IgM: immunoglobulin M, IgA: immunoglobulin A.
Heterogeneity analysis
The heterogeneity in the included studies was assessed utilizing the Cochrane Q, I2, and Tau square tests for the prevalence and association of EBV DNA and EBV VCA, EBNA1, EA/D, IgG, IgA, and IgM in SLE patients. Table 2 presents the results of the heterogeneity tests. The heterogeneity for the prevalence of EBV DNA (Q = 289.77; Phet = 0.00; I2 = 96.89; Tau sqr = 2.75), EBV VCA (IgG: Q = 141.06; Phet = 0.00; I2 = 88.65; Tau sqr = 0.90, IgM: Q = 84.08; Phet = 0.00; I2 = 92.86; Tau sqr = 2.72, IgA: Q = 17.04; Phet = 0.001; I2 = 82.39; Tau sqr = 0.46) and EBNA1 IgG (Q = 36.66; Phet = 0.00; I2 = 70.00; Tau sqr = 0.28) and EA/D IgG (Q = 5.73; Phet = 0.05; I2 = 65.13; Tau sqr = 0.42) demonstrates significant heterogeneity (Table 2). Conversely, EBNA1 IgA (Q = 0.01; Phet = 0.99; I2 = 0.00; Tau sqr = 0.00), EA IgG (Q = 5.94; Phet = 0.11; I2 = 49.54; Tau sqr = 0.18), and EA/D IgA (Q = 0.76; Phet = 0.38; I2 = 0.00; Tau sqr = 0.00) exhibited an absence of heterogeneity. Similarly, for association studies, EBV DNA (Q = 55.65; Phet = 0.00; I2 = 83.82; Tau sqr = 0.97), VCA (IgG: Q = 36.75; Phet = 0.001; I2 = 59.18; Tau sqr = 0.66, and IgA: Q = 18.97; Phet = 0.00; I2 = 84.19; Tau sqr = 0.89), EBNA1 IgG (Q = 19.49; Phet = 0.05; I2 = 43.57; Tau sqr = 0.28), and EA IgG (Q = 16.40; Phet = 0.001; I2 = 81.71; Tau sqr = 1.30) displayed heterogeneity, while other comparison parameters were documented as homogeneous [VCA IgM (Q = 9.22; Phet = 0.16; I2 = 34.92; Tau sqr = 0.19), EBNA1 IgA (Q = 0.95; Phet = 0.62; I2 = 0.00; Tau sqr = 0.00), EA/D IgG (Q = 2.82; Phet = 0.24; I2 = 29.08; Tau sqr = 0.11), EA/D IgA (Q = 0.48; Phet = 0.48; I2 = 0.00; Tau sqr = 0.00), and EA/D IgM (Q = 0.59; Phet = 0.44; I2 = 0.00; Tau sqr = 0.00)] (Table 2). For all analyses, the random effect model was employed.
Prevalence of EBV DNA and immunoglobulins in healthy subjects
To investigate the prevalence of EBV DNA and immunoglobulins, we analyzed data from 28 studies, consisting of 3926 healthy controls, using the CMA v4 software. The prevalence of EBV DNA in the healthy controls was 27.1% (95% CI, 11.4–51.9%). The seroprevalences of EBV VCA IgG, IgA, and IgM antibodies in healthy controls were 89.3% (95% CI, 83.2–93.3%), 18.9% (95% CI, 9.7–33.6%), and 7.8% (95% CI, 2.2–24.1%), respectively. In contrast, the seroprevalence of EBV EBNA1 IgG and IgA antibodies in healthy controls was 89.3% (95% CI, 84.7–92.7%) and 4.1% (95% CI, 2.9–5.7%), respectively. Moreover, the seroprevalence of EBV EA/D IgG, IgA, and IgM antibodies in healthy controls was 9.7% (95% CI, 4.1–21.4%), 6.9% (95% CI, 2.0–21.2%), and 7.6% (95% CI, 4.0–13.9%), respectively. Approximately 13.4% (95% CI, 7.9–21.9%) of healthy subjects were seropositive for EBV EA IgG (Figure 2). Forest plots for the Prevalence of EBV DNA and antibodies in healthy controls. A meta-analysis was conducted to ascertain the prevalence of Epstein-Barr Virus (EBV) DNA and antibodies against EBV within a population of healthy controls. This analysis was executed using CMA v4 software. Forest plots illustrate the pooled event rates, 95% confidence intervals (CI), and the number of positive cases relative to the total number of samples included in the prevalence study. The specific components analyzed include: (a) EBV DNA, (b) Viral Capsid Antigen (VCA) IgG, (c) VCA IgA, (d) VCA IgM, (e) Epstein-Barr Nuclear Antigen 1 (EBNA1) IgG, (f) EBNA1 IgA, (g) Early Antigen/Diffuse (EA/D) IgG, (h) EA/D IgA, (i) EA/D IgM, and (j) Early Antigen (EA) IgG.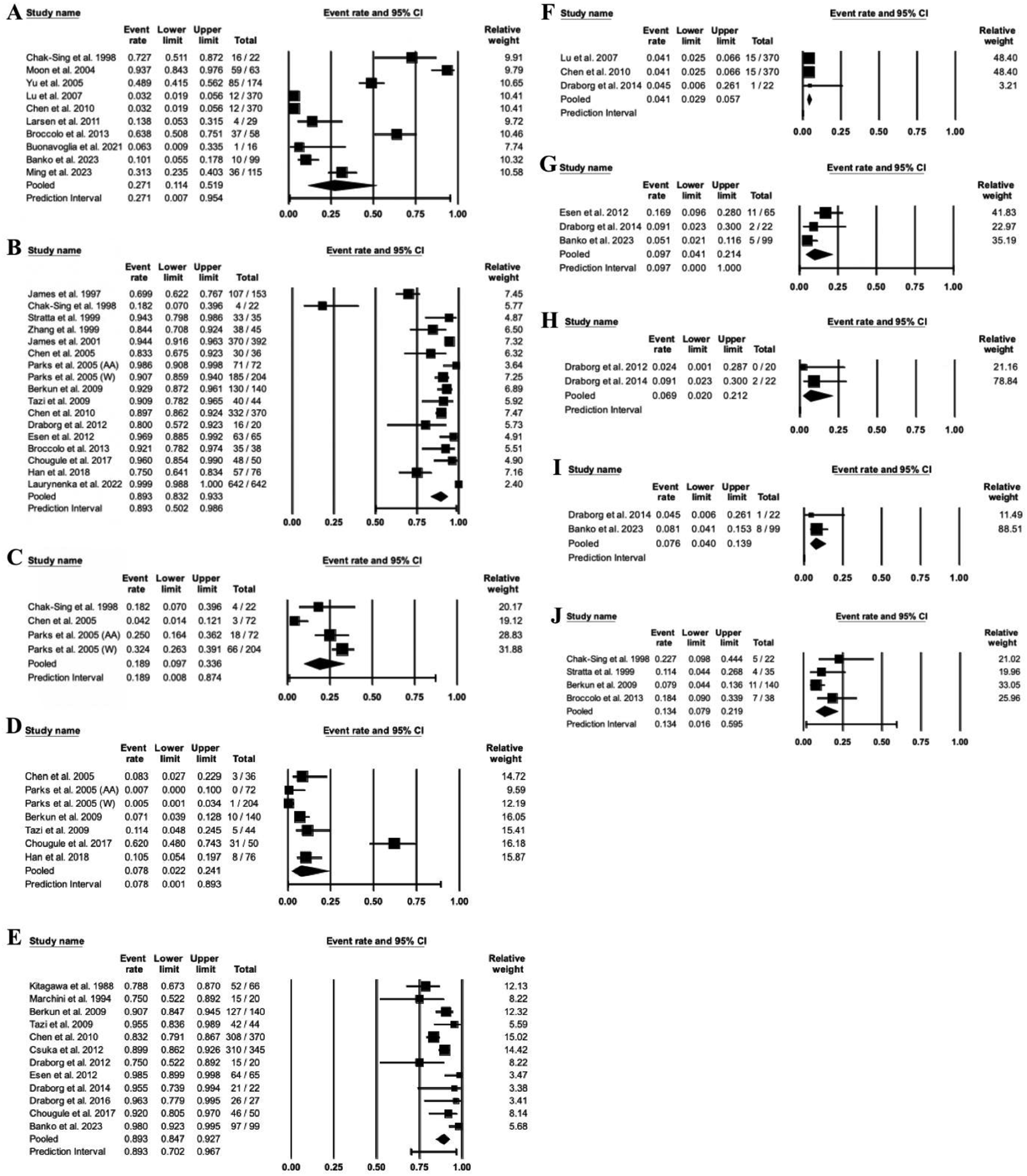
Association of EBV DNA and immunoglobulins with SLE
Furthermore, we explored a possible link between EBV infection and SLE development. A total of 28 studies were found eligible for the analysis, comprising 2968 patients and 3926 controls exploring the prevalence of EBV DNA and related immunoglobulins in patients and controls. As shown in Figure 3, the prevalence of EBV DNA (pooled OR: 3.94, 95% CI = 1.94–7.98, z value = 3.81, p = .00) and immunoglobulins such as VCA IgG (pooled OR: 2.31, 95% CI: 1.31–4.05, z value = 2.91, p = .004), VCA IgA (pooled OR: 5.45, 95% CI: 1.92–15.47, z value = 3.19, p = .001), VCA IgM (pooled OR: 2.02, 95% CI: 1.13–3.60, z value = 2.38, p = .01), EBNA1 IgA (pooled OR: 10.40, 95% CI: 6.51–16.61, z value = 9.79, p = .00), EA/D IgG (pooled OR: 8.56, 95% CI: 4.25–17.22, z value = 6.02, p = .00), EA/D IgA (pooled OR: 23.81, 95% CI: 5.55–102.14, z value = 4.26, p = .00), EA/D IgM (pooled OR: 3.49, 95% CI: 1.56–7.80, z value = 3.05, p = .002) and EA IgG (pooled OR: 4.14, 95%CI: 1.19–14.41, z value = 2.23, p = .02) were significantly higher in SLE patients than the healthy controls indicating an essential role of EBV infections in the pathogenesis of SLE. However, the occurrence of EBNA1 IgG was also higher in SLE patients than in healthy controls, but the difference was not statistically significant (Figure 3). Forest plots for the association of EBV DNA and antibodies with the development of SLE. A meta-analysis was conducted to determine the association between EBV DNA and antibodies against EBV in patients with SLE and healthy controls. This analysis was executed using CMA v4 software. Forest plots depict the pooled odds ratio, 95% confidence intervals (CI), the number of positive cases relative to the total number of samples in both cases and controls, as well as the z value and p values. A p value of less than 0.05 was considered statistically significant. The specific components analyzed include: (a) EBV DNA, (b) Viral Capsid Antigen (VCA) IgG, (c) VCA IgA, (d) VCA IgM, (e) Epstein-Barr Nuclear Antigen 1 (EBNA1) IgG, (f) EBNA1 IgA, (g) Early Antigen/Diffuse (EA/D) IgG, (h) EA/D IgA, (i) EA/D IgM, and (j) Early Antigen (EA) IgG.
Sensitivity analysis
Sensitivity analysis was used to investigate the robustness of the meta-analysis results by excluding one study at a time, performing the meta-analysis, and comparing the new analysis results with the original meta-analysis’s observations. A significant deviation between the latest analysis (after the exclusion of one study) and the original analysis indicates the non-reliability of the meta-analysis, and the results must be interpreted with caution. In contrast, the minimal deviation among them reveals that the results of the meta-analysis are robust. In the present study, sensitivity analysis showed that the prevalence of EBV DNA and immunoglobulins in healthy controls and their association with SLE indicated minimal deviations from the results of the parent meta-analysis, suggesting the robustness of the observations. (Supplemental Figures 4 and 5).
Discussion
The present systematic review and meta-analysis aimed to explore the association between EBV infection and SLE by synthesizing data from previously published articles that investigated EBV prevalence in healthy subjects and compared the differential positive rates between SLE patients and healthy controls. This study provides compelling evidence for a significant association between EBV infection and SLE, further strengthening the earlier findings that EBV infection may act as a key environmental trigger in SLE pathogenesis.8,9
Data on the prevalence of EBV DNA and antibodies against EBV in healthy controls revealed several interesting findings. EBV DNA was present in more than one-fourth of the healthy population, indicating that most healthy individuals harbor latent EBV. Furthermore, the high seroprevalence rates of EBV VCA IgG and EBNA1 IgG (89%) indicate past and persistent EBV infection. On the other hand, the lower positivity rates of EBV VCA IgM (7%) and EBV EA IgG (12%) indicate that active or recent EBV reactivation is uncommon in healthy subjects. Notably, the mismatch in the EBV DNA positivity rate and EA IgG seropositivity further highlights the role of another mechanism in viral shedding and antibody response in asymptomatic subjects.
The present study deciphered an essential association between EBV infections and SLE manifestations as a higher rate of EBV DNA and EBV-specific immunoglobulins were observed in patients compared to the controls. These observations align with earlier reports suggesting that the EBV infection may trigger SLE development in genetically susceptible individuals.42–44 The capacity of EBV to remain inside the host for lifelong 45 and, in certain conditions, reactivate themselves may facilitate chronic inflammation and lead to the production of autoantibodies and immune dysregulations, 46 which are the characteristics of SLE. The possible mechanisms by which EBV infections are linked with the pathogenesis could be attributed to molecular mimicry, bystander activation, and epitope spreading, which could trigger autoimmune responses.1,3 EBV nuclear antigens, especially EBNA-1, show significant homology with human proteins, which may induce cross-reactive autoantibodies, a process that could contribute to the development of autoimmunity in genetically predisposed individuals.4,5 Harley et al., (2018) demonstrated that EBNA-2 occupies several SLE risk loci, providing further support for EBV’s role in altering the immune response in SLE patients. Moreover, the association of EBV antibodies, particularly VCA and EA/D IgA and IgM, with SLE suggests that recent or reactivated EBV infections may be involved in SLE’s progression, possibly due to EBV’s capacity to persist in latent form within the immune system. 6
Interestingly, our meta-analysis did not find a significant association between EBNA1 IgG and SLE, which may be attributed to the high background seroprevalence of this antibody in the general population. 10 EBNA1 IgG is known to persist for life after EBV infection, often in a latent state, making it challenging to differentiate between EBV exposure and active infection in terms of its association with SLE. In contrast, the more variable antibodies, such as EA/D IgG, IgA, and IgM, appear to be more predictive markers of EBV reactivation and are significantly associated with SLE.
The findings of our meta-analysis also highlight some limitations in the current literature. While significant heterogeneity was observed in some of the analyses, particularly regarding antibody prevalence, this variability may arise from differences in study populations, sample sizes, detection methods, and geographical variations. The substantial heterogeneity in the EBV IgG and IgM antibody prevalence among studies suggests the need for standardized methodologies and the requirement for more extensive multicenter studies to confirm these associations. 13 Furthermore, although we used NOS guidelines to ensure the quality of the included studies, variation in quality remains a limitation, emphasizing the importance of high-quality, well-designed future studies. 11
This study underscores the importance of considering EBV as a contributing factor in SLE. However, the high prevalence of EBV in the general population complicates direct causal inference. Future research should focus on understanding the mechanisms underlying EBV reactivation in SLE patients and identifying specific immune responses that may predispose individuals to autoimmunity post-EBV infection. Additionally, prospective studies examining the impact of EBV vaccination or antiviral treatments on SLE incidence could offer valuable insights into potential prevention strategies.
Conclusion
In conclusion, our meta-analysis reinforces the association between EBV infection and SLE, providing a more comprehensive understanding of EBV’s role in SLE pathogenesis. These findings support the hypothesis that EBV infection, particularly its reactivation markers, may play a significant role in SLE development and progression. Further research is essential to clarify the underlying mechanisms and explore therapeutic interventions targeting EBV to potentially mitigate SLE risk and progression.
Supplemental Material
Supplemental material - Epstein-Barr virus infection and its association with systemic lupus erythematosus: Systematic review and meta-analysis
Supplemental material for Epstein-Barr virus infection and its association with systemic lupus erythematosus: Systematic review and meta-analysis by Shovit Ranjan, Sunil Kumar, Hrushikesh Nayak and Aditya K Panda in Lupus
Supplemental Material
Supplemental material - Epstein-Barr virus infection and its association with systemic lupus erythematosus: Systematic review and meta-analysis
Supplemental material for Epstein-Barr virus infection and its association with systemic lupus erythematosus: Systematic review and meta-analysis by Shovit Ranjan, Sunil Kumar, Hrushikesh Nayak and Aditya K Panda in Lupus
Footnotes
Acknowledgment
The authors would like to thank all faculty and staff of the Department of Biotechnology for their support whenever required.
Author contributions
Shovit Ranjan: Literature search, analysis, interpretation, writing first draft of the manuscript. Sunil Kumar: data analysis and interpretations, preparation of the first draft of the manuscript. Hrushikesh Nayak: data analysis and interpretations, preparation of the first draft of the manuscript. Aditya K Panda: inception of the idea, overall supervision, finalization of the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Data Availability Statement
Data will be made available on request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.