
Abstract
Amniotic epithelial cells (AECs) have immunomodulatory and anti-inflammatory properties that may improve outcomes in cell transplantation. However, their effect on islet engraftment after intraportal co-transplantation remains unclear. We evaluated the impact of co-transplanting syngeneic 600 islet equivalents (IEQs) with human AECs (hAECs) via the portal vein in a rat streptozotocin-induced diabetes model. The co-transplantation (Co-Tx) group showed normalization of blood glucose levels within 7 days after transplantation, sustained normoglycemia thereafter, and achieved a higher diabetes reversal rate than controls (100% vs. 71.4%, p < 0.01). Serum CXCL1 levels were significantly lower in the Co-Tx group indicating suppression of early inflammatory responses. Thrombin-antithrombin complex (TAT) levels also tended to be lower, raising the possibility of attenuation of the instant blood-mediated inflammatory reaction (IBMIR). In contrast, no significant differences were observed in VEGF levels or intrahepatic microvascular density. Co-transplantation with hAECs enhances islet engraftment likely through suppression of early inflammation, highlighting their potential as an adjunctive cellular therapy in islet transplantation.
Introduction
Type 1 diabetes mellitus (T1D) is a chronic disease caused by progressive loss of pancreatic β-cells, leading to insulin deficiency and the need for lifelong insulin therapy. 1 Islet transplantation offers a minimally invasive and effective treatment option for patients with severe T1D.2,3 However, widespread clinical application remains limited due to several challenges, including a shortage of suitable donors, low islet yield from donor pancreata, 4 and early graft loss following transplantation. 5 In particular, early islet damage after intraportal transplantation, caused by inflammation, hypoxia, and instant blood-mediated inflammatory reaction (IBMIR), significantly impairs engraftment. IBMIR is a rapid, innate immune response characterized by platelet activation, coagulation cascade initiation, and neutrophil recruitment upon direct contact between transplanted islets and recipient blood. Improving islet engraftment remains a key priority for advancing this therapy.
Human amniotic epithelial cells (hAECs), derived from the innermost layer of the amniotic membrane, are perinatal stem cells that exhibit multiple advantageous properties for transplantation.6–9 They express low levels of classical HLA class I molecules, lack HLA class II expression, and express the immunomodulatory molecule HLA-G. 10 hAECs produce a range of anti-inflammatory and immunosuppressive cytokines, inhibit T cell proliferation, and have been shown to promote epithelialization, angiogenesis, and tissue regeneration.7,11 Importantly, hAECs are non-tumorigenic and can be isolated in large numbers from discarded placental tissue using non-invasive procedures, making them ethically and practically attractive for clinical applications. 8
Mesenchymal stem cells and adipose-derived stem cells have been extensively studied as adjunct cell therapy in islet and hepatocyte transplantation due to their immunomodulatory and angiogenic capabilities.12–15 However, unlike these adult-derived stem cells, hAECs offer distinct advantages, including superior accessibility, a more favorable immunological profile, and lower tumorigenic risk. 8 Furthermore, the anti-inflammatory and immunomodulatory properties of hAECs may contribute to suppression of early inflammatory responses such as IBMIR. Despite these promising features, the impact of hAECs on islet engraftment via intraportal co-transplantation has not been previously evaluated in vivo.
Given the clinical relevance of the intraportal transplantation route and the unique characteristics of hAECs, this study aimed to investigate whether co-transplantation of syngeneic islets with hAECs improves islet engraftment in a rat intraportal transplantation model. We hypothesized that hAECs would enhance islet engraftment primarily by suppressing early post-transplant inflammation, including IBMIR and C-X-C motif chemokine ligand 1 (CXCL1)-mediated inflammatory signaling.
Materials and methods
Experimental animals
Male Lewis rats (9–12 weeks old; Japan SLC, Shizuoka, Japan) were used as syngeneic islet donors and recipients. All animal experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals 16 and were approved by the Tohoku University Expert Committee on Animal Experiments (Approval No.: 2019 MdA-191-03). Islet transplantation was performed under inhalational anesthesia with isoflurane (Viatris Inc., Canonsburg, PA, USA) to minimize animal distress.
Islet isolation
Islet isolation was performed as previously described. 17 Briefly, pancreata were perfused with collagenase solution via the common bile duct, digested, and purified using a density gradient. Islets were cultured overnight in RPMI-1640 medium (Thermo Fisher Scientific Inc.) containing 5.5 mmol/L glucose, 1% penicillin-streptomycin and 10% fetal bovine serum at 37°C in a humidified atmosphere of 5% CO2. Isolated islets were transplanted the following day.
Amniotic membrane collection, hAEC isolation, and thawing
Written informed consent was obtained from all mothers one day prior to elective cesarean section. Immediately after placenta removal, surface blood was rinsed off with sterile saline, and the amnion, located on the superficial fetal layer of the placenta, was carefully separated. The tissue was washed twice with saline and stored in a 50 mL centrifuge tube (Corning Inc., Kennebunk, ME, USA). Amniotic membranes were digested with 5 mL/g TrypLETM Select Enzyme (10×; Thermo Fisher Scientific Inc.) to isolate epithelial cells. This procedure was performed according to a previously established protocol. 18 Isolated cells were cryopreserved at 1.0 × 107 cells per cryovial in NutriFreez® D10 Cryopreservation Medium (Sartorius Stedim Biotech S.A., Goettingen, Germany). Cryopreserved hAECs were thawed using the ThawSTAR® system (BioLife Solutions Inc., Bothell, WA, USA) and used for transplantation after confirming cell viability greater than 80% by trypan blue exclusion. Thawed hAECs were directly used for transplantation experiments without further in vitro expansion or passaging.
Islet transplantation
Diabetes was induced in recipient Lewis rats by intravenous injection of streptozotocin (Sigma Aldrich Co., LLC, St. Louis, MO, USA) via the penile vein at a dose of 70 mg/kg, seven days prior to transplantation. Diabetes was confirmed when non-fasting blood glucose levels exceeded 350 mg/dL on two consecutive occasions. In the control group, 600 islet equivalents (IEQs) in 300 μL of medium were transplanted into the portal vein. In the co-transplant (Co-Tx) group, 600 IEQs were mixed with 1.0 × 106 hAECs in 300 μL medium and transplanted into the portal vein (Figure 1). Protocol for islet-hAEC transplantation.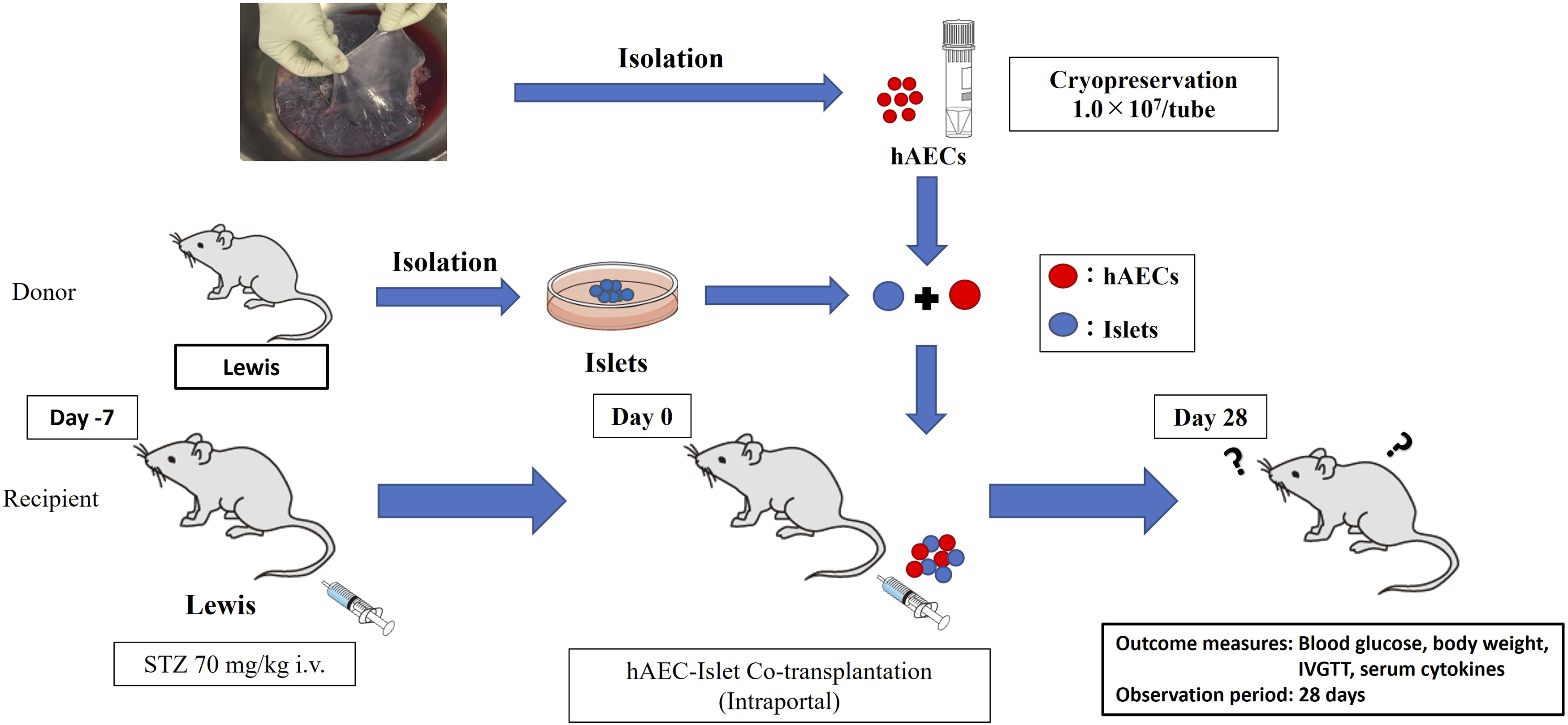
In vivo experimental groups
The in vivo experimental design is illustrated in Figure 1. Diabetes reversal was defined as sustained blood glucose levels below 200 mg/dL thereafter. Blood samples were collected on days 0, 1, 3, 5, and 7 after transplantation. Blood glucose and body weight were monitored two to three times per week until day 28 post-transplantation. An intravenous glucose tolerance test (IVGTT) was performed between days 28 and 35. Rats were euthanized between days 35 and 42, and liver specimens were collected.
Intravenous glucose tolerance test (IVGTT)
Rats were fasted for 14 hours prior to the test. A 30% glucose solution (Otsuka Pharmaceutical, Tokyo, Japan) was administered intravenously via the penile vein at a dose of 1.0 g/kg body weight. Blood glucose levels were measured at 5, 10, 20, 30, 60, 90, and 120 minutes after administration.
Determination of serum cytokines
Serum cytokine levels were measured using the MILLIPLEX® Rat Cytokine/Chemokine Panel 27 Plex Premix (Merck KGaA, Darmstadt, Germany) with the Bio-Plex® 200 system (Bio-Rad Laboratories, Inc., Hercules, CA, USA), according to the manufacturers’ instructions.
Immunohistochemical analysis
Recipient rats were sacrificed at 3 hours, 3 days, 7 days, and 35-42 days post-transplantation for histological evaluation. Separate recipient animals were used for each time point. Liver sections were subjected to hematoxylin and eosin staining and immunohistochemically stained with the following primary antibodies: anti-human leukocyte antigen (HLA)-ABC (ab70328, Abcam plc, Cambridge, UK), anti-insulin (ab181547, Abcam plc), anti-CD31 (ab182981, Abcam plc), and anti-CD4 (ab237722, Abcam plc).
Determination of thrombin-antithrombin (TAT) complex levels
Blood plasma samples were serially collected from the same recipient animals at 0, 15, 30, 60, and 180 minutes after islet transplantation from the Co-Tx group (n = 4) and the control group (n = 7). TAT complex levels were measured by enzyme-linked immunosorbent assay (ELISA) using a Rat TAT Complexes ELISA Kit (Novus Biologicals, Centennial, CO, USA).
Cytokine measurement in co-culture
Direct co-culture was performed using 600 IEQs, with or without 1.0 × 106 hAECs in standard islet culture medium for 72 hours. For indirect co-culture, islets and hAECs were physically separated using a cell culture insert (Thermo Fisher Scientific). Cytokine concentrations in culture supernatants were measured in the direct co-culture, indirect co-culture, and control groups using the MILLIPLEX® Rat Cytokine/Chemokine Panel 27 Plex Premix assay.
Statistical analysis
Data are expressed as mean ± standard deviation. Analyses were performed using JMP Pro 17 (SAS Institute Inc., NC, USA). Mixed-effects models were used to analyze longitudinal changes in blood glucose and serum cytokines. The Wilcoxon rank-sum test was used for between-group comparisons of cytokine levels at each time point. All tests were two-tailed, and p < 0.05 was considered statistically significant.
Ethical approval
All procedures involving human and animal samples were approved by the Ethics Committee of Tohoku University Graduate School of Medicine (Approval No.: 2018-1-203; approved on June 18, 2018) and the Tohoku University Expert Committee on Animal Experiments (Approval No.: 2019 MdA-191-03; effective from April 1, 2019). Written informed consent was obtained from all mothers prior to placenta collection.
Results
Blood glucose and body weight after transplantation
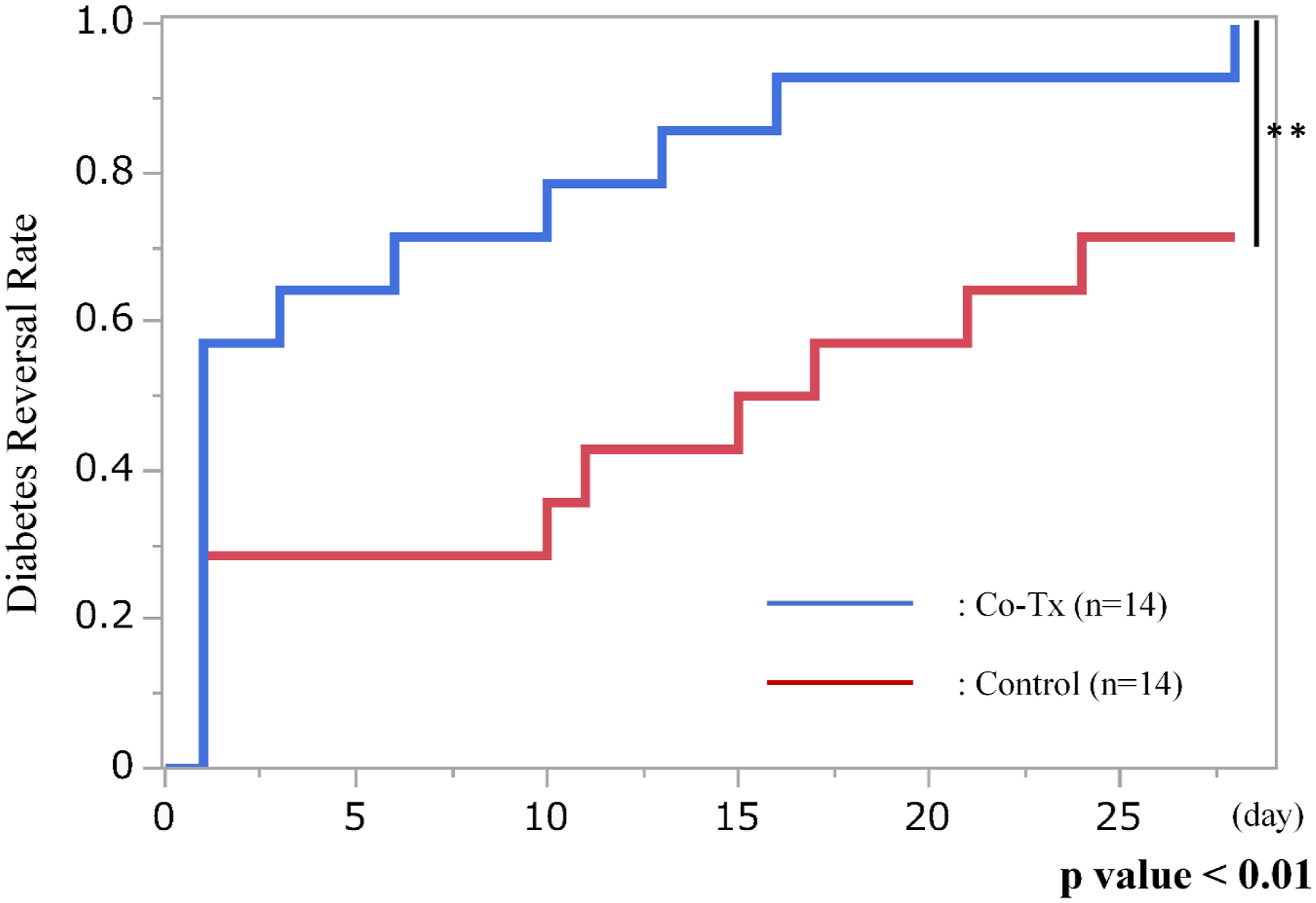
Blood glucose levels over the 28-day period following transplantation are shown in Figure 2(a) and (b). Compared with the control group, the Co-Tx group exhibited consistently and significantly lower blood glucose levels throughout the observation period (p = 0.041 by mixed-effects model). At day 28 post-transplantation, all rats in the Co-Tx group (14/14, 100%) achieved diabetes reversal compared with 10 of 14 rats (71.4%) in the control group (p < 0.01) (Figure 3). The rate of body weight gain did not differ significantly between the two groups during the 28-day period (Co-Tx: 24.1 ± 1.56% vs. Control: 22.6 ± 2.14%; p = 0.621). Blood glucose trends after transplantation. (a): Blood glucose levels of individual rats during the 28 days following transplantation. Rats in the Co-Tx group showed consistently lower glucose levels than those in the control group, indicating improved glycemic control. (b): Mean blood glucose levels during the first 28 days were significantly lower in the Co-Tx group compared to the control group (*p < 0.05). Statistical comparisons between groups at individual post-transplant time points are indicated in the figure, with a significant difference observed on day 13 after transplantation. Diabetes reversal rate after islet transplantation.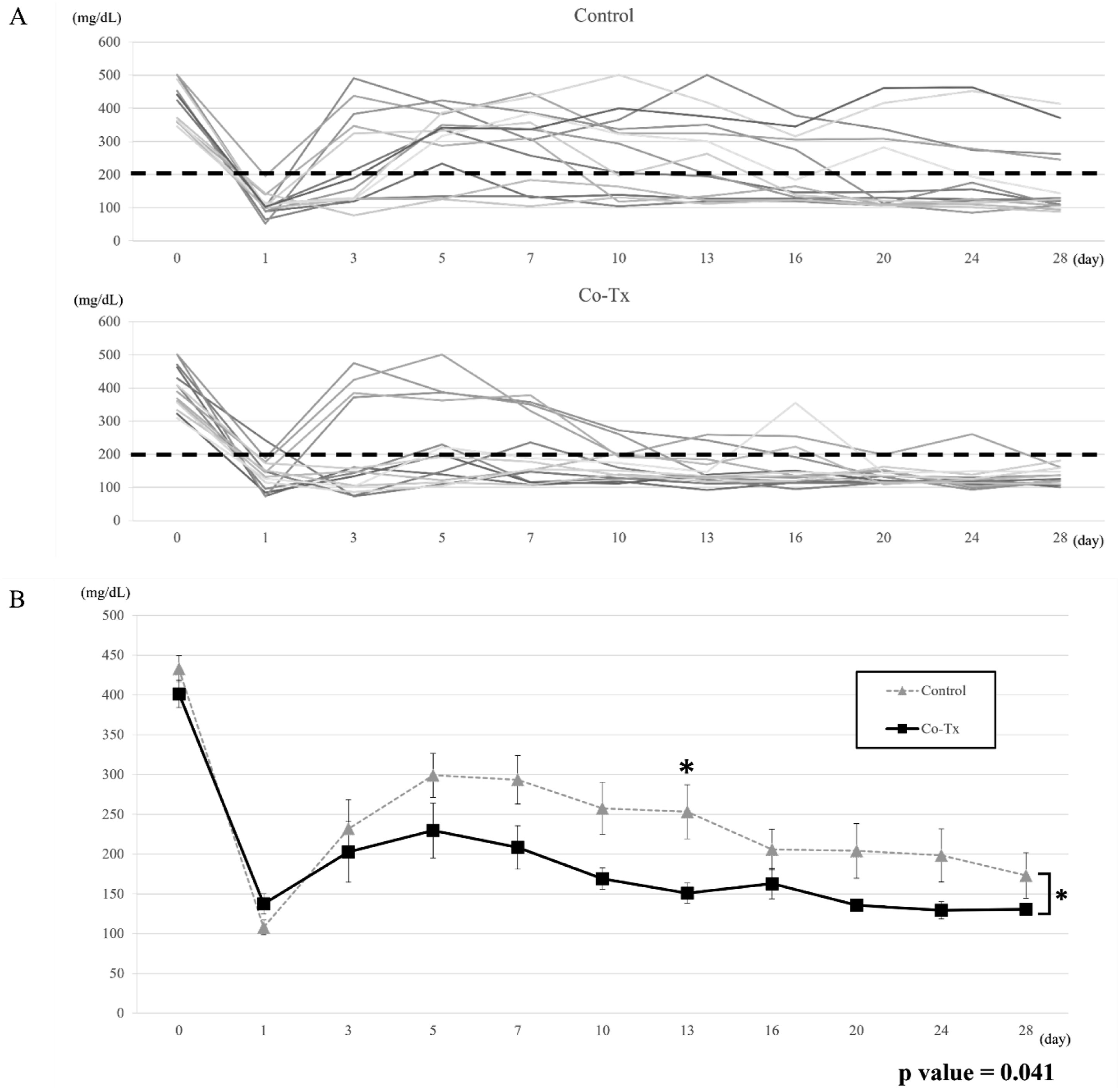
Intravenous glucose tolerance test (IVGTT)
Blood glucose responses during the IVGTT performed 28–35 days after transplantation are shown in Figure 4(a) and (b). No significant differences were observed between the Co-Tx and control groups in glucose levels at any individual time point or in the area under the curve (AUC) (Co-Tx: 30083 ± 1703.7 vs. Control: 31112 ± 2004.7 mg·min/dL; p = 0.623). Results of intravenous glucose tolerance test (IVGTT). (a): Blood glucose levels measured during IVGTT performed 28–35 days after transplantation are shown. No significant differences in blood glucose responses were observed between the Co-Tx and control groups. (b): Area under the curve (AUC) for blood glucose levels from 0 to 120 minutes during IVGTT (performed on days 28–35 post-transplantation). No significant difference was observed between the Co-Tx and control groups.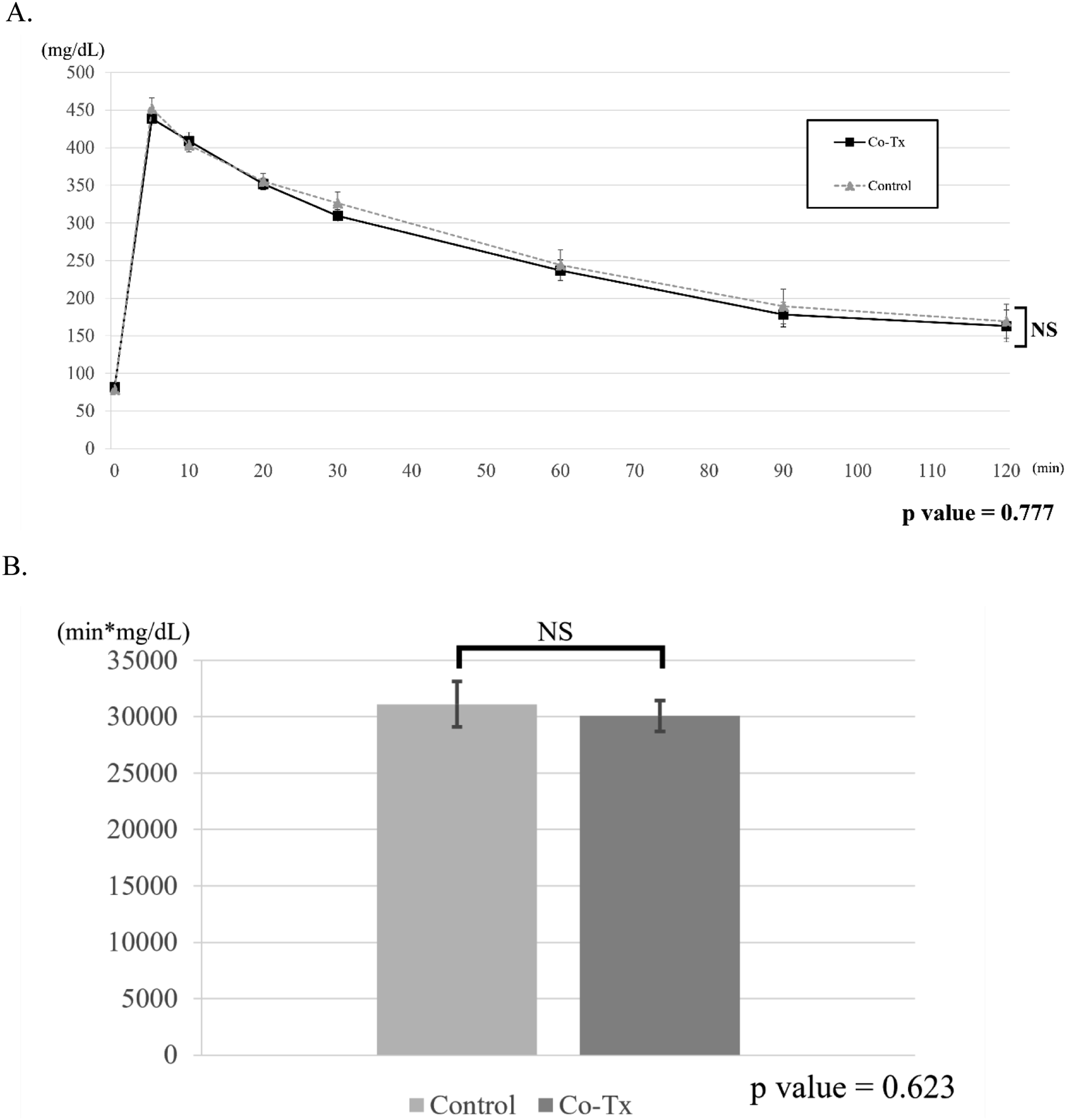
Serum cytokine levels
Serum cytokine trends during the early post-transplant period are shown in Figure 5. Among the measured cytokines, only growth-regulated oncogene/keratinocyte chemoattractant (GRO/KC, also known as CXCL1) exhibited a significantly different overall trend between groups within the first 7 days (p = 0.027). GRO/KC levels were significantly lower in the Co-Tx group only on day 1 (p = 0.011). On day 7, levels of interleukin (IL)-4, IL-5, and IL-12p70 were significantly lower in the Co-Tx group than in the control group (IL-4: p = 0.041; IL-5: p = 0.005; IL-12p70: p = 0.011). IL-10 level also tended to be lower in the Co-Tx group, although the difference was not statistically significant (p = 0.064, respectively). Analysis of serum cytokine levels.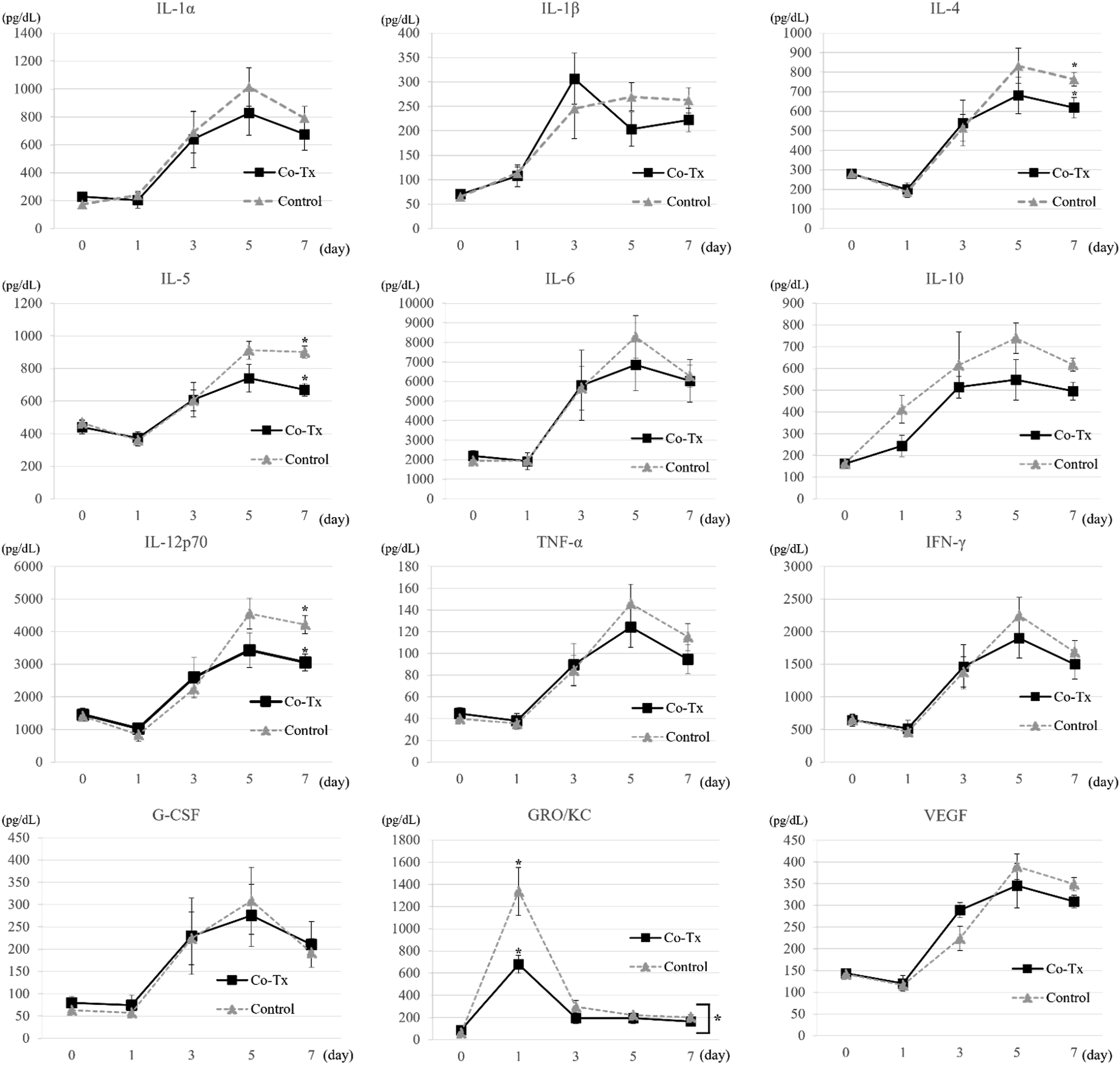
Immunohistochemical analysis
Livers harvested at 3 hours, 3 days, 7 days, and 35–42 days post-transplantation were subjected to hematoxylin-eosin staining and immunostaining for insulin, HLA-ABC, CD31, and CD4 (Figure 6(a)–(d)). No HLA-ABC-positive hAECs were detected around the engrafted islets. CD31 staining, used to assess peri-islet angiogenesis, and CD4 staining, used to evaluate inflammatory cell infiltration, showed no appreciable differences between the two groups. However, quantitative analysis of CD31-positive vascular density and CD4-positive cell infiltration was not performed, which represents a limitation of this study. Histological and immunohistochemical findings in the liver following islet-hAEC transplantation.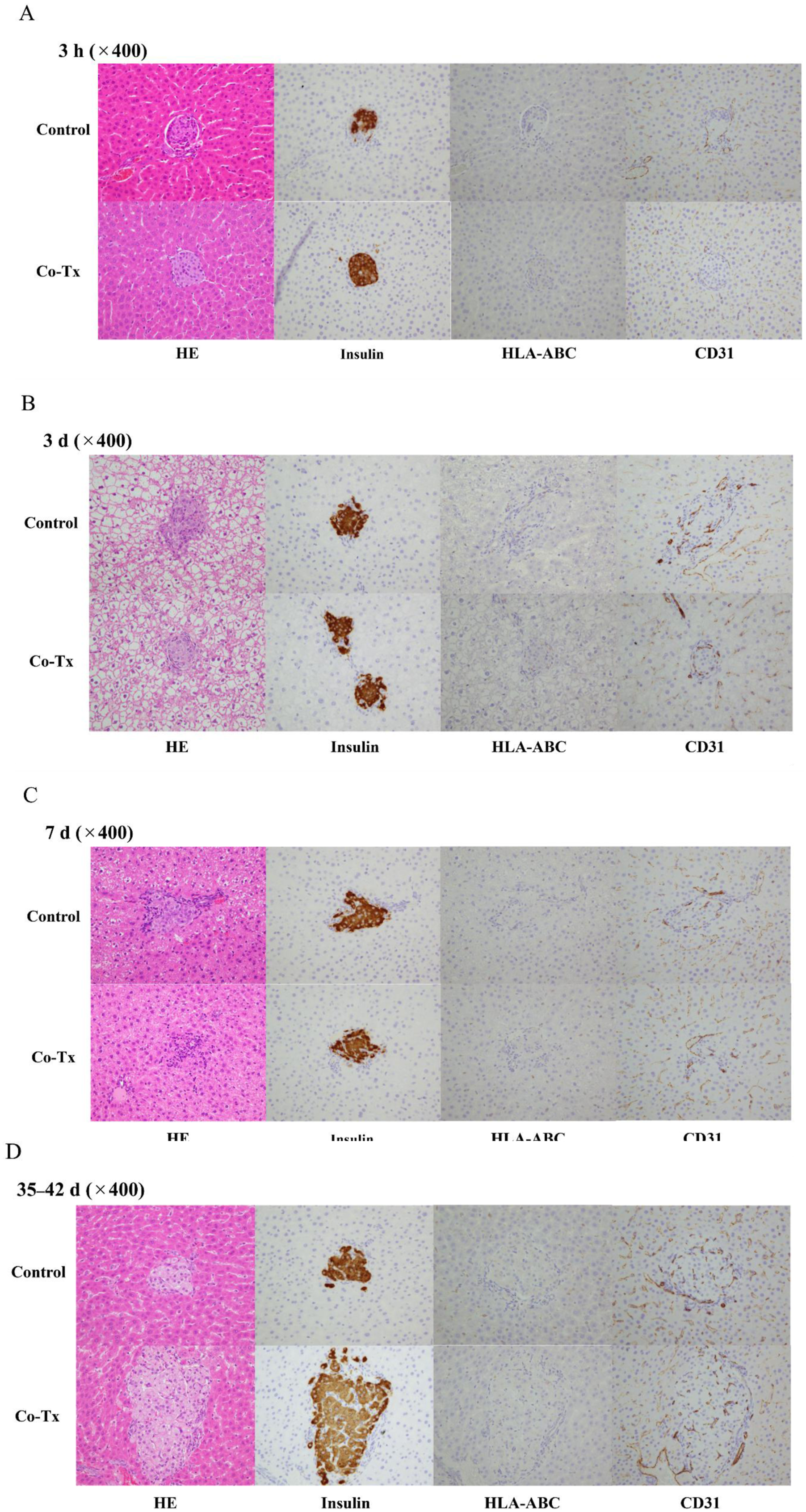
Thrombin–antithrombin (TAT) complex levels
Plasma TAT levels over time are shown in Figure 7. TAT levels peaked at 15 minutes after transplantation in both groups and then declined. The peak TAT level tended to be lower in the Co-Tx group (46.8 ± 4.4 ng/mL) than in the control group (103.3 ± 46.8 ng/mL), but the difference was not statistically significant. This may be partly attributed to the substantial inter-individual variability observed in the control group. Plasma thrombin–antithrombin complex (TAT) levels.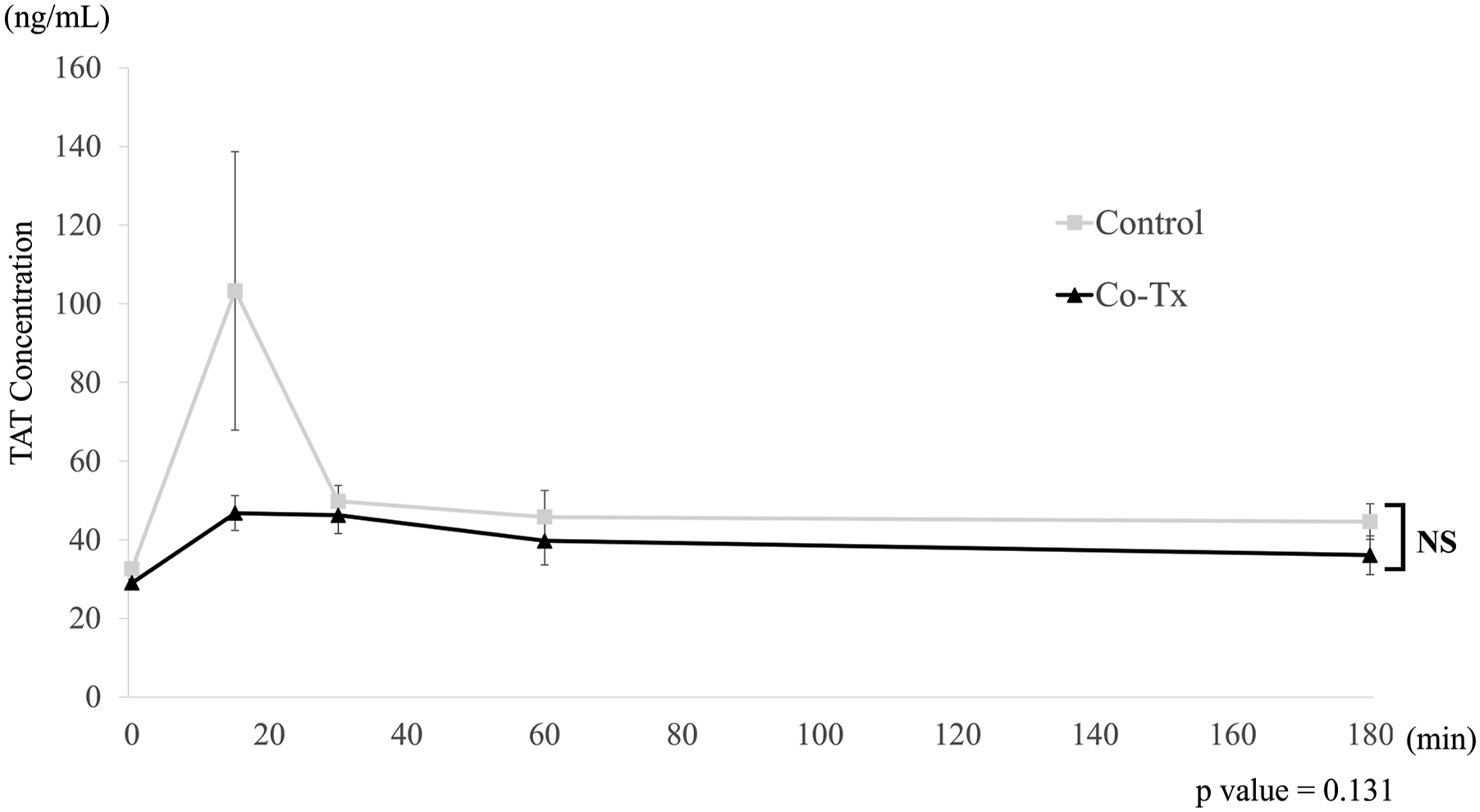
Cytokine levels in in vitro co-culture
Compared with the control group, GRO/KC levels were significantly lower in the direct co-culture group (p = 0.002), consistent with the in vivo findings. IL-4 (p = 0.020) was also significantly suppressed in the direct co-culture group compared with the control group. No significant difference was observed in IL-12p70 levels in vitro, despite the difference noted in vivo. When comparing direct and indirect co-culture conditions, direct co-culture showed a greater suppressive effect on inflammatory cytokine production; however, GRO/KC and MIP-2 were the only cytokines that showed statistically significant differences between these two conditions (p = 0.011, 0.022) (Figure 8). Determination of cytokine levels in co-culture supernatants.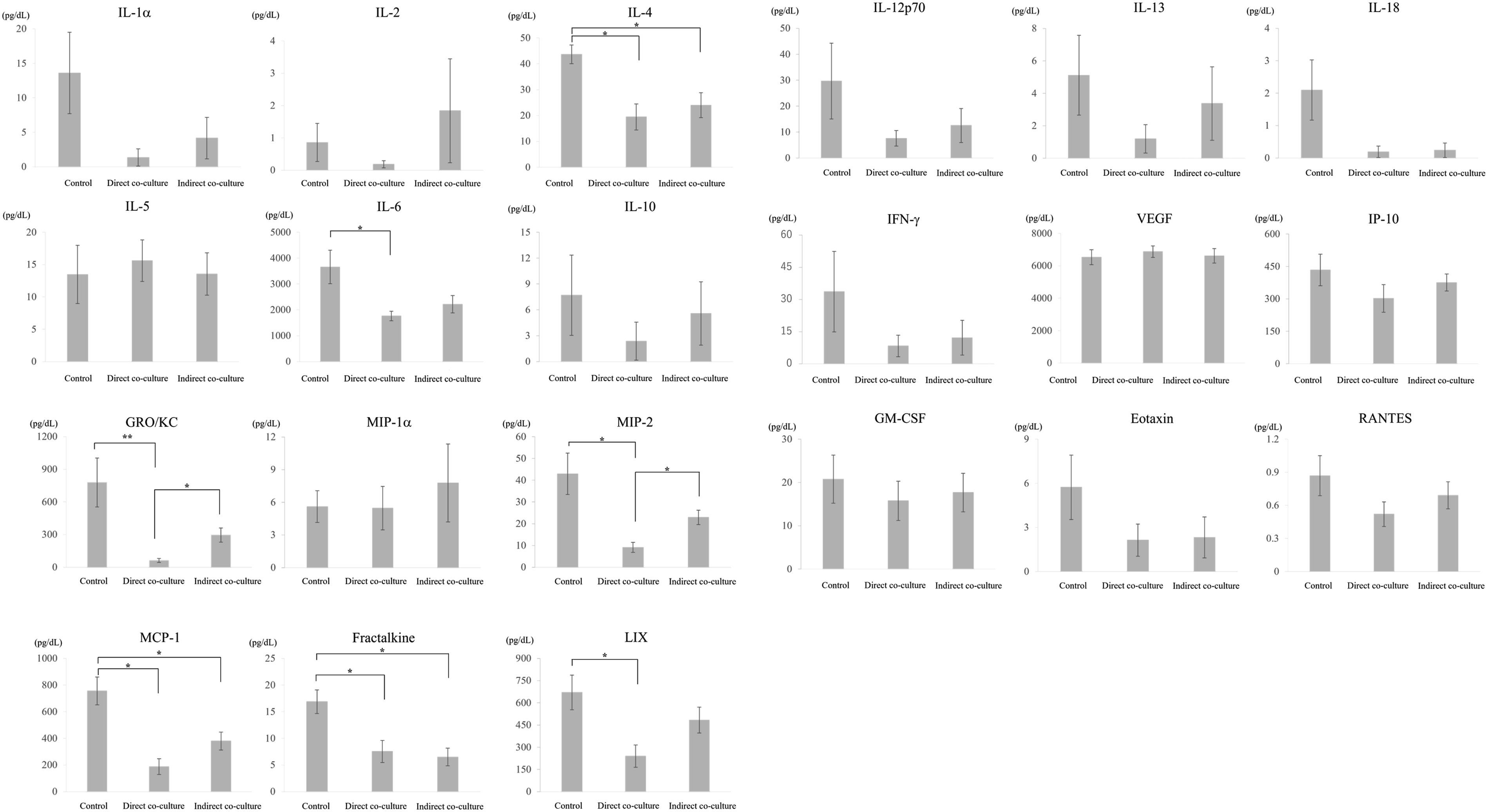
Discussion
In the field of cell transplantation, inflammation, ischemia, and immune rejection are considered the three major factors that prevent islet engraftment.2,3 The success of treatment largely depends on how effectively these three barriers can be controlled. While rejection can be managed to some extent through the use of immunosuppressive agents, this approach has limitations, including drug-induced side effects and the necessity for lifelong administration. 19 hAECs possess several beneficial properties for transplantation, including anti-inflammatory, angiogenesis-promoting, and immunomodulatory effects.7,20,21 Based on these characteristics, we initiated this study with the expectation that hAECs would enhance islet engraftment.
This study demonstrated that syngeneic islet-hAEC co-transplantation promotes islet engraftment and improves the diabetes reversal rate. To our knowledge, this is the first study to demonstrate that intraportal islet-hAEC co-transplantation improves islet engraftment efficiency. Based on the observed enhancement in engraftment, we investigated the underlying mechanisms, focusing on inflammation, ischemia, and immune rejection.
It is estimated that 50–70% of intraportally transplanted islets are destroyed by IBMIR22,23 and by the gut-liver axis, a component of the liver’s innate immune system, even before they are affected by acquired immune responses such as alloimmunity and autoimmunity. 2 Therefore, controlling this innate immune response in the liver is critical for improving islet engraftment. In the early post-transplant period, we compared serum inflammatory cytokine levels and found a significant difference only in CXCL1 (GRO/KC), a chemokine that promotes neutrophil migration and contributes to inflammation. 24 CXCL1 is rapidly released from transplanted islets immediately after transplantation, and previous studies have reported that suppressing CXCL1 signaling can improve transplantation efficiency.25,26 In the present study, CXCL1 levels peaked on the first day after transplantation, consistent with previous reports. 25 These findings suggest that hAEC co-transplantation may suppress CXCL1-mediated inflammatory responses, which could contribute to reduced neutrophil recruitment and improved islet engraftment. Although no significant differences were observed in other cytokines, overall levels of inflammatory cytokines were lower in the Co-Tx group. Interestingly, IL-10, an anti-inflammatory cytokine, was also reduced, suggesting hAECs may exert an immunomodulatory effect by suppressing systemic inflammation. Consistent with these in vivo findings, our in vitro co-culture study also demonstrated that co-culture of islets with hAECs suppressed CXCL1 levels. In addition, MIP-2, another neutrophil-associated chemokine, was significantly reduced in the direct co-culture condition, suggesting that direct interaction between islets and hAECs may contribute to the suppression of inflammatory signaling. Although no apparent differences in CD4-positive cell infiltration were observed between the groups, xenogeneic hAECs were transplanted into immunocompetent rats in the present study. Therefore, the xenogeneic origin of hAECs may have transiently influenced local immune and inflammatory responses during the very early post-transplant period.
IBMIR is triggered upon direct contact between the graft and blood and represents a major barrier to successful islet transplantation due to early graft loss.22,23 In our previous work, we reported that hAECs express tissue factor (TF), which is thought to promote IBMIR. 18 Despite their TF expression, plasma TAT levels in the Co-Tx group showed a non-significant trend toward lower levels shortly after transplantation. Although this finding does not provide definitive evidence, it raises the possibility that hAECs may modulate IBMIR during the very early post-transplant period. Previous studies have reported that hAECs can suppress complement activation, including inhibition of key complement components. 27 In addition to complement regulation, modulation of platelet activation may also contribute to IBMIR. A major limitation of this study is that complement activation and platelet activation were not directly evaluated. Further studies are required to clarify the relative contribution of complement- and platelet-mediated pathways to the potential effects of hAECs on IBMIR.
Islets are highly oxygen-dependent tissues, utilizing approximately 5–15% of total pancreatic blood flow. 28 During islet transplantation, islets are disconnected from their native oxygen supply following isolation. 29 For several days post-transplantation, the grafted islets rely solely on oxygen diffusion from the portal blood, which has relatively low oxygen tension. 2 Prolonged exposure to hypoxic conditions leads to necrosis and apoptosis, particularly in the central regions of larger islets. 30 In studies involving subcutaneous islet transplantation, increased oxygen tension has been shown to enhance islet engraftment. 31 Therefore, early and stable revascularization of graft islets through angiogenesis is considered crucial for promoting successful engraftment. Given the angiogenesis-promoting effect of hAECs, 32 we hypothesized that co-transplantation would enhance peri-islet vascularization around grafted islets. However, no marked difference in peri-islet vascularization, as assessed by CD31 staining, was observed between the two groups, although gradual vascular development around the grafts occurred in both. This finding was consistent with the serum cytokine assay, which showed no significant difference in VEGF levels between the groups. These findings suggest that suppression of early inflammation may have contributed more prominently to the observed improvement in engraftment than apparent changes in peri-islet vascularization as assessed by CD31 staining.
There are several limitations in this study. In clinical settings, most islet transplantations involve allogeneic grafts derived from deceased donors. One limitation of the present study is that a syngeneic transplantation model was used. Therefore, the influence of alloimmune responses on graft survival could not be evaluated, and the beneficial effects of hAEC co-transplantation observed in the present study may have been overestimated. Future studies using allogeneic transplantation models are warranted. The present study was primarily based on in vivo experiments, which made it difficult to assess local inflammatory responses in the liver, although systemic inflammatory changes could be monitored through serum cytokine levels. Furthermore, because xenogeneic human cells were transplanted into immunocompetent rats, the xenogeneic origin of hAECs may have transiently triggered inflammatory responses during the very early post-transplant period, despite their reported anti-inflammatory and immunomodulatory properties. This possibility should be considered when interpreting the inflammatory findings in the present study. In addition, HLA-ABC staining was employed to detect hAECs in liver tissue; however, since hAECs exhibit weak expression of HLA Class I molecules, their detection might have been insufficient. Previous reports suggested that the adhesion of hAECs around islets may promote angiogenesis.33,34 Therefore, additional analysis of the spatial relationship between islets and hAECs is warranted.
Further investigation is needed to determine whether the observed enhancement in islet engraftment is attributable to modulation of very early inflammatory responses, particularly innate immune mechanisms specific to the liver. This may help elucidate the mechanisms underlying early graft failure. Moreover, the effects on adaptive immunity also warrant further investigation.
Conclusion
This study demonstrated that hAECs promote islet engraftment and improve the reversal rate of diabetes. The beneficial effects of hAECs are likely attributable primarily to suppression of early inflammation, with possible modulation of IBMIR following transplantation (Supplemental material available online).
Supplemental material
Supplemental material - Amniotic epithelial cells enhance islet engraftment by suppressing early inflammation in intraportal transplantation
Supplemental material for Amniotic epithelial cells enhance islet engraftment by suppressing early inflammation in intraportal transplantation by Kaoru Okada, Kazuaki Tokodai, Miyako Tanaka, Yuki Watanabe, Hitomi Okita, Takako Ito, Masatoshi Saito, Michiaki Unno, Toshio Miki, Masafumi Goto, and Takashi Kamei in Cell Transplantation.
Footnotes
Acknowledgments
This work was supported by The Clinical Research Promotion Program for Young Investigators of Tohoku University Hospital, Public Trust Surgery Research Fund, Japan IDDM network, and Takeda Science Foundation. We are grateful to Experimental Animal Facility, Tohoku University Graduate School of Medicine for animal care. We thank Biomedical Research Unit of Tohoku University Hospital for technical support, and the Pathology section of the Biomedical Research Core, Tohoku University Graduate School of Medicine, for assistance in preparing tissue specimens.
Ethical considerations
All procedures involving human and animal samples were approved by the Ethics Committee of Tohoku University Graduate School of Medicine (Approval No.: 2018-1-203; approved on June 18, 2018) and the Tohoku University Expert Committee on Animal Experiments (Approval No.: 2019 MdA-191-03; effective from April 1, 2019). Written informed consent was obtained from all mothers prior to placenta collection.
Consent to participate
Written informed consent was obtained from all mothers one day prior to elective cesarean section.
Author contributions
Kaoru Okada: Participated in the performance of the research, the writing of the paper and data analysis. Kazuaki Tokodai: Participated in research design, obtaining funds and supervision of the thesis writing. Miyako Tanaka: Contributed in the collection of the research materials. Yuki Watanabe: Contributed in the collection of the research materials. Hitomi Okita: Contributed in the cell isolation and preservation of the research materials. Takako Ito: Contributed in the cell isolation and preservation of the research materials. Masatoshi Saito: Contributed the research materials. Michiaki Unno: Participated in supervision of thesis preparation. Toshio Miki: Participated in research design and supervision of thesis preparation. Masafumi Goto: Participated in supervision of thesis preparation. Takashi Kamei: Participated in supervision of thesis preparation. Disclosure: The authors of this manuscript have no conflicts of interest to disclose as described by Cell Transplantation.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Japan Society for the Promotion of Science (JSPS KAKENHI) Grant Number: 22K08745, Clinical Research Promotion Program for Young Investigators of Tohoku University Hospital, Kawano Masanori Memorial Public Interest Incorporated Foundation for Promotion of Pediatrics, The Japan Foundation for Pediatric Research, Public Trust Surgery Research Fund. The Japan Society for Organ Preservation and Biology, TERUMO LIFE SCIENCE FOUNDATION, Takeda Science Foundation, Japan IDDM network.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, Kazuaki Tokodai, upon reasonable request.
Use of AI-assisted tools
Artificial intelligence (AI)-assisted tools (ChatGPT, OpenAI) were used for English language refinement and preparation of the graphical abstract during manuscript preparation. No scientific data, analyses, or results were generated or modified using AI. The authors reviewed and verified all outputs and take full responsibility for the content of the manuscript.
Supplemental material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.