
Abstract
Background
Androgenic alopecia (AGA) is a common hair follicle miniaturization disease driven by androgens. Fibroblasts, especially dermal papilla (DP) cells, are considered the core of the pathogenesis of AGA, but there is a lack of systematic synthesis of evidence.
Objective
To systematically review the role of fibroblasts (DP cells and dermal sheath cells) in the pathogenesis of AGA, with a focus on androgen signaling transduction, paracrine signaling, disruption of stem cell niche, inflammation, and fibrosis.
Data sources
PubMed and Web of Science, covering the period from August 23, 2025 to May 28, 2026, including human basic research, animal model studies, and clinical pathology studies related to AGA fibroblast function. Two reviewers independently screened literature, extracted data, and evaluated the risk of bias.
Result
Through retrieval, a total of 1592 articles were identified, and 43 articles were ultimately included. DP cells consistently express high levels of androgen receptor (AR) and 5α-reductase type II. Activation of androgens (dihydrotestosterone) in DP cells leads to: (1) downregulation of Wnt/β-catenin signaling and upregulation of TGF-β/BMP pathway; (2) Changes in secretion profile (decrease in VEGF/IGF-1 and increase in DKK-1/TGF-β1); (3) DP cell aggregation behavior is impaired; (4) Destruction of hair follicle stem cell activation; (5) Inducing inflammation and fibrosis around hair follicles. The evidence for dermal sheath cells as a DP cell bank is still limited.
Conclusion
DP cells are the main transducers of androgen signaling in AGA, driving follicle miniaturization through multiple interrelated mechanisms. The current evidence supports DP cell dysfunction as a therapeutic strategy.
1. Introduction
1.1. Clinical and pathological characteristics of androgenetic alopecia
Androgenic alopecia (AGA) is a common genetic hair loss disease driven by androgens, manifested in different male and female patterns.1–4 Although it does not endanger life, it has a profound impact on psychological and social health as well as quality of life, making it a public health issue.5–7 Pathologically, the characteristic of AGA is the gradual miniaturization of hair follicles, with the final hair transforming into fine hairs.8,9 This is caused by the disorder of the hair cycle, which shortens the growth period and prolongs the rest period, resulting in hair gradually becoming thinner and shorter, unable to fully cover the scalp.10,11
While numerous narrative reviews have described AGA pathogenesis, a systematic synthesis of the evidence supporting the central role of fibroblasts—particularly dermal papilla (DP) cells—is lacking. Follicular miniaturization in AGA occurs unevenly on the scalp, and occipital and temporal follicles typically resist this process. 12 This regional difference implies fundamental differences in the response of hair follicles to microenvironmental signals, 13 the key regulatory factor being a specialized fibroblast population—the dermal papilla (DP) cells. 14 Therefore, a systematic review is needed to critically appraise and synthesize the available evidence on how fibroblasts mediate androgen effects and drive follicular miniaturization.
1.2. Fibroblast populations within the hair follicle structure
As a complex micro organ, hair follicles rely on precise and dynamic interactions between epithelial cells and mesenchymal cells for their development, growth, and cyclical regeneration. 15 During this process, two specific fibroblast populations—DP cells and dermal sheath (DS) cells—form the “mesenchymal unit” of the follicle.16,17 They are not only as important structural scaffolds, but also as central hubs regulating signal transduction.
The DP cells are a unique cluster of mesenchymal embedded in hair follicles, serving as the “command center” of hair follicular.18,19 Its core function is to secrete key growth factors (such as VEGF, FGF7, IGF-1), cytokines, and extracellular matrix molecules, thereby directing epithelial cell behavior and controlling the hair cycle.20–24 It is critical that DP cells highly express androgen receptor (AR) and 5α-reductase type II, making them the main sites for receiving and amplifying androgen signals.25,26 By altering the DP cells, androgens ultimately determine the fate of hair follicle, positioning the DP cells at the center of AGA pathogenesis.27,28 It is worth noting that the aggregative behavior of DP cells in vitro reflects this functional centrality,29,30 and their quantity and activity directly determine the size and type of hair follicles, making them the ultimate executor of follicular enlargement or miniaturization.
The perifollicular DS cells, composed of longitudinally arranged fibroblasts, envelops the outer root sheath. While traditionally viewed as a structural scaffold providing mechanical support,31,32 the DS cells is now recognized as a dynamic component with multiple active functions. It serves as a source of signaling molecules for communication with epithelial and DP cells32,33 and,crucially, as a reservoir for DP cells, which can be activated to form a new DP cells during anagen initiation—a process vital for follicular regeneration.34,35 Furthermore, the DS harbors multipotent stem cells implicated in follicle repair and homeostasis.31,36
In short, the DP cells and DS cells do not exist in isolation, but collaboratively shape the microenvironment that regulates hair follicle growth, morphogenesis, and circulation through complex interactions. In AGA, dysfunction of DP cells is a triggering factor for follicular miniaturization, while DS cells may also contribute to this pathological progression by participating in signaling regulation and cell recruitment process.
This review will systematically elaborate on the central role of fibroblasts in AGA, focusing on their biological properties, androgen signal transduction, mechanisms in driving follicular miniaturization, and the ensuing therapeutic strategies.
2. Methods
2.1. Protocol and registration
This systematic review was conducted and reported in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 statement. The protocol was registered on Open Science Framework (registration number: [DOI: 10.17605/OSF.IO/GVK5Q]).
2.2. Information sources and search strategy
The system search was conducted in PubMed and Web of Science, with a search period from the establishment of the database to May 28, 2026. The search term combination revolves around the two main themes of “androgenic alopecia” and “fibroblast”, using a combination of keywords and free words without limiting the research type or language. The specific search formula can be found in Appendix A.
2.3. Inclusion and exclusion criteria
Develop the following standards based on the PICOS framework:
Population (P): scalp tissue of patients diagnosed with AGA, AGA animal models (induced by testosterone/dihydrotestosterone), or hair follicle cells derived from AGA patients (especially DP cells, dermal fibroblasts, and hair sheath fibroblasts).
Intervention (I): Androgens (testosterone or dihydrotestosterone (DHT))
Control (C): Healthy controls, hair follicles/cells in non-hair loss areas, untreated normal cells or animals.
Outcome (O): Degree of hair follicle miniaturization, hair growth parameters (density, length, cycle), fibroblast proliferation/apoptosis/aging, extracellular matrix deposition, signaling pathway activity (TGF beta, Wnt, CXCL12, etc.).
Research type (S): Original research, including in vitro cell experiments, animal experiments, human tissue research, and clinical trials. Exclude reviews, systematic reviews, meta-analyses, conference abstracts, case reports (sample size<3 and no controls), editorials, hypothetical papers.
2.4. Selection process
Two reviewers (first author and second author) independently conduct literature screening, and any disagreements are resolved through discussion or consultation with a third reviewer. All retrieved records were exported to EndNote X9 for duplicate removal. The screening process is divided into two stages:
Initial screening (title and abstract): First, remove duplicate records, and then determine the title and abstract based on the inclusion and exclusion criteria mentioned above. If the information is not clear enough to exclude it, it will enter the full-text screening.
Re screening (full text): Obtain the full text of the literature that passed the initial screening and reevaluate its eligibility for inclusion. Detailed record of the reasons for exclusion.
The selection process is summarized in a PRISMA 2020 flow diagram.
2.5. Data collection and data items
Use a pre designed standardized Excel spreadsheet to extract key information for inclusion in the literature, including: first author, publication year, study type, sample source (species, AGA region, control region), fibroblast subtypes (such as DPC, dermal fibroblasts, hair sheath fibroblasts), intervention/exposure conditions (DHT/testosterone concentration, treatment time), outcome measures (specific detection methods, quantitative results, effect direction), and bias assessment information. The data extraction is independently completed and cross checked by two reviewers.
2.6. Risk of bias assessment
The Newcastle Ottawa Scale (NOS) was used to evaluate human tissue research; Animal experiments were conducted using the SYRCLE bias risk tool; In vitro cell studies evaluate quality based on custom criteria such as passage number, androgen treatment regimen, and control settings.
2.7. Effect measures and synthesis methods
Meta analysis was not conducted due to significant heterogeneity in cell sources, processing conditions, and outcome indicators included in the study. Mainly using narrative integration, the evidence is summarized in layers according to core mechanism themes (signal transduction, fibrosis, aging, regenerative disorders, treatment strategies). Summarize the trends of multiple studies with consistent effect directions and discuss conflicting results.
3. Results
3.1. Study selection
The PRISMA flow diagram (Figure 1) summarizes the study selection process. The database search yielded a total of 2361 records. After removing duplicates, 1592 records were screened by title and abstract. Firstly, 37 non-English literatures were excluded, followed by 209 literature that did not fit the research type (including 97 reviews, 31 comments, 45 conference abstracts, and 36 case reports). Among the remaining literature, 620 studies with mismatched cell types were excluded (including 509 using fibroblasts from non-hair follicle sources and 111 not specifically using dermal papilla cells or dermal sheath cells), followed by 412 studies without androgen intervention (not using DHT, testosterone, or 5α-reductase activity), and finally 271 studies that did not report functional outcomes of dermal papilla cells (such as proliferation/apoptosis, signaling pathway activity, gene/protein expression, secretion factor levels, cell aggregation ability, etc.) were excluded. Finally, a total of 43 original studies (including in vitro cell experiments, animal experiments, and clinical sample studies) met all inclusion criteria and entered the meta-analysis. PRISMA flow diagram.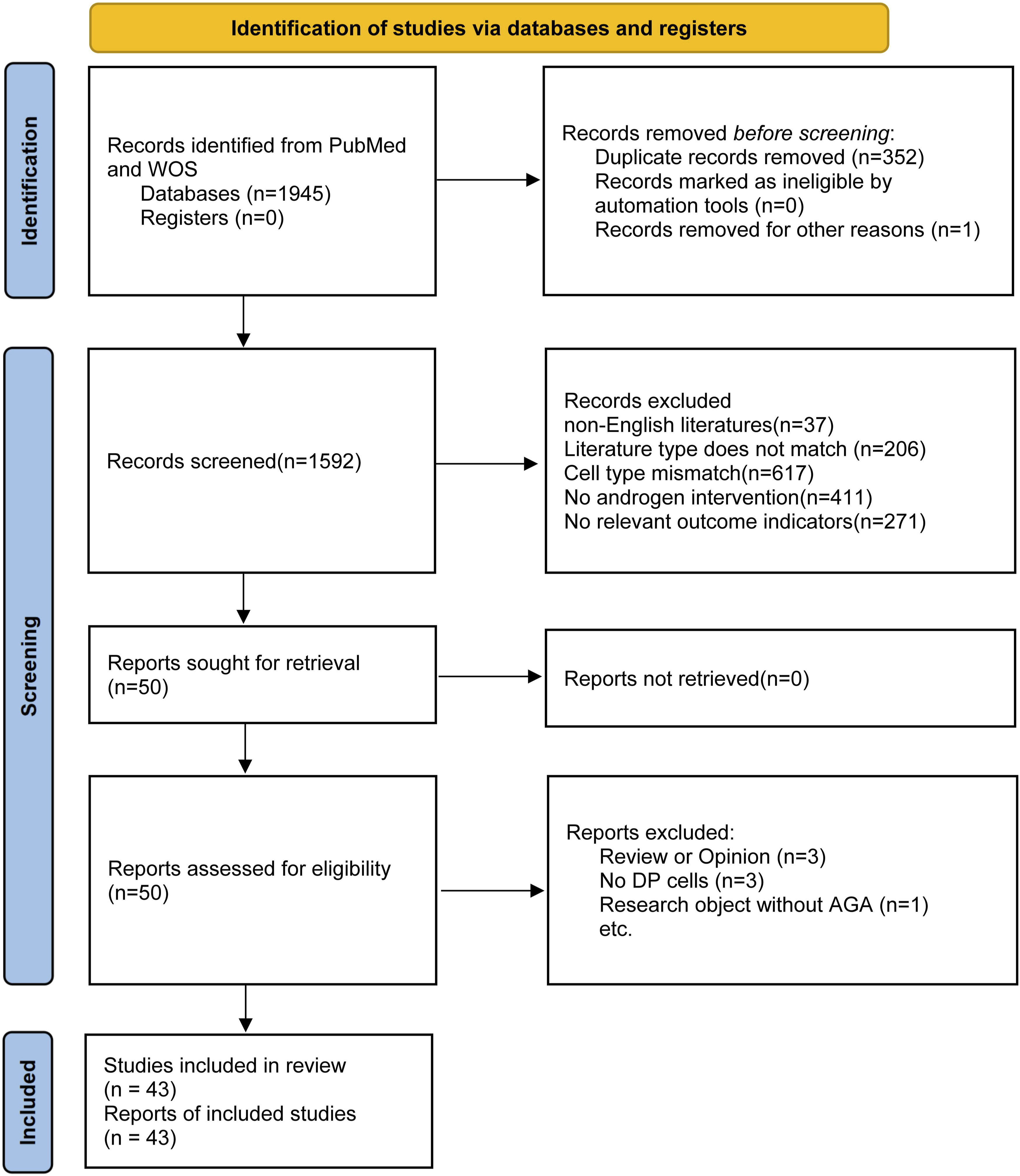
3.2. Characteristics of included studies
The 43 original studies included mainly focused on in vitro cell experiments (65.1%), with some studies combined with animal experiments or clinical sample validation; All studies used hair follicle DP cells or dermal sheath cells from human or animal sources, with human primary DP cells being the most commonly used (accounting for 67%). Androgen intervention is mainly based on DHT (81%), with a concentration range of 0.1 nM to 100 μ M, and a few studies have used testosterone or 5α-reductase inhibitors. The outcome measures include cell proliferation/viability (65%), apoptosis (35%), signal pathway protein expression (51%), gene expression (44%), and in vivo hair growth parameters (28%); The analysis of signaling pathways mainly focuses on Wnt/β-catenin (37%), TGF-β/Smad (23%), PI3K/Akt (19%), and MAPK/ERK (16%). About 35% of the literature provides extractable data such as mean and standard deviation in the abstract. Overall, these studies systematically explored the regulatory effects of DHT and androgen signaling on DP cell function, providing sufficient experimental evidence for subsequent meta-analyses.
3.3. Fibroblasts
3.3.1 Unique properties of DP cells
DP cells have unique biological characteristics that make them the “command center” of hair follicles. 37 Firstly, they exhibit characteristics of mesenchymal stem cell, including self-renewal and pluripotency. 38 This inherent potential is activated during the transition from dormancy to growth, enabling DP cells to drive epithelial stem cell proliferation and differentiation, thereby initiating a new growth cycle.19,39
Secondly, DP cells act as a highly active secretion centers, producing large number of key signaling molecules such as VEGF, FGF7, FGF10, IGF-1, BMPs, Wnt ligands, and extracellular matrix components.20,40–43 These molecules form a precise paracrine network that regulates the behavior of epithelial cell, ultimately determining the growth rate, shaft thickness, and cycle progression of hair. The subtle changes in this secretion pattern can shift follicle from growth to miniaturization.17,19,39,44
Thirdly, DP cells exhibit unique spontaneous aggregation behavior in vitro. These three-dimensional spheres reflects their functional integrity and is crucial for maintaining their hair inducing ability.23,45–49 In AGA, this aggregation ability has been severely compromised; DP cells become dispersed, leading to functional decline, secretion imbalance, and inability to maintain a robust growth phase, which is the core event of miniaturization. Therefore, aggregation characteristics are both functional markers and key indicators for evaluating the physiological and therapeutic status of DP cells. 50
3.3.2. Functions of perifollicular fibroblasts
The DS cells that encapsulate follicle provides crucial structural and mechanical support.36,51–53 In addition to this static function, DS is also a dynamic cell bank with multiple functions. It is an important source of signaling molecules that communicate with epithelial cells and the DP.54,55
Its most critical function is to serve as a reservoir for DP cells. During the transition from telogen to anagen, specific DS cells are activated, proliferated, migrated, and aggregated at the nascent hair bulb to participate in forming a new, functional DP—a essential process for successful follicular regeneration.56,57 Therefore, the activities and health statue of the DS are crucial for supplementing DP.
In AGA, the functional decline of the DS is significant. DS cells may be indirectly affected by dysregulated androgen signaling, thereby affecting their efficacy as cellular reservoirs. 58 This damage may lead to insufficient supply of healthy progenitor cells for DP. exacerbating DP dysfunction and accelerating follicular miniaturization. Therefore, DS and DP form a collaborative functional unit, and the interruption of their interaction is a key driving force for the progress of AGA.
3.4. Fibroblasts and androgen signal transduction
3.4.1. Expression of AR
The pathogenesis of AGA revolves around the abnormal response of hair follicles to androgens, exhibiting significant cellular specificity.59,60 Numerous studies have confirmed that DP cells are the main direct target of androgen action. 61 The key evidence for this conclusion lies in the cellular distribution pattern of the AR: DP cells consistently exhibit high levels of AR expression, while the surrounding hair follicle epithelial cells (such as hair stromal cells and outer root sheath keratinocytes) show significantly lower level of AR expression.62,63
This differential distribution of AR expression determines the pathway of androgen signaling. Circulating androgens, mainly testosterone, penetrate the capillary network around follicles and are absorbed by DP cells. 64 In these cells, the highly expressed 5α-reductase type II enzyme catalyzes the conversion of testosterone to the more potent dihydrotestosterone (DHT). Subsequently, DHT binds with high affinity to the extensively expressed AR in the nucleus, forming a DHT-AR complex.65,66 This complex undergoes conformational changes, dimerizes, and binds to specific androgen response elements (AREs) in nuclear DNA, thereby initiating or repressing the transcription of downstream target genes.67,68
Therefore, the DP unit serve as a “signal transducer”. They receive DHT and convert it into a local cellular responses, specifically, an alteration of their secretory profile—which in turn regulates the behavior of the entire follicular unit. In contrast, epithelial cells exhibit the least direct response to androgens due to their low AR expression levels. Their behavioral changes are largely indirectly driven by changes in the secretion factors of DP cells. This highly specific pattern of AR expression establishes the molecular foundation for DP cells to act as trigger and signal amplifier in AGA pathophysiology. It also explains why follicular miniaturization is an orderly process driven primarily by dysfunction of the mesenchymal “command center”.
3.4.2. Response of DP cells following androgen activation
After DHT binds to AR highly expressed in the nucleus of DP cells, the DHT-AR complex produced acts as a key transcriptional regulator, triggering a series of profound cellular responses and ultimately disrupting the steady-state balance of the hair follicle microenvironment. 66 This process begins with a global change in gene expression profiles within the cell nucleus. The DHT-AR complex translocate into the nucleus and specifically binds to the androgen responsive AREs located in the promoter region of target gene. Its function is similar to a “molecular key”, directly initiating or inhibiting the transcription of a series of key genes, thereby reshaping the biological function of DP cells.
The core consequence of this transcriptional reprogramming is the disruption of the balance between signaling pathways that determine the fate of hair follicle. Under steady-state conditions, the growth promoting (e.g., Wnt/β-catenin) and anti-growth (e.g., TGF-β, BMP) pathways are in dynamic balance, maintaining a normal follicular cycle.20,69 However, sustained androgen activation can severely disrupt this balance. The DHT-AR signaling pathway upregulates multiple anti-growth pathways: it effectively activates the TGF-β1 and BMP pathways, which can drive the follicle into the catagen phase prematurely. 20 At the same time, it inhibits or antagonizes key growth promoting pathways, most notably by inhibiting the activity of the Wnt/β-catenin pathway—the primary driver for initiating and sustaining the anagen phase.70,71
The ultimate and most direct manifestation of these changes in gene expression and signaling pathways is the pathological changes in the secretion profile of DP cells. Dysfunctional DP cells transition from a “pro-growth center” into a “growth inhibiting signal sources”. The net effect of its secreted factors shifts from promoting to inhibiting growth: on one hand, the expression of key factors that support papilla vascularization and hair matrix cell proliferation—such as VEGF and IGF-1—is reduced; on the other hand, the secretion of hair growth-inhibiting factors, such as TGF-β1 and DKK-1 (a potent Wnt signaling inhibitor), is significantly increased. This altered secretory profile acts via paracrine signaling on surrounding epithelial cells, directly resulting in a shortened anagen phase, impaired hair matrix cell proliferation, and ultimately driving the follicle toward irreversible miniaturization. Therefore, the response of DP cells following androgen activation fundamentally represents a pathological transformation of their functional identity, serving as the molecular initiation point for the process of follicular miniaturization.
3.5. Mechanism of follicular miniaturization induced by fibroblast dysfunction
3.5.1. Imbalance in signal transduction and disruption of intercellular communication
The homeostasis of hair follicle depends on a fine dynamic balance between pro-growth and anti-growth signaling pathways. In AGA, dysfunctional DP cells disrupt this balance, tilting the follicular growth program toward suppression and thereby initiating the miniaturization process. 14 The imbalance in signal transduction represents the core molecular mechanism underlying follicular miniaturization.
The fundamental reversal of the signaling pathway is the molecular core of this process. Dysfunctional DP cells undergo pathological changes in their secretion profile, transitioning from a “growth promoting center” to a “growth inhibiting signaling source”. This manifests as a synergistic “riple blow”: firstly, the Wnt/β-catenin pathway, which is crucial for initiating and maintaining growth, is significantly inhibited by the secretion of antagonists such as DKK-1. 72 Secondly, growth inhibitory signaling pathways such as TGF-β1 and BMP are continuously activated and amplified.20,73 Among them, TGF-β1 can not only directly induce apoptosis of hair germ cells and drive hair follicles to enter the degenerative phase prematurely, but also act as a key “stop signal” to forcibly shorten the growth phase. The imbalance between these pathways fundamentally reverses the microenvironmental signals of hair follicles.
The imbalance of this signal directly leads to the collapse of the intercellular communication network. On the one hand, the instruction transmission between DP cells and epithelial cells is ineffective: due to the weakening of pro proliferative signals (such as FGF7) and the enhancement of pro apoptotic signals (such as TGF-β1), the proliferation of dermal cells is impaired, and stem cells in the bulge region remain quiescent due to the inability to receive correct activation instructions.21,74,75 The communication failure between the “command center” and the “execution unit” directly disrupted the regenerative ability of hair follicles. On the other hand, the connection between hair follicles and external support systems is severed: DP cells secrete factors such as VEGF to maintain the ability of the surrounding capillary network, leading to insufficient blood supply to the hair follicles; Its signal communication with peripheral nerve endings is also impaired. This puts hair follicles in an unfavorable microenvironment of “malnutrition” and “signal islands”, further exacerbating growth disorders.
Therefore, the dysfunction of DP cells promotes the irreversible miniaturization of hair follicles through the imbalance of internal signaling pathways and the breakdown of external communication support systems.
3.5.2. Destruction of the stem cell niche
The regenerative ability of the hair follicle comes from their powerful stem cell system. Hair follicle stem cells (HFSCs) are located in the bulge region and serve as a reservoir for periodic hair follicle regeneration. 76 However, a key finding is that HFSCs themselves are insensitive to androgens because their surface expressed AR levels are extremely low. This indicates that androgens cannot directly instruct stem cell activation. On the contrary, the behavior of HFSCs is primarily governed by their microenvironment—specifically, the signals originating from DP cells. Consequently, in AGA, the dysfunction of DP cells directly leads to the disruption of the stem cell niche, thereby severing the fundamental driver of hair follicle regeneration.
In a healthy hair follicle, as a new anagen phase initiates, functionally competent DP cells send a series of potent activation signals to the bulge region. The Wnt/β-catenin pathway acts as the “starter switch” in this process. Concurrently, signals that maintain stem cell quiescence, such as BMP signaling, are correspondingly downregulated. 77 This precisely timed combination of signals functions like an accurate “departure command”, prompting HFSCs to migrate downward, proliferate, and differentiate into new follicular components, ultimately forming a thick terminal hair.
In the pathological state of AGA, this activation mechanism fails completely. As DP cells are the primary target of androgen action, their functional disruption leads to a fundamental alteration in their secretory profile. On one hand, the secretion of key activating signals is reduced, while the expression of Wnt pathway inhibitors increases, resulting in a weakened or even absent “starter switch” signal. On the other hand, the signaling environment that suppresses stem cell activity may be reinforced. The ultimate consequence is that DP cells become incapable of delivering an effective and correct activation command to HFSCs. 78 Without receiving the crucial “departure” signal, the HFSCs are forced into a prolonged state of quiescence and cannot be recruited to the new hair bulb to participate in hair shaft formation.
The direct consequence of this failed stem cell activation is the “congenital deficiency” of the newly formed hair follicle. Due to a reduced available progenitor cell pool and weakened downstream proliferative and differentiation drive, the follicular structure rebuilt with each subsequent cycle becomes progressively smaller and more shallow. Consequently, the hair shaft gradually thins, transitioning from terminal hair to vellus hair. This indicates that follicular miniaturization does not stem from the depletion of stem cells, but rather from the “silencing” of stem cells caused by DP cells dysfunction. Therefore, restoring DP cells function and rebuilding a healthy stem cell niche represent crucial strategic objectives for reversing follicular miniaturization and restoring the hair follicle’s regenerative capacity.
3.6. Inflammation and fibrosis
The process of follicular miniaturization is not confined solely to cell-autonomous dysfunction but is accompanied by pathological remodeling of its surrounding microenvironment. 79 Among these changes, chronic inflammation and subsequent fibrosis constitute critical pathological features in the advanced stages of AGA. This process acts like a “sturdy cage” encasing the hair follicle, ultimately leading to its complete structural destruction.
Dysfunctional DP cells once again act as the primary driver in this process. Under sustained androgen stimulation, activated DP cells secrete a range of pro-inflammatory factors and chemokines, such as Interleukin-6 (IL-6) and Monocyte Chemoattractant Protein-1 (MCP-1). These signaling molecules recruit immune cells, including macrophages and T-lymphocytes, to the perifollicular region, triggering a mild, chronic, and subclinical inflammatory infiltrate. 80 Although this inflammatory response is less acute than classical folliculitis, it operates like a chronic, “simmering at a low heat”, continuously releasing pro-inflammatory and pro-fibrotic signals. This persistent assault causes chronic damage to hair follicle stem cells and proliferative cells, further disrupting the normal hair follicle cycle.
The persistent inflammatory response ultimately initiates a tissue repair/fibrosis program. In this process, the excessively secreted TGF-β1 by DP cells plays a central role. TGF-β1 is not only a potent growth-inhibitory factor but also one of the most powerful pro-fibrotic factors. It activates the perifollicular fibroblasts, causing them to transform into myofibroblasts and massively synthesize and deposit extracellular matrix components, primarily collagen types I and III. 81 The normally loose and elastic perifollicular connective tissue is gradually replaced by rigid, dense fibrous tissue.
Thus, inflammation and fibrosis represent the terminal stage of a vicious cycle: dysfunctional DP cells trigger inflammation, which drives fibrosis, and the resulting fibrosis physically strangles the hair follicle. This process clarifies that AGA is not merely an endocrine disorder, but also an inflammatory and fibrotic disease characterized by specific microenvironmental alterations. This understanding provides a theoretical foundation for developing anti-inflammatory and anti-fibrotic treatment strategies. Figure 2 illustrates the central role of fibroblasts in androgenetic alopecia. The central role of fibroblasts in androgenetic alopecia. (Drawn by the authors using PowerPoint).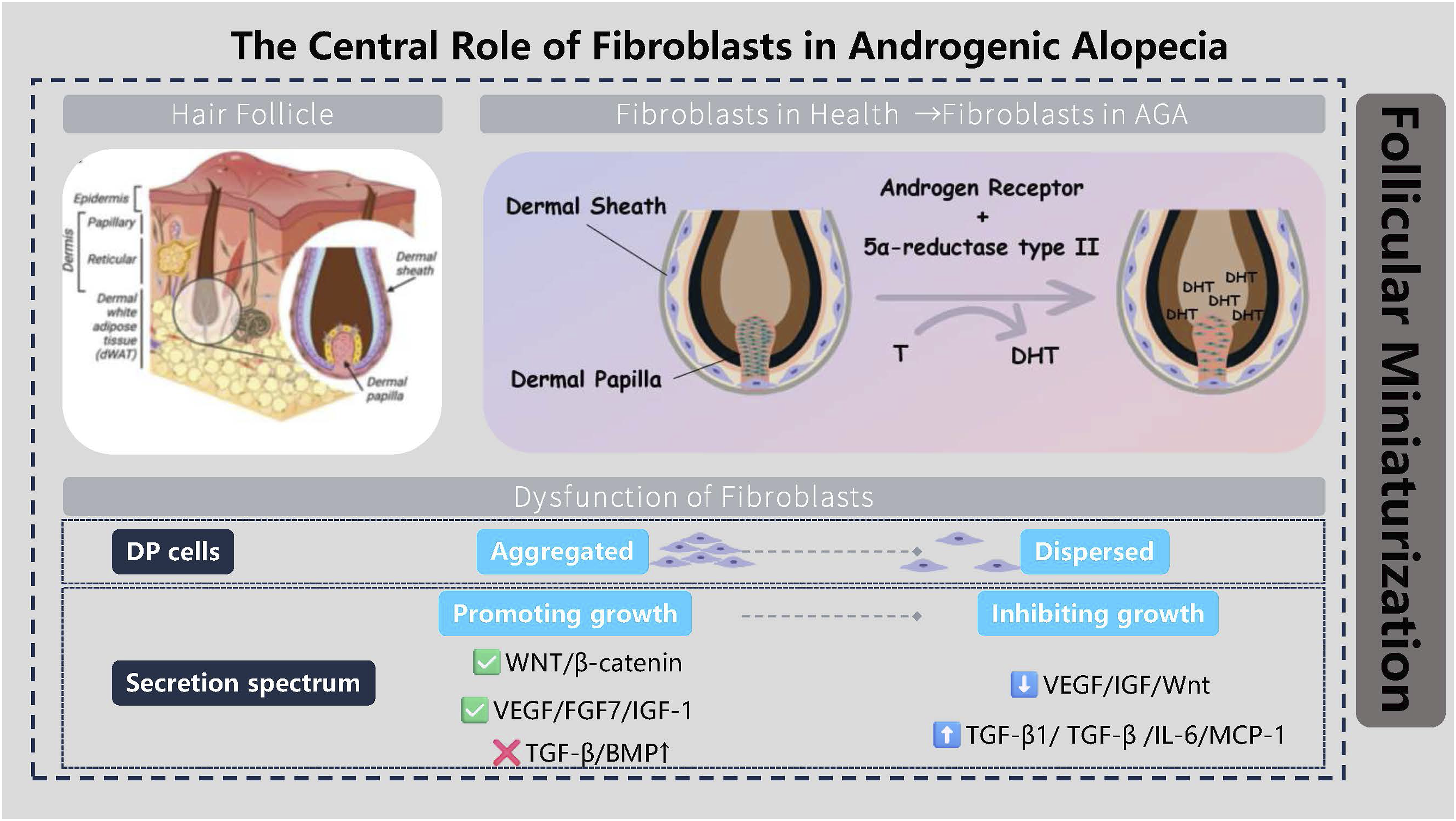
3.7. Therapeutic strategies targeting fibroblasts and future directions
3.7.1. Indirect effects of existing treatments
The first-line medications currently widely used in clinical practice for AGA primarily exert their therapeutic effects by modulating or influencing the functional state of DP cells, although their mechanisms of action do not directly target fibroblasts. These therapies indirectly intervene in the abnormal activation of DP cells through different pathways, thereby slowing the progression of follicular miniaturization.
Finasteride and dutasteride, as specific inhibitors of 5α-reductase, operate by systemically reducing levels of DHT in the scalp and circulation. 3 By inhibiting the key enzyme responsible for converting testosterone into the highly active DHT, these drugs effectively interrupt the pathological signaling source that chronically stimulates DP cells. When the concentration of DHT in the DP cells microenvironment decreases significantly, the overactivated AR signaling pathway is “unburdened”. This leads to beneficial adjustments in the gene expression and secretory profiles of DP cells—for instance, potentially reducing the expression of pro-apoptotic factors like TGF-β1—thereby normalizing the hair growth cycle. Consequently, these medications essentially work by removing the abnormal stimulus, creating an “unburdened” or “resting” microenvironment for the dysfunctional DP cells and indirectly restoring some of their normal function. However, this therapy cannot reverse pre-existing DP cells functional decline or perifollicular fibrosis, and the therapeutic effects diminish upon discontinuation as the stimulatory source returns.
In contrast, minoxidil’s mechanism of action more directly targets the local microenvironment. 3 Although its precise molecular targets are not fully elucidated, substantial evidence suggests it acts as a potassium channel opener and vasodilator, directly or indirectly influencing DP cells. Studies indicate that minoxidil promotes VEGF expression in DP cells, significantly improving blood supply to the follicular bulb and thereby providing enhanced nutrition and oxygen to the follicle. Concurrently, it likely exerts effects through multiple pathways: prolonging epithelial cell survival, exerting mild anti-inflammatory effects, and directly stimulating DP cells proliferation. Thus, minoxidil can be regarded as a “functional enhancer” for DP cells or a microenvironment modulator. By creating more favorable growth conditions, it partially counteracts the negative signals induced by androgens, thereby helping maintain the hair follicle in a healthier state.
In summary, the strategy of current mainstream pharmacological treatments for AGA—whether systemically removing the pathogenic signal (finasteride/dutasteride) or locally improving the microenvironment (minoxidil)—ultimately converges on correcting the functional imbalance of DP cells. This clinically confirms the central role of DP cells in AGA pathogenesis and simultaneously highlights the limitations of current therapies: they are primarily indirect and palliative, rather than directly repairing or replacing functionally compromised DP cells. This very limitation defines a clear direction and creates an urgent need for next-generation therapeutic strategies that target the fibroblasts themselves.
3.7.2. Emerging research directions and therapies
Building on the profound understanding of the central role of fibroblasts—particularly DP cells—in AGA, the research focus is shifting from traditional hormonal interventions toward innovative strategies that directly target and rejuvenate these critical cells. These emerging therapeutic approaches aim to fundamentally rebuild a healthy follicular microenvironment, offering new promise for the treatment of AGA.
Cell therapy represents one of the most promising directions.49,82 Its core concept involves “replacing” or “replenishing” functionally compromised DP cells. Researchers isolate healthy DP cells from a small number of donor hairs and expand them via 3D culture in vitro to restore their inherent high aggregative behavior and hair-inductive capability. These cultured, healthy DP cells are then transplanted into the balding scalp. It is expected that the transplanted healthy DP cells can establish a functional “signaling command center” in vivo. By secreting the correct network of factors, they can reactivate resident epithelial stem cells within the follicle, thereby inducing de novo hair follicle formation or reversing the miniaturization of existing follicles. Similarly, strategies leveraging DS cells—exploiting their inherent role as a reservoir for DP cells—are also under investigation for transplantation purposes.
Compared to whole-cell transplantation, exosome therapy represents a more refined and safer “cell-free” strategy. 83 Exosomes are nanoscale vesicles secreted by cells, carrying bioactive substances from their parent cells such as proteins, messenger RNAs, and microRNAs. Exosomes derived from DP cells are believed to encapsulate the key signaling instructions that enable DP cells to induce hair regeneration. The extraction, purification, and topical application of these exosomes can mimic the function of healthy DP cells by delivering pro-growth signals (e.g., activating the Wnt pathway) to downstream target cells, while simultaneously circumventing potential issues associated with live cell transplantation, such as immune rejection and ethical concerns.
At the molecular level, precisely targeting key signaling pathways within DP cells is a major focus of drug development. For instance, developing small-molecule agonists to specifically activate the Wnt/β-catenin pathway in DP cells, or employing antagonists to suppress the overactive TGF-β pathway, aims to directly modulate the “signaling switches” responsible for follicular miniaturization and restore pathway homeostasis. This strategy demands that the therapeutic agents exhibit high targeting precision and specificity to minimize off-target effects on other tissues and cells.
More forward-looking tissue engineering strategies aim to “de novo construct” hair follicles in vitro. By co-culturing highly viable DP cells, epidermal stem cells, and other components in a three-dimensional system using various biocompatible scaffold materials, researchers have successfully generated organoids or miniature hair follicles in the laboratory capable of producing hair fibers. Although this technology remains in its early stages and faces significant challenges—such as achieving synchronized hair cycling and controlling growth orientation—it provides the ultimate blueprint for achieving true “hair follicle regeneration” and transplantation.
Finally, targeting the pathological endpoint of perifollicular fibrosis in advanced AGA, anti-fibrotic therapy represents an important adjuvant strategy. Exploring the use of approved or novel anti-fibrotic agents—such as monoclonal antibodies or small-molecule drugs targeting TGF-β signaling—aims to degrade the excessive extracellular matrix and alleviate the physical constriction of hair follicles, thereby creating a more permissive microenvironment for follicular regeneration. This strategy could potentially be combined with other regenerative therapies to establish conditions for restoring vitality to severely fibrotic areas of alopecia.
3.7.3. Challenges of drug delivery systems
The development of novel therapies targeting hair follicle fibroblasts highly relies on efficient drug delivery systems for their efficacy. The core challenge currently faced is how to effectively penetrate the skin barrier with therapeutic drugs (such as small molecules, growth factors, nucleic acids, or cell products), accurately target and enrich the DP cells area deep in the hair follicle, while maintaining a sufficient duration of action locally to reshape the hair follicle microenvironment. The penetration efficiency, targeting specificity, controllable release ability, and feasibility of production of delivery systems are key bottlenecks that determine the successful translation of many cutting-edge therapies.
4. Conclusion
In summary, fibroblasts, especially dermal papilla cells, play a central role in the pathogenesis of androgenic alopecia by regulating androgen signaling transduction, paracrine signaling networks, stem cell microenvironment, and intercellular communication. These emerging therapies targeting fibroblasts—spanning cellular, exosomal, signaling pathway, and tissue engineering approaches—form a multi-pronged research spectrum aimed at fundamentally restoring hair follicle function. Although existing treatment strategies can indirectly regulate its function, their effectiveness is limited. The breakthroughs in future research directions will heavily rely on the application of new technologies. Single cell sequencing technology can reveal the heterogeneity of different cell subpopulations within DP cells and their surrounding microenvironments with unprecedented resolution, accurately identify specific cell states and their characteristic molecular labels that play a key role in disease occurrence, and thus discover new therapeutic targets. At the same time, gene editing technologies such as CRISPR-Cas9 have shown the potential to directly correct pathogenic signaling pathway abnormalities in DP cells, such as precise regulation of the expression or function of key genes such as AR and Wnt/β-catenin, providing a revolutionary strategy for fundamentally reversing hair follicle miniaturization. These cutting-edge technologies, along with cell therapy, exosomes, and tissue engineering mentioned in the article, together form a multi-level future treatment blueprint aimed at fundamentally restoring hair follicle function. They not only demonstrate significant therapeutic potential but have also further reinforced the central role of fibroblasts in both the pathogenesis of AGA and the exploration of novel treatments.
Footnotes
Acknowledgments
We are grateful to Nanjing Medical University for their financial support.
Ethical considerations
Ethics approval is not required for this literature review as it does not involve new human or animal subjects.
Author contributions
Conceptualization: Shanshan Yu; Methodology: Jie Ji; Formal analysis and investigation: Yingying Wu, Jingyun Li; Writing - original draft preparation: Shanshan Yu, Yingying Wu; Writing - review and editing: Hui Yan, Yongfang Xu, Yutong Shi; Funding acquisition: Shanshan Yu; Supervision: Jun Li.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported financially by General Project of Science and Technology Development Fund of Nanjing Medical University (NMUB20240092).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
No new data were generated or analyzed in this study. All information is drawn from cited literature and references listed at the end of the manuscript.
Trial registration
DOI: 10.17605/OSF.IO/GVK5Q.