
Abstract
Stromal vascular fraction (SVF)-based therapies and autologous fat grafting have emerged as promising regenerative strategies due to their pro-angiogenic, immunomodulatory, and trophic properties. However, despite encouraging preclinical and clinical findings, therapeutic outcomes remain highly heterogeneous, with marked variability in graft retention and functional efficacy between patients. Increasing evidence suggests that this variability cannot be explained solely by procedural factors or cellular composition, but may also depend on host-related immune and microenvironmental determinants. This review explores the biological mechanisms governing SVF engraftment and introduces the emerging concept of “SVF therapy resistance,” defined as the failure of autologous regenerative therapies resulting from maladaptive interactions between transplanted stromal cells and the host tissue environment. Particular attention is given to sterile inflammation, innate immune activation, and early graft–host interactions. Following transplantation, tissue injury and ischemia induce the release of danger-associated molecular patterns (DAMPs), triggering neutrophil recruitment, macrophage activation, complement signaling, and inflammatory remodeling. While controlled inflammatory responses may support tissue repair and angiogenesis, excessive neutrophil activation, neutrophil extracellular trap (NET) formation, persistent pro-inflammatory macrophage polarization, and impaired vascular adaptation may compromise graft survival and regenerative efficacy. The review further discusses how SVF processing, inflammatory priming, stromal cell heterogeneity, and donor-related factors—including obesity, aging, metabolic dysfunction, and chronic inflammation—may influence therapeutic responsiveness. Emerging evidence from mesenchymal stromal cell biology suggests that stromal cells are highly sensitive to inflammatory licensing and microenvironmental cues. Candidate biomarkers and immune profiling strategies capable of identifying responders and non-responders to SVF-based therapies are also reviewed. Finally, these mechanisms are discussed in spinal cord injury, a condition characterized by chronic inflammation and vascular dysfunction. Overall, this review proposes a translational framework linking innate immunity, sterile inflammation, angiogenesis, and stromal cell heterogeneity to the variability of SVF therapy outcomes, highlighting the need for personalized regenerative medicine approaches.
Keywords
• Host innate immunity critically influences SVF graft survival and regenerative outcomes. • Neutrophil activation and NET formation may impair early graft integration and vascularization. • Macrophage polarization balance appears central to successful tissue remodeling after SVF therapy. • Hypoxia and angiogenic adaptation are key determinants of long-term graft retention. • The concept of “SVF therapy resistance” is proposed to explain inter-individual therapeutic variability. • Immune profiling and biomarker-based stratification may support personalized regenerative medicine approaches. • Spinal cord injury represents a high-risk inflammatory microenvironment for SVF therapeutic failure.Highlights
Introduction
Autologous fat grafting and stromal vascular fraction (SVF)-based therapies have emerged as promising approaches in regenerative medicine, with applications ranging from soft tissue reconstruction to the treatment of chronic wounds and neurological disorders. The regenerative potential of SVF relies on a heterogeneous cellular population including adipose-derived stromal cells, endothelial progenitors, pericytes, and immune cells, which collectively contribute to tissue repair through paracrine signaling, angiogenesis, immunomodulation, and trophic support.1–4 More broadly, mesenchymal stromal cells have been extensively investigated as therapeutic tools in regenerative medicine due to their immunomodulatory and reparative properties.5,6
Despite encouraging preclinical and clinical results, a major limitation of SVF-based therapies remains the substantial variability in outcomes, with inconsistent graft retention and heterogeneous therapeutic efficacy observed across patients.7,8 While technical factors and procedural variability may partially account for these discrepancies, growing evidence suggests that host-related biological determinants play a central role in shaping regenerative outcomes. Increasing attention has therefore been directed toward the interactions between transplanted stromal cells and the host immune microenvironment.
Recent advances in mesenchymal stromal cell biology have demonstrated that regenerative efficacy is highly dependent on the surrounding inflammatory context. Rather than acting as constitutively active therapeutic agents, mesenchymal stromal cells appear to require functional “licensing” by inflammatory signals to exert their immunomodulatory and reparative functions.9,10 In this framework, the host immune environment is not merely a passive recipient of cell therapy but an active determinant of therapeutic efficacy. Systemic inflammation, metabolic dysfunction, and obesity-related immune alterations may profoundly influence stromal cell behavior and tissue integration.11,12
Among the host-related factors potentially influencing graft fate, innate immunity has emerged as a critical early regulator of engraftment. Following transplantation, neutrophils are rapidly recruited to the graft site where they participate in debris clearance and inflammatory signaling. However, excessive neutrophil activation may impair tissue integration through the release of reactive oxygen species (ROS), proteolytic enzymes, and neutrophil extracellular traps (NETs).13–15 Beyond their antimicrobial role, NETs have increasingly been implicated in sterile inflammation, endothelial dysfunction, thrombosis, and impaired tissue repair, potentially compromising early graft vascularization and survival.16,17
Subsequently, macrophages orchestrate the resolution or persistence of inflammation and play a pivotal role in tissue remodeling after fat grafting. While pro-inflammatory M1 macrophages are associated with tissue degradation and graft resorption, alternatively activated M2 macrophages support angiogenesis, adipogenesis, extracellular matrix remodeling, and regenerative integration.18,19 Importantly, macrophage polarization is strongly influenced by systemic metabolic and inflammatory conditions. Obesity and metabolic dysfunction have notably been associated with persistent pro-inflammatory macrophage activation, potentially creating a microenvironment less permissive to regenerative processes.11,20,21
In parallel, hypoxia and angiogenesis constitute central determinants of graft survival. Following transplantation, adipose tissue is immediately exposed to ischemic stress due to disruption of its native vascular network. 22 While transient hypoxia may activate adaptive and pro-angiogenic pathways through hypoxia-inducible factor signaling, prolonged or dysregulated hypoxia can exacerbate inflammation, adipocyte death, fibrosis, and graft resorption.23–25 The efficiency of neovascularization therefore appears to represent a critical step determining long-term graft retention.
Importantly, variability in regenerative outcomes may also arise from intrinsic heterogeneity within SVF cellular populations themselves. Adipose-derived stromal cells exhibit substantial differences in proliferative capacity, immunomodulatory properties, and angiogenic potential depending on donor characteristics and tissue origin.26–28 Such heterogeneity may further interact with patient-specific inflammatory and metabolic profiles, contributing to the existence of “responders” and “non-responders” to regenerative therapies.
These challenges may be particularly relevant in spinal cord injury (SCI), a condition characterized by persistent inflammation, microvascular dysfunction, oxidative stress, and a chronically hostile regenerative microenvironment. 29 Although SVF-based approaches have demonstrated promising neuroprotective and regenerative potential in experimental SCI models,30,31 therapeutic efficacy may remain highly dependent on host immune and vascular conditions. In this context, regenerative medicine increasingly converges with precision medicine approaches aimed at integrating patient-specific biological characteristics into therapeutic decision-making. 32
In this context, we propose the concept of “SVF therapy resistance,” defined as the failure of autologous regenerative therapies resulting from maladaptive interactions between transplanted SVF cellular populations and the host immune–microenvironmental landscape. This review aims to synthesize current evidence linking innate immunity, angiogenesis, hypoxia, and stromal cell heterogeneity to regenerative outcomes, while exploring the emerging implications of immune profiling and precision regenerative medicine in SVF-based therapies.
Materials & methods
Search strategy
A structured literature search was conducted to identify studies investigating SVF-based therapies, fat grafting, innate immune responses, and factors influencing graft survival and regenerative outcomes. Electronic databases including PubMed, Scopus, and Web of Science were systematically searched for articles published in English up to March 2026.
Search terms combined keywords related to adipose-derived regenerative therapies and host immune mechanisms, including: “stromal vascular fraction”, “fat grafting”, “adipose-derived stem/stromal cells”, “innate immunity”, “neutrophils”, “macrophages”, “NETs”, “angiogenesis”, “hypoxia”, “inflammation”, and “regenerative medicine”. Boolean operators (AND, OR) were used to refine the search strategy.
Additional references were identified through manual screening of reference lists from relevant articles and reviews.
Study selection
Titles and abstracts were screened according to their relevance to the objectives of the review. Full-text articles were subsequently assessed for eligibility based on predefined inclusion and exclusion criteria.
Priority was given to studies exploring: • mechanisms of host–graft interaction, • innate immune responses following fat grafting or SVF administration, • angiogenic and hypoxic determinants of graft survival, • mesenchymal stromal cell heterogeneity and immunomodulation, • and translational implications for regenerative medicine and SCI.
Both preclinical and clinical studies were considered. The overall literature search and study selection workflow is presented in Figure 1. Study selection workflow for the narrative mechanistic review.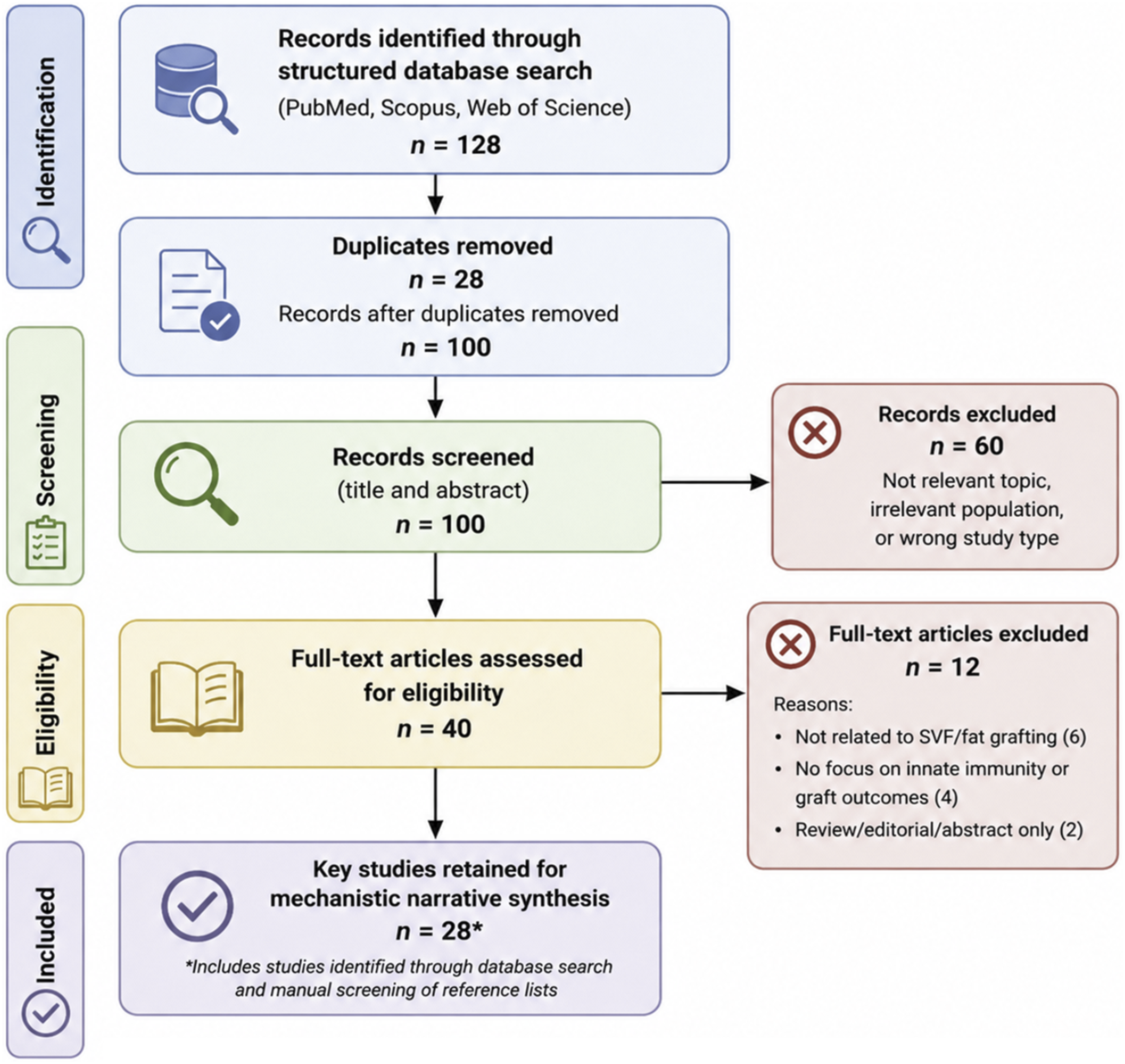
Inclusion and exclusion criteria
Studies were included if they: • investigated SVF, adipose-derived stromal/stem cells, or fat grafting; • explored mechanisms related to innate immunity, inflammation, angiogenesis, or tissue remodeling; • addressed factors influencing graft survival or regenerative efficacy; • included experimental, translational, or clinical data relevant to regenerative medicine.
Studies were excluded if they: • were unrelated to adipose-derived regenerative therapies; • focused exclusively on non-regenerative or unrelated biological contexts; • consisted solely of conference abstracts, editorials, or non-peer-reviewed material.
Data extraction and synthesis
Data were extracted qualitatively with a focus on mechanistic insights related to: • early inflammatory responses, • neutrophil and macrophage dynamics, • NET formation, • angiogenesis and hypoxia, • stromal cell heterogeneity, • and host-related determinants of therapeutic variability.
Given the heterogeneity of study designs and reported outcomes, a narrative synthesis approach was adopted. The literature was organized into thematic sections to support the development of the proposed concept of “SVF therapy resistance”.
Reporting approach
This work was designed as a narrative and hypothesis-generating review integrating experimental, translational, and clinical evidence related to host determinants of SVF therapy outcomes.
Review
Conceptual framework of SVF therapy failure
From graft failure to regenerative failure
Despite the growing clinical use of SVF-based therapies, therapeutic outcomes remain highly heterogeneous across patients and indications. Traditionally, unsuccessful outcomes have been interpreted primarily through a technical perspective, including insufficient graft retention, inadequate harvesting procedures, poor vascularization, or suboptimal cell processing methods.3,7,9 However, accumulating evidence suggests that regenerative failure cannot be explained solely by procedural variability and likely reflects a more complex interaction between the graft, the host microenvironment, and the biological properties of the SVF product itself.
Importantly, autologous SVF therapies differ fundamentally from conventional transplantation paradigms. Because the graft originates from the patient, classical alloimmune rejection mechanisms are largely absent. Nevertheless, transplanted tissues and cells remain exposed to intense inflammatory, ischemic, and metabolic stresses immediately after implantation. The success of engraftment therefore depends not only on cell survival but also on the capacity of the host environment to support vascular integration, immune resolution, and tissue remodeling.
In this context, regenerative failure should be viewed as a dynamic and multifactorial process rather than a simple absence of therapeutic efficacy. Early ischemic injury, dysregulated innate immune activation, impaired angiogenic adaptation, and altered stromal cell functionality may collectively compromise graft integration and long-term regenerative outcomes. Importantly, these mechanisms are likely interconnected and mutually reinforcing rather than isolated phenomena.
Defining “SVF therapy resistance”
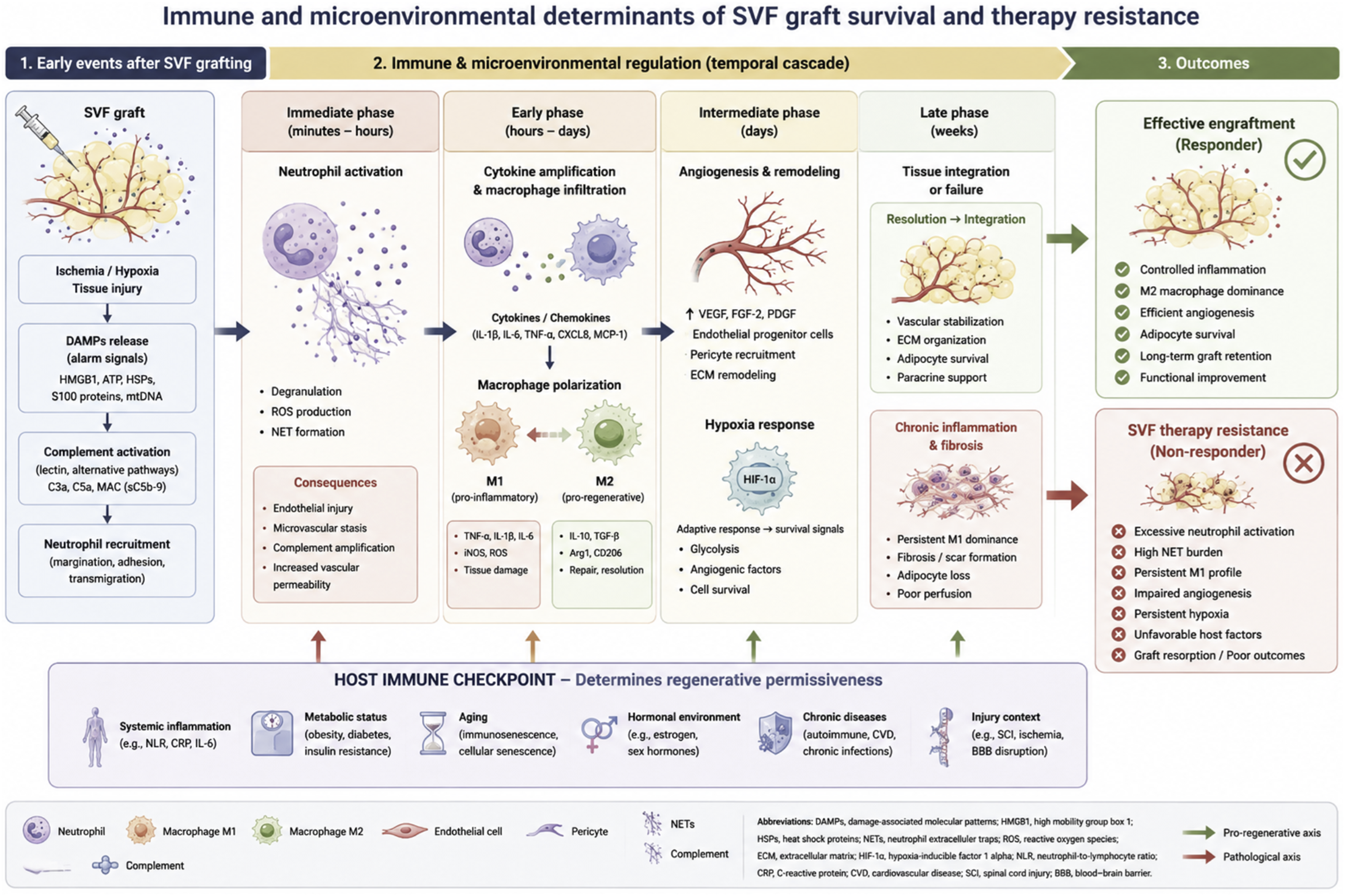
We propose that the concept of “SVF therapy resistance” may provide a useful operational framework to characterize patients or biological contexts associated with reduced responsiveness to SVF-based regenerative therapies. The proposed interactions between innate immunity, hypoxia, angiogenesis, and host-related determinants contributing to SVF therapy resistance are summarized in Figure 2. Immune and microenvironmental determinants of SVF graft survival and therapy resistance.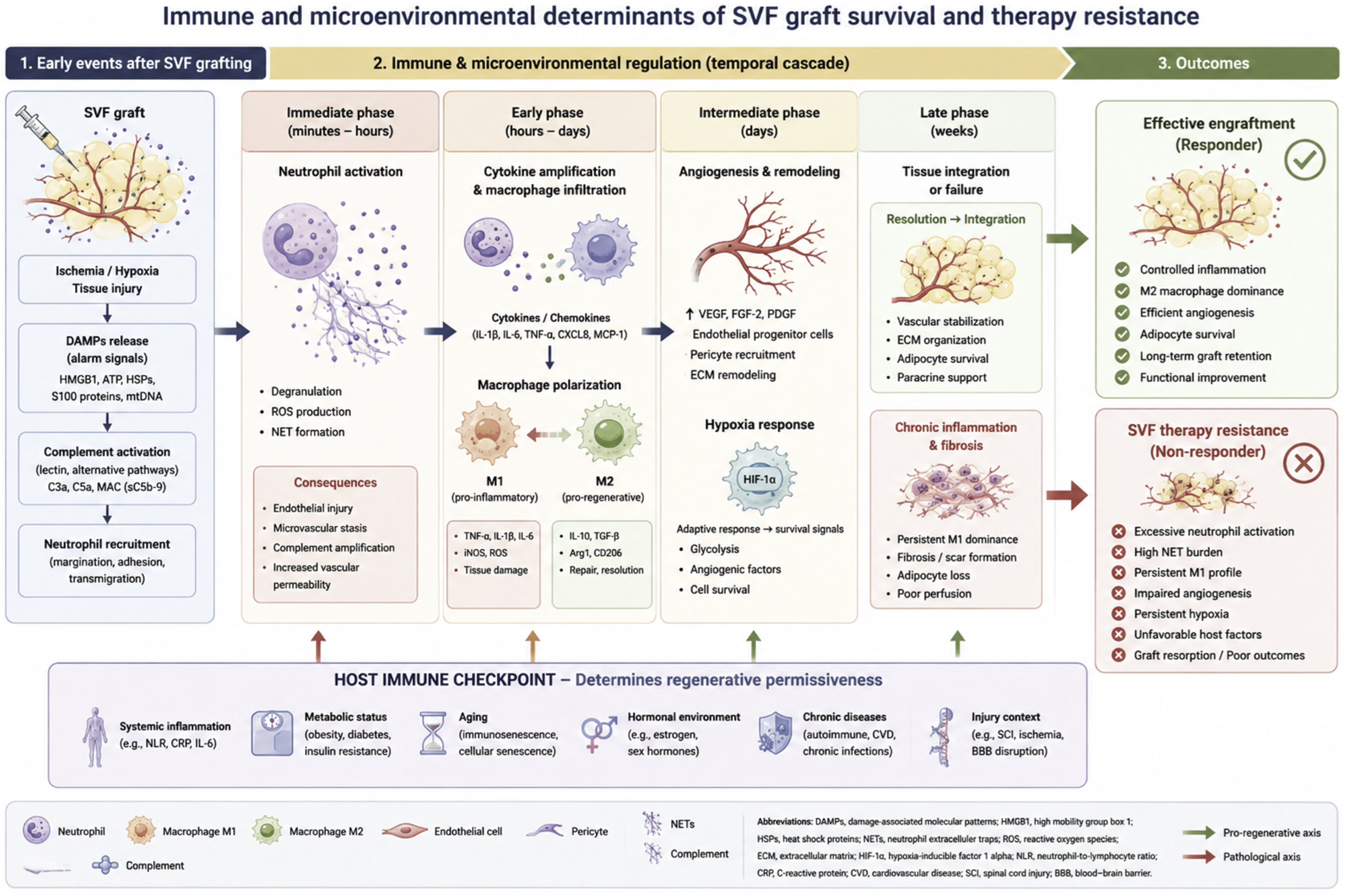
SVF therapy resistance can be defined as: a state in which maladaptive host–graft interactions prevent effective engraftment, integration, or regenerative activity of autologous SVF-based therapies despite technically adequate cell delivery.
Importantly, this concept does not imply classical immune rejection. Instead, it reflects the inability of the local and systemic microenvironment to support regenerative processes following transplantation. Such maladaptive responses may involve excessive inflammation, defective angiogenesis, persistent hypoxia, endothelial dysfunction, impaired immune resolution, or intrinsic cellular dysfunction within the transplanted SVF product.
Proposed features distinguishing responders and non-responders to SVF-based therapies.
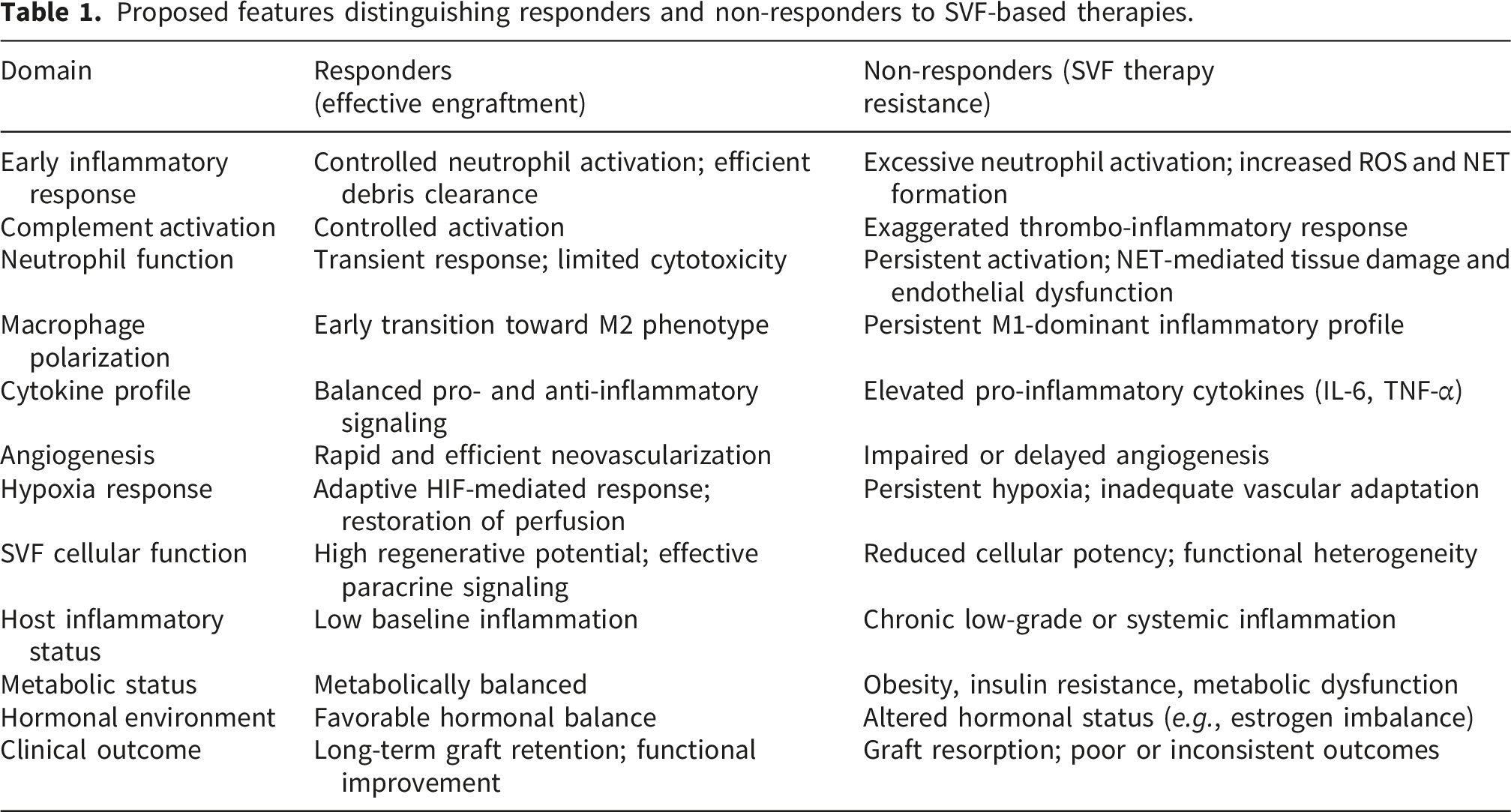
This framework may help explain why some patients exhibit limited clinical benefit despite apparently standardized procedures and adequate cell yields. It also provides a rationale for considering regenerative responsiveness as a biological continuum rather than a binary responder/non-responder state.
A multifactorial model of regenerative resistance
Current evidence suggests that regenerative outcomes in SVF therapy are influenced by at least four major interconnected domains: (i) Host immune and inflammatory status
Baseline inflammatory tone, neutrophil activation, macrophage polarization dynamics, oxidative stress, and systemic inflammatory disorders may critically influence graft fate.10–20 (ii) Angiogenic and microvascular competence
Successful graft integration requires rapid restoration of tissue perfusion. Impaired angiogenesis, endothelial dysfunction, and defective hypoxic adaptation may compromise graft survival.8,21,22 (iii) Cellular quality and SVF heterogeneity
The regenerative properties of SVF products depend on complex and variable cellular compositions, including ADSCs, endothelial progenitors, immune cells, and stromal populations.23–26 Aging, obesity, metabolic dysfunction, and chronic inflammation may alter these cellular functions. (iv) Procedural and biophysical determinants
Mechanical trauma during harvesting, enzymatic digestion, endotoxin contamination, complement activation, oxygen diffusion limitations, and graft geometry may independently modulate inflammatory responses and cell viability.
Rather than acting independently, these domains likely interact continuously throughout the engraftment process. For example, impaired angiogenesis may exacerbate hypoxia, which in turn amplifies innate immune activation and promotes additional tissue injury. Similarly, poor cellular quality may reduce resilience to inflammatory stress and ischemic conditions.
Distinguishing technical failure from biological resistance
An important distinction should be made between: • procedural or technical failure, • product-related insufficiency, • and true host-mediated regenerative resistance.
Technical failures may result from inadequate harvesting, poor injection technique, excessive mechanical stress, or insufficient cell viability. Product-related insufficiency may reflect altered SVF composition or low regenerative potency. In contrast, biological resistance refers to maladaptive host-graft interactions occurring despite technically adequate procedures and acceptable product quality.
This distinction is essential because it directly influences therapeutic strategies. While technical optimization may improve procedural consistency, overcoming biological resistance may require targeted modulation of inflammation, vascular responses, immune signaling, or cellular conditioning.
Toward precision regenerative medicine
The concept of SVF therapy resistance supports a transition from empirical regenerative approaches toward precision regenerative medicine. Rather than assuming equivalent responsiveness across patients, future strategies may require: • pre-therapeutic immune profiling, • characterization of inflammatory and angiogenic status, • assessment of SVF cellular quality, • and identification of biomarkers predictive of regenerative response.
Such approaches may help identify patients at higher risk of therapeutic failure and facilitate the development of personalized interventions aimed at improving engraftment and regenerative efficacy.
Ultimately, understanding regenerative failure as a systems-level interaction between host biology, graft characteristics, and procedural determinants may provide a more realistic and clinically relevant framework for optimizing SVF-based therapies.
Innate immunity as an early determinant of SVF engraftment
Early inflammatory responses following SVF grafting
Immediately after transplantation, adipose-derived grafts and SVF-containing products are exposed to profound ischemic and mechanical stress resulting from harvesting, processing, and implantation procedures. 7 This early injury phase leads to adipocyte disruption, extracellular matrix damage, and the release of danger-associated molecular patterns (DAMPs), thereby initiating a sterile inflammatory response within the graft microenvironment.
Although this inflammatory phase is necessary for tissue remodeling and debris clearance, excessive or dysregulated activation of innate immunity may critically compromise graft survival. The earliest stages of engraftment therefore represent a narrow biological window during which the balance between adaptive repair and uncontrolled inflammation determines long-term regenerative outcomes.
Importantly, innate immune activation after SVF transplantation does not occur in isolation but rather develops in a complex ischemic environment characterized by hypoxia, oxidative stress, endothelial dysfunction, and microvascular instability. These processes collectively shape the inflammatory landscape surrounding the graft.
Neutrophils and acute inflammatory injury
Neutrophils are among the first immune cells recruited to sites of tissue injury following fat grafting and SVF administration. 10 Their physiological role includes phagocytosis of necrotic debris, secretion of proteolytic enzymes, production of ROS, and orchestration of early antimicrobial defense mechanisms.
Experimental studies have demonstrated that disturbed neutrophil behavior during the early post-grafting period may negatively affect long-term graft retention. 10 Excessive neutrophil activation may amplify oxidative stress, damage endothelial structures, and exacerbate local ischemia. High ROS production can impair stromal cell viability, alter extracellular matrix integrity, and contribute to microvascular dysfunction.
Beyond simple inflammatory amplification, neutrophils may directly influence angiogenic responses. Excessive neutrophil-derived proteases and oxidants can disrupt endothelial integrity and impair neovascularization, thereby limiting oxygen and nutrient delivery to transplanted tissues during the critical early engraftment phase.
These observations suggest that neutrophil activity constitutes a central checkpoint in the balance between regenerative integration and inflammatory tissue injury.
Neutrophil extracellular traps (NETs) and sterile inflammatory toxicity
Among the mechanisms implicated in neutrophil-mediated tissue injury, neutrophil extracellular traps (NETs) have emerged as particularly relevant in regenerative contexts.11–14 NETs are extracellular chromatin structures decorated with histones, proteases, and antimicrobial proteins released by activated neutrophils during a specialized process termed NETosis. 11
Although NET formation contributes to antimicrobial defense, excessive NETosis has increasingly been implicated in sterile inflammatory diseases, thrombosis, endothelial injury, and microvascular dysfunction.12–14 Experimental evidence suggests that NETs may impair tissue perfusion through several mechanisms, including: • endothelial cytotoxicity, • platelet activation, • complement activation, • microvascular thrombosis, • and amplification of local inflammatory signaling.
In ischemic tissues, excessive NET formation may therefore critically compromise graft revascularization and oxygen delivery. NET-mediated obstruction of microcirculation may be particularly detrimental during the early post-transplantation period, when graft survival is highly dependent on rapid restoration of tissue perfusion.
Furthermore, NETs may contribute to persistent inflammatory amplification by sustaining macrophage activation and cytokine release. This creates a self-reinforcing inflammatory loop potentially unfavorable to regenerative integration.
Importantly, systemic conditions such as obesity, metabolic syndrome, diabetes, aging, and chronic inflammatory states have all been associated with altered neutrophil behavior and increased NET formation.17,18,26 Such baseline inflammatory predispositions may partly explain inter-individual variability in regenerative responses following SVF therapies.
Processing-related inflammatory priming
In addition to host-related immune determinants, procedural and processing-related factors may independently contribute to inflammatory activation following SVF administration. Mechanical trauma during liposuction, enzymatic digestion, centrifugation, washing procedures, and resuspension media may alter both cellular integrity and the inflammatory profile of the final SVF product.33,34
Importantly, tissue disruption and cell damage occurring during harvesting and processing can induce the release of damage-associated molecular patterns (DAMPs), thereby amplifying sterile inflammatory signaling independently of baseline host inflammatory status.35–37 In parallel, residual endotoxin contamination or lipopolysaccharide (LPS) exposure introduced during enzymatic processing, washing solutions, or handling procedures may further activate innate immune pathways through Toll-like receptor 4 (TLR4)-mediated signaling. 38
Even low-grade endotoxin exposure may enhance neutrophil activation, promote NETosis, stimulate pro-inflammatory macrophage polarization, and impair endothelial function and microvascular perfusion. In this context, mesenchymal stromal cells themselves may adopt distinct inflammatory phenotypes depending on environmental stimulation, shifting toward either pro-inflammatory (MSC1) or immunosuppressive (MSC2) functional states. 39 Moreover, interactions between transplanted stromal cells and blood components may activate complement and coagulation cascades, contributing to thrombo-inflammatory responses after transplantation.40,41 Similar inflammatory mechanisms have also been described in foreign body reactions to biomaterials and transplanted tissues. 42
These observations suggest that inflammatory priming associated with SVF processing may partially contribute to the proposed concept of SVF therapy resistance and should be distinguished from purely host-mediated biological resistance mechanisms.
Macrophage polarization and tissue remodeling
Following the initial neutrophil-dominated phase, macrophages become the major orchestrators of tissue remodeling and inflammatory resolution. Macrophages exhibit remarkable functional plasticity and can adopt a broad spectrum of activation states depending on local environmental cues.15,16
Classically activated M1 macrophages produce pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, thereby promoting inflammatory amplification and tissue degradation. In contrast, alternatively activated M2 macrophages support tissue repair through: • secretion of anti-inflammatory mediators, • stimulation of angiogenesis, • extracellular matrix remodeling, • adipogenesis, • and resolution of inflammation.
The balance between M1 and M2 polarization appears to represent a major determinant of graft fate. Persistent M1-dominant inflammation is associated with chronic tissue injury, fibrosis, and graft resorption, whereas timely transition toward an M2-dominant environment promotes vascular integration and long-term tissue stability.15,16
Importantly, macrophage polarization is strongly influenced by systemic host factors, including obesity, insulin resistance, metabolic dysfunction, hormonal status, and chronic low-grade inflammation.17–20,25 Metabolic dysfunction may drive distinct pro-inflammatory macrophage phenotypes characterized by altered cytokine production and impaired reparative activity. 25
These findings further support the concept that baseline host immune status may critically modulate regenerative responsiveness.
Crosstalk between innate immunity and angiogenesis
Innate immune responses and angiogenesis are closely interconnected processes during graft integration. Macrophages—particularly M2-like phenotypes—represent important sources of pro-angiogenic mediators including vascular endothelial growth factor (VEGF), transforming growth factor-β (TGF-β), and matrix remodeling enzymes.
Conversely, excessive neutrophil activation, NET formation, oxidative stress, and persistent inflammatory signaling may impair endothelial function and disrupt angiogenic processes. Thus, inflammatory dysregulation may indirectly compromise graft survival by limiting neovascularization.
The temporal coordination between inflammation and vascular remodeling therefore appears critical. Controlled inflammatory activation may support tissue adaptation and repair, whereas persistent or exaggerated innate immune responses may shift the microenvironment toward chronic injury and regenerative failure.
Innate immunity and the concept of SVF therapy resistance
Collectively, current evidence suggests that innate immunity should not be viewed merely as a secondary consequence of tissue transplantation but rather as a central regulator of SVF engraftment outcomes.
Variability in neutrophil activation, NET formation, macrophage polarization, oxidative stress responses, and inflammatory resolution mechanisms may create distinct host inflammatory phenotypes associated with differential regenerative capacity. In some patients, maladaptive innate immune responses may generate a persistently hostile microenvironment preventing effective graft integration despite technically adequate procedures and sufficient cell delivery.
Such observations support the hypothesis that dysregulated innate immunity constitutes one of the major biological foundations underlying the proposed concept of SVF therapy resistance.
Hypoxia, angiogenesis, and microvascular adaptation
Hypoxia as an unavoidable early stressor
Following transplantation, adipose tissue grafts and SVF-containing products are immediately exposed to a hypoxic environment due to disruption of their native vascular supply. 7 During the early post-implantation phase, oxygen and nutrient diffusion become critically limited, exposing transplanted cells to severe metabolic stress before adequate neovascularization can occur.
This ischemic period represents one of the principal bottlenecks of graft survival. Experimental studies have demonstrated that adipocyte death occurs rapidly after transplantation when vascular reconnection is insufficient. 7 Inadequate oxygenation promotes apoptosis, necrosis, lipid cyst formation, fibrosis, and progressive graft resorption. 8
Importantly, hypoxia should not be viewed solely as a deleterious phenomenon. Moderate and transient hypoxic exposure may activate adaptive cellular programs capable of supporting regenerative responses. In contrast, prolonged or severe ischemia may exceed the adaptive capacity of both grafted cells and host tissues, leading to irreversible tissue injury.
Thus, the biological outcome of hypoxia largely depends on the balance between adaptive vascular remodeling and persistent ischemic stress.
HIF signaling and adaptive angiogenic responses
Cellular adaptation to hypoxia is primarily mediated through stabilization of hypoxia-inducible factors (HIFs), which regulate the transcription of genes involved in angiogenesis, metabolism, cell survival, and tissue remodeling.20,21
Among HIF-regulated mediators, vascular endothelial growth factor (VEGF) plays a central role in promoting neovascularization and restoration of tissue perfusion. VEGF stimulates endothelial proliferation, vascular permeability, and microvascular remodeling, thereby facilitating oxygen delivery to ischemic tissues.
Experimental studies have shown that hypoxic preconditioning of adipose-derived stromal cells may enhance:
Hypoxia-preconditioned ADSCs exhibit increased secretion of VEGF and other trophic mediators, suggesting that controlled activation of hypoxic pathways may represent a therapeutic strategy to improve engraftment.
However, excessive or prolonged hypoxia may instead trigger inflammatory amplification, mitochondrial dysfunction, oxidative stress, and apoptotic signaling. Consequently, successful graft integration depends not simply on hypoxic exposure itself but rather on the efficiency of adaptive angiogenic responses.
Angiogenesis and adipose tissue integration
Angiogenesis and adipogenesis are tightly interconnected biological processes. 8 Adequate vascularization is required not only for the survival of pre-existing adipocytes but also for the maintenance, differentiation, and metabolic activity of newly regenerated adipose tissue.
The establishment of a functional microvascular network therefore constitutes a decisive step in long-term graft integration. Rapid neovascularization supports: • oxygen delivery, • nutrient exchange, • waste clearance, • stromal cell survival, • and tissue remodeling.
Conversely, delayed or insufficient vascular adaptation results in persistent ischemia and progressive graft deterioration. Histologically, poorly vascularized grafts are frequently characterized by fibrosis, oil cyst formation, calcification, and chronic inflammatory infiltration.
Importantly, angiogenic efficiency may vary substantially between individuals. Aging, obesity, endothelial dysfunction, diabetes, smoking, and chronic inflammatory states have all been associated with impaired vascular regenerative responses.17,25 Such host-related variability may critically influence the capacity to support graft integration after SVF transplantation.
Diffusion-limited ischemia and spatial graft heterogeneity
An additional but often underappreciated determinant of graft survival relates to the spatial organization and diffusion limitations within transplanted adipose tissue. SVF-containing grafts are structurally heterogeneous and may contain regions exposed to markedly different oxygenation conditions during the early post-transplantation period.
Cells located within the central core of dense graft clusters may experience prolonged ischemia before adequate neovascularization occurs. Because oxygen diffusion is limited to relatively short distances, central regions may undergo severe hypoxia, necrosis, and lipid degeneration despite apparently adequate peripheral vascularization. These necrotic zones may subsequently release DAMPs and inflammatory mediators, thereby amplifying innate immune activation and secondary tissue injury.
Importantly, diffusion-limited hypoxia may create a self-reinforcing loop linking ischemia, inflammation, endothelial dysfunction, impaired angiogenesis, and graft resorption. In this context, some manifestations currently interpreted as “host-mediated SVF therapy resistance” may partly reflect unfavorable graft architecture and biophysical constraints rather than exclusively immunological determinants.
These observations highlight the importance of considering graft geometry, cell density, and oxygen diffusion dynamics as additional modulators of regenerative outcomes.
Interactions between inflammation and angiogenesis
Inflammation and angiogenesis are deeply interconnected processes during regenerative tissue remodeling. Controlled inflammatory responses contribute to vascular adaptation by promoting endothelial activation and secretion of pro-angiogenic mediators.
Macrophages—particularly M2-like reparative phenotypes—represent key regulators of angiogenesis through the production of VEGF, TGF-β, matrix metalloproteinases, and extracellular remodeling factors.15,16 In this context, properly coordinated immune activation may facilitate vascular integration and tissue repair.
In contrast, excessive inflammatory activation may severely impair microvascular adaptation. Persistent neutrophil activation, oxidative stress, endothelial injury, and NET formation may disrupt endothelial integrity and promote microvascular dysfunction.10–14 NET-associated thrombosis and capillary obstruction may further compromise tissue perfusion during the critical early post-grafting period.
Thus, angiogenic failure may not only result from insufficient pro-angiogenic signaling but also from excessive inflammatory injury impairing vascular stability.
Biophysical constraints and oxygen diffusion limitations
Beyond biological signaling pathways, graft survival is also constrained by fundamental biophysical limitations. Oxygen diffusion within transplanted adipose tissue is intrinsically restricted, particularly in large graft volumes or densely packed cellular environments.
Cells located beyond effective oxygen diffusion distances may rapidly undergo hypoxic injury before vascular reconnection can occur. Consequently, graft geometry, injection technique, tissue dispersion, and local microarchitecture may substantially influence tissue viability independently of immune mechanisms.
Mechanical compression, elevated interstitial pressure, and impaired microcirculatory flow may further exacerbate ischemic stress. These factors suggest that regenerative outcomes depend not only on cellular composition and host biology but also on spatial and biophysical properties of the graft itself.
Importantly, these biophysical constraints may interact synergistically with inflammatory mechanisms. Hypoxia may amplify inflammatory signaling, while inflammation-induced endothelial dysfunction may worsen tissue ischemia, thereby creating self-reinforcing cycles of regenerative failure.
Angiogenic failure and SVF therapy resistance
Collectively, current evidence indicates that successful SVF engraftment critically depends on the ability to transition from an ischemic environment toward stable vascular integration.
Patients with impaired angiogenic competence, endothelial dysfunction, chronic inflammation, or defective hypoxic adaptation may exhibit reduced regenerative responsiveness despite technically adequate procedures. In such contexts, persistent ischemia may maintain a hostile microenvironment characterized by oxidative stress, inflammatory amplification, and progressive tissue injury.
These observations support the hypothesis that angiogenic insufficiency and defective microvascular adaptation constitute major mechanistic components of the broader framework of SVF therapy resistance.
Rather than representing isolated phenomena, hypoxia, inflammation, vascular remodeling, and cellular survival likely form an integrated biological network determining whether transplanted SVF products successfully integrate or undergo progressive regenerative failure.
Procedural and non-sterile inflammatory determinants
Procedural injury as an underrecognized determinant of regenerative outcomes
Although host-related biological factors are increasingly recognized as critical determinants of SVF therapy efficacy, procedural variables may independently influence graft survival and inflammatory responses. SVF-based therapies involve multiple processing steps—including harvesting, aspiration, centrifugation, washing, enzymatic digestion, filtration, and reinjection—that can substantially affect cellular integrity and microenvironmental signaling.
Historically, procedural optimization has primarily focused on maximizing cell yield and viability. However, accumulating evidence suggests that tissue processing itself may actively shape inflammatory and regenerative responses. Mechanical stress, enzymatic exposure, biomaterial interactions, and contamination-related inflammatory signaling may all contribute to early graft injury independently of host baseline characteristics.
Consequently, regenerative failure should not be interpreted exclusively as a host-mediated phenomenon but rather as the result of dynamic interactions between: • host biology, • graft characteristics, • and procedural inflammatory determinants.
Mechanical stress and tissue processing-related inflammation
Adipose harvesting and SVF preparation expose cells to substantial mechanical stress. Liposuction-induced shear forces, aspiration pressure, centrifugation, filtration, and repeated manipulation may induce cellular membrane disruption, mitochondrial injury, and extracellular matrix fragmentation.33,34
These mechanical insults may lead to the release of intracellular contents and DAMPs, thereby amplifying sterile inflammatory signaling immediately after transplantation.35,36 Damaged adipocytes and stromal cells may release: • mitochondrial DNA, • ATP, • histones, • heat shock proteins, • and oxidized lipids, which collectively contribute to innate immune activation. Circulating mitochondrial DAMPs have notably been shown to trigger inflammatory responses resembling pathogen-induced activation despite the absence of infection.35,36
Mechanical trauma may also alter endothelial integrity and impair the survival of fragile stromal cell populations. Excessive tissue manipulation has been associated with reduced adipocyte viability and impaired long-term graft retention in experimental and clinical studies. 33
Importantly, tissue processing methods may differentially influence inflammatory activation. Enzymatic SVF isolation, mechanical fragmentation, and nanofat preparation generate products with distinct cellular compositions, extracellular matrix content, and inflammatory profiles. Such differences may partly explain variability in regenerative outcomes observed across studies and clinical applications. 34
Enzymatic digestion and inflammatory signaling
Collagenase-based enzymatic digestion remains widely used for isolation of cellular SVF products. While enzymatic processing improves cell yield and facilitates stromal cell extraction, it may also modify extracellular matrix architecture and alter cellular interactions within the graft microenvironment.
Residual enzymatic activity, matrix degradation products, and altered cell-surface signaling may potentially contribute to inflammatory activation after transplantation. In addition, enzymatic digestion may remove structural extracellular matrix components that normally provide mechanical protection and trophic support to transplanted cells.
Importantly, mesenchymal stromal cells are themselves highly responsive to inflammatory stimuli. Toll-like receptor (TLR) signaling pathways, particularly TLR3 and TLR4, have been shown to modulate mesenchymal stromal cell immunoregulatory functions and alter their interaction with immune cells. 38 Furthermore, exposure to inflammatory stimuli may polarize mesenchymal stromal cells toward distinct functional phenotypes, including pro-inflammatory or immunosuppressive states. 39
These observations suggest that procedural inflammatory signals may directly influence the biological behavior of transplanted SVF cells and alter their regenerative potential.
Importantly, enzymatic and mechanical SVF products should not be considered biologically equivalent. Mechanically derived stromal vascular tissue (SVT) or nanofat preparations retain substantial extracellular matrix content and microstructural organization, whereas enzymatically isolated SVF represents a more dissociated cellular suspension.
43
These differences may influence: • cell survival, • inflammatory responses, • vascular integration, • and tissue remodeling dynamics.
Such distinctions may become particularly relevant in highly inflammatory environments such as spinal cord injury, where graft–host interactions are strongly influenced by tissue architecture and inflammatory priming.
Endotoxin contamination and innate immune priming
An additional underrecognized determinant of regenerative outcomes involves low-grade endotoxin exposure and non-sterile inflammatory signaling associated with tissue processing procedures.
Even minimal endotoxin contamination may strongly activate innate immune pathways through Toll-like receptor 4 (TLR4) and CD14 signaling, leading to amplification of:
• IL-6,
• TNF-α,
• ROS production,
• neutrophil activation,
• and macrophage polarization.
Importantly, mesenchymal stromal cells and adipose-derived stromal cells are highly sensitive to inflammatory priming. Exposure to inflammatory microenvironments may profoundly alter their immunomodulatory properties, cytokine secretion profiles, and extracellular vesicle signaling. 39
Thus, regenerative therapies intended to suppress inflammation may paradoxically become sources of inflammatory activation when exposed to procedural contaminants or excessive tissue injury.
Although clinically significant contamination is uncommon under appropriate manufacturing conditions, low-level inflammatory priming may still contribute to inter-study variability and inconsistent therapeutic responses, particularly in already inflamed or ischemic tissues.
Complement activation and innate immune recognition
Following transplantation, SVF products may also interact directly with plasma proteins and complement pathways. Cellular surfaces, extracellular vesicles, matrix fragments, and damaged membranes may trigger complement activation and opsonization, thereby facilitating innate immune recognition.
Therapeutic mesenchymal stromal cells have been shown to activate complement cascades and interact dynamically with circulating blood components following administration.
43
Similar mechanisms may occur after SVF transplantation and contribute to: • neutrophil recruitment, • macrophage activation, • endothelial dysfunction, • and thromboinflammatory responses.
In highly vascularized environments, innate immune activation may resemble forms of instant blood-mediated inflammatory reaction (IBMIR), initially described in transplantation biology. 40 Such reactions may impair microvascular perfusion and contribute to early graft injury.
Furthermore, adsorption of plasma proteins onto transplanted cellular products may generate a dynamic “protein corona” capable of modifying immune recognition and inflammatory signaling. More broadly, biomaterial-associated inflammatory responses have been shown to critically influence macrophage behavior and tissue integration processes. 41
Although these mechanisms remain insufficiently characterized in SVF-based therapies, they may represent important contributors to regenerative variability.
Resuspension media and microenvironmental modulation
The local physicochemical environment surrounding transplanted SVF products may further influence inflammatory and regenerative responses. Osmolarity, ionic composition, protein content, and carrier materials may all affect cellular viability and immune activation.
Different resuspension media—including saline, platelet-rich plasma, albumin-containing solutions, or biomaterial scaffolds—may differentially modulate:
• endothelial interactions,
• cytokine signaling,
• oxidative stress,
• and cell survival.
Similarly, local biomaterial properties may influence macrophage polarization and foreign-body responses. 41 Even subtle differences in injection matrices or carrier materials may therefore contribute to variability in regenerative outcomes.
These observations reinforce the idea that regenerative efficacy depends not only on cellular composition but also on the surrounding procedural microenvironment.
Procedural inflammation versus host-mediated resistance
A critical distinction should be made between:
• technical failure,
• procedure-induced inflammatory injury,
• product-related insufficiency,
• and true host-mediated regenerative resistance.
In many cases, therapeutic failure is likely multifactorial and results from overlapping contributions of these mechanisms. Excessive procedural trauma may amplify inflammatory signaling and worsen ischemic stress, thereby mimicking or exacerbating host-related resistance phenomena.
Consequently, the concept of SVF therapy resistance should not be interpreted as assigning therapeutic failure exclusively to patient biology. Instead, regenerative outcomes likely emerge from complex host–graft–procedure interactions occurring within highly dynamic inflammatory and vascular microenvironments.
Recognizing this complexity is essential for developing rational regenerative strategies. Optimizing procedural techniques, minimizing inflammatory priming, preserving cellular integrity, and controlling microenvironmental stressors may be just as important as host immune modulation itself.
Ultimately, integrating procedural determinants into the broader framework of SVF therapy resistance provides a more comprehensive and biologically coherent model of regenerative failure.
Immune profiling, biomarkers, and precision regenerative medicine
Candidate biomarkers and assessment tools for predicting SVF therapy outcomes.
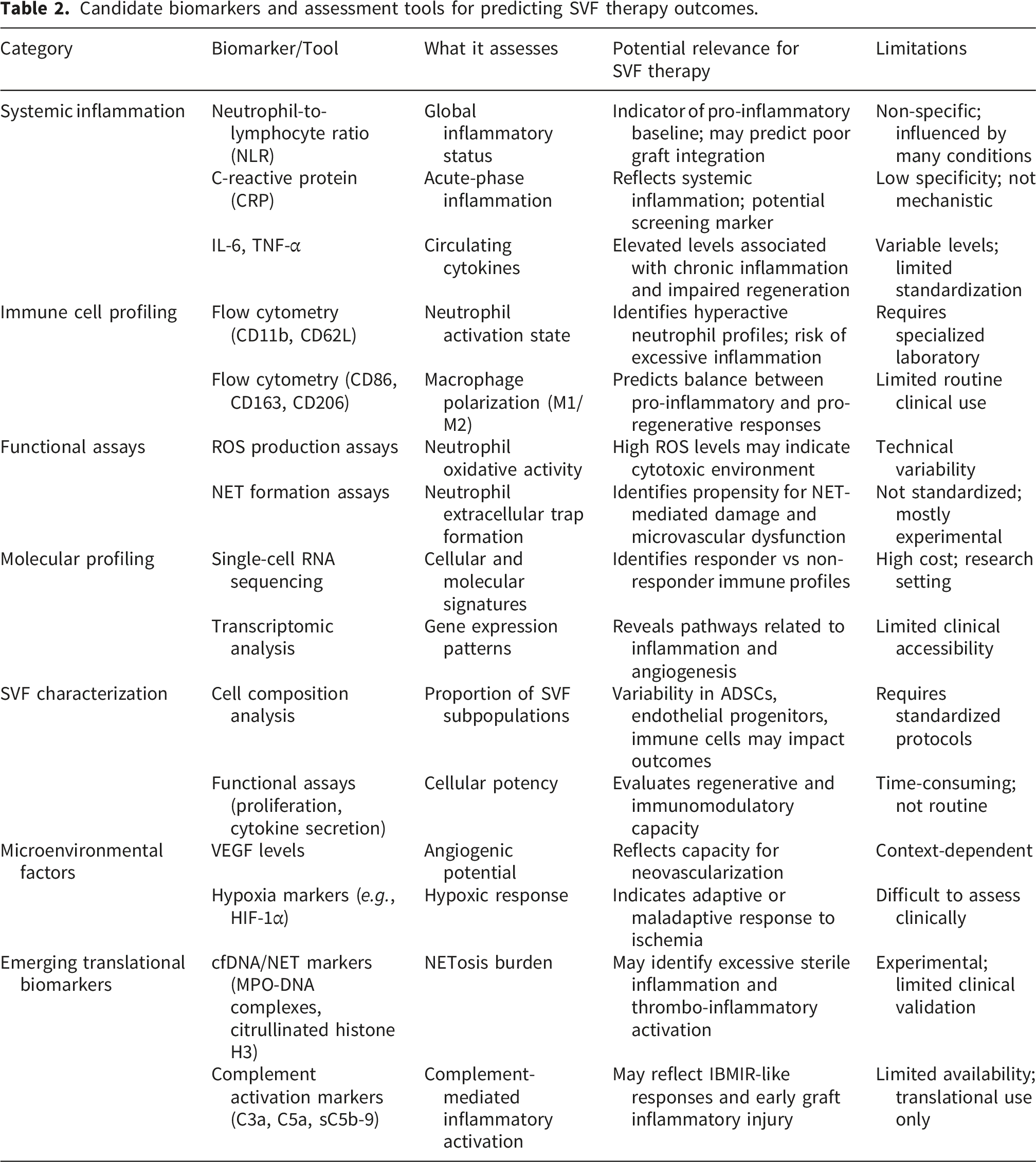
One of the major limitations of current SVF-based therapies is the absence of reliable tools capable of predicting therapeutic response. Despite increasing clinical use, regenerative interventions are still largely applied according to standardized protocols without accounting for inter-individual biological variability. However, growing evidence suggests that regenerative efficacy is highly dependent on host immune status, inflammatory balance, vascular adaptation, and intrinsic SVF cellular functionality.6,9,10 In this context, the concept of “SVF therapy resistance” implies the existence of biologically distinct responder and non-responder profiles, supporting the emergence of a precision regenerative medicine approach. 32
At the clinical level, systemic inflammatory biomarkers may provide initial insights into the host microenvironment. Chronic low-grade inflammation associated with obesity and metabolic dysfunction has been shown to alter macrophage polarization, endothelial function, and tissue repair capacities.11,12,20 Simple inflammatory markers such as the neutrophil-to-lymphocyte ratio (NLR), C-reactive protein (CRP), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α) may therefore help identify patients with a pro-inflammatory baseline state potentially unfavorable to graft integration.
Beyond circulating biomarkers, immune cell profiling may offer a more mechanistic evaluation of regenerative responsiveness. Flow cytometry-based analyses enable characterization of neutrophil activation states, macrophage polarization, and inflammatory immune signatures. 10 In particular, the balance between pro-inflammatory and pro-regenerative macrophage phenotypes appears critical for tissue repair and graft remodeling.18,19,44 Mantovani et al. highlighted the remarkable plasticity of macrophages during tissue repair processes, emphasizing their central role in the transition between inflammation and regeneration. 44 Similarly, Frangogiannis demonstrated that successful tissue repair requires tightly regulated temporal coordination between inflammatory activation and resolution pathways, suggesting that dysregulated inflammatory responses may compromise regenerative outcomes.
Neutrophil functional profiling may represent another important area of investigation. Excessive neutrophil activation and pathological NET formation have been associated with endothelial dysfunction, impaired angiogenesis, and tissue damage.13–17 Since sterile inflammatory responses are largely triggered by DAMP release following tissue injury and adipocyte disruption,35–37 variations in innate immune activation may significantly contribute to the heterogeneity of SVF therapy outcomes. These observations are particularly relevant considering that adipose tissue harvesting, processing, and grafting themselves induce mechanical stress and tissue injury capable of activating inflammatory cascades.22,33–37
Recent advances in mesenchymal stromal cell biology have further demonstrated that MSCs are not functionally homogeneous populations. Krampera described MSC “licensing” as a dynamic process through which inflammatory signals modulate immunoregulatory functions. 9 Similarly, Waterman et al. proposed the concept of MSC polarization into pro-inflammatory MSC1 and immunosuppressive MSC2 phenotypes depending on microenvironmental stimuli. 39 Liotta et al. further demonstrated that Toll-like receptor activation may profoundly alter MSC immunomodulatory properties through disruption of Notch signaling pathways. 38 Together, these findings strongly support the concept that the host inflammatory environment directly influences SVF biological behavior and therapeutic efficacy.
Importantly, increasing evidence suggests that SVF cellular heterogeneity itself may significantly affect regenerative outcomes. Functional variability between stromal cell populations influences proliferative capacity, cytokine secretion, angiogenic activity, and immunomodulatory potential.26–28 Phinney emphasized that mesenchymal stromal cells exhibit substantial functional heterogeneity with important implications for cell therapy reproducibility and therapeutic potency. 45 Wilson et al. further highlighted the complexity and multiplicity of mesenchymal stromal cell populations, emphasizing the need for more refined characterization and standardization strategies in regenerative medicine applications. 46
Single-cell transcriptomic technologies now offer unprecedented opportunities to characterize immune and stromal cell heterogeneity at high resolution. Papalexi and Satija demonstrated how single-cell RNA sequencing enables detailed mapping of immune cell functional states and inflammatory microenvironments. 47 Such approaches may facilitate identification of molecular signatures associated with responder and non-responder profiles in SVF-based therapies. Transcriptomic analyses may notably help identify pro-inflammatory states, angiogenic deficiencies, dysfunctional stromal populations, and maladaptive immune responses associated with therapeutic resistance.
In parallel, interactions between transplanted stromal cells and blood components may also influence therapeutic outcomes. Moll et al. demonstrated that mesenchymal stromal cells may trigger innate immune activation and complement-mediated inflammatory responses following systemic exposure. 43 Similarly, Moberg et al. described how tissue factor expression can initiate thrombo-inflammatory reactions capable of impairing cellular engraftment. 40 Although these mechanisms have been primarily investigated in systemic cellular therapies, they may also contribute to early inflammatory events occurring after SVF transplantation. More broadly, Anderson et al. described how biomaterial-associated foreign body reactions are driven by coordinated innate immune activation and macrophage responses, 41 further supporting the concept that implanted biological products are strongly influenced by host immune recognition mechanisms, even in autologous settings.
Collectively, these observations support a transition from empirical regenerative medicine toward biologically informed therapeutic strategies. Rather than applying identical protocols to all patients, future regenerative approaches may require integration of immune profiling, inflammatory biomarkers, metabolic assessment, and functional SVF characterization to guide therapeutic decision-making. Trounson and McDonald emphasized that one of the major challenges limiting stem cell translation remains the inability to predict therapeutic variability and optimize patient selection. 48 In this context, precision regenerative medicine approaches integrating host biology and cellular functionality may represent a critical step toward improving the consistency and efficacy of SVF-based therapies. 32
Beyond innate immune markers, adaptive immune regulation may also contribute to regenerative responsiveness. Regulatory T cells (Tregs), characterized by CD25 and FOXP3 expression, play a central role in controlling excessive inflammation and supporting tissue repair. Experimental evidence suggests that Tregs may promote macrophage polarization toward pro-regenerative phenotypes and facilitate resolution of inflammation.
In chronic inflammatory conditions such as SCI, impaired Treg function or reduced CD25-mediated immunoregulation may contribute to persistent inflammatory activation and defective tissue remodeling. Consequently, Treg-associated markers may represent additional candidate biomarkers for identifying permissive versus non-permissive host microenvironments in SVF-based therapies.
SVF therapy in spinal cord injury (SCI): A high-risk context for therapeutic resistance
The hostile inflammatory and microvascular environment associated with SCI and its potential implications for SVF therapy resistance are illustrated in Figure 3. Spinal cord injury as a high-risk microenvironment for SVF therapy resistance.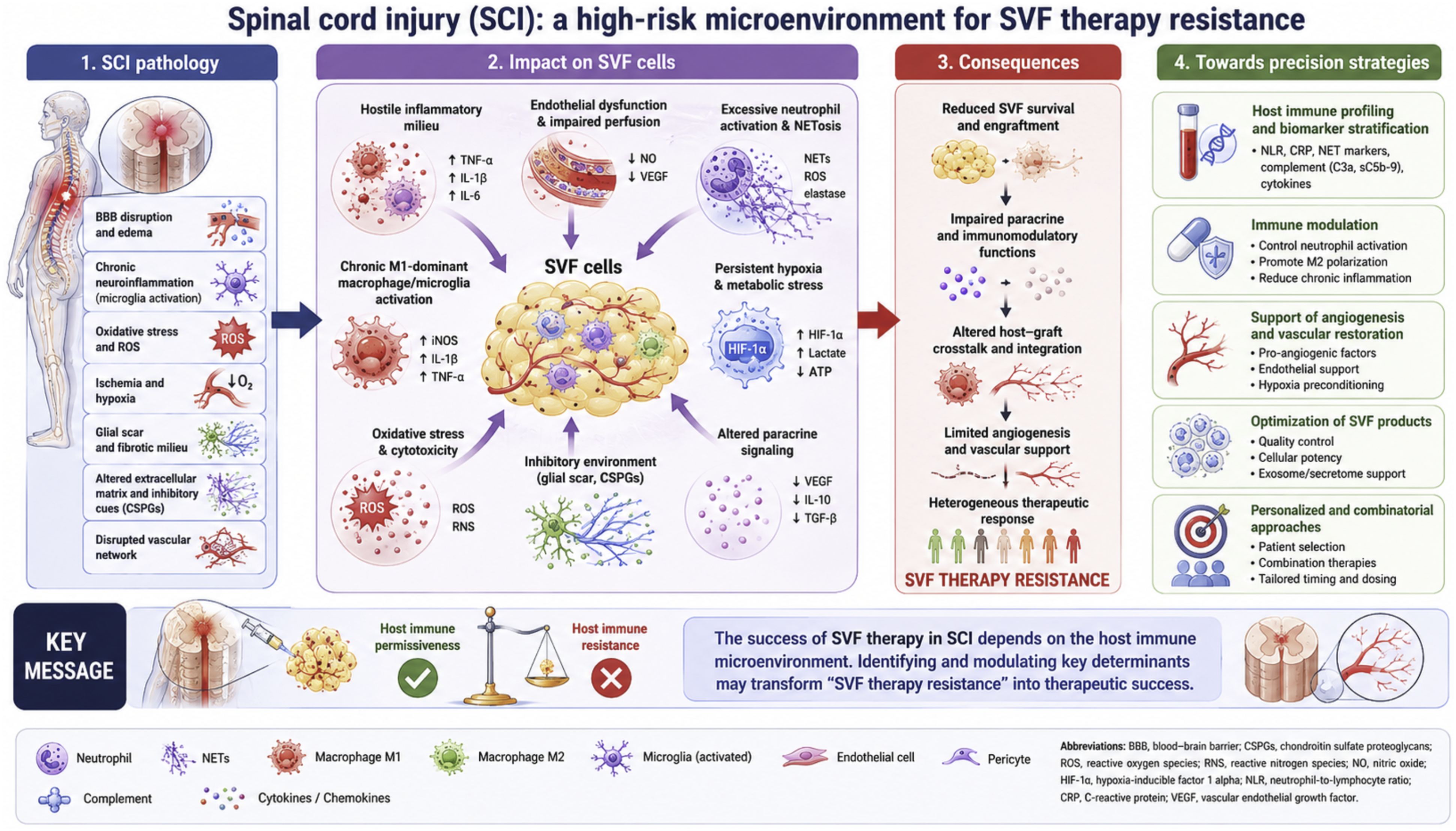
SCI represents one of the most challenging indications for regenerative medicine due to the profound and persistent alterations of the local and systemic microenvironment following injury. 29 Beyond the initial mechanical trauma, SCI triggers a complex cascade of secondary injury mechanisms including disruption of the blood–spinal cord barrier, ischemia, oxidative stress, excitotoxicity, immune activation, and progressive neurodegeneration. These processes collectively create a hostile inflammatory environment that may severely compromise the efficacy of cell-based regenerative therapies, including SVF-based approaches.
Innate immune activation plays a central role in SCI pathophysiology. Immediately following injury, neutrophils rapidly infiltrate the damaged spinal cord and contribute to secondary tissue injury through the release of proteases, ROS, and inflammatory mediators. 29 Similar to peripheral tissue injury, excessive neutrophil activation and NET formation may further exacerbate endothelial dysfunction, microvascular impairment, and tissue destruction.13–17 Persistent sterile inflammation driven by DAMP release from injured neural tissue additionally amplifies inflammatory signaling pathways and contributes to chronic immune dysregulation.35–37
Macrophages and activated microglia subsequently become dominant regulators of the post-traumatic inflammatory environment. However, unlike physiological tissue repair where inflammation progressively resolves, SCI is frequently characterized by prolonged persistence of pro-inflammatory macrophage phenotypes.29,44 This sustained inflammatory state promotes axonal degeneration, glial scar formation, and impaired tissue remodeling, thereby limiting regenerative capacity. Since successful SVF engraftment appears highly dependent on the transition toward pro-regenerative immune responses,18,19,44 the chronically inflammatory SCI microenvironment may represent a particularly unfavorable setting for regenerative therapies.
Hypoxia and vascular dysfunction constitute additional major barriers to successful regeneration after SCI. Disruption of spinal cord microcirculation induces prolonged ischemia and impaired oxygen delivery, contributing to neuronal death and defective tissue repair. 29 Although hypoxia may transiently activate adaptive angiogenic pathways through HIF-mediated signaling,23–25 chronic ischemia within the injured spinal cord often results in insufficient vascular remodeling and persistent metabolic stress. Such conditions may critically impair survival, integration, and paracrine activity of transplanted SVF cells.
Despite these limitations, SVF-based therapies remain attractive candidates for SCI treatment due to their multimodal mechanisms of action. SVF contains heterogeneous cellular populations capable of exerting immunomodulatory, angiogenic, neuroprotective, and trophic effects.1,2,6 Adipose-derived stromal cells notably secrete cytokines, growth factors, and extracellular vesicles capable of modulating inflammation and supporting tissue repair.4,6 Importantly, increasing evidence suggests that MSCs immunomodulatory functions are highly dynamic and strongly influenced by the host inflammatory environment.9,38,39 This concept may be particularly relevant in SCI, where the pathological microenvironment may alter or impair the regenerative behavior of transplanted cells.
Preclinical studies have provided encouraging results regarding SVF-based therapies in SCI models. Ertlen et al. demonstrated that immediate autologous SVF transplantation following spinal cord contusion improved functional sensorimotor recovery in rats, supporting the therapeutic potential of SVF in acute SCI settings. 30 More recently, refined translational large-animal models have further improved the experimental evaluation of regenerative approaches in SCI. 31 However, despite promising experimental findings, significant variability in therapeutic response remains a major challenge in regenerative neurology.
Importantly, the intrinsic heterogeneity of SVF products may further contribute to inconsistent outcomes in SCI applications. Differences in harvesting techniques, processing methods, and cellular composition can substantially influence regenerative potential.33,34,42 In particular, Nonnarath and Serratrice recently emphasized the critical biological differences between enzymatically isolated SVF and mechanically derived stromal vascular tissue or nanofat preparations in the context of SCI applications. 42 Such distinctions may significantly influence inflammatory interactions, angiogenic potential, and therapeutic efficacy and security. 49
The concept of “SVF therapy resistance” may therefore be particularly relevant in SCI. Patients exhibiting persistent systemic inflammation, impaired vascular adaptation, metabolic dysfunction, or maladaptive immune responses may be less likely to respond favorably to autologous regenerative therapies. Similarly, chronic SCI stages characterized by established fibrosis, glial scarring, and prolonged neuroinflammation may represent biologically resistant environments for cellular engraftment and repair.
In this context, precision regenerative medicine approaches may become essential for optimizing future SCI therapies. 32 Integrating immune profiling, inflammatory biomarkers, microenvironmental characterization, and functional SVF analysis could help identify patients most likely to benefit from regenerative interventions. Moreover, combining SVF transplantation with strategies aimed at modulating inflammation, enhancing angiogenesis, or improving cell survival may help overcome some mechanisms of therapeutic resistance.
Ultimately, SCI may represent one of the clearest examples illustrating how host immune and microenvironmental factors critically determine regenerative success. Understanding and overcoming these barriers will likely be essential for fully harnessing the therapeutic potential of SVF-based therapies in neurological disorders.
Clinical implications and future directions
The growing recognition of host-dependent determinants of SVF efficacy has important implications for the future development of regenerative medicine. Current SVF-based therapies are still largely performed using standardized protocols with limited consideration of patient-specific biological variability. However, accumulating evidence suggests that regenerative outcomes are strongly influenced by the complex interplay between innate immunity, angiogenesis, metabolic status, and intrinsic cellular functionality.6,9,10,44 Consequently, a “one-size-fits-all” approach may not be optimal for regenerative therapies.
One major implication of the proposed concept of “SVF therapy resistance” is the need to integrate pre-treatment biological assessment into clinical practice. Evaluating inflammatory status, immune activation, vascular function, and metabolic condition may help identify patients at increased risk of poor graft integration or suboptimal therapeutic response. Simple inflammatory biomarkers such as NLR, CRP, IL-6, and TNF-α could potentially serve as preliminary screening tools, while more advanced immune profiling approaches may provide deeper mechanistic characterization in selected patients.
Importantly, these considerations may also help explain part of the variability observed across clinical studies investigating SVF and fat grafting procedures.7,8,34 Differences in patient inflammatory status, obesity-related immune dysfunction, hormonal environment, aging, or chronic disease may substantially influence therapeutic responsiveness independently of procedural technique. Such factors may partly account for the inconsistent clinical outcomes frequently reported in regenerative medicine trials.6,48
The host microenvironment itself may also become a therapeutic target. Strategies aimed at modulating excessive inflammation, reducing NET formation, promoting macrophage polarization toward pro-regenerative phenotypes, or improving angiogenesis may help optimize graft survival and tissue integration.18,19,44 Similarly, preconditioning approaches designed to enhance cellular resistance to hypoxia and inflammatory stress may further improve regenerative efficacy.24,25
Recent advances in mesenchymal stromal cell biology additionally suggest that SVF therapeutic activity is highly dynamic and context-dependent.9,38,39 Rather than functioning as passive regenerative products, stromal cells continuously interact with inflammatory and microenvironmental signals. Consequently, future therapeutic strategies may require better control of these host–cell interactions in order to maximize regenerative potential.
Standardization of SVF preparation also remains a major challenge. Variability in harvesting methods, processing techniques, enzymatic digestion, mechanical isolation, and cellular composition may substantially influence biological activity and therapeutic consistency.33,34,42 In particular, the distinction between enzymatically isolated SVF and mechanically derived stromal vascular tissue or nanofat preparations remains critical, especially in neurological applications. 42 Improved characterization and harmonization of SVF products will therefore likely be necessary for future clinical translation.
Emerging technologies such as transcriptomics, proteomics, metabolomics, and single-cell RNA sequencing may provide powerful tools for identifying predictive molecular signatures associated with therapeutic responsiveness.27,47 Such approaches could facilitate development of integrated predictive models combining: • immune profiling, • inflammatory biomarkers, • angiogenic potential, • metabolic status, • and SVF cellular functionality.
Ultimately, these strategies may support the emergence of precision regenerative medicine approaches tailored to individual biological profiles. 32
However, several limitations must also be acknowledged. First, many mechanisms discussed in this review remain hypothetical and require prospective clinical validation. Direct evidence linking specific immune profiles or inflammatory biomarkers to SVF therapeutic failure remains limited. Second, the majority of currently available studies are preclinical, exploratory, or observational, limiting the ability to establish definitive causal relationships. Third, the substantial heterogeneity of SVF products and clinical protocols complicates interpretation and comparison across studies.
Future research should therefore focus on prospective translational studies integrating: • immune monitoring, • molecular profiling, • functional SVF characterization, • and standardized clinical outcome assessment.
Mechanistic studies exploring the interactions between neutrophils, macrophages, endothelial cells, and stromal cell populations will also be essential to better understand the biological determinants of graft survival and regenerative efficacy.
Finally, highly inflammatory and ischemic conditions such as spinal cord injury may represent particularly relevant models for studying mechanisms of SVF therapy resistance.29–32 These pathological contexts may provide valuable insights into how hostile microenvironments influence regenerative outcomes and how targeted interventions could potentially overcome therapeutic resistance.
Overall, moving from empirical regenerative medicine toward biologically informed and personalized therapeutic strategies may represent one of the most important future directions for SVF-based therapies. A deeper understanding of host–graft interactions will likely be essential to improve therapeutic consistency, optimize patient selection, and fully unlock the regenerative potential of SVF technologies.
Limitations
Several limitations should be acknowledged regarding the present review. First, this work was designed as a narrative and hypothesis-generating review rather than a systematic meta-analysis. Consequently, the proposed concept of “SVF therapy resistance” remains primarily conceptual and is supported by indirect evidence derived from heterogeneous experimental, translational, and clinical studies.
Second, the available literature on SVF-based therapies remains highly heterogeneous in terms of harvesting procedures, processing methods, cellular composition, administration routes, and clinical indications. Such variability limits direct comparisons between studies and complicates the identification of standardized determinants of therapeutic response.
Third, many mechanistic insights discussed in this review originate from related fields including mesenchymal stromal cell biology, transplantation immunology, biomaterial-associated inflammation, wound healing, and ischemia–reperfusion injury. Although these mechanisms are biologically relevant to SVF therapies, direct evidence specifically demonstrating their role in human SVF graft failure remains limited.
In addition, host-related determinants such as systemic inflammation, metabolic dysfunction, hormonal status, and immune heterogeneity are dynamic and multifactorial processes that may vary considerably between patients and disease contexts. The absence of validated biomarkers currently limits the clinical applicability of patient stratification approaches.
Finally, procedural variables including harvesting techniques, enzymatic or mechanical processing, washing procedures, and potential inflammatory priming of SVF products may independently influence therapeutic outcomes and remain insufficiently standardized across studies. Future prospective studies integrating immune profiling, molecular biomarkers, and standardized SVF characterization will be required to validate the proposed framework and clarify the respective contributions of host-, product-, and procedure-related determinants of regenerative efficacy.
Conclusions
SVF-based therapies represent a promising and rapidly evolving field within regenerative medicine. Their therapeutic potential relies on the combined effects of heterogeneous stromal and immune cellular populations capable of promoting angiogenesis, immunomodulation, and tissue repair. However, despite encouraging experimental and clinical results, substantial inter-individual variability remains a major limitation to their broader clinical translation.
This review highlights the central role of host-related factors in determining regenerative outcomes. Graft survival and therapeutic efficacy appear to depend not only on the intrinsic properties of transplanted SVF cells, but also on the dynamic interactions established with the host immune and microenvironmental landscape. Early innate immune responses, particularly neutrophil activation and NET formation, emerge as critical determinants of tissue integration and vascular preservation. Subsequently, macrophage polarization, angiogenic adaptation, and hypoxia responses further shape the balance between regeneration and graft failure.
Collectively, these mechanisms support the emerging concept of “SVF therapy resistance,” defined as the failure of autologous regenerative therapies due to maladaptive interactions between transplanted cellular products and the host biological environment. Unlike classical immune rejection, this phenomenon likely reflects the convergence of chronic inflammation, impaired vascular adaptation, metabolic dysfunction, and intrinsic cellular heterogeneity.
Importantly, increasing evidence suggests that SVF therapeutic activity is highly context-dependent and dynamically influenced by inflammatory signaling pathways. Variability in SVF composition, harvesting methods, processing techniques, and host immune status may therefore contribute substantially to inconsistent clinical outcomes. These observations further emphasize the limitations of standardized “one-size-fits-all” regenerative approaches.
The integration of immune profiling, inflammatory biomarkers, molecular characterization, and functional SVF assessment may help identify responder and non-responder profiles, thereby supporting the development of precision regenerative medicine strategies. Emerging technologies such as single-cell transcriptomics and advanced molecular profiling may further improve understanding of host–graft interactions and facilitate the identification of predictive therapeutic signatures.
These concepts may be particularly relevant in highly inflammatory and ischemic conditions such as spinal cord injury, where hostile microenvironments likely represent major barriers to successful regeneration. In such contexts, overcoming mechanisms of SVF therapy resistance may become essential for improving therapeutic efficacy.
Ultimately, future progress in regenerative medicine will likely depend on the ability to move beyond empirical therapeutic approaches toward biologically informed and personalized strategies integrating both cellular and host-related determinants of response.
Understanding regenerative failure as a dynamic interaction between host immunity, angiogenic competence, stromal cell functionality, and procedural inflammatory priming may represent a critical step toward the development of precision regenerative medicine strategies in SVF-based therapies.
Footnotes
Author contributions
CN & NS contribute equally to this work.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Artificial intelligence declaration
The authors declare that they have not use AI-generated work in this manuscript.