
Abstract
General intelligence (g) represents the factors that influence performance across all academic and cognitive domains. The search for these factors has been ongoing for more than a century and has focused on the brain and cognitive systems that support learning and problem solving. In recent decades, it has become clear that the factors that influence academic and cognitive performance extend to general health and to successful aging in adulthood. The implication is that there may be one or several fundamental processes that influence the functioning of all biological systems, not simply the brain. The functioning of mitochondria is well situated as one of the processes that might unify intelligence, health, and aging. These organelles are located within cells and are the primary producers of cellular energy, among other functions. Energy availability, in turn, is the lowest common denominator needed for the development, maintenance, and optimal functioning of all biological systems. Here, I review the relations among intelligence, health, and aging and outline how the efficiency of mitochondrial functioning can link them together.
Well over a century ago, Spearman (1904) discovered positive correlations among cognitive and academic domains and concluded “that all branches of intellectual activity have in common one fundamental function (or group of functions)” (p. 285), which he termed general intelligence, or g. Since that time, researchers have assumed that the fundamental bases of g must reside in some constellation of brain and cognitive processes that facilitate learning and comprehension across domains. The associated research programs have yielded important insights into the abilities that contribute to intelligence (Holyoak & Thagard, 1997) and the underlying cognitive and brain systems that support them (Jung & Haier, 2007; Shipstead, Harrison, & Engle, 2016). In recent decades, however, it has become clear that the factor common to all branches of intellectual activity must, at least in part, extend beyond brain and cognition. This is because the positive correlations among cognitive and academic measures extend to many indexes of health and aging (Deary, Weiss, & Batty, 2010). There may be one or several basic biological mechanisms that broadly influence individual differences in human health, aging, and brain and cognitive functioning.
The identification of any such mechanisms will not only deepen our understanding of the nexus of relations among intelligence, health, and aging but might also lead to treatments to ameliorate environmental compromises to them. One candidate is the functioning of the evolutionarily ancient mitochondria, small organelles that reside within cells. The mitochondria are the primary producers of cellular energy (i.e., adenosine triphosphate, or ATP) and have been implicated in myriad physical, neurological, and psychiatric diseases (e.g., Grimm, Mensah-Nyagan, & Eckert, 2016; Picard et al., 2015) and in normal aging (Lane, 2011). Here, I provide a brief overview of the relations among intelligence, health, and aging and outline some of the reasons why mitochondrial functions could provide a parsimonious explanation of them. The basic structure on which my argument is based is illustrated in Figure 1.
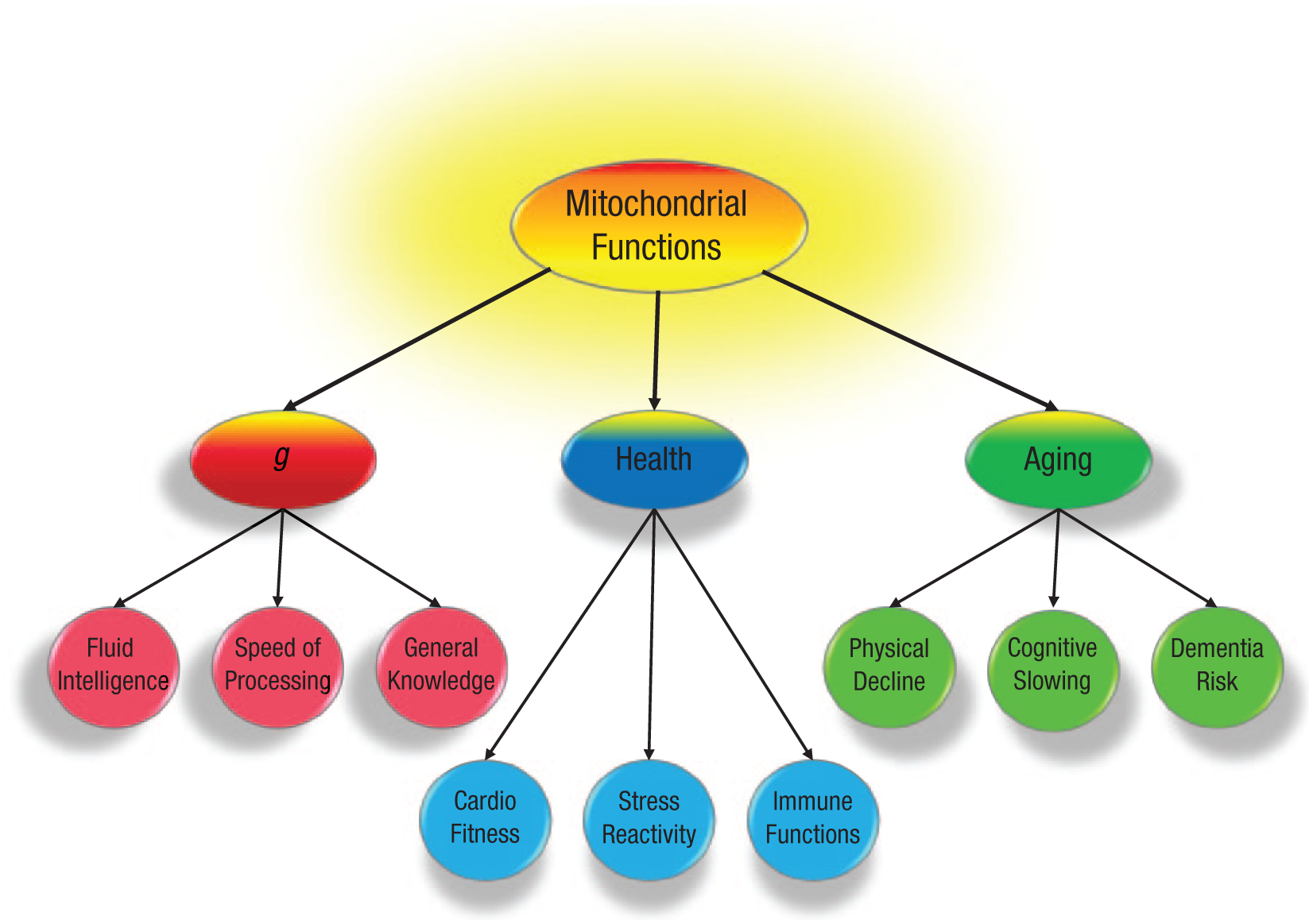
Schematic illustrating the proposed theory that mitochondrial functions are the common factor contributing to cognitive abilities (as indexed by general intelligence, or g), health, and normal aging in adulthood.
Intelligence, Health, and Aging
The relations among intelligence, health, and aging have been well established by Deary and his colleagues (2010). One of the most comprehensive demonstrations of these relations is based on more than 70,000 people who were followed from childhood to the age of 79 years (Cˇukić, Brett, Calvin, Batty, & Deary, 2017). The key finding was that each 1-standard-deviation increase in intelligence at age 11 was associated with about a 20% reduction in the odds of dying before the age of 79. More than half of the individuals in the bottom 20% of intelligence at age 11 had died before age 79, whereas nearly 2 out of every 3 people in the top 20% were still alive at this age. These results reflect a much broader relation between intelligence and general physical health, including indicators as diverse as handgrip strength (Meincke, Hansen, Mortensen, Osler, & Mortensen, 2016) and risk of coronary disease (Batty, Mortensen, Nybo Andersen, & Osler, 2005). These patterns hold with control of socioeconomic status and health-related behaviors and are correlated with cognitive changes associated with normal aging.
Research in cognitive aging suggests nuanced environmental and genetic influences on age-related declines in cognition (Deary, 2000; Salthouse & Ferrer-Caja, 2003; Zavala et al., 2018), as well as larger declines in some domains (e.g., fluid intelligence) than others (e.g., memory for word sequences; Verhaeghen & Salthouse, 1997). Even with environmental influences (e.g., occupational complexity) on cognitive aging, there also seems to be a common biological influence on age-related declines in intelligence, visual and auditory acuity, and sensorimotor skills (e.g., balance), suggesting “aging changes in the physiological state of the brain” (Lindenberger & Baltes, 1994, p. 352). Lubinski and Humphreys (1992) suggested that the relation between cognition and health might be due to “organismic superiority” (p. 111), whereas Deary and colleagues (2010) called this the body-integrity hypothesis: “This idea, which is often rather vaguely articulated . . . demands a search for other possible markers of system integrity—other measurable indicators of bodily and brain efficiency” (p. 63).
Why Mitochondria?
Mitochondria are an ideal candidate for at least partially explaining bodily and brain efficiency, and through this the links among intelligence, health, and aging. Mitochondria carry their own set of DNA that are inherited from the individual’s mother and interact with nuclear DNA to produce the majority of energy that supports cellular functions; they also contribute to immune functions, hormone synthesis, and many other processes (Koch, Josefson, & Hill, 2017). Following conception, the same initial cache of up to 100,000 mitochondria in the unfertilized egg is randomly distributed among the cell lines that will form all of the physical systems in the developing fetus (Stewart, Freyer, Elson, & Larsson, 2008). Potentially deleterious or salubrious variants of mitochondria are thus distributed throughout the body, although tissue-specific configurations eventually emerge. Still, the initial pool of mitochondria will be highly similar in all cell lines and thus could easily contribute to individual differences in the development and functioning of all physical systems. This is one reason mitochondrial functioning is now recognized as a key contributor to overall health (Picard, Wallace, & Burelle, 2016) and to the emergence or progression of a wide array of diseases, including neurodegenerative diseases (Grimm et al., 2016).
One key function of mitochondria is energy production; specifically, the pathway that uses glucose to create ATP. The process is called cellular respiration and involves the transport of energy-carrying electrons within mitochondria. There is individual variation in the efficiency of ATP production and in the tendency for electrons to “escape” during the transport process and bind with oxygen molecules. The latter generate reactive oxygen species (ROS). These ROS are important for a variety of normal cellular functions but at excessive levels can bind to and damage DNA, cell membranes, and proteins needed for many biochemical processes. The damage resulting from excess ROS is called oxidative stress and has been suggested as a potential contributor to the relations among intelligence, health, and aging (Gottfredson & Deary, 2004). Mitochondrial DNA are particularly vulnerable to this stress because they are located near the complexes that create ROS (Brand & Nicholls, 2011). The associated degradation of mitochondria reduces their capacity to produce cellular energy and through this compromises the functioning of the broader system within which these cells are situated.
In other words, there is a cycle whereby energy generation creates ROS, which in turn gradually undermines the subsequent ability to produce energy, among other compromises to mitochondrial functions (e.g., immunity, inflammation). Individual differences in the rate at which this use-it-and-lose-it cycling occurs are influenced by the efficiency of ATP production and control of ROS but gradually contribute to the normal aging process and age-related changes in cognition and health (Harman, 1972; Lane, 2011; Sun, Youle, & Finkel, 2016). Critically, the rate at which this occurs can also be influenced by a variety of environmental factors, such as toxin exposure and chronic psychosocial stress (Caito & Aschner, 2015; Picard et al., 2016). Whatever the cause, declines are often noticed first in energy-demanding systems, such as the brain and heart. Normal functioning can be maintained as long as ATP production is above their basal requirements, but functions can decline precipitously once energy production drops below this threshold or under conditions that require a rapid increase in energy demands. One result is that chronic early stressors or early toxin exposures might not manifest until later in life, when the associated degradation in mitochondrial functioning combines with normal age-related declines to produce accelerated aging.
How Might Mitochondria Contribute to Intelligence?
Human cognitive abilities are hierarchically organized, and Carroll’s (1993) seminal work suggests that they are spread across three meaningful levels. At the lowest level are specific abilities or knowledge bases, and these are unified by higher-order strata that reflect processes common to several specific competencies. Examples of second-order abilities are fluid intelligence, speed of processing, and general knowledge, as can be seen in Figure 1. General knowledge, for instance, can be decomposed into specific domains, such as mathematics or world history, and more basic processes involved in memory and concept formation result in positive correlations among knowledge in these areas. Second-order abilities are unified by g, which explains up to 50% of the covariation among cognitive and academic tests. The essence of g is vigorously debated (e.g., Kovacs & Conway, 2016), is almost certainly multifaceted, and is probably “not a mental or cognitive process or one of the operating principles of the mind, such as perception, learning, or memory” (Jensen, 1998, pp. 94–95).
Indeed, brain-imaging and genetic studies indicate that systems at multiple levels likely contribute to g (Lee et al., 2018; Santarnecchi, Emmendorfer, & Pascual-Leone, 2017; Savage et al., 2018). One can also think of these as being hierarchically organized, as shown in Figure 2. Much of the associated research is focused on networks of brain regions that support executive function, attentional control, and explicit problem solving; these networks are represented by the distributed-brain-networks ring in Figure 2 (Jung & Haier, 2007). Local brain networks allow for fine-grain processing within spatially close brain regions (e.g., visual system) and support sensitivity to subtle variation in perceptual cues (Melnick, Harrison, Park, Bennetto, & Tadin, 2013). Genes related to neuron and glia (cells that support neurons) functioning are also related to g and may contribute to the availability of the neurotransmitters needed for neural communication (Tseng et al., 2015).
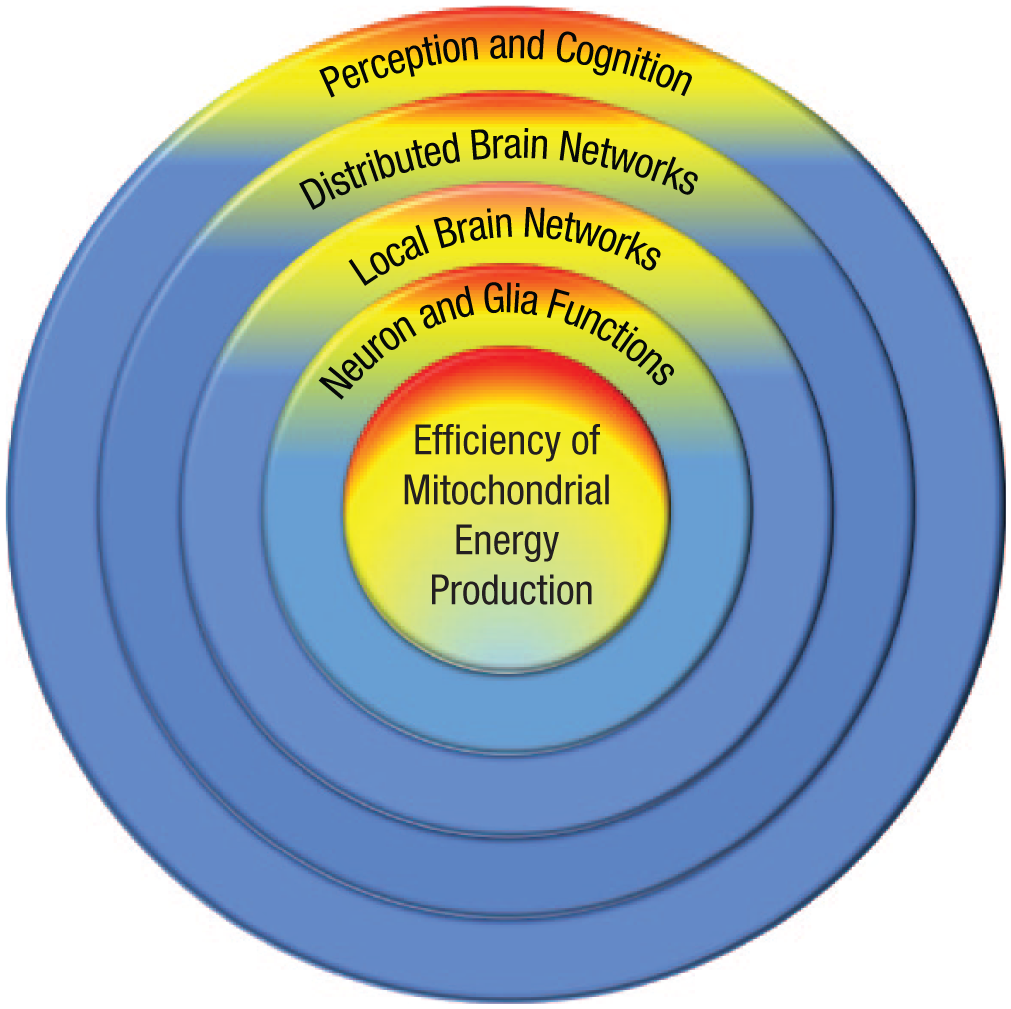
Illustration showing the hierarchy of the multiple cognitive and brain systems that contribute to general intelligence, or g. Deficits at lower biological levels (inner rings) will compromise functioning at higher levels (outer rings), with the efficiency of mitochondrial energy production anchoring higher-level systems and linking them to health and aging.
The brain is one of the most energy-demanding organs in the body, and the lowest common denominator supporting the functioning of all brain systems is the availability of ATP. Zhu et al. (2012), for instance, estimated that the human brain consumes about 5.7 kg of ATP per day; this is more than the weight of the brain itself, a level that is possible because of the recycling of these molecules. Individual neurons at rest consume about 4.7 billion ATPs per second (Zhu et al., 2012), and consumption may increase three and a half times with the propagation of an action potential (Kann & Kovács, 2007). Mitochondria are at the center of the systems shown in Figure 2 because of these exorbitant energy requirements.
There is a potentially important asymmetry in the relations shown in Figure 2, namely, that optimal functioning at outer rings is dependent on the integrity of all of the systems represented by inner rings, but not vice versa. Focal injury to a brain region that contributes to working memory (e.g., dorsolateral prefrontal cortex) would disrupt the ability to engage in systematic problem solving but would not compromise lower-level processes outside of this damaged area. In contrast, systematic declines in mitochondrial energy production, as occurs with normal aging (Sun et al., 2016), will resonate through all levels and ultimately result in declines across perceptual and cognitive domains. Moreover, broad declines in energy availability should result in functional declines that are directly proportional to the complexity of the supporting brain networks (Bullmore & Sporns, 2012). By analogy, consider that it takes more energy to heat a 4,000-square-foot house than a 1,500-square-foot house. A drop in available energy will be noticed first in the larger house and result in a larger drop in ambient temperature relative to the smaller house. In fact, if available energy is sufficient for the latter, then a change in energy availability will go unnoticed. Distributed brain networks are analogous to the larger house, and this might be why age-related declines are larger in more-complex cognitive abilities (e.g., fluid intelligence) than in less-complex cognitive abilities (e.g., verbal recall; Verhaeghen & Salthouse, 1997).
In any case, the importance of mitochondrial functioning for various aspects of health, including risk of neurodegenerative diseases, has motivated the development of methods to better study them (Strack & Usachev, 2017) and interventions to promote their functioning. Given the complexity of mitochondria, developing such treatments is not straightforward, but there are some early promising results (Henderson et al., 2009), as well as many null results (Caldwell, Yao, & Brinton, 2015). Perhaps the latter are not too surprising, given the complexity of mitochondrial functioning and the focus on populations at risk for severe cognitive deficits such as Alzheimer’s disease. These groups are identified after they have suffered some degree of neurological damage at biological levels above mitochondrial functioning (outer rings in Fig. 2), and this damage might not be fully reversible even if energy production is fully restored. We are in the early stages of the development of these types of treatments, but advances in this area may eventually prove useful for the amelioration of environmental and age-related compromises to cognition and health more broadly.
Conclusions and Future Directions
The function or group of functions identified by Spearman (1904) as general intelligence extends, at least in part, beyond the relations among academic and cognitive domains into many facets of general health, disease risk, and successful aging in adulthood (Deary et al., 2010; Lubinski & Humphreys, 1992). To date, identification of the factors that link these disparate domains has proven elusive. The proposal here is that the multiple functions of the evolutionarily ancient mitochondria, including cellular energy production (Lane, 2011), may provide at least part of this link.
Indeed, Picard et al. (2016) placed mitochondrial functioning at the heart of cellular and physiological health and at the core of many physical, psychiatric, and neurological diseases, as well as normal aging. It is only a small step to incorporate g into this nexus of relations. The approach may be useful for identifying genetic and environmental influences on the development of intelligence and rate of age-related declines in cognition, and the emerging mitochondrial therapies for specific diseases (e.g., Alzheimer’s) might prove to be broadly useful, specifically, for the amelioration of environmental and age-related compromises to health and cognition.
Recommended Reading
Deary, I. J., Whiteman, M. C., Starr, J. M., Whalley, L. J., & Fox, H. C. (2004). The impact of childhood intelligence on later life: Following up the Scottish mental surveys of 1932 and 1947. Journal of Personality and Social Psychology, 86, 130–147. Provides a description of the Scottish Mental Surveys and the relation between intelligence in childhood and later health
Geary, D. C. (2018). Efficiency of mitochondrial functioning as the fundamental biological mechanism of general intelligence (g). Psychological Review, 125, 1028–1050. Provides a detailed discussion of the relations among mitochondrial functioning and intelligence, health, and aging and discussion of the sex difference in variability in intelligence.
Picard, M., Juster, R. P., & McEwen, B. S. (2014). Mitochondrial allostatic load puts the ‘gluc’ back in glucocorticoids. Nature Reviews Endocrinology, 10, 303–310. Provides a review of how chronic stress can result in mitochondrial dysfunction and compromised health.
Raz, N., & Daugherty, A. M. (2018). Pathways to brain aging and their modifiers: Free-radical-induced energetic and neural decline in senescence (FRIENDS) model – A minireview. Gerontology, 64, 49–57. Provides a review of agerelated changes in brain functions and their potential relation to declines in mitochondrial functioning.