
Abstract
Background
To investigate whether the mechanism underlying the anti-inflammatory effects of electroacupuncture (EA) at ST36 involves dopamine (DA) and its receptor and whether it is mediated by the vagus nerve in a rat model of intestinal ischaemia-reperfusion (I/R) injury.
Methods
Rats were subjected to gut ischaemia for 30 min and then received EA for 30 min with or without abdominal vagotomy or intraperitoneal administration of butaclamol (D1 receptor antagonist) or spiperone (D2 receptor antagonist). Plasma levels of DA and tumour necrosis factor (TNF)-α were assessed 1 or 4 h after reperfusion. Myeloperoxidase (MPO) activity and malondialdehyde (MDA) content in intestinal tissues were assessed using enzyme-linked immunosorbent assay (ELISA) kits. Intestinal tissue injury was assessed by observation of the pathological lesions and permeability to 4 kDa fluorescein isothiocyanate (FITC)-dextran.
Results
EA significantly increased levels of DA and lowered levels of TNF-α. EA also inhibited intestinal levels of MPO and MDA and intestinal tissue injury and decreased intestinal permeability to FITC-dextran. Abdominal vagotomy and intraperitoneal administration of butaclamol (but not spiperone) inhibited the effects of EA.
Conclusion
These findings suggest that EA at ST36 could attenuate intestinal I/R-induced inflammatory injury and that the underlying mechanism may involve EA-induced increases in levels of DA, mediated by the vagus nerve and D1 receptors.
Keywords
Introduction
Intestinal ischaemia-reperfusion (I/R) is a common form of organ damage in surgical practice, which can be caused by acute blood loss, shock, disseminated intravascular coagulation (DIC), intestinal obstruction, ischaemic disease of the mesenteric vessels, severe or multiple trauma and other factors. Studies have shown that intestinal I/R not only causes local tissue damage in the gastrointestinal tract, but that it can also cause a series of reactive changes in the monocyte/macrophage system due to the displacement of bacteria and toxins after intestinal barrier dysfunction, and release a large number of inflammatory mediators and cytokines, even causing multiple organ dysfunction syndrome (MODS).1,2 Thus, clinical and basic scientific research into the mechanisms underlying intestinal I/R injury and potential preventive measures is important. However, to date, there is no generally accepted, effective pharmacological or non-pharmacological treatment option to reduce intestinal I/R injury.
Animal experiments and clinical studies have shown that acupuncture can significantly improve immunity and attenuate rising levels of pro-inflammatory factors during the inflammatory response.3–5 Previous studies have shown that acupuncture at ST36 (Zusanli) has a regulatory effect on gastrointestinal function and gastrointestinal tissue inflammation6–8 and that these effects involve the vagus nerve.6,7,9 A growing literature reports that electroacupuncture (EA) at ST36 activates the cholinergic anti-inflammatory pathway and has anti-inflammatory effects.10–12 Moreover, recent studies suggest that the anti-inflammatory effect of EA at ST36 is related to the activation of dopamine (DA) receptors independent of cholinergic pathway stimulation. However, whether EA can suppress intestinal ischaemia-induced inflammatory injury through stimulation of DA receptors is unclear.
We hypothesised that EA at ST36 exerts an anti-inflammatory action by activating the vagus nerve, stimulating the adrenal gland to release DA and inhibiting the release of cytokines. We used a rat model of intestinal I/R injury to investigate the putative protective effect of EA at ST36 and determine whether it is related to the excitement or blockade of DA or its receptors, to further explore the foundation for EA treatment of ischaemic and inflammatory diseases.
Methods
Animals
A total of 84 male Sprague-Dawley (SD) rats (12 weeks old, weighing 250 ± 20 g) were purchased from the Experimental Animal Centre of Military Medical Sciences of the Chinese People’s Liberation Army (PLA). The rats were housed in mesh cages in a room maintained at 25°C under a 12/12 h light–dark cycle. Rats were fasted overnight but allowed free access to water up to 4 h before surgery. All research protocols were approved by the Committee of Scientific Research of the First Hospital Affiliated with the General Hospital of PLA, China. The experiment was conducted in compliance with the Guide for Care and Use of Laboratory Animals of the National Research Council, China.
Intestinal ischaemia model
Rats were anaesthetised by injection of pentobarbital sodium (50 mg/kg; Sigma, USA), and the abdomen was shaved and washed with 10% povidone iodine. The rats were secured on a heating pad to maintain appropriate body temperature during anaesthesia. A laparotomy was performed to expose the superior mesenteric artery (SMA), across which a microvascular clip was placed for 30 min. Thereafter, the clip was removed to allow reperfusion for 1 or 4 h, and the animals were randomly assigned to experimental groups (see the following). 13
Neurectomy
Rats were anaesthetized and then the abdomen and posterolateral femora were shaved and washed with 10% povidone iodine. A 2 cm upper midline laparotomy was performed to identify the gastroesophageal junction and expose the dorsal and ventral vagus nerve on the distal oesophagus using a Phenix XLT165-LB stereomicroscope (Phenix Optical Instrument Group Company, China).
EA at ST36
Both hind limbs were shaved and the skin was disinfected. EA stimulation of both limbs was performed by inserting unipolar stainless steel needle electrodes (Huatuo, China), with a diameter of 0.25 mm and length of 12 mm, at ST36 bilaterally, advancing them to a depth of 7 mm and connecting them to an EA apparatus (HANS, LH202H). Electrical stimulation was applied using a continuous pulse mode for 30 min with an intensity of 2 mA, pulse width of 50 µs and frequency of 10 Hz.
Experimental groups and treatments
A total of 84 SD rats were randomly divided into the following seven groups: sham (S) group, sham + EA (SE) group, intestinal I/R (IR) group, I/R + EA (EA) group, I/R + EA + vagotomy (VX) group, I/R + EA + butaclamol (BC) group and I/R + EA + spiperone (SP) group. According to two different observation times, each group was divided into two subgroups: 1 and 4 h after injury (n = 6 each) for detection. The I/R model was established by SMA occlusion for 30 min, followed by reperfusion for 1 or 4 h. EA stimulation at bilateral ST36 was performed in EA, VX, BC and SP groups for 30 min as described earlier. Rats in the sham group underwent laparotomy without SMA occlusion or other manipulation. Rats in the SE group underwent laparotomy without SMA occlusion but received EA. Rats in the EA group received EA immediately after ischaemia, and those in the model group received no treatment. Rats in the VX group underwent vagotomy of the dorsal and ventral vagus nerve on the distal oesophagus prior to EA at ST36 immediately after the ischaemia. Rats in the BC group underwent intraperitoneal (i.p.) administration of butaclamol (1 µg/kg, Sigma; a standard D1 receptor antagonist) before 1 h ischaemia. Rats in the SP group underwent i.p. administration of spiperone hydrochloride (1 mg/kg, Sigma; a D2 receptor antagonist) before 1 h ischaemia.
Blood and intestinal tissue sampling
Rats were terminally anaesthetised with 2% isoflurane inhalation and sacrificed by exsanguination via the abdominal aorta at 1 or 4 h after reperfusion. Systemic blood was drawn by abdominal aortic puncture and segments of intestine were harvested and immediately homogenised on ice with 1 mL denaturing or non-denaturing lysis buffer for enzyme-linked immunosorbent assay (ELISA).
Detection of DA and TNF-α levels in plasma
Systemic blood obtained by abdominal aortic puncture was anti-coagulated with heparin and centrifuged to yield plasma. DA and tumour necrosis factor (TNF)-α levels in the plasma were assessed using commercially available ELISA kits in accordance with the protocols provided by the manufacturer (Cloud-Clone Corp., USA; Nanjing Jiancheng Corp., China).
Determination of intestinal myeloperoxidase and malondialdehyde activity
Intestinal myeloperoxidase (MPO) activity was determined using an MPO fluorometric activity assay kit (Sigma-Aldrich, USA) according to the manufacturer’s instructions. The activity of MPO in the intestine was calculated according to the following formula: MPO (U/weight grammes) = (determination optical density (OD) value–control OD value)/11.3 × sample volume (g). Intestinal malondialdehyde (MDA) was determined using an MDA content kit (Nanjing Jiancheng Corp., China) according to the manufacturer’s instructions. The content of MDA in the intestine was calculated according to the following formula: MDA (nmol/mgProt) = {((determination tube absorbance – blank tube absorbance)/(standard tube absorbance – blank tube absorbance)) ×standard concentrations}/protein content.
Intestinal epithelial permeability
Animals were anaesthetised with inhaled 2% isoflurane 1 or 4 h after reperfusion. A midline laparotomy was performed and a 5 m segment of distal ileum was isolated between silk ties. A 500 µL solution containing 4-kDa FITC-dextran (25 mg/mL; Sigma, USA) was injected into the lumen of the isolated segment of intestine. The bowel was returned to the abdominal cavity and the abdomen was closed; 30 min after injection of the FITC-dextran, systemic blood was drawn by abdominal aortic puncture. Plasma fluorescence was measured using a fluorescence spectrophotometer (Synergy 2 Multi-detection Microplate Reader; Biotek, USA).
Histopathology
Segments of the distal ileum were fixed in 10% buffered formalin, embedded in paraffin and sectioned. Haematoxylin and eosin staining of the intestine was performed.
Statistical analysis
Data were analysed using the Statistical Package for the Social Sciences (SPSS) version 17.0 (SPSS Inc., Chicago, IL, USA), and all results were expressed as mean ± SEM. One-way ANOVA was used to compare across all groups and post hoc Student–Newman–Keuls (SNK) tests were used to examine pairwise comparisons. Differences were considered to be statistically significant when p ≤ 0.05.
Results
EA at ST36 increased plasma DA concentration
Figure 1 illustrates the plasma concentrations of DA, which were significantly decreased in the IR group compared with the S and SE groups at 1 h (21.4 ± 1.92 vs 36.7 ± 4.35 and 38.1 ± 3.25 pg/mL, respectively; p < 0.001) and 4 h (6.8 ± 0.81 vs 49.8 ±8.62 and 43.9 ± 5.35 pg/mL, respectively; p < 0.001). Meanwhile, compared with the IR group, EA at ST36 significantly restored the level of DA at 1 h (28.7 ± 1.89 vs 21.4 ± 1.92 pg/mL; p < 0.001) and 4 h (10.6 ± 1.02 vs 6.8 ± 0.81 pg/mL; p < 0.001). However, the effect of EA was abolished in the VX and BC groups. DA levels in the SP group were equivalent to the EA group (p > 0.05). These findings suggest that EA at ST36 increases DA concentration after intestinal I/R injury via activation of the vagus nerve and D1 receptors. DA concentration in the IR4h subgroup was reduced to a further degree than that in the IR1h group, but was similarly inhibited by EA.
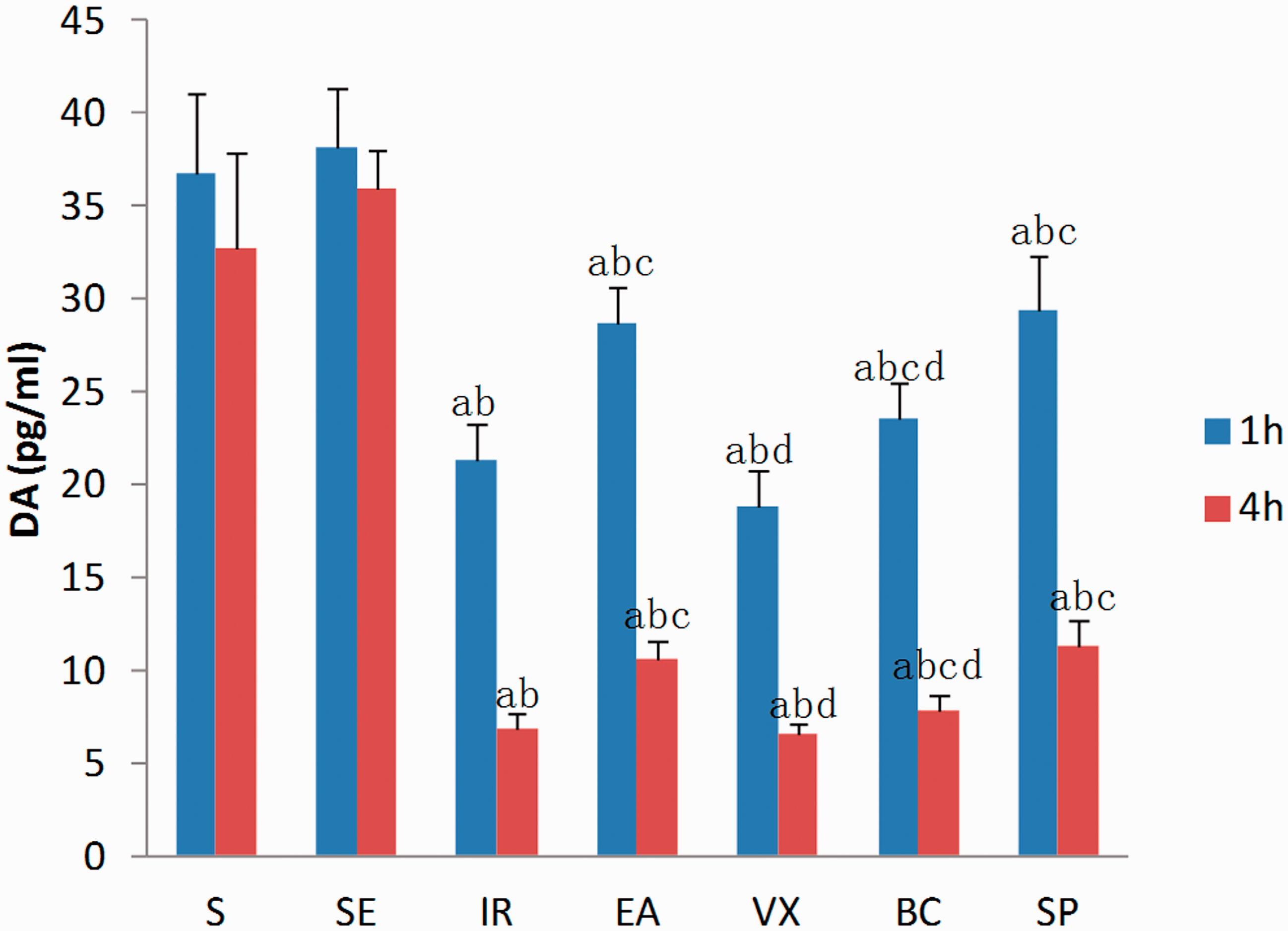
Plasma concentration of dopamine (DA), expressed as pg/mL, at 1 and 4 h after ischaemia-reperfusion (I/R).
EA at ST36 lowered TNF-α levels in plasma
Plasma levels of TNF-α were low in the S group and SE group (Figure 2). The level of TNF-α was significantly elevated after intestinal I/R injury. Serum TNF-α concentration was maximal at 1 h and significantly reduced at 4 h following I/R. EA lowered TNF-α levels in plasma (p < 0.05). The abscission of the vagus nerve and use of a D1 receptor antagonist inhibited the effect of EA. There were no statistically significant differences between the SP group and EA group (p > 0.05). These findings suggest that EA at ST36 decreases TNF-α concentration after intestinal I/R injury by activating the vagus nerve and that the effect of EA may be mediated through the D1 receptor.
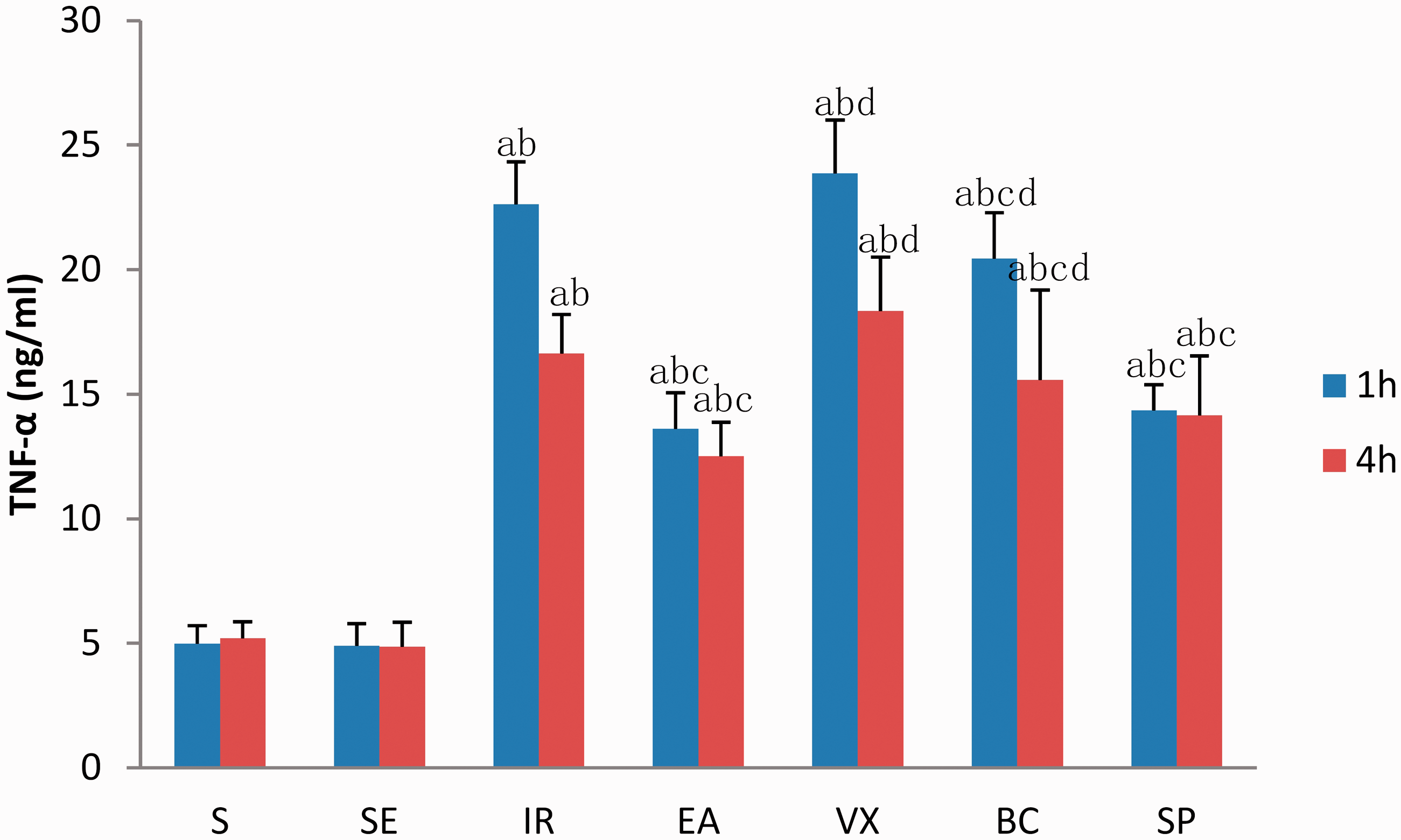
Plasma concentration of tumour necrosis factor (TNF)-α, expressed as ng/mL, at 1 and 4 h after ischaemia-reperfusion (I/R).
EA at ST36 decreased intestinal MPO and MDA
Table 1 illustrates the activity of MPO and MDA in the intestine. Compared with the IR group, EA at ST36 inhibited intestinal activity of MPO at 1 and 4 h (both p < 0.001) and MDA at 4 h (IR4h vs EA4h p < 0.001) but not 1 h (IR1h vs EA1h p > 0.05) after I/R injury, and the effect was more obvious at 4 h (37% and 39% reductions for MPO and MDA, respectively) than at 1 h (27% and (non-significant) 9% reductions for MPO and MDA, respectively). MDA activity in the IR4h subgroup was 1.4 times that of the IR1h subgroup. By contrast, MDA activity in the EA4h subgroup was only 94% of that in the EA1h subgroup. These results indicate that EA could inhibit the activity of MDA and MPO, with the largest protective effect at the earlier time point. The effect of EA weakened after abdominal vagotomy or i.p. administration of butaclamol. By contrast, i.p. administration of spiperone had no effect. Accordingly, we suspect that the effects of EA were related to the vagus nerve and D1 receptors.
Intestinal activity of myeloperoxidase and malondialdehyde activity.
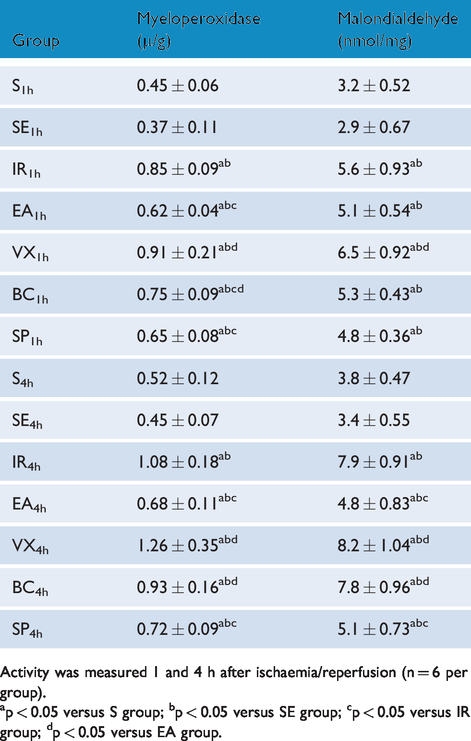
Activity was measured 1 and 4 h after ischaemia/reperfusion (n = 6 per group).
ap < 0.05 versus S group; bp < 0.05 versus SE group; cp < 0.05 versus IR group; dp < 0.05 versus EA group.
EA at ST36 lowered intestinal permeability
Animals in the IR group had an increase in permeability, when compared with sham-operated animals. EA at ST36 protected against increased intestinal permeability (EA vs IR group: 66.6 ± 8.19 vs 221.9 ±22.51 ng/mL at 1 h; 386.7 ± 12.31 vs 611.4 ±28.79 ng/mL at 4 h; Figure 3). However, when abdominal vagotomy was performed or butaclamol was administered before reperfusion, the intestinal permeability of animals undergoing EA at ST36 was indistinguishable at 1 h (235.6 ± 13.26 and 231.8 ± 21.6 vs 66.6 ± 8.19 ng/mL) and 4 h (604.6 ± 21.67 and 635.8 ± 25.45 vs 386.7 ± 12.31 ng/mL). These data indicate that EA at ST36 can only offer protection to the gut in the presence of intact neuroenteric innervation and that EA might exert this effect via the D1 receptor.
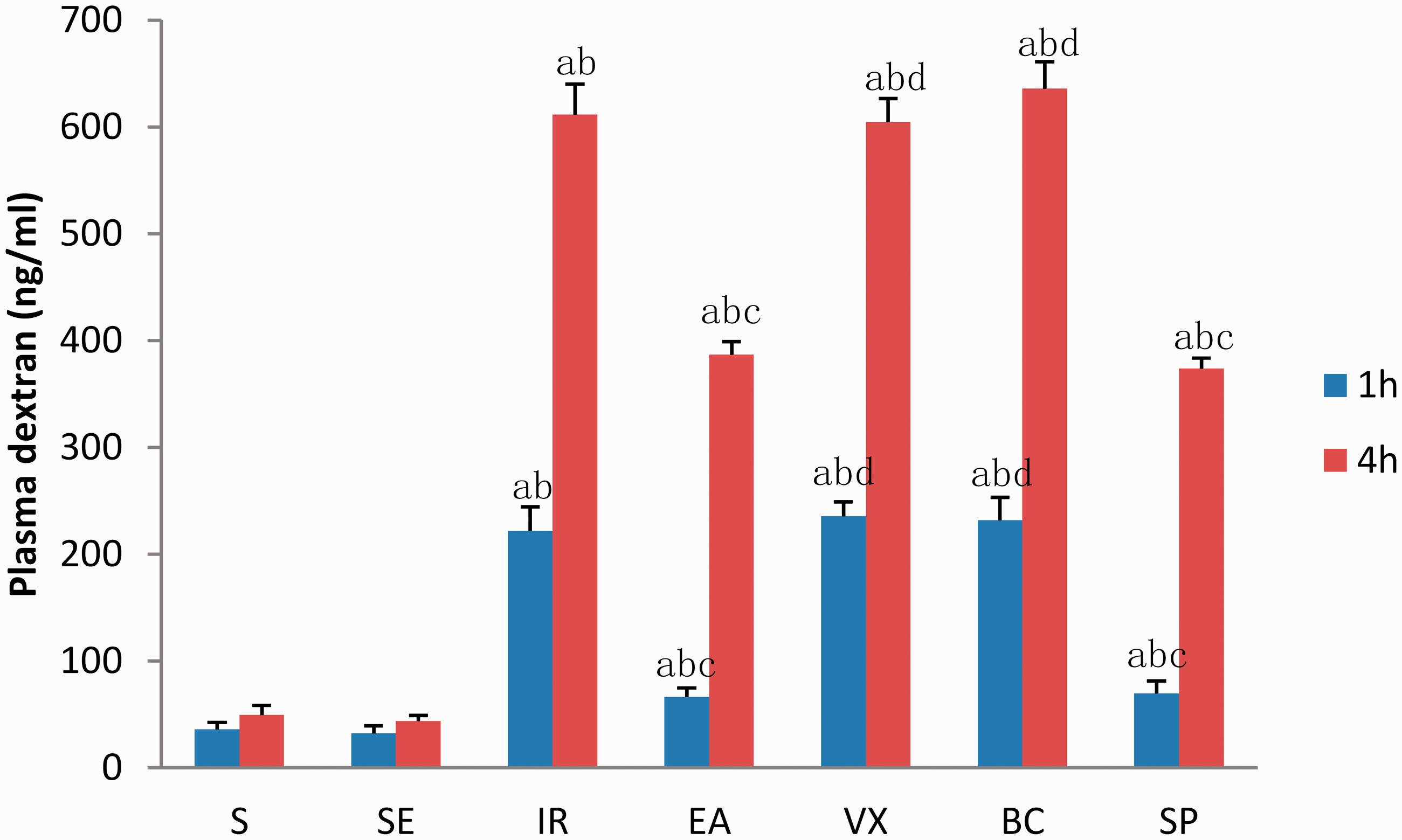
Intestinal permeability to 4-kDa fluorescein isothiocyanate (FITC)-dextran at 1 and 4 h after ischaemia-reperfusion (I/R).
EA at ST36 decreases intestinal injury
In the S and SE groups, intestinal epithelial villi exhibited clear and complete structure with no defects, epithelial shedding or clastic formation (Figure 4, S and SE). At 4 h after I/R injury, oedema, haemorrhage, atrophy and tissue disorders were observed in the pathological sections of intestine. Interstitial villi demonstrated inflammatory cell infiltration in great quantities and villous epithelial cells had become necrotic or fallen off (Figure 4, IR). Compared to the model group, the injury observed in the EA group was subjectively much milder. In the EA group, intestinal epithelial villi had clear and complete structures with no defects (Figure 4, EA). The effect of EA in the VX and BC groups was weakened (Figure 4, VX and BC) and pathological sections from the SP group were not obviously different compared with the EA group (Figure 4, SP).
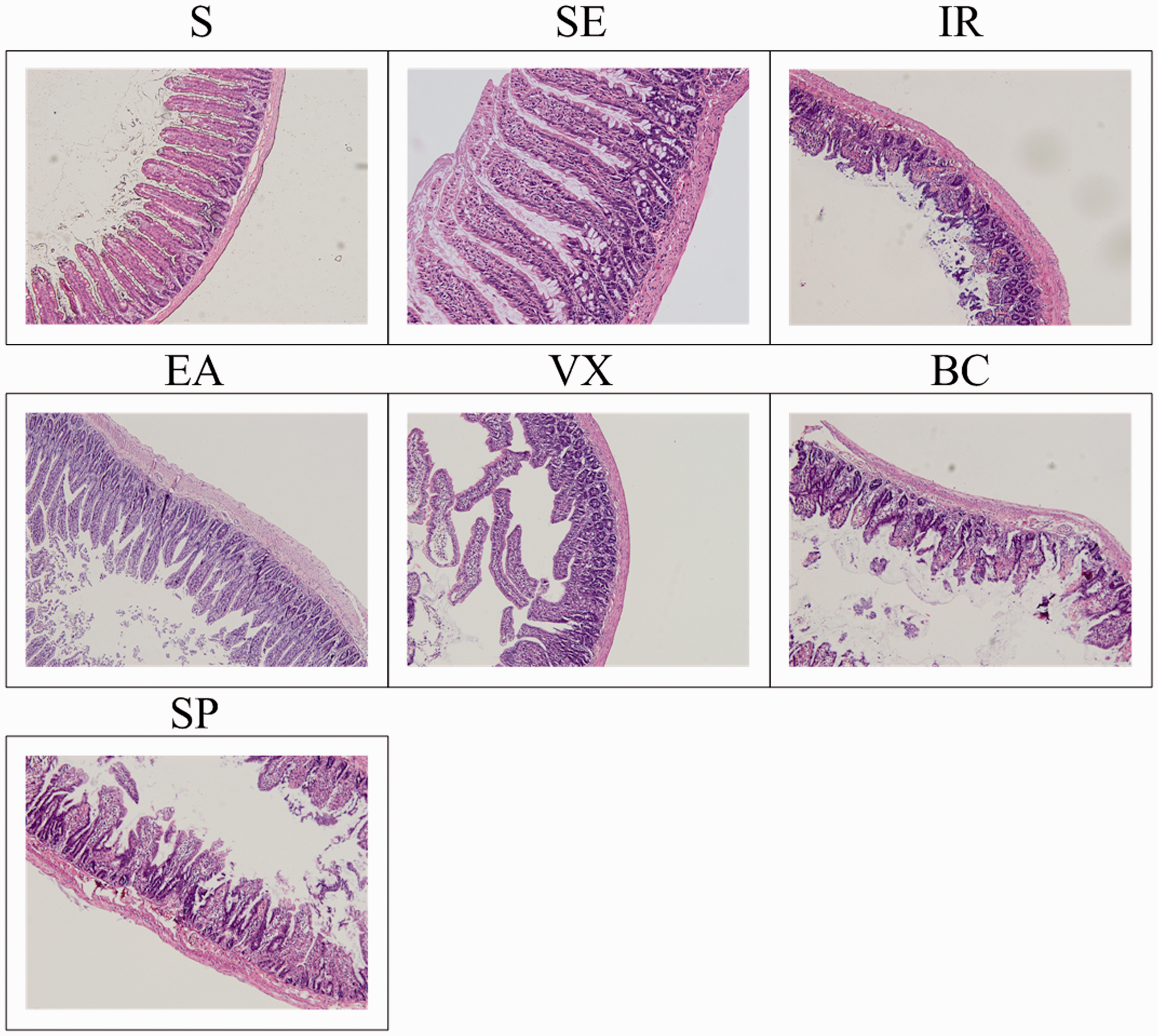
Representative intestinal histology at 4 h after ischaemia-reperfusion (I/R). All images were taken at 200× magnification. Black scale bar = 5 µm.
Discussion
Normal intestinal mucosa has strong mechanical, immune and biological barrier functions. Intestinal I/R may not only lead to intestinal ischaemia and mucosal cell damage, but also to the systemic inflammatory response syndrome (SIRS) and MODS.14,15 Experiments have shown that EA at ST36 could inhibit synthesis of TNF-α, reduce immunosuppression caused by trauma, and inhibit sepsis, peritonitis and other inflammatory reactions in animal models. Studies have shown that EA at ST36 has an influence on the vagal central nuclei – the vagus nerve dorsal nucleus, the solitary nucleus and nucleus ambiguus. We have performed a large number of experiments to show that the anti-inflammatory effects of EA at ST36 are related to the vagus nerve.11,16,17 The latest research suggests EA at ST36 plays an anti-inflammatory role by exciting the vagus nerve, adjusting the adrenal medulla to release DA and thereby inhibiting the production of cytokines. 18 Previous studies have confirmed that DA and its receptor have anti-inflammatory effects.18,19 Furthermore, DA can alleviate the inflammatory response and damage caused by I/R,20–22 the anti-inflammatory effect of which is achieved by regulating the secretion of inflammatory cytokines, including TNF-α, interleukin (IL)-1β and IL-6. 18
DA is a neurotransmitter that regulates the function of multiple tissues. DA acts as an agonist at DA receptors, which are divided into D1-like receptors (D1 and D5) and D2-like receptors (D2, D3 and D4). 17 Previous studies have confirmed that DA and its receptors exert anti-inflammatory effects, that DA can alleviate the damage caused by I/R and inflammation, and that its anti-inflammatory effect is achieved by regulation of the secretion of inflammatory cytokines, including TNF-α, IL-1β and IL-6. Intracellular cyclic adenosine monophosphate (cAMP) can inhibit the formation of TNF complexes.23,24 Stimulation of D1 receptors can increase intracellular cAMP levels, and the stimulation of D2 receptors can decrease intracellular cAMP levels. DA pretreatment can reduce the damage caused by kidney I/R.25,26
TNF-α is a key factor leading to multiple organ damage after intestinal I/R. 27 It appears early in the cycle, peaks quickly, induces the generation and release of a variety of secondary cellular cytokines, such as IL-1, IL-6 and IL-8, and then forms the so-called ‘waterfall effect’. TNF-α not only directly results in tissue damage but can also lead to the generation of oxygen-free radicals, which in turn lead to the formation of a cell membrane lipid peroxide and its degradation product, MDA. MDA content is a good index reflecting oxygen-free radical damage. 28 MPO is a specific enzyme found in polymorphonuclear leukocytes, and the increase of MPO activity in tissue prompts the production of oxygen-free radicals in great numbers. 29 In this experiment, we found that EA at ST36 can significantly reduce TNF-α concentration and the activity of MPO and MDA, and it could lower the serum TNF-α concentration to exert an anti-inflammatory role and thus protect visceral function.
In our experiments, we found that EA at ST36 has anti-inflammatory and organ protective effects. Compared with the IR group, EA increased DA levels and inhibited the production of TNF-α, which reduced MDA and MPO activity, and exerted a protective effect on the intestine. After vagotomy or i.p. administration of butaclamol, a standard D1 receptor antagonist, the protective effect of EA weakened or disappeared, thus we consider the protective effect of EA may be mediated through the vagus nerve and D1 receptor. These results are consistent with the latest research, which suggests that EA at ST36 plays an anti-inflammatory role by exciting the vagus nerve and adjusting the adrenal medulla to release DA and thereby inhibit the production of cytokines. 18 Acetylcholine is the principal neurotransmitter of the vagus nerve and vagal immune modulation requires the α7 nicotinic acetylcholine receptor (α7nAChR),30,31 and yet it is possible that α7nAChR is not required for EA-induced inhibition of serum TNF-α levels or induction of DA. Overall, this study explored the anti-inflammatory effects of EA at ST36 through DA-related mechanisms and provides a theoretical foundation for the EA treatment of ischaemic and inflammatory diseases.
In conclusion, when severe injury occurs, the blood supply to the intestinal tract is sharply reduced, which results in gut barrier dysfunction that may lead to SIRS and MODS. Previous research has demonstrated the anti-inflammatory and protective effects of EA on intestinal tissue, but the mechanism is unclear. This experiment indicates that the anti-inflammatory effects of EA at ST36 are related to the vagus nerve and D1 receptors. Activation of DA receptor may be a new mechanism of action underlying the inhibitory effect of EA on serum TNF-α levels besides α7nAChR. Further studies are needed to explore the mechanisms by which EA inhibits serum TNF-α levels and induces DA.
Footnotes
Contributors
YML and GCX contributed equally to this study. SH, YML and GCX designed the research. YML, YLD, HML, WHX and FBT performed the research. SH, YML and HW analysed the data. YML, SH, XS and HW prepared and revised the manuscript. All authors have read and approved the final version of the manuscript accepted for publication.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This study was supported by the National Natural Science Foundation of China (grant no. 81774422).
Ethical approval
This study received ethical approval from Institutional Animal Care and Use Committee of the First Hospital Affiliated to the Chinese PLA General Hospital.
Licence Statement
The Corresponding Author has the right to grant on behalf of all authors and does grant on behalf of all authors, an exclusive licence (or non-exclusive for government employees) on a worldwide basis to permit this article (if accepted) to the published in AiM editions and any other BMJPGL products to exploit all subsidiary rights, as set out in our licence (![]() ).
).
Provenance and peer review
Not commissioned; externally peer reviewed.