
Abstract
Objective:
This aim of the study was to investigate whether electroacupuncture (EA) can mitigate osteoporosis (OP) in an ovariectomy (OVX)-induced rat model by affecting the ferroptosis-related pathway.
Methods:
Thirty-two 12-week-old female Sprague-Dawley rats were randomly allocated into four groups: CONTROL, SHAM, OVX and EA. We examined the effects of OVX and EA on markers of bone resorption and formation, iron content, malondialdehyde (MDA) levels, and the expression of ferritin heavy chain 1 (FTH1), solute carrier family 7 member 11 (SLC7A11) and glutathione peroxidase 4 (GPX4). In addition, femoral morphology and immunofluorescence were analysed.
Results:
The OVX group exhibited a significant increase in bone resorption markers, iron and MDA levels, with a notable decrease in alkaline phosphatase (ALP) levels, FTH1, SLC7A11 and GPX4 expression, and impaired trabecular architecture. EA significantly reduced bone resorption markers, iron and MDA content while substantially elevating ALP levels and FTH1, SLC7A11 and GPX4 expression. At the same time, EA improved trabecular architecture.
Conclusion:
EA ameliorated OVX-induced OP by decreasing iron accumulation and activating the SLC7A11/GPX4 axis.
Introduction
Osteoporosis (OP) is a systemic metabolic bone disease. With the continuous acceleration of global population ageing, both the incidence of OP and the resultant public health burden have shown a significant upward trend.1–4 In the face of this escalating public health challenge, pursuing highly effective and safe strategies for prevention and treatment has become a crucial global healthcare concern.
As an age-related degenerative disease, OP is fundamentally driven by an imbalance between bone formation and resorption. 5 This metabolic disequilibrium is heavily influenced by factors such as sex hormone deficiency, calcium loss and impaired vitamin D homeostasis.6,7 Current treatment strategies include both nonpharmacological and pharmacological approaches, but each has limitations in efficacy, safety concerns and individual variability.8,9 There is a vital need for safe and effective alternative and complementary interventions.
Multiple meta-analyses have confirmed the beneficial effects of acupuncture in the treatment of OP.10,11 Furthermore, its clinical value has been demonstrated in improving early-stage osteopenia 12 and alleviating pain from osteoporotic fractures 13 through a unique holistic regulatory approach. Mechanistic studies in postmenopausal animal models have demonstrated that acupuncture effectively mitigates osteoporotic bone loss by modulating key signaling pathways, such as Wnt/β-catenin and OPG/RANKL, while simultaneously restoring estrogen levels and regulating histone acetylation.14–17 However, the exact mechanisms of action underlying the beneficial effects of acupuncture on OP remain unclear.
Recent studies have shown that ferroptosis is associated with osteoblast dysfunction and abnormal osteoclast activity. 18 This form of cell death is characterised by iron dependence, lipid peroxidation and disruptions in the antioxidant defence system, primarily involving solute carrier family 7 member 11 (SLC7A11) and glutathione peroxidase 4 (GPX4). Ferritin heavy chain 1 (FTH1) manages the oxidation and storage of Fe2+, while malondialdehyde (MDA) is a product of lipid peroxidation.19–21 Research indicates that natural compounds can inhibit ferroptosis by targeting the SLC7A11/GPX4 axis, or reducing iron overload, aiding in the reduction of bone loss in osteoporotic rats.22,23
Accordingly, we hypothesised that electroacupuncture (EA) may ameliorate OP by modulating ferroptosis-related pathways. The aim of this study was to test whether an EA intervention can mitigate bone loss by regulating ferroptosis-related pathways using the ovariectomy (OVX)-induced Sprague-Dawley (SD) rat model of OP, and potentially provide evidence for its application in OP.
Methods
Animals and grouping
All procedures in this study received approval from the Experimental Animal Welfare and Ethics Committee of Nanjing Agricultural University (PZW2025020). Thirty-two female SD rats (12 weeks old and weighing 200 ± 20 g) were acquired from Speidord Biotechnology Co. Ltd (Suzhou, China). The rats were housed in a specific pathogen-free laboratory under controlled conditions (22 ± 2°C, 50%–60% humidity, 12 h light/dark cycle) and had access to standard feed and water. The rats were divided into four groups (n = 8 each): the CONTROL group (that received no intervention), the SHAM group (that was subjected to a dorsal incision with adipose tissue excision only), the OVX group (that underwent bilateral OVX for OP modelling) and the EA group (that underwent OVX surgery followed by EA).
Electroacupuncture intervention
Rats in the CONTROL, SHAM and OVX groups were fed routinely for 12 weeks without intervention, while the EA group received EA treatment using 0.25 mm × 25 mm stainless steel needles (Suzhou Medical Appliance Factory Co. Ltd, Suzhou, China) (Figure 1(A)) at the following sets of traditional acupuncture point locations: set A (CV4 and SP6) and set B (BL23 and ST36) (Figure 1(B)). For set A, the electrodes were connected to the needles inserted at CV4 and unilateral SP6 (alternating sides). For set B, the electrodes were connected to the needles inserted at ipsilateral BL23 and ST36 on both sides. Treatments alternated daily between set A and set B for 20 min over 5 days, followed by 2 rest days. EA was administered with a device set to a stimulation intensity of 2 mA and a frequency of 2/15 Hz (alternating every 3 s). The pulse width was automatically adjusted by the device within a range of 0.2–0.6 ms to match the output frequency. The depth of needle insertion was determined based on the thickness of the skin and subcutaneous fat of the rats. Specifically, needles were inserted into the muscle layer at ST36 (5 mm), while a relatively shallower depth was applied at SP6 (3 mm) due to the limited subcutaneous tissue. For CV4 and BL23, the insertion depth (4 mm and 3 mm, respectively) was carefully controlled to avoid potential injury to the underlying internal organs.

Experimental schedule. (A) Bilateral ovariectomy (OVX) for the osteoporosis model was established on day 8 and electroacupuncture (EA) treatment was administered from weeks 3 to 14. (B) Schematic representation of traditional acupuncture point locations. For set A, electrodes were paired between CV4 and unilateral SP6 (alternated daily). For set B, electrodes were connected between ipsilateral BL23 and ST36.
Enzyme-linked immunosorbent assay
After the treatment was completed, the rats were anaesthetised with 3% sodium pentobarbital (0.1 mL/100 g), and blood was drawn from the abdominal aorta. After standing, the serum was separated by centrifugation at 1000g for 10 min at 4°C. Bone γ-carboxyglutamic acid (Gla) protein (BGP), procollagen type I N-terminal propeptide (PINP) and C-terminal telopeptide of type I collagen (β-CTx) were measured using enzyme-linked immunosorbent assay (ELISA) kits (JX-6757A, JX-6614A1 and JX-7079A1, respectively; Junhang Biotechnology, Yancheng, China). Absorbance was read at 450 nm, with data expressed as ng/mL based on standard curves. Following blood collection, the rats were euthanised by exsanguination, and femurs and tibias were isolated for further analysis.
Micro-computed tomography
Quantitative analysis of bone microstructure was performed on the fixed femoral samples using a micro-computed tomography (micro-CT) system (SkyScan 1176, Bruker, Kontich, Belgium). The region of interest (ROI) was defined as a 2-mm segment of the distal femur, starting 2 mm proximal to the growth plate and extending along the long axis of the bone. CTAn software (v.1.20, Bruker, Kontich, Belgium) was used to quantitatively analyse the ROI, calculating bone volume/total volume (BV/TV), trabecular separation (Tb.Sp), trabecular number (Tb.N), bone mineral density (BMD) and structure model index (SMI). Subsequently, three-dimensional reconstruction and visualisation were performed using CTVox software (v.3.3.0, Bruker).
Colorimetric assay
Serum and tibial tissue iron ion levels were measured using a colorimetric assay. Iron ion levels were determined using an iron ion assay kit (AKIC001M, Boxbio, Beijing, China) following the manufacturer’s instructions. Absorbance was read at 520 nm, and the results were expressed in μmol/L.
Thiobarbituric acid assay
After grinding and homogenising the tibial tissue, the supernatant was collected and MDA content was determined using an MDA assay kit (S0131S, Beyotime Biotechnology, Shanghai, China), following the manufacturer’s protocol. Absorbance was measured at 532 nm. The content was calculated using a standard curve and expressed as μmol/g.
Bioinformatics analysis
Gene expression profiles were obtained from the NCBI Gene Expression Omnibus (GEO, accession number: GSE68313), a publicly available database. The GEO dataset GSE68313 included samples from 11 SHAM-operated and 12 ovariectomized rats. Data underwent background correction, quantile normalisation and logarithmic transformation. Differential expression analysis was conducted with the limma R package, using a threshold of 1.5-fold change and a false discovery rate (FDR) < 0.05. Pathway enrichment analysis for differentially expressed genes (DEGs) was performed through the clusterProfiler R package, focusing on ferroptosis-related genes sourced from FerrDb. Paired samples were compared using a paired t-test, and key regulatory factors, such as GPX4 and Tfrc, were analysed. Spearman’s rank correlation coefficients were calculated between ferroptosis-related and osteoclast marker genes (e.g. Acp5, Ctsk and Mmp9) and heatmaps illustrated co-expression networks.
Immunofluorescent staining
Femoral tissue from each group of rats was collected and fixed in 4% paraformaldehyde (PFA) and then transferred to ethylenediaminetetraacetic acid (EDTA) for decalcification. The femurs were embedded in paraffin and sliced into 5 μm sections using a microtome. After incubating in blocking solution for 1 h, the sections were treated overnight at 4°C with the following mouse or rabbit primary antibodies: anti-alkaline phosphatase (ALP) (1:500, DF6225, Affinity Biosciences, Cincinnati, OH, USA), anti-cathepsin K (1:200, 11239-1-AP, Proteintech Group, Inc., Rosemont, IL, USA), anti-FTH1 (1:200, 83428-1-RR, Proteintech Group, Inc.) and anti-GPX4 (1:200, 67763-1-Ig, Proteintech Group, Inc.). The following day, coralite488 (CL488)-conjugated goat anti-mouse immunoglobulin (Ig)G H&L (1:800, SA00013-1, Proteintech Group, Inc.) and fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG H&L (1:800, SA00003-2, Proteintech Group, Inc.) secondary antibodies were added for 1 h at room temperature in the dark, followed by a 4’,6-diamidino-2-phenylindole (DAPI) solution (BL105B, Biosharp, Hefei, China) counterstain for cell nuclei. Images of all sections were captured using an inverted fluorescence microscope (Axio Vert A1, Carl Zeiss Microscopy GmbH, Jena, Germany) and mean fluorescence intensity was analysed using ImageJ (National Institutes of Health (NIH), Bethesda, MD, USA).
Western blotting
Total protein was extracted from bone tissue using a lysis buffer containing protease inhibitors. The lysate was incubated on ice and the supernatant was collected. Total protein concentration was determined using a bicinchoninic (BCA) protein assay kit (P0011, Beyotime Biotechnology). Absorbance was measured at 562 nm with a multi-function microplate reader (EnVision, PerkinElmer, Waltham, MA, USA). Sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE) (5X) protein loading buffer was added to the collected protein sample, and followed by denaturation in a metal bath. In total, 20 μg of protein was loaded and separated by SDS-PAGE on a 12% gel. Proteins were transferred to polyvinylidene fluoride membranes and blocked. The membranes were then incubated overnight at 4°C with the following mouse primary antibodies: anti-SLC7A11 (1:1000, HA721868, HUABIO, Hangzhou, China), anti-FTH1 (1:1000, 83428-1-RR, Proteintech Group, Inc.), anti-GPX4 (1:2000, 67763-1-IG, Proteintech Group, Inc.) and the internal reference anti-β-actin (1:20000, 66009-1-IG, Proteintech Group, Inc.). The second day, the membranes were incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG H&L secondary antibodies (1:5000, SA00001-1, Proteintech Group, Inc.) for 1 h at room temperature. Enhanced chemiluminescence (ECL) substrate working solution (P0018S, Beyotime Biotechnology) was used for development and image acquisition. Quantitative analysis was performed using ImageJ (NIH) with β-actin as an internal reference.
Statistical analysis
All data are expressed as the mean ± standard deviation (SD). Bar graphs were generated using GraphPad Prism version 10 (GraphPad software, La Jolla, CA, USA). Groups were compared using one-way analysis of variance (ANOVA) with a post hoc Dunnett’s test for pairwise comparisons. p < 0.05 was considered statistically significant.
Results
EA ameliorated OVX-induced osteoporosis
Figure 2 shows the results of the micro-CT analysis to evaluate the impact of EA on bone microstructure. The OVX group showed a significant loss of femoral trabecular bone compared with the CONTROL and SHAM groups, confirming successful model establishment. The EA group exhibited increased femoral trabeculae relative to the OVX group (Figure 2(A)). As shown in Figure 2(B), (C) and (F), there were significant decreases in BV/TV, BMD and Tb.N in the OVX group compared with the CONTROL and SHAM groups (p < 0.05). Meanwhile, these parameters were markedly higher in the EA group (p < 0.05). In addition, Tb.Sp was significantly elevated in OVX versus CONTROL and SHAM groups (both p < 0.05), but EA treatment markedly reduced Tb.Sp (Figure 2(D), p < 0.05). The SMI of the OVX group indicated severe microstructural degeneration and was significantly higher than in the CONTROL and SHAM groups (p < 0.05). However, the SMI value in the EA group did not significantly differ (Figure 2(E), p > 0.05). Overall, these findings show that OVX induced OP in our rat model and that EA significantly alleviated several markers of the condition.
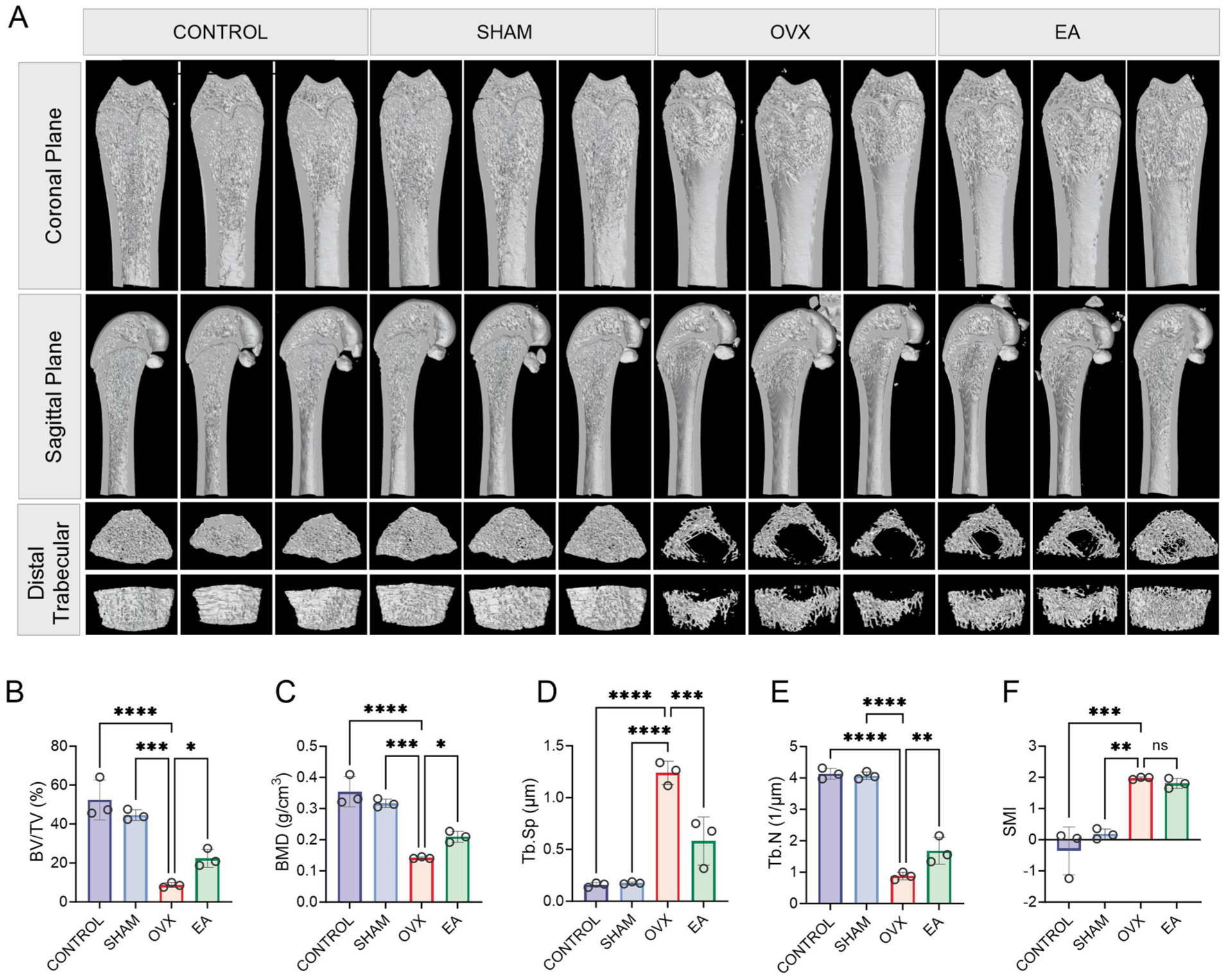
Effects of electroacupuncture (EA) on femoral microstructure in ovariectomy (OVX)-induced osteoporosis model rats. (A) 3D micro-computed tomography images of femurs from CONTROL, SHAM, OVX and EA groups, displaying various views. (B–F) Bone microstructure parameters: bone volume fraction (BV/TV), bone mineral density (BMD), trabecular separation (Tb.Sp), trabecular number (Tb.N) and structure model index (SMI). Data are presented as mean ± SD. n = 3 per group. Significant differences (p < 0.05) are marked with asterisks.
EA reversed OVX-induced bone metabolic imbalance by inhibiting osteoclast activity and enhancing local osteoblast activity
Serum β-CTx levels were significantly higher in the OVX group than in the CONTROL and SHAM groups (both p < 0.05). EA treatment lowered β-CTx levels compared with no treatment in OVX rats (Figure 3(A), p < 0.05), indicating that EA inhibited OVX-induced osteoclast activity.
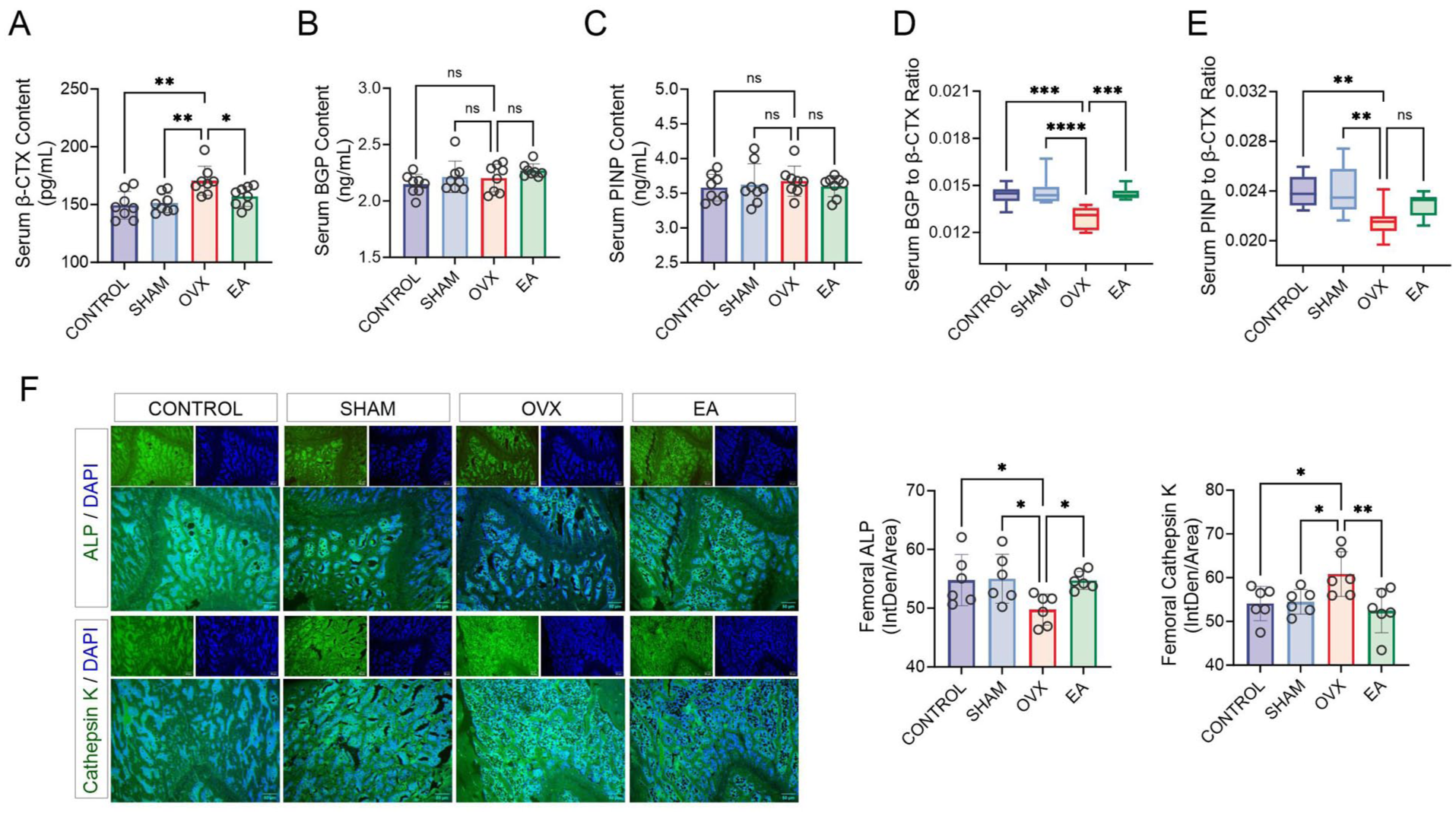
Effects of electroacupuncture (EA) treatment on serum bone markers and histomorphology. (A–E) Analysis of serum indicators: C-terminal telopeptide of type I collagen (β-CTx), bone γ-carboxyglutamic acid (Gla) protein (BGP), procollagen type I N-terminal propeptide (PINP), BGP/β-CTx ratio and PINP/β-CTx ratio. n = 8 per group. (F) Fluorescent analysis of alkaline phosphatase (ALP) and cathepsin K in femoral tissue. n = 6 per group.
There were no significant differences in BGP or PINP between the groups (Figure 3(B) and (C), p > 0.05). However, the BGP/β-CTx and PINP/β-CTx ratios were significantly lower in the OVX group, confirming increased bone resorption activity. EA treatment markedly increased the BGP/β-CTx ratio (Figure 3(D), p < 0.05), suggesting a relative restoration of the balance between resorption and formation. However, the PINP/β-CTx ratio did not significantly differ in the EA group (Figure 3(E), p > 0.05).
Immunofluorescent staining (Figure 3(F)) showed significantly lower ALP intensity in the OVX group compared with the CONTROL and SHAM groups (p < 0.05) and substantially higher intensity in the EA group (p < 0.05). Conversely, cathepsin K levels were significantly elevated in the OVX group and reduced in the EA group (p < 0.05). These results indicate that EA counteracts bone metabolic imbalance by inhibiting osteoclast activity and restoring osteoblast function.
OVX upregulated osteoclast gene expression and inhibited ferroptosis pathway-related genes
Figure 4 presents the differential gene expression analysis of the GSE68313 dataset. As depicted in Figure 4(A), the similar gene expression distributions in the OVX and SHAM groups allowed for further analysis. There were no significant differences in mRNA levels of key osteoblast-related genes between the groups (Figure 4(B), p > 0.05). However, as indicated in Figure 4(C), osteoclast-related genes had significantly higher expression in the OVX group compared with the SHAM group (p < 0.05), suggesting that increased osteoclast activity contributes to the bone metabolic imbalance induced by OVX.
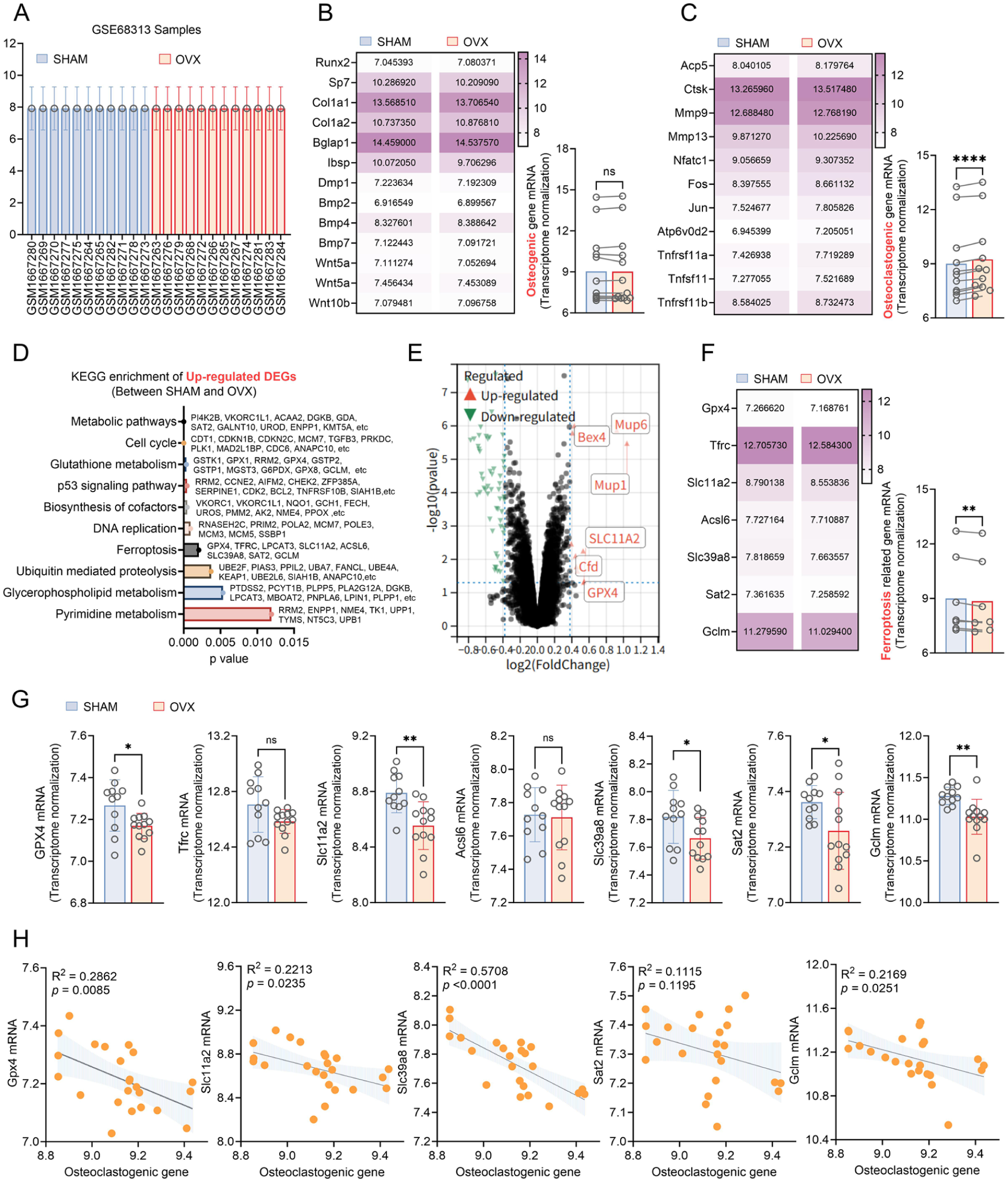
Differential gene expression in the GSE68313 dataset. (A) Normalisation between SHAM and OVX (ovariectomy) groups. (B) Key osteoblast gene expression. (C) Key osteoclast gene expression. (D) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of upregulated differentially expressed genes (DEGs). (E) Volcano plot of DEGs. Red dots indicate significantly upregulated genes, blue dots indicate significantly downregulated genes, and grey dots indicate genes that are not significantly different. Dashed lines mark the significance threshold. (F) and (G) Ferroptosis pathway gene expression analysis. (H) Correlation analysis between characteristic genes enriched in the ferroptosis pathway and osteoclast-related gene expression.
Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis (Figure 4(D)) revealed that DEGs were upregulated in the SHAM group and significantly enriched in the ferroptosis pathway (p < 0.05), indicating suppressed ferroptosis-related gene expression under the OVX condition. As shown in Figure 4(E), there was substantial upregulation of Mup6, Mup1, SLC11A2, Cfd and GPX4 in the SHAM group (p < 0.05). As illustrated in Figure 4(F), there were markedly lower levels of upregulated DEGs in the OVX group (p < 0.05), particularly for GPX4, SLC11A2, Slc39a8, Sat2 and Gclm, whereas Tfrc and Acsl6 showed no significant differences (Figure 4(G), p > 0.05). These findings suggest that OVX significantly downregulated key genes in the ferroptosis inhibitory pathway, particularly the core inhibitors GPX4, Sat2 and Gclm, indicating that cellular capacity to resist lipid peroxidation may have been impaired. Simultaneously, SLC11A2 and Slc39a8, which are involved in iron homeostasis, were also significantly downregulated, which may reflect a compensatory response to the iron metabolic imbalance induced by OVX.
As displayed in Figure 4(H), there was a notably negative correlation between the mRNA levels of GPX4, SLC11A2, Slc39a8 and Gclm with osteoclast-related gene expression (p < 0.05), whereas Sat2 showed no linear relationship. The present results demonstrate a negative correlation between osteoclast activity and the expression of ferroptosis-related genes. The correlation was strongest for Slc39a8 and second strongest for GPX4, indicating that increased bone resorption is associated with diminished ferroptosis inhibitory activity. Although Slc39a8 showed the strongest correlation, given the indispensable core role of GPX4 as the ultimate effector in clearing lipid peroxides and mediating ferroptosis, we ultimately selected GPX4 as the primary entry point for our subsequent research.
EA inhibited OVX-induced ferroptosis by modulating iron accumulation and the SLC7A11/GPX4 axis
The results of ferroptosis-related assays are summarised in Figure 5. The iron content of tibial tissue and serum of the OVX group was significantly higher compared with the CONTROL and SHAM groups and was decreased in the EA group (Figure 5(A) and (B), all p < 0.05). Immunofluorescent staining (Figure 5(C)) showed that GPX4 levels were significantly reduced in the OVX group but increased in the EA group. FTH1 showed a similar trend, with significant decreases in the OVX group and increases following EA treatment; these changes were further confirmed by quantitative analysis of fluorescence intensity (Figure 6).
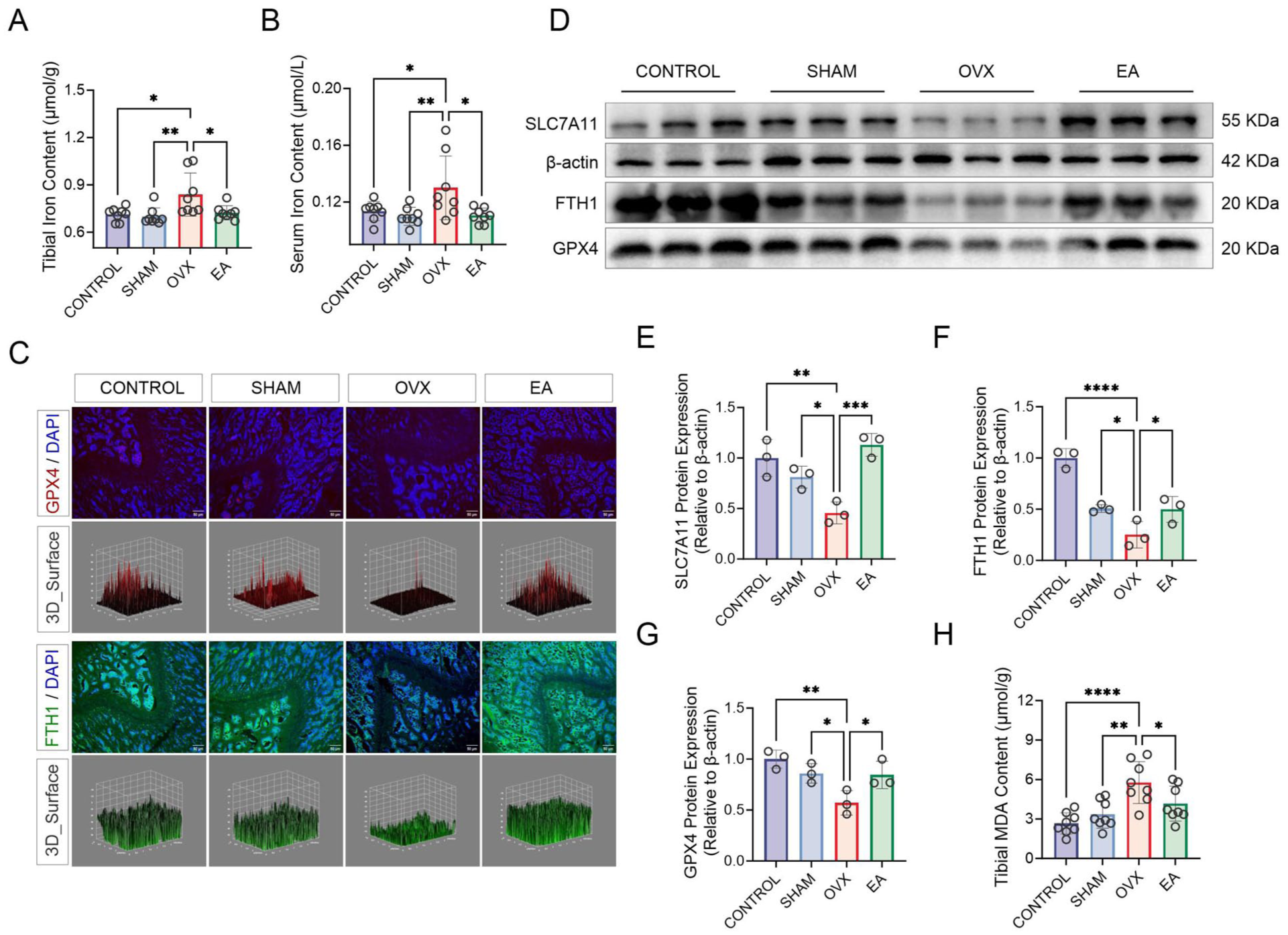
Electroacupuncture (EA) improves ovariectomy (OVX)-induced osteoporosis by regulating proteins related to ferroptosis. (A) and (B) Measurement of iron levels in tibial tissue and serum (n = 8 per group). (C) Immunofluorescence images of glutathione peroxidase 4 (GPX4, red) and ferritin heavy chain 1 (FTH1, green) in femoral tissue. 4’,6-diamidino-2-phenylindole (DAPI)-stained nuclei (blue) are also shown. Corresponding 3D surface plots are included. n = 6 per group. (D)–(G) Western blot analysis of solute carrier family 7 member 11 (SLC7A11), FTH1 and GPX4 protein expression. n = 3 per group. (H) Measurement of malondialdehyde (MDA) levels. n = 8 per group.
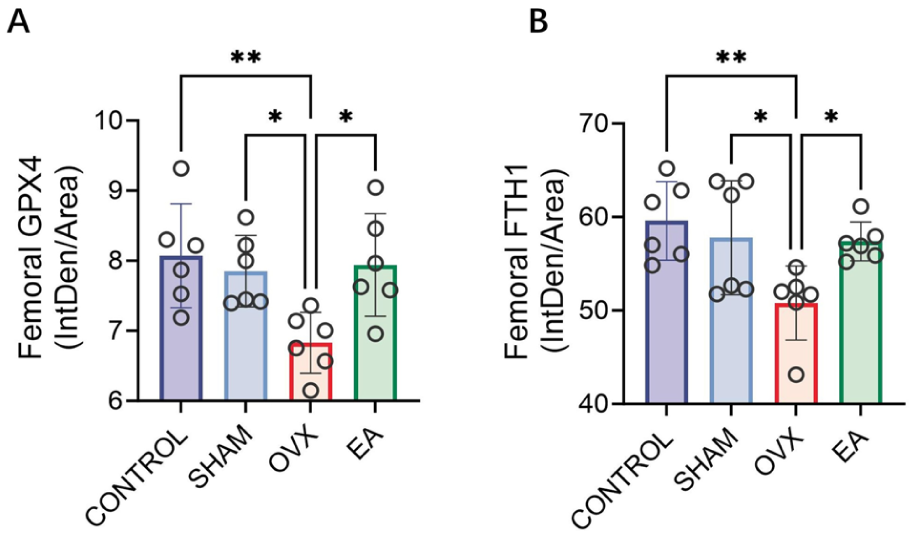
Quantitative analysis of glutathione peroxidase 4 (GPX4) and ferritin heavy chain 1 (FTH1) immunofluorescence staining. Statistical analysis of the mean fluorescence intensity (integrated density/area) for GPX4 (A) and FTH1 (B) in femoral tissues across the four groups. n = 6 per group.
Western blot analysis (Figure 5(d)) showed significantly weaker expression of SLC7A11, FTH1 and GPX4 in the OVX group compared with the CONTROL and SHAM groups, while the EA group displayed markedly increased levels. The data presented in Figure 5(e)–(g) confirm these results, with significant reductions in the expression of these proteins in the OVX group and increases in the EA group (all p < 0.05). Moreover, MDA levels in tibial tissue were markedly elevated in the OVX group and significantly decreased by EA treatment (Figure 5(h), p < 0.05).
These results suggest that OVX regulates iron accumulation and inhibition of the SLC7A11/GPX4 axis, leading to ferroptosis. EA intervention effectively reversed these effects, apparently protecting against OVX-induced OP.
Discussion
As a vital component of Traditional Chinese Medicine (TCM), acupuncture can regulate organ function and restore homeostatic balance by stimulating somatic tissues through needle insertion and manipulation. Due to its high safety profile, acupuncture shows great promise for the prevention and treatment of OP.24–26 However, the exact mechanisms behind its therapeutic effects in OP are still not fully understood.
In the present study, we selected traditional acupuncture point locations for EA based on TCM theory.27,28 Consistently, recent evidence also indicates that BL23, CV4, ST36 and SP6 are among the most frequently used traditional acupuncture point locations in the treatment of OP, as summarised in a comprehensive review by Tian et al. 29 We established an OVX-induced OP model using female SD rats, which displayed decreased BMD and impaired trabecular architecture. After EA treatment, we observed significant improvements in BMD and morphological parameters of trabecular bone. This finding aligns with previous results reported by Shu et al., 17 who found that EA similarly improved BMD and trabecular bone morphology in OVX rats, confirming the effectiveness of EA in alleviating OVX-induced OP.
In the OVX group, the significant rise in β-CTx and the upregulation of cathepsin K indicate a high-turnover osteoporotic state, mainly driven by increased bone resorption. BGP and PINP levels showed no significant differences, whereas immunofluorescence staining revealed a substantial decrease in ALP expression, suggesting suppressed osteoblast activity. Predominant resorption appeared to result in a decline in serum bone turnover balance indices (BGP/β-CTx and PINP/β-CTx ratios), confirming a negative balance and net bone loss.
EA treatment significantly suppressed β-CTx levels and cathepsin K expression, indicating reduced osteoclast activity and bone resorption. This finding is consistent with a prior study by Zhou et al., 30 which also reported a significant reduction in β-CTx and cathepsin K following EA treatment. The BGP/β-CTx ratio was notably increased in the EA group, suggesting improved bone metabolic balance. Immunofluorescent staining showed that EA also promoted ALP expression in bone tissue, enhancing osteoblast function. This observation supports the findings of a study by Fan et al., 15 in which EA increased ALP levels, reflecting that EA promotes osteogenic activity. Thus, EA demonstrates a dual role in inhibiting bone resorption while supporting local osteogenic activity.
Based on the genomic findings, we utilised the ferroptosis-inducing OVX model to confirm the role of ferroptosis in this process. The OVX group showed a significant increase in iron levels and a notable decrease in the iron storage protein FTH1, confirming iron accumulation. In addition, SLC7A11 and GPX4 expression decreased, while MDA levels increased, indicating that inactivation of the SLC7A11/GPX4 axis led to ferroptosis. However, the EA intervention significantly reduced iron levels and increased FTH1 expression, reversing the accumulation. Moreover, the EA group also exhibited higher SLC7A11 and GPX4 levels and lower MDA, demonstrating that EA mitigated OVX-induced bone ferroptosis by restoring the SLC7A11/GPX4 axis.
In conclusion, this study found that EA significantly enhanced OVX-induced OP in rats by decreasing bone resorption through inhibition of β-CTx and cathepsin K while promoting local osteogenic activity. EA also reduced iron accumulation and restored the SLC7A11/GPX4 axis, effectively decreasing ferroptosis and improving OP. To our knowledge, our study is the first to connect the mechanism of EA in OP treatment to the ferroptosis pathway, potentially offering new targets for prevention and treatment. However, our findings remain correlative rather than causal, given the absence of any direct pharmacological intervention (e.g. use of ferroptosis inhibitors or activators), which is a limitation. To establish the necessity of the ferroptosis pathway in the mechanism of EA treatment of OP, in the future, we plan to conduct validation experiments, such as knocking out key genes in the SLC7A11/GPX4 axis, and explore the precise upstream signalling pathways that potentially govern GPX4 activation by EA.
Footnotes
Acknowledgements
The authors would like to thank Tong-kang Guan and Bo-Yi Zhu from the Jiangsu Province Geriatric Hospital and Chu-qiong Zhang from the Huai’an Hospital of Traditional Chinese Medicine for providing assistance during this experiment.
Contributors
G.O. and C.C.Z. conceived and designed the experiments. C.C.Z. analysed all the data and was responsible for writing the manuscript. All authors read and approved the final version of the manuscript accepted for publication.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the Jiangsu Provincial Cadre Healthcare Bureau Project (grant no. BJ23018) and the Nanjing Medical University Science and Technology Development General Project (grant no. NMUB20240121).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.