
Abstract
Background
Neurodevelopmental disorders like autism are on the rise, with the role of genetics and epigenetics being increasingly cited as reasons or contributors. The environmental factors were found to be important as epigenetic modifiers, and pregnant mothers are incrementally exposed to many of these environmental factors, like aluminium (Al), known to damage neurons, even before delivery. However, its presence and persistence inside the central nervous system (CNS), leading to changes in the expression of behaviours, need further research. Hence, this animal study was conceived.
Purpose
To evaluate the impact of early dietary exposure to Al in the brains of foetuses of pregnant rats and its subsequent effects on the neurobehaviour in their pups.
Methods
Pregnant Wistar rats were divided into four groups (Gr1 receiving injection tetanus toxoid; Gr2 receiving tetanus toxoid + soy; Gr3 receiving tetanus toxoid + valproic acid and Gr4 receiving tetanus toxoid + soy + valproic acid). A few pups were sacrificed, and their brain Al and α-synuclein levels were assessed. Others were allowed to grow, and their behaviour, α-synuclein and Al levels were analysed.
Results
α-synuclein was low in groups 3 and 4, with group 4 pups with abnormal socialisation behaviours exhibiting the lowest levels, suggesting ongoing neuronal injury. Brain Al in pups sacrificed immediately after birth suggests prenatal Al exposure, with the highest values in group 4, though all had elevated Al levels. Gr4 had impaired socialisation. Higher sociability indices were noted in groups 1 and 2 (0.36 and 0.3) compared to groups 3 and 4 (0.09 and 0.04). Groups 1 and 2 pups demonstrated intact social memory and novelty seeking. There was also a negative correlation between brain Al and α-synuclein levels and the socialisation index.
Conclusion
Environmental factors like Al once enter the CNS remain in the brain even when invisible in blood and can induce changes in the growing brain, behaviour and socialisation.
Keywords
Introduction
Autism spectrum disorder (ASD) is a chronic neurodevelopmental disorder with considerable genetic basis, the susceptibility of which is enhanced by epigenetic alterations from the environment. Recent research points out that prenatal exposure to valproic acid (VPA) increases the likelihood of autism, especially those rodent experiments that exhibited typical features of ASD. 1 This robust autism model has good face construct and predictive validity and provides a more accurate representation of ‘idiopathic autism’ of environmental/epigenetic origin compared to transgenic models based on single gene mutations. 2 Prenatal VPA exposure alters the expression of several genes in the cortices of male pups. 3
α-synuclein has been found to be reduced in many neurodegenerative disorders, including ASD. It is a small protein from neuronal cells that helps in synaptic function and neurotransmission. Though a soluble protein, its aggregation happens, reducing its levels, during neuronal insults. The neuronal damage is evident from the alterations of α-synuclein levels in ASD. Environmental epigenetic factors like aluminium (Al) are toxic to humans and are present not only in drinking water (usual levels below 0.2 mg/L) but also in most natural foodstuffs, such as potatoes, spinach and tea, at significant concentrations (typically below 5 mg/kg) and on cooking utensils and foils. A recent report by the American Academy of Pediatrics emphasised the critical need for research elucidating the health hazards associated with elevated Al in infant formula and highlighted an urgent requirement for investigations into Al levels in preterm infants and those with compromised renal function. 4 Al contamination with concentrations ranging from 100 to 430 µg/L was found in 30 infant formulae, both ready-to-drink milk and milk powders, with two soy-based milk products having 656 and 756 µg/L. The intake of Al from infant formulae was high (non-soy-based infant formulae: 100–300 µg/day; soy-based milk: up to 700 µg/day). 5 The soy-based and soy milk-based formulae had a mean Al concentration of 5 mg/kg (0.6 mg/L). 6 A recent study on infant formulae revealed that soy formulae intended for infants with poor weight gain contained the highest Al content (153.5–1,956.3 µg/L), followed by specialised formulas for preterm and intrauterine growth restriction (IUGR) infants (49.9–249.4 µg/L). 7
The use of Al as an adjuvant to enhance vaccine immunogenicity has been a subject of debate. 8 Analyses of Al exposure following the introduction of six additional vaccines revealed a 25% increase in Al exposure from 2000 to 2005. With the 2011 Centre for Disease Control and Prevention (CDC, USA) recommendation that pregnant women can receive vaccines containing Al, the possibility of increased prenatal Al exposure has increased. 9 Despite compelling evidence to the contrary, numerous reviews and comprehensive prospective studies have dismissed the association between measles, mumps and rubella (MMR) vaccines and the aetiology of autism, though most were epidemiological or retrospective.10–13 The potential for Al to remain in the central nervous system (CNS) and act as the focal point for sustained low-grade neuronal inflammation, even when cleared quickly from the circulatory system, remains unexplored.
Therefore, it is essential to investigate the combined, long-term impact of Al deposition, along with other environmental factors that influence the nervous system during organogenesis and subsequent development. Such investigations are essential to elucidate the role of Al as an epigenetic gene-modifying agent in ASD. This may explain the lack of identifiable mutations in the majority of patients, with less than 30% of ASD cases having mutations. Consequently, the influence of Al obtained through diet and alternative sources during neurogenesis in conjunction with prenatal valproate exposure, simulating a non-genetic clinical prototype of ASD, was examined.
This study aimed to evaluate the likelihood and potential impact of early dietary exposure to Al in the brains of foetuses of pregnant rats treated with VPA and its subsequent effects on the neurobehaviour of their pups.
The objectives of the study were:
To determine whether Al exposure during gestation results in the persistent presence of metals in the postnatal brain.
To determine whether Al exposure in gravid rats induces alterations in the foetal brain, thereby establishing a persistent foundation for neuroinflammation that affects neurodevelopment throughout the lifespan.
To evaluate whether Al exposure in conjunction with clinical doses of VPA results in impaired social communication behaviours in animal models.
To investigate whether combined exposure leads to alterations in α-synuclein expression in pups subjected to prenatal VPA and Al exposure.
Materials and Methods
Animals and Ethical Approval
Pregnant Wistar rats at gestational day 6, obtained from the Biogen Animal Facility in Bangalore, were maintained under a standard light-dark cycle at an ambient temperature of 25°C ± 2°C and humidity of 50%–60%, with ad libitum access to chow pellets and water. After 6 days of acclimatisation to their home cages, this study was conducted following the guidelines stipulated by the Committee of Control and Supervision of Experiments on Animals (CCSEA, India) and was approved by the Institutional Animal Ethics Committee (SU/CLAIR/RD/002/2022). The Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines were adhered to throughout the animal research and documentation process.
Procedure
Only pregnant Wistar rats with an average weight of 250 ± 10 g, confirmed by the presence of a vaginal plug and detection of semen through microscopic examination of a vaginal specimen, were included in the study. The pregnant rats were divided into four groups of three rats each: group 1 received an injection of Tetanus Toxoid (TT; 0.5 mL) in addition to normal feeds (control); group 2 received an injection of TT and soya feed ad libitum; group 3 received an injection of TT, normal feeds and VPA (370 mg/kg) and group 4 received an injection of TT, soya feeds and VPA (370 mg/kg) administered orally as a solution. Sodium valproate (Sigma–Aldrich, USA) was dissolved in 0.9% saline to achieve a concentration of 100 mg/mL at pH 7.3. The dosage was adjusted according to the body weight of pregnant rats on the day of enrolment. Between gestational days 10 and 13 (E10–E13), mother rats in the groups consuming 3, and 4 received VPA at a dose equivalent to 50 mg/kg/day for humans [50 × 7.4 mg/kg/day (conversion factor for rats) = 370 mg/kg for rats], from the day pregnancy was confirmed. This was intended to better simulate human consumption than the previously validated autism model experiments in which the rats were given 600 mg/kg/day VPA between gestational days 10 and 13.2, 13 Accordingly, 92.5 mg of VPA was administered to each 250 g pregnant rat (370 mg × 250 g/1,000 g). Rats in the TTS and TTVS groups were provided with 1 mL of soymilk daily, in addition to their standard pellet diet. This volume (1 mL) was meticulously selected to give the rats the intended nutritional benefits while avoiding adverse effects.
The groups were isolated throughout pregnancy and were monitored for well-being, including checking metabolic parameters. Immediately after birth, three pups from each group were randomly selected and euthanised for brain Al analysis. The remaining pups were allowed to remain with their mothers. They were subjected to social behaviour experiments at 2.5 weeks of life (equivalent to 1.87 human years/684 human days), after which they were sacrificed and brain tissue analysed for Al content. Blood samples were also analysed for α-synuclein levels.
Behavioural Testing
Sociability Test
The sociability test with an unfamiliar and familiar rat was developed and tested with multiple changes in 2.5-week-old Wistar rats. The test was conducted in a cage (75 × 75 × 50 cm) with three compartments (25 × 25 cm) interconnected through a square opening (10 × 10 cm) at the centre bottom of the middle partitions for the animals to pass through. Behavioural testing was performed between 9:00
The three-chamber paradigm test, known as Crawley’s sociability and preference for social novelty protocol, was used for the testing (procedure adapted from Kaidanovich-Beilin et al., 2011). 14 The experiment consisted of evaluating the time spent by the rats to freely choose to spend time in any of the three compartments/boxes during two experimental sessions. In session 1, the time spent by the rats with a novel conspecific was measured, and in session 2, the preference for a novel conspecific was measured. Session 1 tested the social affiliation of rats. The experimental rats could choose to spend more time around the wired restraint cup containing the stranger1 rat or move to the compartment with the ‘inanimate mouse model’ to avoid contact (detailed procedure in Annexure 1).
Monitoring of Sessions
Session 1: Recorded the duration and number of contacts between the experimental rat and the restraint-wired cup with either the ‘stranger1’ rat or an inanimate rat model. Rats could choose to spend time around the cup with stranger1 or move to the compartment with the toy rat. Active contact included direct contact or stretching within 3–5 cm around the cup. Noted behaviours such as walking, self-grooming, freezing (lack of movement >5 s), jumping, repetitive actions and the duration and number of times they fully entered each compartment.
Session 2: Testing of social novelty/preference was done by placing a second normal rat (‘stranger2’) inside the wire containment cup in the previously empty (opposite) chamber. The aforementioned parameters were monitored, and the behaviours and interactions of experimental rats with ‘stranger1’ and ‘stranger2’ were differentiated.
The ratio of entries into the animal compartment, as well as the ratio of entries into the toy rat compartment to the total number of entries into both compartments, was calculated. The total duration of the recording was also noted. Two distinct aspects of social behaviour were determined by this experiment: social affiliation/motivation and social memory/novelty. Sociability was defined as ‘the propensity to spend time with another rat, as compared to time spent alone in an identical but empty chamber’. ‘Preference for social novelty was defined as the propensity to spend more time with an unfamiliar rat than with a familiar rat’. The coding was done to easily recognise and document different animal behaviours as shown in Table 1 (Annexure 2). The participant recording the behavioural parameters was blinded to the group selection and treatment conditions, and the information was decoded later.
Animal Surgery and Sample Preparation
After measuring the sociability of the different groups of pups, at the end of 2 weeks, they were sacrificed under fluothane anaesthesia. The brain was dissected by a sagittal incision on the head, samples isolated and preserved in plastic containers to prevent contamination by particulate matter and metallic substances. Brain samples were homogenised to ensure uniformity by sectioning samples into smaller samples of 0.3–0.5 g, desiccated and digested in concentrated nitric acid and filtered and diluted to a suitable concentration for analysis.
Al Measurement
The diluted sample was aspirated into an inductively coupled plasma optical emission spectroscopy (ICP-OES) and nebulised into a fine mist using an Ultrasonic Nebulizer U-500AT+. The mist was introduced into a plasma torch, wherein the elevated temperature excited the Al atoms, emitting light at the characteristic wavelengths of 308.22 and 396.152 nm (which yields the highest intensity), detected and quantified using a spectrometer. The intensity of the emitted light was compared with that of the calibration curve to determine the Al concentration.
α-synuclein Measurement
Blood samples from the central vessels were collected in ethylenediaminetetraacetic acid (EDTA) tubes and stored at −80°C until the day of sandwich enzyme-linked immunosorbent assay (ELISA) analysis for α-synuclein at a wavelength of 450/620 nm using an ELISA machine (Bio-Rad Evolis; Rat SNCa ELISA Kit ELK Biotechnology Co. Ltd., Denver, CO 80202 USA, Cat No.: ELK7204).
Statistical Analysis
Statistical analysis was conducted using SPSS version 21 (IBM Corp., Armonk, NY, USA) for data analysis. The mean of the α-synuclein values of rats in each group was calculated in pg/mL, and the significance of the difference in levels between the groups was analysed. The mean of triplicate Al concentrations with the lowest RDS was taken to increase the reliability and validity of the results.
Analysis of Sociability Index (SI)
The total number and duration of contacts between experimental rats and inanimate restraint cups or cups with stranger1 (session I) and between the experimental rats and the cups with stranger1 versus stranger2 (session II) were calculated. The mean duration of contact was calculated for each experimental group (TT, TTV, TTVS and TTS). The total number and duration of alternate behaviours (freezing, self-grooming and walking) were also calculated. The total time spent by the experimental rats in each compartment was also calculated. The significance of the difference (p < .05) in the above parameters between the groups was compared. Session 1 analysis by Whisker plots provided social affiliation and sociability plots depicting mean duration (±SEM) in the chamber with the ‘stranger1’ compared to the opposite chamber (‘inanimate rat’).
Session 2 plots provide the social memory and novelty indices with the mean duration of time (±SEM) in the chamber with the unfamiliar rat from the previous sociability phase (‘stranger1’) and in the opposite chamber with a new unfamiliar rat (‘stranger2’). Significance was calculated (p < .05). Outlier analyses were conducted before reporting. Values that were ±2 standard deviation (SD) from the mean were adjusted to be one unit more extreme than the closest data point in the distribution. The effect sizes for the analysis of variance (ANOVA) were reported as partial eta-squared (η2 p ). Independent samples t-tests were two-tailed, and statistical significance was set at p < .05. After ensuring normality with the Shapiro–Wilk test and homogeneity of variance with the Levene test, repeated ANOVA for assessing the significance of novelty effects of conspecific, groups or interaction was performed, and Tukey’s Honest Significant Difference (HSD) post-hoc test was used to identify specific differences.
Results
Twelve rats gave birth to 66 pups (Gr1–Gr18, Gr2–Gr21, Gr3–Gr15 and Gr4–Gr12 pups). Nine pups were randomly selected from the four groups and were sacrificed at 2.5 weeks (36 brain samples generated: 9 each × 4 groups).
Changes in Biochemical Parameters
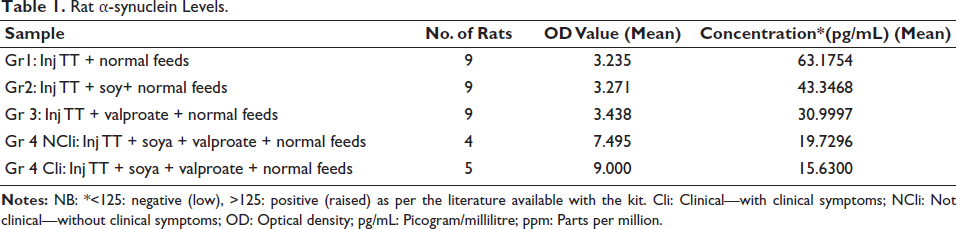
α-synuclein levels in the blood of pups of different groups of mother rats are shown in Table 1.
Rat α-synuclein Levels.
Analysis of Al levels in the brain revealed the following (Table 2):
Al Levels in Rats’ Brains Measured at 396.152 nm Wavelength.
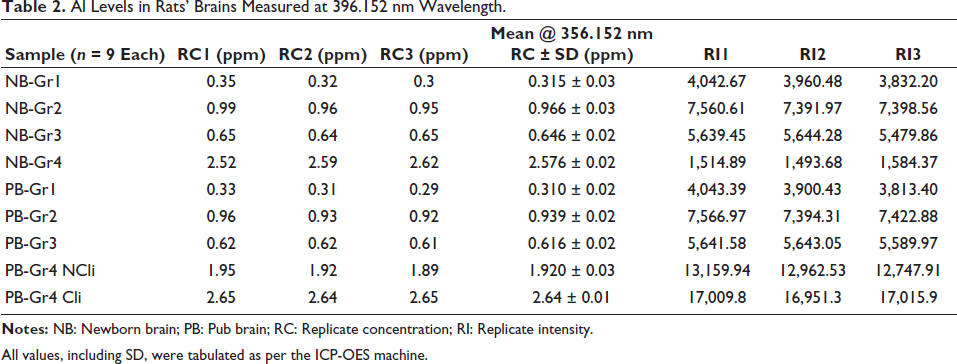
All values, including SD, were tabulated as per the ICP-OES machine.
Session 1: Social Interaction
Rats at 2.5 weeks of age were assessed for their social interactions. Interaction index is the mean time spent by the mouse with stranger1 or rat toy. Interaction time is the total time spent interacting with stranger1 or the rat toy. SI = (time with stranger1 rat − time with toy rat)/total interaction time = for Gr1–Gr4, the values were 0.36, 0.3, 0.09 and 0.04, respectively. Repeated-measures ANOVA to assess the time spent in different chambers showed that there were main effects of conspecific (stranger1 vs inanimate rat toy), main effect of groups (groups 1–4) and an interaction as in Tables 3 and 4.
Analysis of Social Interaction–Total Duration (Stranger1 vs Toy Rat).

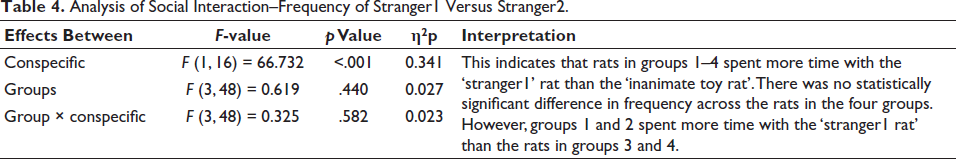
Analysis of Social Interaction–Frequency of Stranger1 Versus Stranger2.
Tukey’s HSD test (post-hoc) revealed that groups 1 and 2 had significantly higher responses with stranger1 compared to toy rat (p < .05) than groups 3 and 4.
When the mean length of time (±SEM) in the chamber with the stranger (‘stranger side’) compared to the opposite chamber (‘inanimate rat toy’) was assessed, Gr3 and Gr4 animals failed to demonstrate a preference for social proximity by spending the same time in both chambers. ***p ≤ .0001 between Gr1 and Gr2 when stranger1 and toy rat response were compared with Gr3 and Gr4, as shown in Figure 1.
Sociability Testing: Session 1.
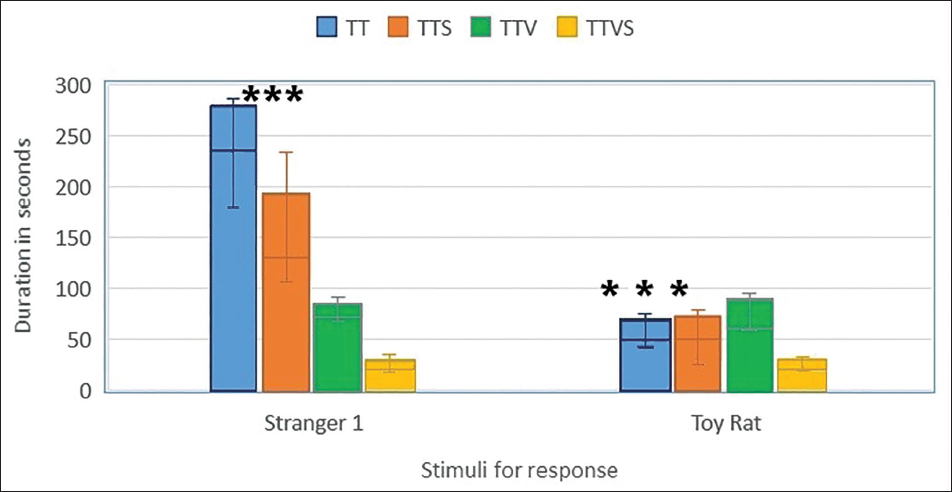
Post-hoc comparisons using Tukey’s HSD test revealed that groups 1 and 2 had significantly higher responses with stranger1 compared to the toy rat (p < .05) than groups 3 and 4. These results indicate that the presence of a conspecific significantly impacts the time spent in different chambers, while the group variables and their interaction did not.
Session 2: Social Novelty (Stranger1 Versus Stranger2)
The two conditions (strangers1 and 2) had nine observations per level of measure (i.e., nine rat behaviours per TT, TTV, TTS and TTVS). Because there are two conditions and four measures, we obtain df as 48 (includes correction factors—software correction applied for errors). Repeated-measures ANOVA to assess the time spent in and the number of times the rat visited different chambers showed that there were main effects of conspecific (stranger1 vs inanimate rat toy), the main effect of groups (groups 1–4) and an interaction (Table 5).
Analysis of Social Novelty.
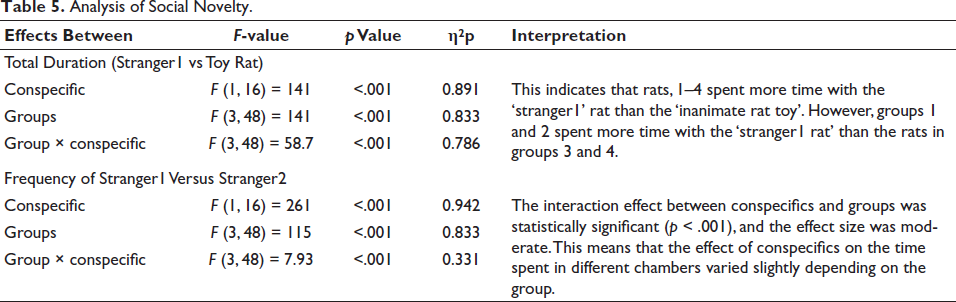
Tukey’s HSD post-hoc comparisons revealed that groups 1 and 2 had significantly higher responses with stranger1 compared to the toy rat (p < .05) than groups 3 and 4.
The mean duration of time (±SEM) in the chamber with an unfamiliar rat from the sociability phase (‘stranger1’) and in the opposite chamber with a new unfamiliar rat (‘stranger2’) are shown in Figure 2.
Social Memory and Novelty: Session 2.
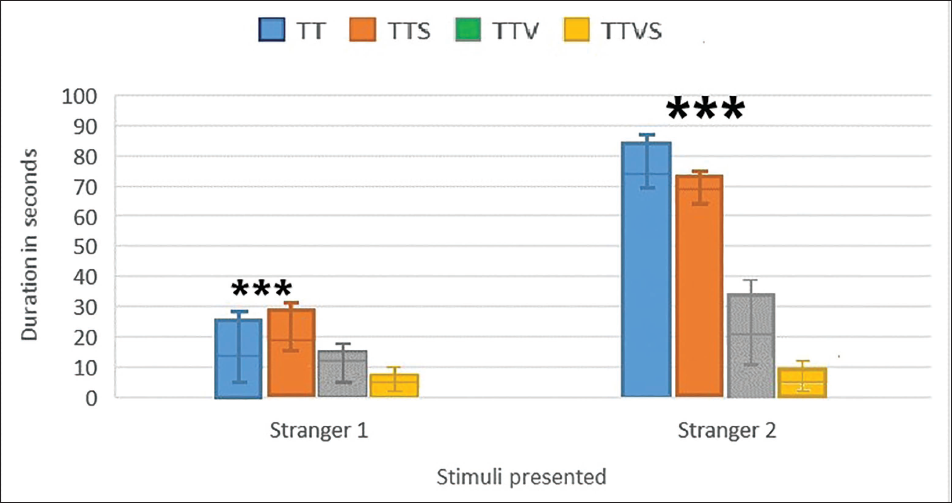
Unlike Gr1 and Gr2, Gr3 and Gr4 animals failed to demonstrate a preference for social novelty by spending the same amount of time in both chambers. ***p ≤ .0001 between Gr1 and Gr2 when strangers1 and 2 responses are compared. Novelty index (NI), (time exploring novel conspecific-time with familiar conspecific/total time), for groups 1–4 were 62.7, 45, 37 and 28.9, respectively.
Correlation
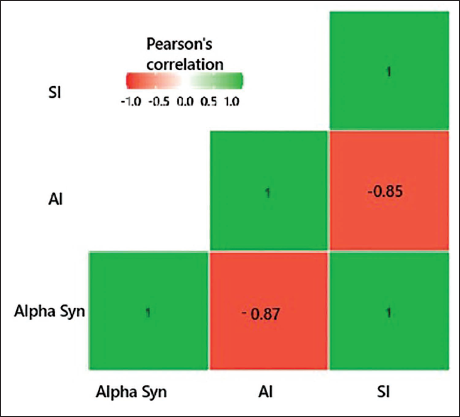
Pearson’s correlation has shown a positive relationship between the α-synuclein levels and SI (r = 0.997; p value = .003); a negative correlation between α-synuclein and AI (r = −0.866; p = .134) and a negative but insignificant correlation between Al and SI (r = −0.849; p = .151), as shown in the heat map (Figure 3).
Correlation Heat Map of Aluminium, Alpha-synuclein, and Sociability Index.
Discussion
This experimental study investigates the presence of Al in brain tissue after ingestion. Al is abundant in food, often exceeding safe levels, and RDA is not established due to its minimal biological role. Processed food, food additives, tea, cocoa powder and vaccine adjuvants also contain Al. Studies report that vaccines, especially MMR vaccines, are safe, with adjuvant Al cleared from the bloodstream within 24–48 h.9, 10 However, a recent study cautioned for Al exposure from catch-up vaccinations in infants who missed vaccinations following post-COVID isolation. 15 VPA, when used inadvertently during pregnancy, is associated with autism-like behaviour in children. Thus, a combined exposure of VPA and Al from the diet may lead to increasing neuronal damage during pregnancy. This study examines the combined effect of exposure to Al through dietary soy formula and VPA during pregnancy using a rodent model.
Pregnant rats were monitored throughout pregnancy for well-being and normal metabolic parameters. The blood α-synuclein level was analysed as previous studies have identified a correlation between α-synuclein and autism, as well as neurodegenerative disorders.16–18 Groups 3 and 4 had lower levels of α-synuclein, with group 4 pups with abnormal socialisation behaviours, exhibiting the lowest levels; those in group 4 who displayed typical social behaviour followed closely. However, none of the subjects exhibited values above the cut-off value of 125 pg/mL, indicating that the values were affected even in the TT-alone groups. This finding is significant, as it suggests ongoing neuronal injury. Since pups were randomly picked up and euthanised, brain Al in them suggests prenatal Al exposure resulting in Al passing through the blood–brain barrier in utero itself. The presence of Al could not be attributed to maternal rat milk or diet, as the pups were euthanised before feeding commenced. To the best of our knowledge, only a few studies have demonstrated the presence of Al in the brain following prenatal exposure.
The brain Al levels of remaining pups when analysed after 2.5 weeks of life showed the highest values in group 4, with significant differences within the group (symptomatic having higher values than asymptomatic pups). All groups exhibited elevated levels of Al, suggesting that Al from food, TT and soya could have contributed to higher levels. The pub brains with prenatal exposure to TT + soy and TT + VPA + soy had higher Al than the normally fed. The Al content of the soya feed was 542 µg/L of milk. Considering the low bioavailability of Al (0.21%), the absorbed amount was approximately 1.14 µg/L. Numerous studies have reported a high Al content in soya milk. 5 The TT vaccine typically contains Al phosphate as an adjuvant, and a single dose (0.5 mL) of the vaccine usually contains approximately 1.25 mg or less of Al. 4 In a study, it was found that deceased autistic adults exhibited higher Al levels in their brains, with concentrations ranging from 0.01 to 22.11 µg/g dry wt, with all five donor brains having Al content considered pathologically significant (≥3.00 µg/g dry wt) when compared with the non-exposed, that is, 0.19 µg/g (0.02–0.40). 19
This study investigated the effects of Al and VPA exposure on social behaviours and found that higher levels of Al, VPA and injection TT-exposed pups had altered social behaviours, including impaired socialisation (p < .0001) with higher SIs in groups 1 and 2 (0.36 and 0.3) compared to groups 3 and 4 (0.09 and 0.04). Pups preferred the stranger1 rat over the toy rat. Group 4 pups showed severe deficiencies in social memory and novelty-seeking behaviours. When pups were not exposed to Al and VPA (Gr1 and Gr2) preferred a new stranger (stranger2), demonstrating intact social memory and novelty seeking, pups exposed to Al and VPA (Gr3 and Gr4) preferred both strangers (strangers1 and 2) equally.
Al- and VPA-exposed rats (groups 3 and 4) with significantly reduced SI spent more time on the toy rat side and less on the familiar side, which is consistent with autism-like behaviours observed in other animal studies utilising VPA rat models of autism. 2 This could be due to oxidative stress and neuronal apoptosis, synaptic dysfunction and neuroinflammation during the brain development, which is maximum in group 4 due to double insult of VPA and Al. The safety of vaccines was based on the presumed difficulty of Al penetrating the blood–brain barrier and its rapid clearance by renal mechanisms, resulting in almost undetectable blood levels.10, 11 However, the tissue Al levels remained unmeasured in these studies; a critical oversight, as tissue concentration could provide more accurate insights into Al’s potential neurotoxic effects. The present study proves that Al continues to remain in the brain tissue, and hence its absence in the blood does not conclusively prove its absence in neural tissues. Vaccines introduce ~4.4 mg of Al in the first 6 months of life, but some studies report higher levels from breast milk and food. 20 Bioavailability and transport of Al across the blood–brain barrier are enhanced when it is injected than when consumed, a factor that is overlooked in epidemiological studies assessing biosafety. 21
Brain Al was negatively correlating α-synuclein and the socialisation index. α-synuclein inversely correlated with autism severity in numerous studies.17–19 Group 4 rats with high brain Al and VPA exposure displayed stereotypical behaviours and decreased socialisation (Supplementary material: Video 1). This experiment provides evidence that, contrary to the VPA-induced autism model at a prenatal exposure of 600 mg/kg, autism can also be induced at therapeutic VPA doses of 50–60 mg/kg/day, and Al enhances the neuronal damage as evidenced by the decreasing α-synuclein. Further, the presence of increased Al levels and decreased α-synuclein levels in the just-born pups before feeding was initiated shows neuronal damage in utero itself, and the same was seen in pups analysed after 2 weeks of age, correlating with the social behaviours also.
Conclusion
In conclusion, there is enough evidence that Al transferred from the maternal blood to the foetus persists in the foetal brain and potentially serves as a source of inflammation or alters neurodevelopment by interacting with other epigenetic factors and neurotoxins such as VPA. Contrary to the prevailing scientific belief that Al is quickly eliminated through renal mechanisms, the present study demonstrated that once Al enters the CNS prenatally, it can remain there for years after birth. Continued neuronal injury can ensue from dietary intake through food items with high levels of Al. This investigation provides a solid foundation for understanding the safety of exposure to small incremental doses of metals like Al in children. The study is limited by the absence of detailed evidence regarding the specific pathways and mechanisms through which neuronal damage occurs due to Al and VPA exposure. However, these aspects have been elucidated in detail in several previous studies. Further research is necessary in human subjects to elucidate the potential roles of epigenetic and neurotoxic factors, such as Al, in producing protracted low-grade in utero neuronal changes extending into the initial years of life.
Footnotes
Abbreviations
Al: Aluminium; Cli: Clinical—with clinical symptoms; Gr: Group; ICP-OES: Inductively coupled optical emission spectroscopy; NB-Gr: Newborn-group; NCli: Not clinical—without clinical symptoms; PB-Gr: Pub group; Pg/mL: Pictogram/millilitres; SEM: Standard error of the mean; TT: Tetanus toxoid; µg/g: Microgram/gram; VPA: Valproic acid; TTS: Inj TT + soy (TTS); TTV: Inj TT + valproate; TTVS: Inj TT + soya + valproate; RDA: Recommended dietary allowances.
Acknowledgements
The authors sincerely acknowledge the services of Mr Silambarasan, research assistant of SIMATS, and Dr Selva Kumar, PSG Institute of Advanced Studies, for their help with ICP-OES for tissue aluminium measurements.
Authors’ Contributions
LDVN was involved in the concept, design, acquisition, analysis, interpretation of data and drafting article. SS was involved in design, analysis of data and critically reviewing the intellectual contents. DSK was involved in concept, acquisition, help in animal work and data interpretation. All authors approved the final version of the article for publication and agreed to be accountable for all aspects of the work and resolved any issues related to its accuracy or integrity.
Note: DSK after the completion of the study had moved on to the Department of Research, Panimalar Medical College, Chennai, Tamil Nadu, India.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
Statement of Ethics
There are no human participants in this article and informed consent is not required. The SIMATS University Institutional Animal Ethics Committee approved the experimental procedures used in this study (approval no. SU/CLAIR/RD/002/2022) in 2022. All animal housing and experiments were conducted in strict accordance with the Institutional Animal Ethical Committee, SIMATS University, following the Committee of Control and Supervision of Experiments on Animals (CCSEA, India) which comply with National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Supplementary Material
Supplementary material for this article is available online.
Annexure 1
Annexure 2: Rat Activity Coding
The coding was done as shown below-
The person recording the behavioral parameters during the test was blinded to the group selection and treatment conditions and the information were decoded later.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.