
Abstract
Background
Autism spectrum disorder (ASD) is a multifactorial neurodevelopmental disorder. Numerous environmental risk factors have been implicated in the development of ASD in human cases, suggesting that multiple aetiological agents may contribute to developing an ASD rodent model. In this study, we have investigated the pathobiological processes and outcomes in a novel rat model of ASD that incorporates clinically relevant prenatal and perinatal insults. Specifically, a gamma-aminobutyric acid modulator, valproic acid (VPA), used for perinatal insults, and postnatal hypoxia was combined to understand the critical stages of brain development in rat pups and to explore potential causal mechanisms underlying ASD.
Purpose
To develop the robust animal model for ASD using VPA and hypoxia as causative agents in combination to test other efficacious medicines for ASD in future.
Method
A total of 16 female and 4 male Wistar rats were selected for mating, and the delivered pups were used for the study. The pregnant female animals were divided into four groups: G1: normal control group, G2: VPA group—received a single injection of VPA intraperitoneal (i.p.) on gestational day (GD) 12.5, G3: hypoxia group, rat pups were subjected to postnatal hypoxia, G4: VPA + hypoxia group, pregnant females received VPA (i.p. on GD 12.5), followed by normal delivery, after which the pups were subjected to postnatal hypoxia. Neurodevelopmental reflex tests, neurobehavioural assessments (open field test and Morris water maze) and three-chamber social interaction tests were conducted on the rat pups. Histopathological evaluation of the pups’ brains was performed.
Results
The VPA, hypoxia and combined VPA + hypoxia groups exhibited significant neurobehavioural alterations, with marked delays in neurodevelopmental reflexes compared to the normal control group. Histopathological examination of the brain revealed a reduction in Purkinje cells within the Purkinje cell layer, along with extensive neuronal degeneration. The severity of both neurobehavioural deficits and histopathological abnormalities was greatest in the combined VPA + hypoxia group compared to the other experimental groups.
Conclusions
These findings suggest that the combination of prenatal VPA exposure and postnatal hypoxia, representing pre- and perinatal insults, induces ASD-like conditions in rat pups that closely resemble the clinical behaviour features and redox imbalance of human ASD. Thus, this combined model could serve as a potential animal model for ASD and could be used for screening pharmacological agents aimed at treating and managing ASD.
Introduction
Autism spectrum disorder (ASD) is a multifactorial neurodevelopmental condition caused by both genetic and non-genetic factors that affects approximately 1 in 31 children aged 8 years. 1 In India, the prevalence is estimated at 61 per 10,000 (1 in 156), 2 whereas Hong Kong reports the highest global prevalence, at 372 per 10,000 (1 in 27). ASD is about four times more common in boys than in girls. It is characterised by restricted and repetitive behaviours, along with impairments in social interaction and communication that typically manifest in early childhood and significantly interfere with daily functioning. 2
ASD presents with substantial clinical heterogeneity, reflecting its multifactorial aetiology. 3 Affected individuals exhibit a wide spectrum of symptoms, functional disabilities, severities and intellectual levels, often accompanied by gastrointestinal, immunological and metabolic comorbidities.4–8 The disorder originates during early embryogenesis, beginning with disruptions in cell proliferation and differentiation, and progresses through prenatal and postnatal stages of brain development. This cascade of events leads to aberrant neural migration, synaptic formation, laminar disorganisation and reduced neural network functioning. 3
Associated symptoms commonly observed in ASD are compulsions, mood disturbances, gastrointestinal problems, anxiety, depression, irritability, aggression, redox imbalance and sleep abnormalities. 9 Currently, clinical diagnosis relies on psychometric tests that evaluate the presence or absence of core behavioural features, with the Diagnostic and Statistical Manual of Mental Disorders serving as the gold standard. 2
To date, the precise aetiology of ASD is highly complex and remains unclear. As the prevalence of ASD continues to rise, there are a number of hypothesised aetiological factors, encompassing both genetic and environmental influences. Given the growing evidence of ASD comorbidity and phenotypic overlap with other neurological disorders, it is essential to explore the underlying neurobiological mechanisms driving ASD phenotypes. Developing robust animal models is critical for this purpose, though interpretations of animal research must be made cautiously.
Administration of valproic acid (VPA) to pregnant rodents at a critical gestational time point is one of the most widely used experimental approaches to model ASD. 10 This method stems from pharmacoepidemiological evidence demonstrating a strong association between maternal exposure to VPA during pregnancy (used for epilepsy, mania or migraine) and the subsequent development of ASD in offspring. However, existing VPA-based models vary in terms of dosage, route of administration, gestational timing and behavioural analysis.
Other investigations further reported associations between ASD and pregnancy complications, particularly those involving foetal hypoxia. 11 Several studies suggest that perinatal asphyxia or hypoxia, which can lead to neonatal encephalopathy in term infants, is associated with later neurodevelopmental disorders. Notably, even moderate neonatal encephalopathy following asphyxia can result in cognitive and sensorimotor impairments. In one clinical study of 239 children who survived neonatal encephalopathy, 4.2% were diagnosed with ASD.
Most of the experimental models of ASD rely on a single causative factor, either chemical or genetic, which limits their construct validity. Behavioural testing alone does not fully capture the complexity of autism pathology. For a successful animal model, construct, face and predictive validity must be achieved by replicating behavioural, neuropathological and biochemical features of ASD. The use of multiple aetiological agents could therefore yield a more representative and reliable model. 12
In the present study, we aimed to develop a novel and robust rodent model of ASD by combining prenatal exposure to VPA with perinatal hypoxic insult, thereby mimicking clinically relevant pre- and postnatal conditions observed in human ASD by enhancing oxidative stress in the brains of experimental animals. This model mimics the disruptions in redox homeostasis, thereby promoting an ASD-like scenario. 13 However, numerous experimental models have employed VPA alone to induce ASD-like features, which reflect the repetitive pattern and social interaction to some extent. To the best of our knowledge, no previous studies have considered a combined hypoxia and VPA approach, which uses two different aetiologies and thereby establishes ASD features in experimental rats. This approach seeks to provide a more comprehensive model for investigating the underlying mechanisms and potential therapeutic interventions for ASD.
Methods
Chemicals
Induction agent, VPA, was procured from Sigma–Aldrich Chemical Pvt. Ltd, Bangalore. All other chemicals and reagents used were of analytical grade.
Laboratory Animals
The study was approved by the Institutional Animal Ethics Committee (IAEC) (Approval No.: IAEC/70/SRIHER/846/2023), Sri Ramachandra Institute of Higher Education and Research, Chennai, and was conducted as per the ethical norms of the Committee for Control and Supervision of Experiments on Animals (CCSEA) and ARRIVE guidelines. Totally, 16 adult female Wistar rats (age: 11–12 weeks) and 4 male Wistar rats weighing ≤200 g were procured from the CCSEA-approved vendor (Invivo Biosciences, Bangalore, CCSEA Registration No: 1165/PO/RcBiBtS/NRc-L/08/CPCSEA). Animals were acclimatised in the study room for 7 days before mating, following which they were permitted to mate in a 2:1 ratio. For breeding purposes, males were used and then were precluded from the experiment. Animals were sustained under a regulated environmental condition of temperature and relative humidity, 19°C–23°C and 30%–70%, respectively. 12–15 air changes/h were maintained throughout the experiment, including a 12 h light artificial photoperiod and 12 h dark. Animals were provided with ad libitum laboratory rodent pelleted feed procured from an authorised vendor, and ultraviolet-treated reverse osmosis water in autoclaved bottles.
Rat Breeding and Weaning
Every morning, female animals were inspected for signs of pregnancy, which were confirmed by checking vaginal smears for spermatozoa or by examining the vaginal plug. Mating confirmation was identified as the 0.5th day of gestation. Pregnant rats were kept apart until their delivery. On postnatal day (PND) 23, the pups were weaned, and both female and male progeny were considered for the study.
Autism Spectrum Disorder Model
After pregnancy, pregnant rats were grouped into four (G1–G4): for G1 (control group), pregnant female rats were administered a single intraperitoneal (i.p.) injection of an equivalent quantity of normal saline at the G12.5 days of gestational time point to serve as the vehicle control. In G2 (VPA group, prenatal), on G12.5 days, 600 mg/kg of sodium VPA dissolved in newly made 0.9% saline was administered intraperitoneally (i.p.) to pregnant female rats. In all groups, pregnant rats were monitored closely on a daily basis till normal delivery. In G3, the hypoxia group, pregnant rats were allowed for normal delivery, and neonates from PND2 to PND6 were subjected to a hypoxia insult by placing in a hypoxic chamber (ProOx P360; Biospherix; USA) with 8% oxygen for 30 min. In G4 (VPA + hypoxia group, pre- and post-insult), pregnant rats were insulted by a single injection of VPA, 600 mg/kg (i.p.) administrated on G12.5 days, and then were allowed for normal delivery. After that, neonates from PND2 to PND6 were subjected to a hypoxia insult with 8% oxygen for 30 min once daily in a hypoxia chamber. For hypoxia exposure, pups were placed in a small container with bedding material and transferred to a preheated hypoxic chamber (ProOx P360; Biospherix, USA) set to 8% oxygen for 30 min. Throughout the exposure period, pups were continuously monitored, and the chamber temperature was maintained at approximately 37°C. After hypoxia exposure, pups were removed from the chamber, monitored and transferred to their respective mothers’ home cages. No mortality or acute stress was evidenced in any pups after hypoxia exposure.
Each group was allocated four adult female rats, and their delivered pups were as follows: Group 1 (normal control), n = 22; Group 2 (VPA), n = 19; Group 3 (hypoxia), n = 22; and Group 4 (VPA + hypoxia), n = 13. Both male and female rat pups were subjected to neurodevelopmental reflex test, open-field test, social interaction test and Morris water maze (MWM) test, and their results were pooled together for further analysis.
After the behavioural test, all rat pups were euthanised using isoflurane, and the brain was collected and fixed in 10% neutral buffered formalin for histopathological analysis.
Neurodevelopmental Reflex Test
Pups in all groups were assessed from PND 3 onwards for neurodevelopmental reflex testing. 14 All reflex tests were performed on each pup until a positive reaction was seen (Figure 1). The reflex test is qualified when a positive result is obtained. Following the favourable response, no additional testing was conducted.
Representative Image of Neurodevelopmental Reflex Test Performed in Wistar Rat Pups: (A) Forelimb Grasping, (B) Hind Limb Grasping, (C) Righting, (D) Hind Limb Placing, (E) Cliff Avoidance, (F) Gait, (G) Auditory Startle, (H) Posture, (I) Eye Opening.
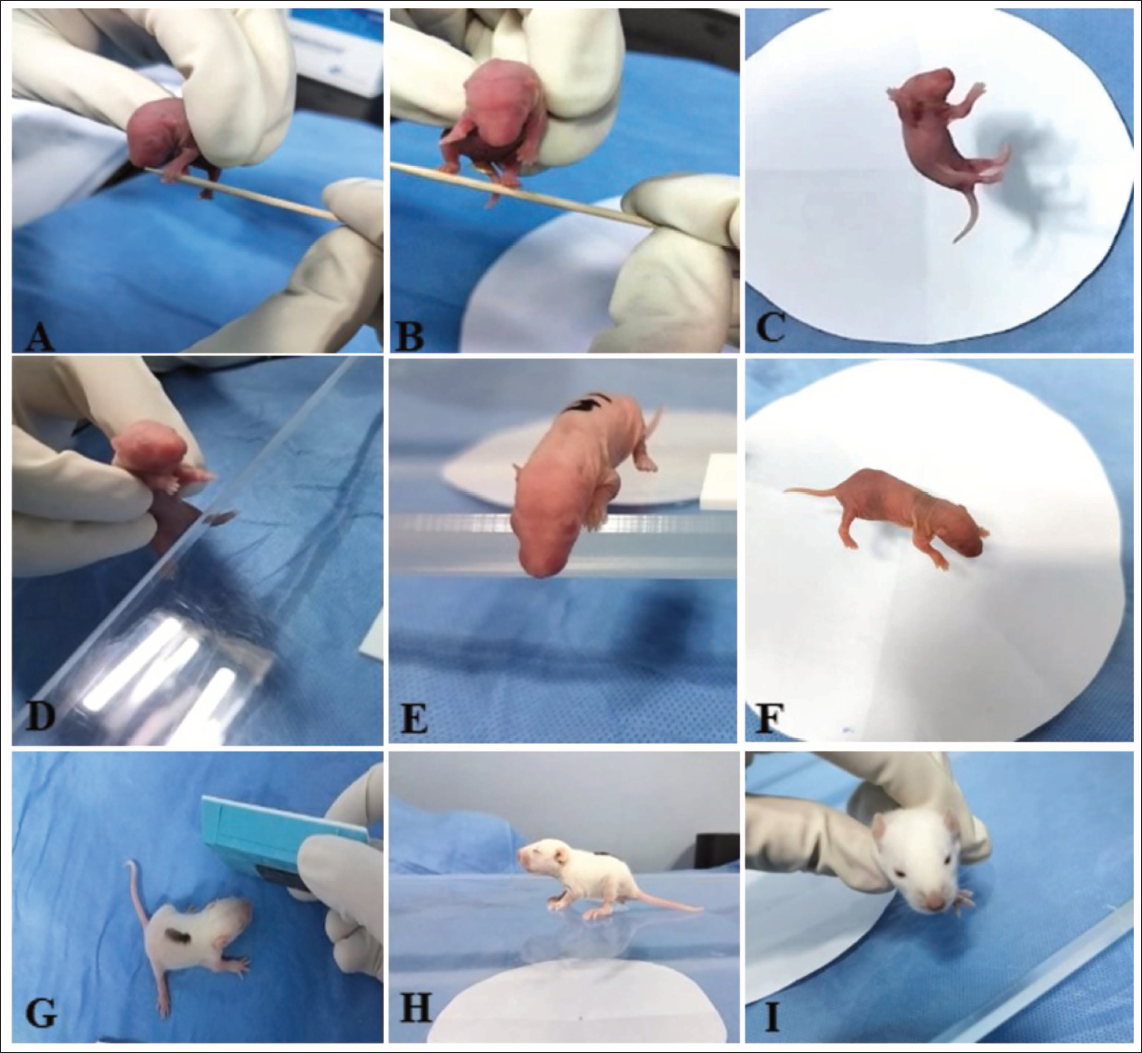
Forelimb Grasping
The forelimb grasping reflex test was conducted on PND 3 to evaluate neuromuscular development and reflex integrity in pups. This test provides an effective measure of sensorimotor function, enabling the assessment of early development of motor and the detection of potential neurological or developmental impairments. During the test, gentle pressure was applied to the forelimbs using a blunt cylindrical rod, eliciting a reflex characterised by forelimb digit flexion. Reflex responses were scored on a three-point scale: 0 for no grasping, 1 for successful grasping with a single forepaw, and 2 for successful grasping with both forepaws.
Hindlimb Grasping
The hindlimb grasping reflex test was also performed on PND 3 to evaluate neuromuscular function and reflex development in rat pups. This test provides a straightforward assessment of hindlimb motor function and reflex integrity, aiding in the detection of developmental or neurological abnormalities. A blunt cylindrical rod was gently applied to the hindlimbs with mild pressure, eliciting a reflex characterised by hindlimb digit flexion. Reflex responses were scored on a scale of 0–2, with a score of 0 indicating no grasping, 1 indicating successful grasping with one hind paw, and 2 indicating successful grasping with both hind paws.
Righting
The righting reflex test was conducted on PND 3 to evaluate the developmental progression of motor coordination and postural control in rat pups. This test provides data on the pup’s neuromuscular development and its ability to perform postural adjustments, offering insight into potential developmental or neurological impairments. Each pup was gently held upright and then released onto a flat surface, allowing it to attempt to orient itself to a normal standing posture. The time taken to complete the task was recorded, with a maximum duration of 15 s allotted for each pup. A positive righting reflex was defined as the pup successfully orienting itself to stand on all four paws. Reflex performance was assessed using a three-point scoring system: a score of 0 was assigned if the pup remained lying on its back, 1 if the pup lay on its side (either left or right) or managed to right itself but with an incorrect posture, and 2 if the pup successfully achieved the correct standing posture.
Hindlimb Placing
The hindlimb placement reflex test was conducted on PND 4 to assess the development of sensorimotor integration and reflexive responses in rat pups. Each pup was gently held vertically by the torso to allow free movement of the hindlimbs. A rough surface, like as the edge of a table, was used to lightly stroke the dorsum of one of the hind paws, stimulating a reflexive response. A correct placing reflex was defined as the withdrawal of the stimulated hind limb, immediately followed by its placement onto the surface. Reflex responses were scored using a three-point scale: a score of 0 indicated the absence of any reflex or movement, 1 indicated the successful placement of one hind paw (either left or right), and 2 indicated the placement of both hind paws. This test provides an effective measure of the maturation of the sensory and motor pathways involved in coordinating hind limb movement, offering valuable insights into the pup’s neuromuscular development and potential impairments in reflex behaviour.
Cliff Avoidance
The cliff avoidance reflex test was initiated on PND 4 to evaluate the development of protective motor responses and spatial awareness in rat pups. This test provides a critical view of the pup’s developing ability to respond to environmental stimuli, as well as its motor coordination and reflexive behaviours, which are essential for avoiding potential hazards. Each pup was carefully placed on a flat surface with its snout and forepaws positioned at the edge of the surface, simulating a ‘cliff’ scenario. This setup was designed to provoke a natural protective response, where the pup perceives the potential risk and turns or moves away from the edge. Reflex performance was assessed using a three-point scoring system: a score of 0 indicated no protective movement or the pup falling off the edge, 1 indicated an attempt to move away from the edge but with limbs hanging over the edge, and 2 indicated a successful and complete movement away from the edge of the surface.
Gait Test
The gait assessment was conducted on PND 6 to evaluate motor coordination and locomotive ability of the pups. This test provides evidence on the pup’s developing motor skills, offering insights into its neuromuscular coordination and locomotive progression at this early stage of life. Each pup was gently placed at the middle of a circle with a diameter of 15 cm on a flat surface and observed for its ability to navigate and move out of the circle within a time frame of 30 s. The goal of the test was to assess the pup’s ability to coordinate forepaw movements effectively to achieve forward locomotion. Reflexive and motor performance was scored using a two-point scale: a score of 0 was assigned within 30 s if the puppy could not move both forepaws outside the circle, indicating a delay or lack of sufficient motor function, while a score of 1 was assigned for a successful gait, defined as the pup moving both forepaws outside the circle within the allotted time.
Auditory Startle
The auditory startle reflex test was conducted on PND 10 to assess the pups’ auditory function and the integration of sensory and motor pathways involved in reflexive responses to sudden stimuli. This test provides critical insights into pups’ sensory-motor development and the maturation of their auditory system at this developmental stage. Each pup was exposed to a sudden, loud noise of sufficient intensity to provoke a startle response, which is a natural reflex triggered by unexpected auditory stimuli. The test aimed to determine whether the pups could perceive the sound and respond appropriately. A positive startle response was characterised by observable movements such as a sudden jerk, twitch or other reflexive motions in reaction to the noise, indicating functional auditory pathways. The scoring system was binary: ‘No’ for the absence of any startle response, suggesting impaired or underdeveloped auditory or motor function, and ‘Yes’ for a positive startle response, confirming the presence of functional auditory reflexes.
Posture
The posture analysis was conducted on PND 12 to evaluate the motor development and postural control of the pups during movement. This assessment provides details on the developmental progression of motor coordination, strength and postural stability in the pups. Each pup was placed on a flat, stable surface and observed for its ability to lift and maintain its abdomen off the ground while moving, a key indicator of maturing motor coordination and strength. A fully mature posture was defined as the abdomen being elevated from the surface or positioned parallel to the body, reflecting the pup’s ability to engage in coordinated and balanced locomotion. The scoring system for posture assessment was based on a three-point scale: a score of 0 indicated the absence of movement or any attempt to lift the abdomen, suggesting a lack of motor maturity; a score of 1 indicated partial posture, where the pup demonstrated some elevation of the abdomen but did not maintain a fully mature posture while moving; and a score of 2 indicated a fully mature posture, with the abdomen lifted and maintained parallel to the body during locomotion.
Opening of Eye
The analysis of eyelid opening was conducted on PND 12 to assess the developmental progress of sensory and motor systems in rat pups, particularly the maturation of the ocular system. During this assessment, the pups were gently observed to determine the extent to which their eyelids had opened, which is a critical milestone in sensory development. The scoring system for eyelid opening was as follows: a score of 0 was given if neither eye was visible, indicating that the eyelids were still closed or only partially open, which is typical during the early stages of postnatal development; a score of 1 was assigned if one eye was visible, suggesting partial opening of the eyelid on one side, indicating some progress toward ocular maturation; and a score of 2 was given when both eyes were visible, signifying that the pup had achieved full eyelid opening, a sign of maturation and readiness for sensory exploration of the environment.
Open Field Test
On PND 29, rat pups were evaluated for their exploratory and motor behaviours in an open-field chamber (60 × 60 × 45 cm) over a 5-min observation period, a widely used test for assessing anxiety-related activity and exploratory behaviour in rodents. During the test, the following behaviours were recorded.
Number of Squares Crossed
Movement in the arena is an indicator of the pup’s activity level and willingness to explore its environment. The chamber floor was divided into a grid of squares, and a count was made every time the pup crossed a square with both its forelimbs. This measure reflects the pup’s locomotor activity, motor coordination and exploratory drive.
Number of Entries to the Central Zone
The central section of the open field was defined by the central four squares, and entries into this zone were recorded. The central zone is often used to assess anxiety-related behaviour, as animals typically avoid this area when stressed or anxious. Increased time spent in, or entries into, the central zone are usually associated with lower anxiety levels and greater exploration, while fewer entries may suggest heightened anxiety or caution.
Number of Rearing
Rearing, where the pup stands on its hind limbs, sometimes leaning against the wall with its forelimbs, was recorded. This behaviour indicates exploratory motivation and the ability to balance and orient itself in space. It can also reflect curiosity and engagement with the environment, as the pup may rear to look around, sniff, or explore the surroundings in a more vertical position.
Grooming
Grooming behaviours were noted, as they can serve as indicators of the emotional or physical state. Grooming in rodents is a natural behaviour that can be used to assess the pup’s comfort level. Excessive grooming can sometimes be indicative of stress, while normal grooming patterns suggest a neutral or positive state.
Faecal Count
This measure can be an indirect indicator of anxiety or stress, as rodents often defecate more frequently under stress or in novel environments.
Urination Occurrence
Urination is a common stress response in rodents when they are placed in unfamiliar or mildly stressful environments, and this measure can provide an idea of the emotional or physiological state of the pup while in a distress situation. Urination of pups, if evidenced or not during the test, was also recorded, with a score of 1 indicating urination and 0 indicating no urination.
This 5-min open field test allows for the observation of a range of behaviours related to motor activity, exploration, anxiety and general well-being. It provides a comprehensive assessment of the pup’s ability to interact with its environment and serves as an indicator of overall health, development and sensory-motor integration at this stage.
Three-chamber Sociability and Social Novelty Preference Tests
Pups were assessed for a social interaction test (3-CST) on PND 31.
15
One of the main characteristics of autism is social interaction impairment. It was evaluated using a three-chambered apparatus that measured 60 × 40 × 25 cm and was separated into three communicating chambers. The animals were allowed to move across the three-chambers thanks to apertures between the compartments. Each of the lateral sections had a transparent cage. Between test animals, the chambers were cleaned with 70% alcohol. The test was divided into three stages:
Habituation phase: Prior to the sociability phase, the tested rats were put in the central compartment and waited for 5 min to explore. Sociability phase: A new rat was placed in a separate transparent cage of the lateral chambers to represent the new rat, while the other transparent cage was leaving empty to represent the new item, starting shortly after the habituation phase. The matter of time spent examining the new or novel object or rat one was analysed to determine the sociability index (SI) after the analysed rat was left for 10 min. Social novelty preference phase: Proceeded for 10 more minutes right after the sociability phase. Rat 2 was a new, unknown rat that was put into the clear cage that was unoccupied in the other chamber. In order to determine whether the tested rat preferred social novelty when interacting with the novel animal (novel rat 2) or the familiar animal (now known as the known rat), the social novelty preference index (SNI) was calculated during this session.
A blind observation was performed, and the interaction time was recorded manually for each phase. The following formulas were used to determine the SI and SNI:

Morris Water Maze Test
The pups’ learning and spatial memory were assessed on PNDs 33, 34 and 35. 16 For the water maze, the animals had already been trained. The pups were put on a pedestal in the middle of the pool with water. The animals undergone three trials in a row. The animals were initially kept on the platform for 20 s. The animal was then carried to one of the four points (north, south, east or west) and supported by hand before being lowered into the water. The animals were allowed to look for the platform for 60 s. To pretrain the animals, the process was carried out twice more. After that, the platform was removed from each animal to conduct a probing trial. For 30 s, the number of times the animal crossed the centre was noted.
Histopathology
At the end (PND 36), the pups’ brains were sacrificed by using isoflurane euthanasia. A 10% neutral buffered formalin solution was used to fix the brain tissue samples. The tissues were sectioned to a thickness of approximately 3–4 µm and immersed in paraffin wax, and stained with haematoxylin and eosin (H&E), toluidine blue and cresyl violet for the purpose of evaluating the brain histopathology. The morphology of the brain tissue was studied by looking at the stained slides under a light microscope. Scoring was done by a pathologist without revealing the group information. The following semi-quantitative grading system was used for histopathological examination of brain tissue: (a) minimal (<10% of tissue affected), (b) mild (approximately 10%–25% of tissue affected), (c) moderate (26%–50% of tissue affected), (d) marked (51%–75% of tissue affected), and (e) severe (>75% of tissue affected).
Statistical Analysis
Values are presented as the mean ± standard deviation in tables and the mean ± standard error of the mean (SEM) in graphical representations. One-way analysis of variance with Tukey’s post hoc multiple comparison test was used to ascertain the statistical significance of the mean difference between the control and induction groups using SigmaPlot 14.5 software. The significance threshold was established as a p value below .05, .01 and .001.
Results
Neurodevelopmental Reflex Test
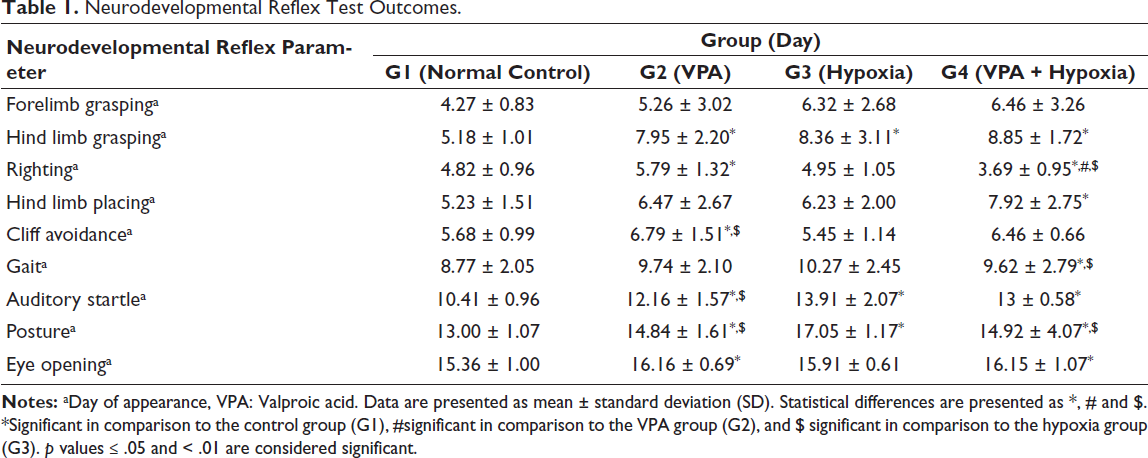
Neurodevelopmental reflex testing was conducted on rat pups beginning on PND 3. The presence of a positive reflex response was recorded and scored (Table 1, Figure 2).
Neurodevelopmental Reflex Test Outcomes.

Forelimb Grasping
A significant difference (p ≤ .05) was observed among the experimental groups in forelimb grasping performance.
Hind Limb Grasping
Rats in the VPA (G2), hypoxia (G3) and combined VPA + hypoxia (G4) groups exhibited significant motor developmental delays in hind limb grasping compared with the normal control (G1) group (p < .001).
Righting
The VPA group (G2) demonstrated a significant delay in righting reflex compared with the normal control group (G1) (p ≤ .001). The combined VPA + hypoxia group (G4) showed a significantly greater delay in righting reflex compared with G1, G2 and G3 (p ≤ .001). No significant difference was found between the hypoxia groups (G3) and the G1 or G2 groups.
Hind Limb Placing
A significant delay (p ≤ .01) in hind limb placing was observed in the combined VPA + hypoxia group (G4) compared with the normal control (G1). No significant differences were noted in G2 (VPA) and G3 (hypoxia) compared with G1.
Cliff Avoidance
The cliff avoidance performance of the VPA group (G2) was significantly slower (p ≤ .001) than that of the normal control (G1) and hypoxia (G3) groups.
Gait
A significant difference in gait development was observed in the combined VPA + hypoxia group (G4) compared with the hypoxia group (G3) and the normal control (G1) (p ≤ .01). Delayed gait acquisition was also evident in both the VPA (G2) and hypoxia (G3) groups relative to the normal control (G1).
Auditory Startle
The VPA (G2), hypoxia (G3) and VPA + hypoxia (G4) groups exhibited significant delays in auditory startle responses compared with the normal control (G1) (p < .001). A significant delay was also observed in G3 (hypoxia) relative to G2 (VPA) (p < .001). However, no significant differences were noted between G4 (VPA + hypoxia), G2 (VPA) and G3 (hypoxia) groups.
Posture
Significant delays in postural reflexes were observed in G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) when compared with the normal control (G1) (p < .001). Additionally, G3 (hypoxia) showed a significant delay compared with both G2 (VPA) and G4 (VPA + hypoxia) (p < .001). No significant difference was observed between G2 and G4.
Opening of Eye
Significant (p < .05) delays in eye opening were observed in both the VPA (G2) and VPA + hypoxia (G4) groups compared with the normal control (G1).
Open Field Test
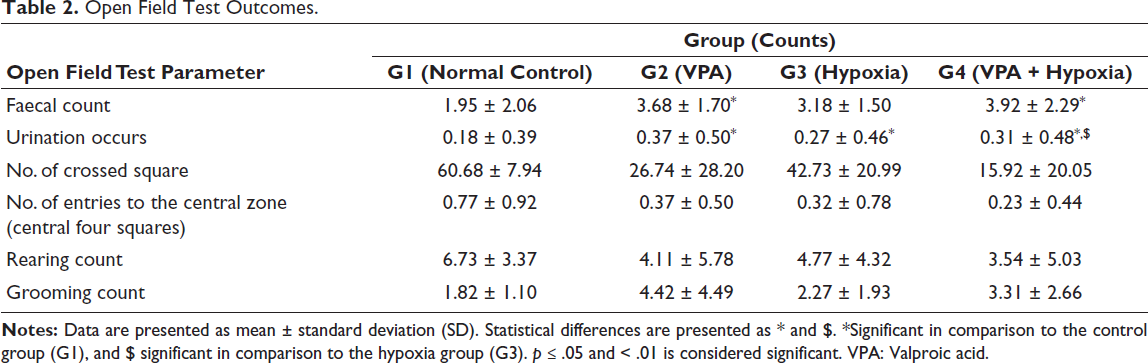
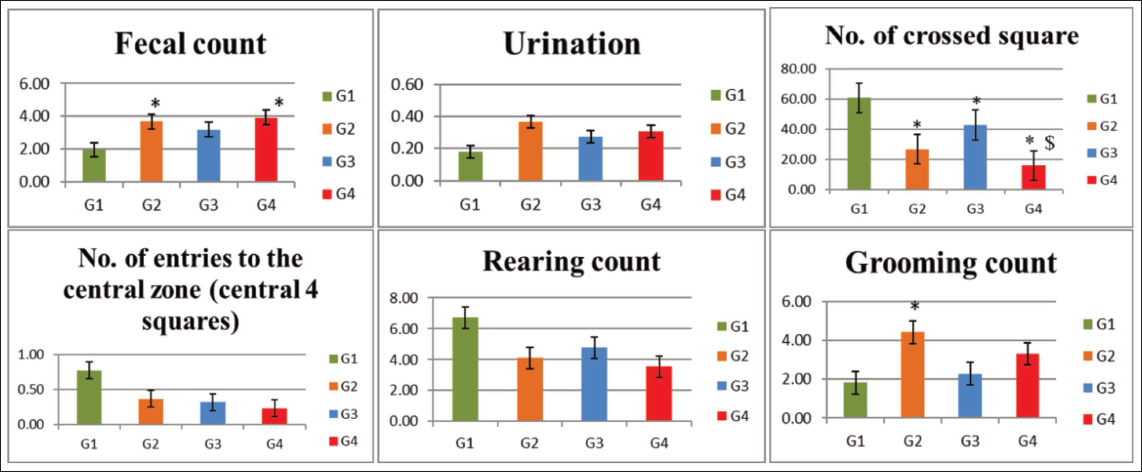
Compared with the normal control group (G1), rats exposed to VPA (G2), hypoxia (G3) and combined VPA + hypoxia (G4) showed a significant reduction in the total number of crossed squares, indicating anxiety-like behaviour (p ≤ .001). The number of crossed squares was significantly (p ≤ .001) lower in the combined exposure group (G4) compared with the hypoxia group (G3).
When compared with the normal control (G1), VPA exposure (G2) resulted in a significant increase in grooming frequency, suggesting enhanced repetitive and stereotyped behaviours (p ≤ .05). Additionally, the number of faecal boli was significantly higher in both the G2 (VPA) and G4 (VPA + hypoxia) groups than in the control group (p ≤ .01). However, no significant differences were observed among the groups in terms of urination frequency, the number of entries into the central zone, or the number of rearing (Table 2 and Figure 3).
Open Field Test Outcomes.
Social Novelty Preference Tests and Three-chamber Sociability
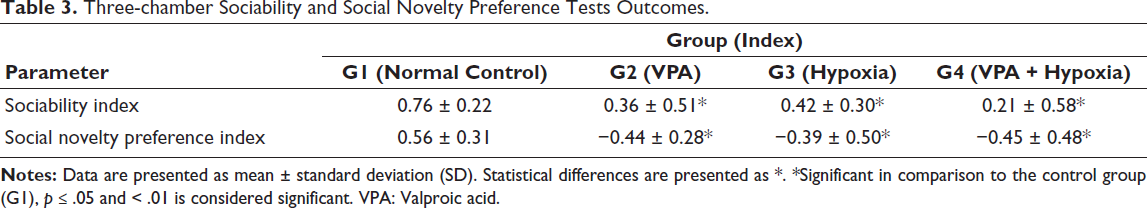
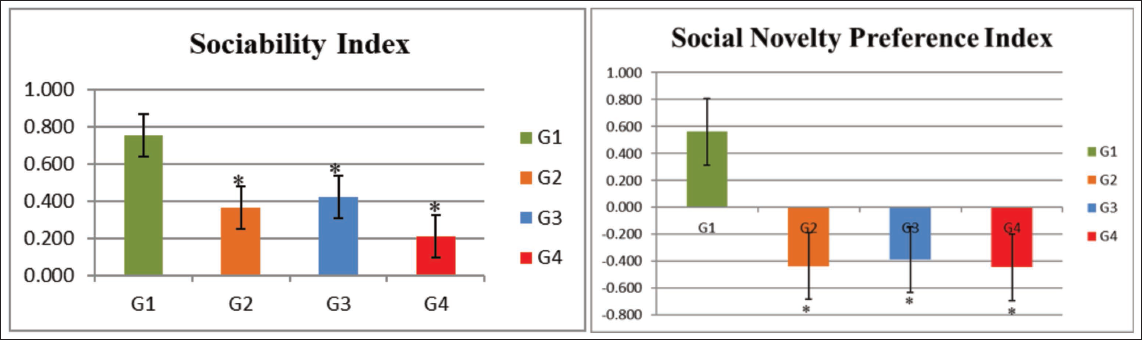
The SI was significantly lower in the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups compared with the normal control (G1) (p ≤ .001).
Similarly, during the social novelty preference phase, the SNI was significantly decreased in the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups relative to G1 (p ≤ .001). The mean values for both sociability and social novelty preference indices were lowest in the combined VPA + hypoxia group (G4), indicating the most pronounced social deficits (Table 3, Figure 4).
Three-chamber Sociability and Social Novelty Preference Tests Outcomes.
Morris Water Maze Test
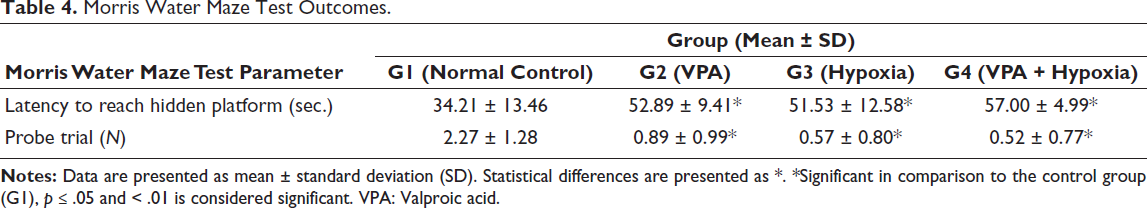
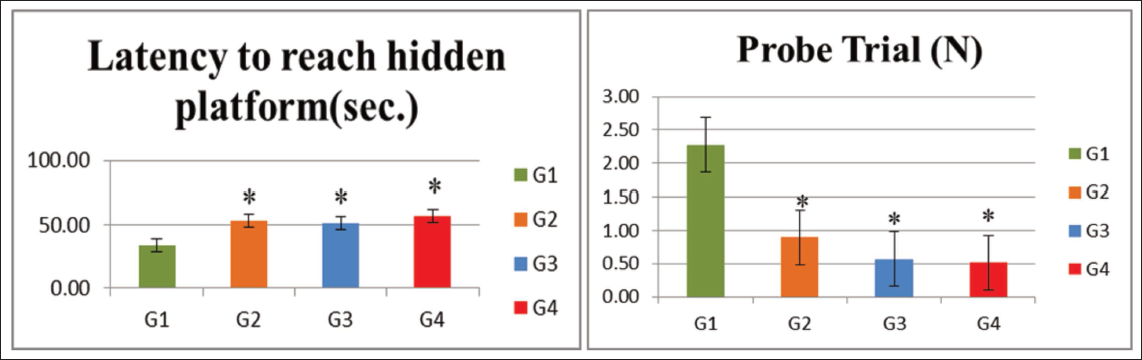
Rats in the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups exhibited significantly (p < .001) longer escape latencies, that is, delays in locating the hidden platform, when compared with the normal control (G1) group, reflecting cognitive impairment.
In the probe trial, the number of times the rats crossed the former platform location within a 30-s period was significantly reduced in G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) compared with G1 (p ≤ .001). Among these, rats in the G4 (VPA + hypoxia) group took the longest time to reach the hidden platform and had the lowest probe trial crossing counts, indicating the greatest level of spatial memory impairment (Table 4, Figure 5).
Morris Water Maze Test Outcomes.
Histopathology Investigation
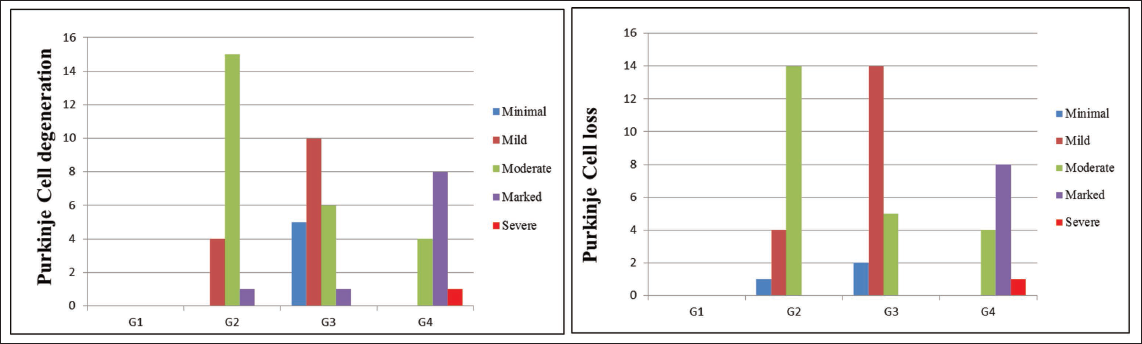
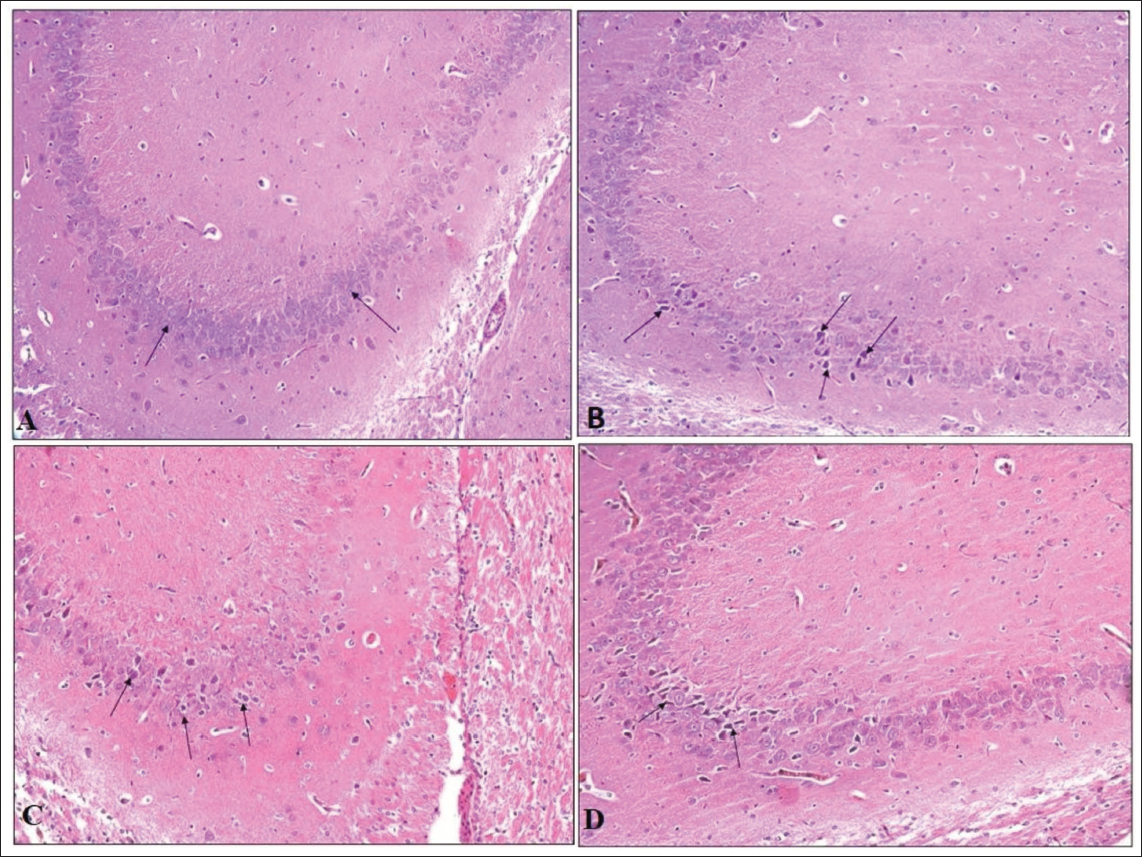
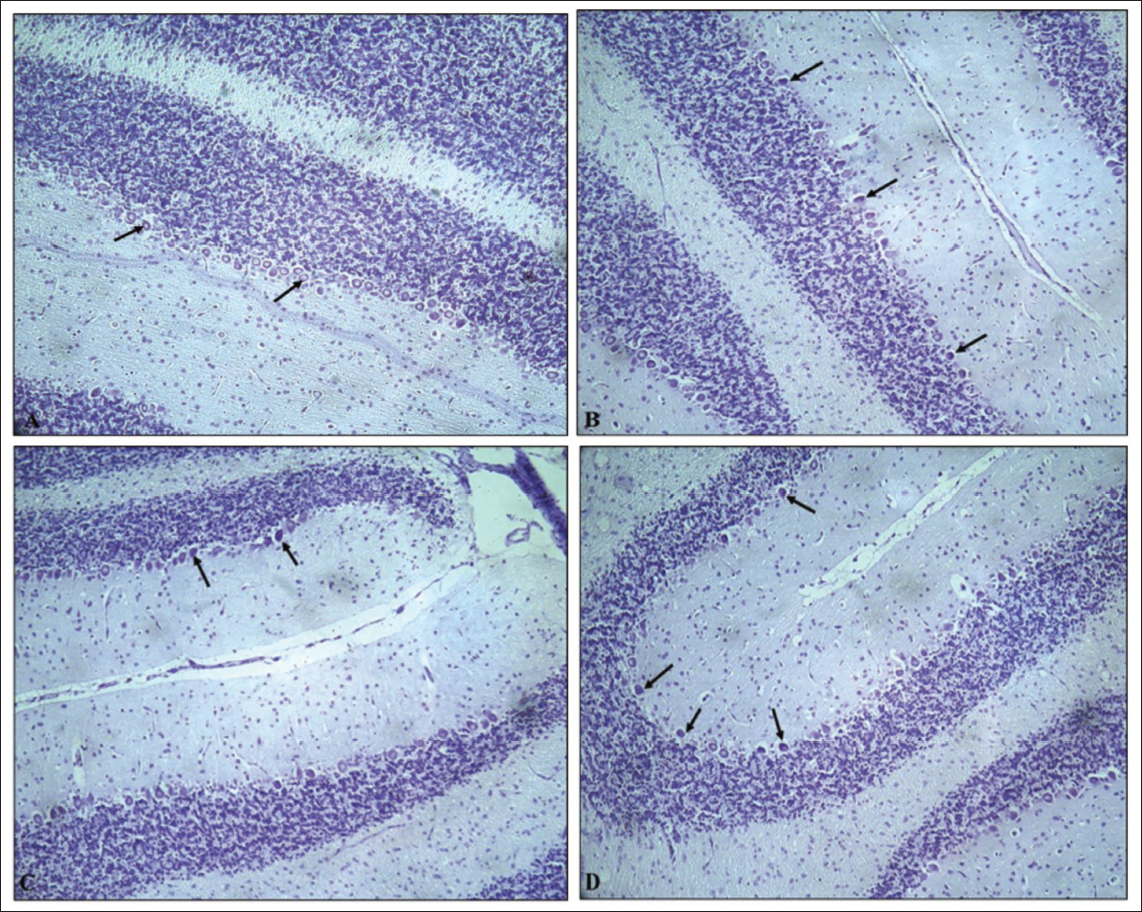
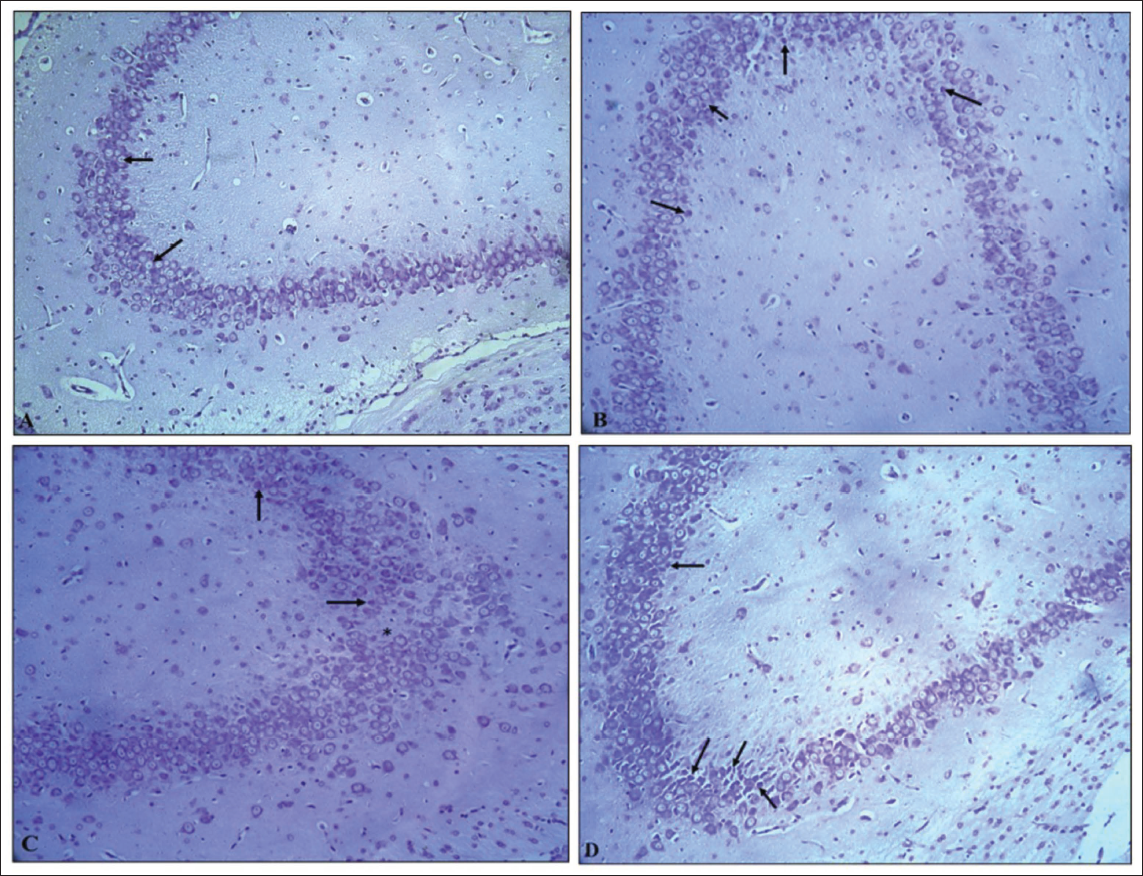
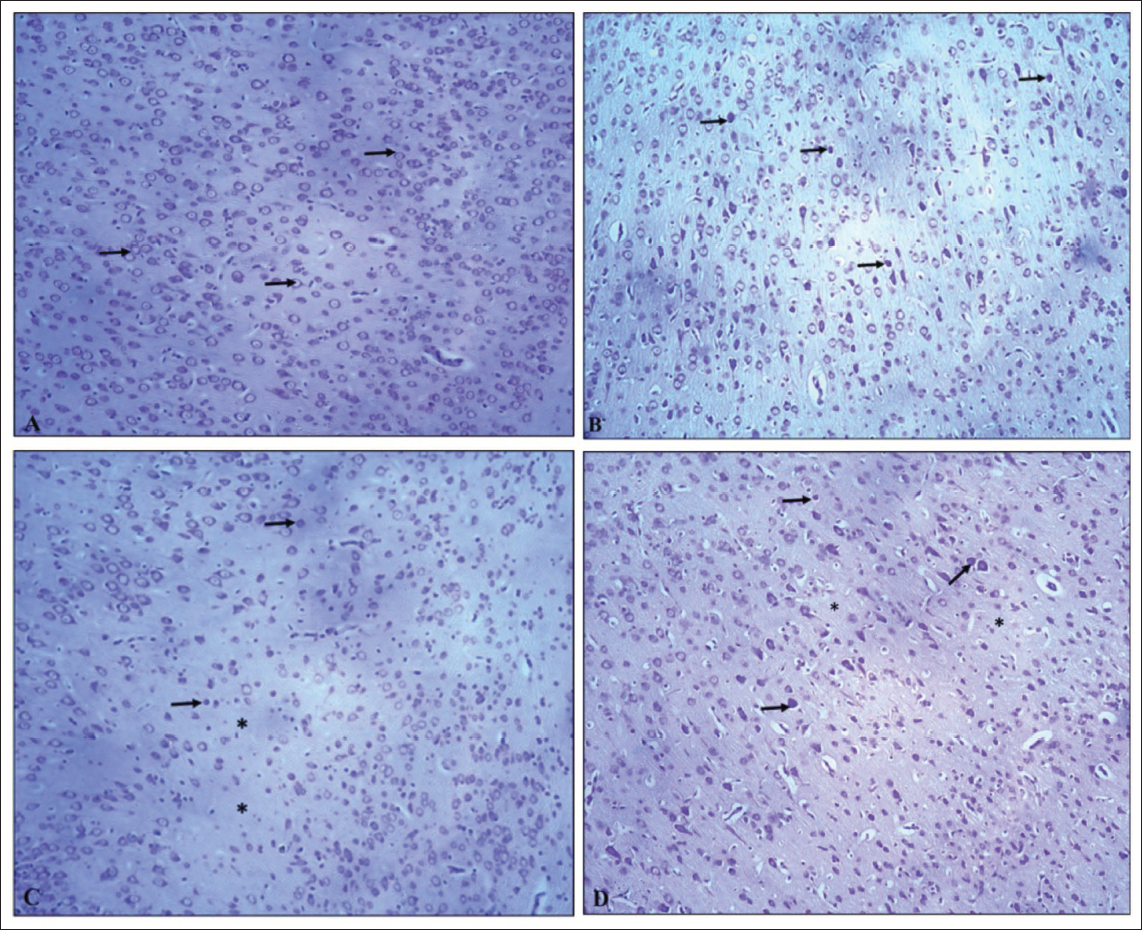
H&E stained pup brain section revealed the inner granular cell layer (GL), middle Purkinje cell layer (PC), and outer molecular layer (ML) comprised the cerebellar cortex of the control group’s brain. Flask-shaped cells with large, rounded, vesicular nuclei in pale cytoplasm comprised the PC layer. The cerebellar cortex of the VPA group (G2) showed mild to marked severity of degenerated PCs, with signs of shrinkage, disfigurement and minimal to moderate severity of loss of PC, along with condensed chromatin. The hypoxia group (G3) showed minimal to marked severity of degenerated PCs, with minimal to moderate severity of loss of PC and spongiosis in the white matter of the cerebellum; the VPA + hypoxia group (G4) showed moderate to severe severity of degenerated PCs, with loss of PC and gliosis (Figures 6 and 7). G4 (VPA + hypoxia) group showed more severity of histopathological findings (PC degeneration and PC loss) compared to G2 (VPA) and G3 (hypoxia) groups (Figure 7). Histopathology of the brain revealed the normal appearance of neuronal cells in the hippocampus of G1 (normal control). G2 (VPA group), G3 (hypoxia) and G4 (VPA + hypoxia) showed neuronal cell degeneration in the hippocampus of the brain (Figure 8).
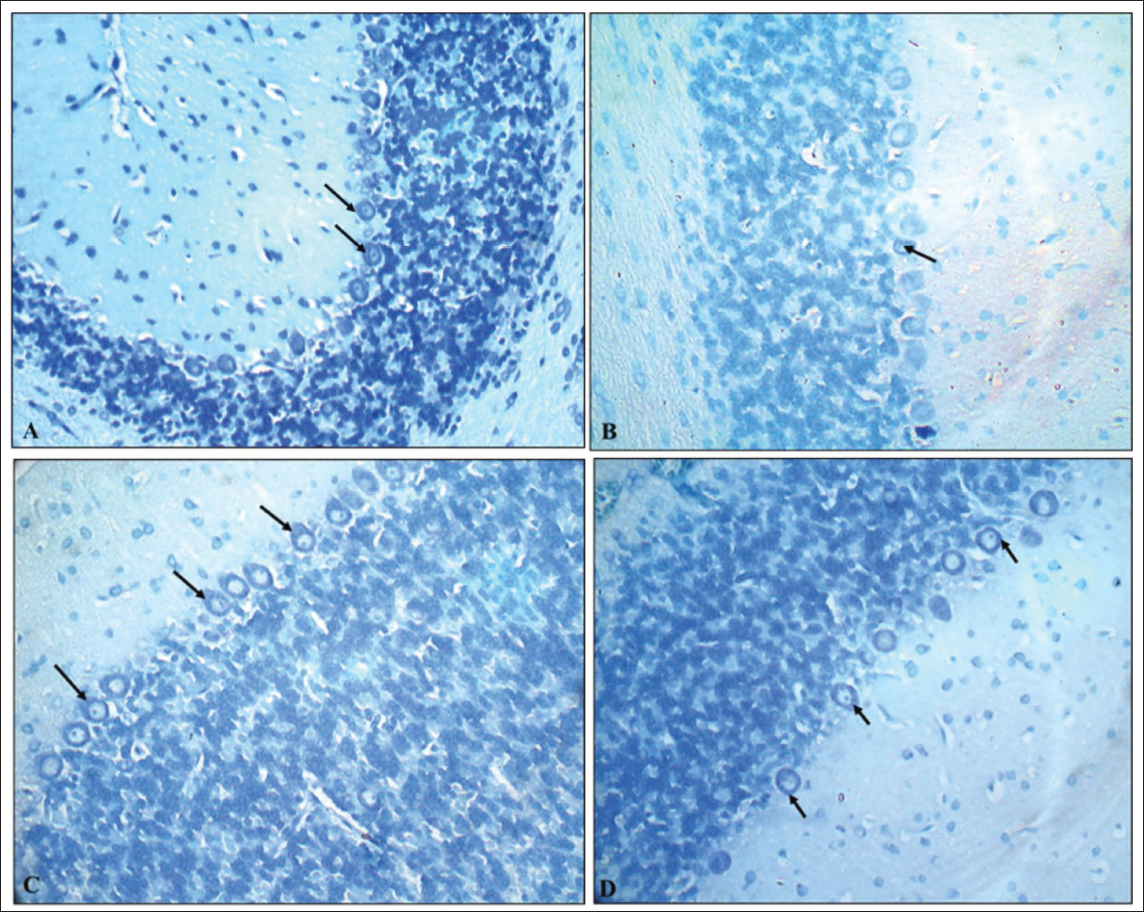
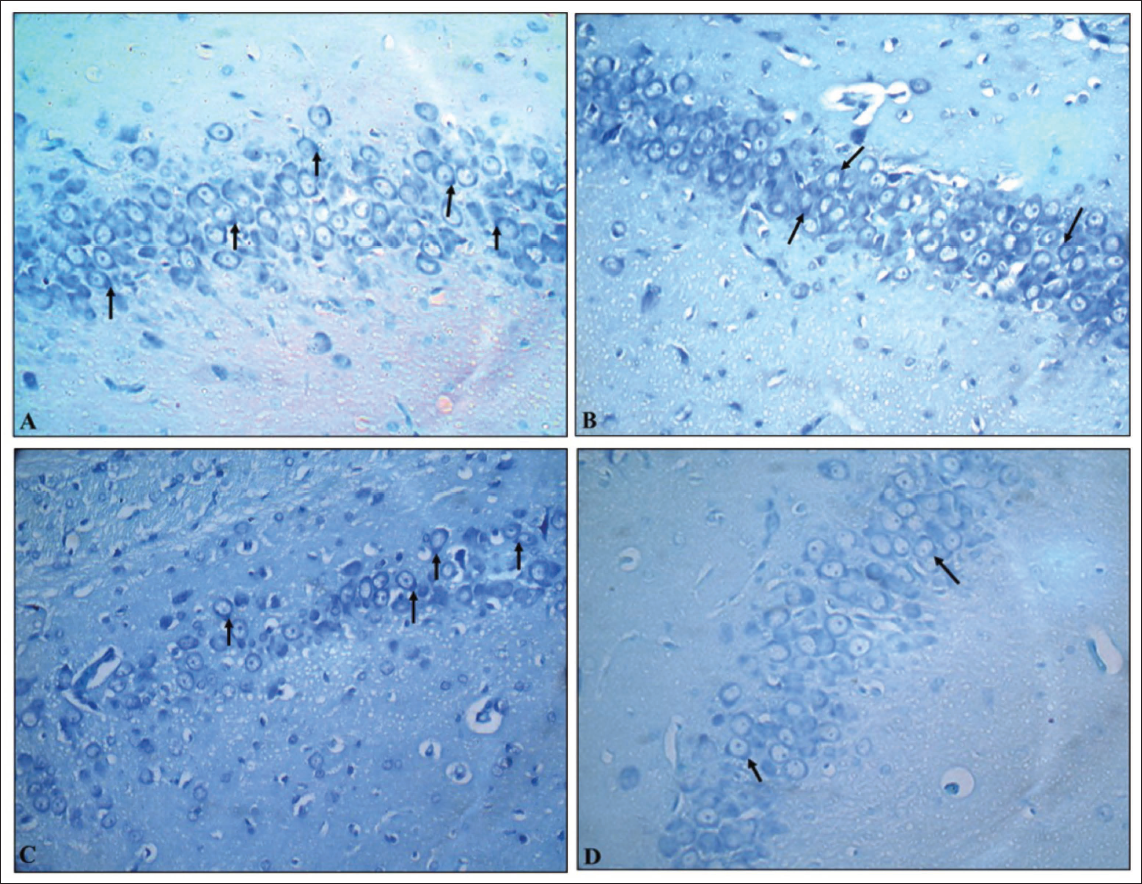
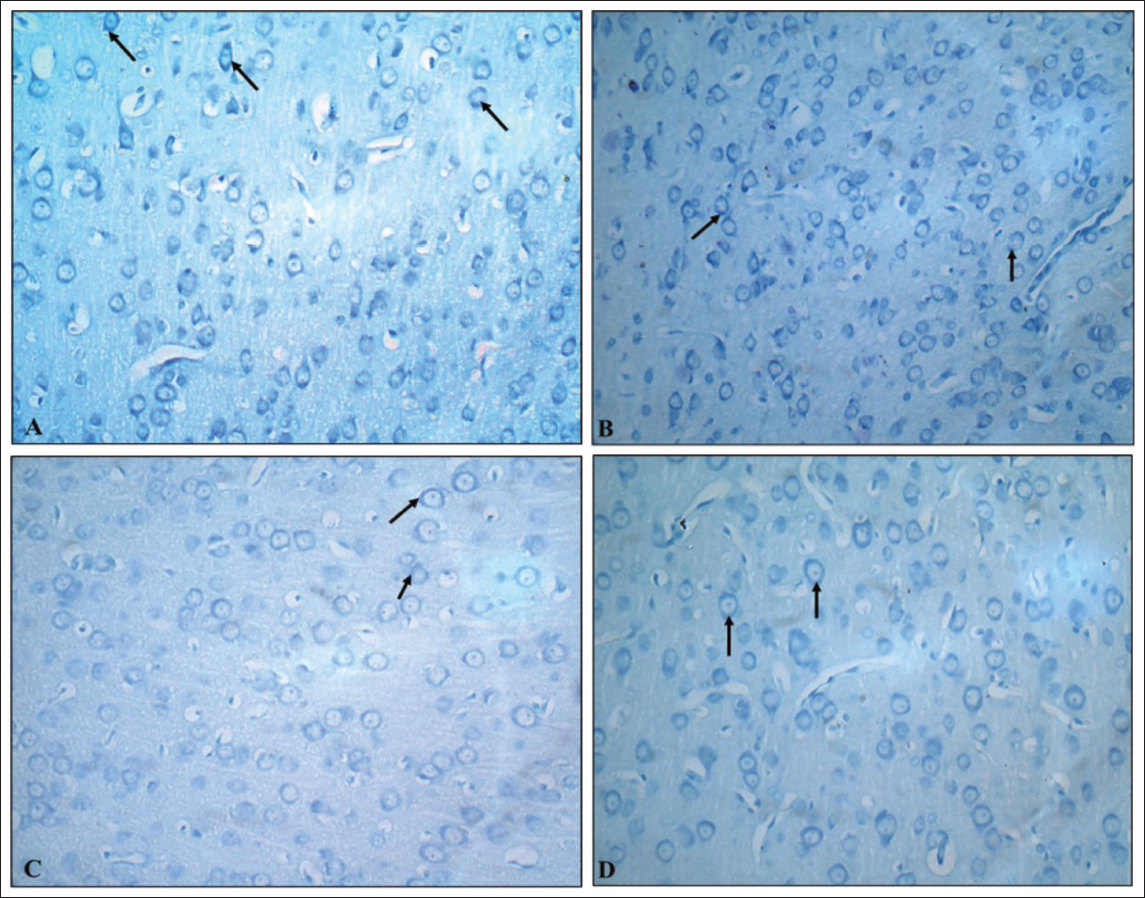
Toluidine blue-stained brain sections of PCs in cerebellar cortex sections of the G1 (normal control) group revealed normal dark blue-stained neurons, while the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups revealed reduced colour intensity when compared to the G1 group (Figure 9). Hippocampal sections (Figure 10) and the cerebral cortex (Figure 11) of the G1 (normal control) group revealed that the majority of neurons in various places were studded with Nissl’s granules, while the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups revealed the least amount of Nissl’s granules in the majority of the neurons.
Cresyl violet-stained brain sections of PCs in the cerebellar cortex, hippocampal and cerebral cortex sections from the G1 (normal control) group revealed normal neuronal cells, while the G2 (VPA), G3 (hypoxia) and G4 (VPA + hypoxia) groups revealed shrunken and darkly stained neurons (Figures 12–14).
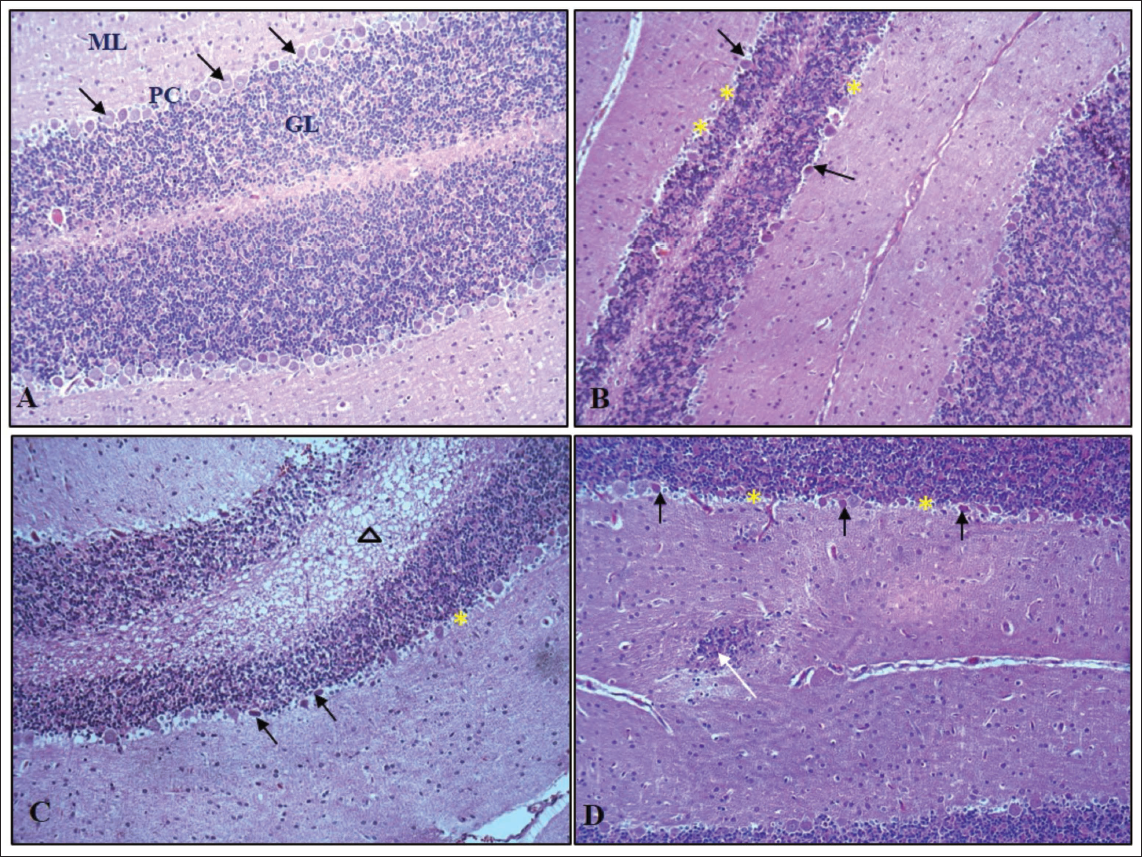
Semi-quantitative Histopathological Grading of Purkinje Cell (PC) in Cerebellum Section of Brain: Minimal: Very Few PC Show Degeneration (Pyknosis, Chromatolysis), <10% of PC Cells Affected and Alignment Mostly Preserved; Mild: Slight Loss and Degeneration of PC (Approximately 10%–25% Affected), Scattered Shrunken or Eosinophilic Cells, Mild Disruption of Linear Arrangement; Moderate: Clear Loss of PC (26%–50% Affected), Noticeable Gaps in the Purkinje Layer, Moderate Vacuolation in Molecular and Granular Layer; Marked: Extensive PC Loss (51%–75% Affected), Severe Cytoplasmic and Nuclear Degeneration, Disorganisation of Cortical Layers; Severe: Almost Complete Loss of PC (>75% Affected), Extensive Vacuolation and Prominent Gliosis.
Discussion
The exact aetiology of ASD remains unknown. Determining the genetic and cellular causes of ASD continues to be a major challenge in clinical research. To comprehensively investigate the underlying mechanisms, it is essential to develop novel rodent models that closely replicate the characteristics of human ASD. Previous studies have shown that certain environmental factors, such as exposure to VPA, may contribute to the development of ASD.17–20 Pregnant rats exposed to VPA (500 mg/kg) on gestational day 12.5 have been shown to give birth to offspring exhibiting autism-like behaviours.10, 21, 22
For the model of neonatal ischaemia or hypoxic brain injury, researchers have developed and reported several animal paradigms that replicate hypoxic or hypoxic–ischaemic conditions.23, 24 Studies have also demonstrated that the behavioural changes induced by hypoxia, such as impaired motor coordination and reduced novel object recognition, depend on both the timing and severity of exposure.25, 26 As is known, hypoxia disrupts several critical biological processes, including brain synaptogenesis, redox imbalance, glial cell activity, blood–brain barrier permeability, and the regulation and clearance of neuromediators.27–29 Perinatal hypoxic episodes have been linked to excitotoxicity, learning impairments, depression-like behaviours, altered spatial memory and neuronal cell death.30–34 Previous animal studies have also reported sex-specific differences, with male and female rats responding differently to hypoxic conditions and exhibiting varying degrees of behavioural deficits.11, 35 Male neonates are generally more susceptible to hypoxia–ischaemia and tend to display greater behavioural and cognitive impairments compared to females.36–39
In the present study, three paradigms were employed and compared to develop autism in rats: VPA, hypoxia and combined VPA plus hypoxia exposure. The current findings support the hypothesis that a hypoxic insult amplifies VPA-induced ASD-like behaviours in offspring rats. Attenuation and amplification in neurodevelopmental reflexes, social interaction, repetitive and stereotyped behaviours, anxiety, cognitive function and cerebellar and hippocampal morphology in rats by VPA, hypoxia and their combination were altered between the paradigms. Prenatal VPA exposure, as well as hypoxia alone and combined exposure, produced clinically relevant ASD-like behaviours in offspring, including reduced social interaction, increased anxiety, heightened repetitive behaviours in the open-field test, and cognitive impairments in the MWM test. These behavioural alterations are consistent with previous experimental studies with a few changes.21, 40–48 However, the combined VPA and hypoxia model exhibited more pronounced behavioural deficits and neuroanatomical disruptions, making it particularly suitable for studying the severity and spectrum of ASD phenotypes.
Neurodevelopmental reflex testing revealed that the VPA + hypoxia group displayed the most pronounced phenotype compared with the groups exposed to VPA or hypoxia alone. This finding further supports that hypoxia exacerbates neurodevelopmental delays in rat offspring prenatally exposed to VPA, aligning with previous reports. 15
The current study demonstrated that prenatal exposure to VPA, hypoxia, and their combination led to significant behavioural alterations, as observed in behaviour tests. These results expand upon earlier findings from the classical VPA-only model, which reported reduced locomotor activity but did not explore other behavioural aspects, such as grooming or anxiety-related defecation. 49 Our study provides a more comprehensive behavioural profile and highlights the compounding effect of dual insults.
In the MWM test, all experimental groups exhibited cognitive impairments compared with the controls. The escape latency was significantly longer in the VPA, hypoxia and VPA + hypoxia groups, indicating impaired spatial learning. The most severe deficits were observed in the VPA + hypoxia group, suggesting an additive effect of the combined prenatal insults. Moreover, the probe trial showed a reduced number of target-zone crossings in the VPA + hypoxia group, reflecting poor memory retention. While previous VPA-only studies also reported learning deficits, 49 the current findings emphasise that hypoxia alone impairs cognition and that its combination with VPA exacerbates these effects, thereby enhancing the construct validity of this multifactorial ASD model.
Collectively, the comparison reveals that, while the classical VPA model reliably demonstrates deficits in locomotion and learning, the addition of hypoxia in the present study produces a more robust and multifaceted ASD phenotype. The expanded behavioural assessments, including grooming and faecal output, offer deeper insight into ASD-like features, while the pronounced cognitive impairments observed in the combined model further support its suitability as a high-fidelity animal model for investigating both behavioural and neurocognitive aspects of ASD.
Previous studies using propionic acid (PPA) to induce ASD-like behaviours examined baseline activity before treatment and found no initial differences between the PPA and control (phosphate-buffered saline) groups, allowing researchers to attribute subsequent changes directly to the treatment. 50 While that model focused on verifying treatment-induced effects under controlled conditions, our study instead models the impact of prenatal environmental insults that more closely mimic real-life risk factors for ASD.
Histopathological analysis revealed PC degeneration and loss in the PC layer, basket cell degeneration in the cerebellar ML, and neuronal degeneration in both the cerebral cortex and hippocampus in the VPA, hypoxia and VPA + hypoxia groups. These findings are consistent with earlier studies.15, 51–53 Cerebellar damage has been associated with neurodevelopmental delays, a key feature in autistic children, as demonstrated in multiple clinical and experimental investigations.17, 41, 54–56 The combined VPA + hypoxia group showed more severity of histopathological findings (PC degeneration and PC loss in the cerebellum) compared to the VPA and hypoxia group. The cerebellum plays a crucial role in higher-order brain functions, and lesions within its cortico-subcortical connections can lead to motor, verbal, cognitive, emotional and social impairments.57–61
Overall, both the VPA and hypoxia models demonstrate that specific interventions can elicit ASD-like behaviours. However, our findings go further, showing that the combination of VPA and hypoxia produces more severe and complex behavioural and histological alterations than either insult alone. This combined model is, therefore, especially valuable for exploring how multiple early-life risk factors interact in ASD and for testing potential therapeutic strategies targeting these multifactorial origins.
Conclusions
Our results demonstrate that combined prenatal exposure to VPA and hypoxia produces ASD-like conditions in rat offspring, characterised by more severe neurobehavioural changes and histopathological lesions compared with the individual exposures alone. This combined model could serve as a novel and reliable animal model for ASD, closely mimicking postnatal conditions in humans. It holds promise for evaluating potent therapeutic interventions to prevent or mitigate autism-related neurodevelopmental deficits in the future.
Footnotes
Abbreviations
ASD: Autism spectrum disorder; G: Group; GD: Gestational day; H&E: Haematoxylin and eosin; HYP: Hypoxia; i.p.: Intraperitoneal; MWM: Morris water maze; OFT: Open field test; VPA: Valproic acid.
Acknowledgements
The authors are thankful to Dr S. Thanikachalam, Director, Centre for Toxicology and Developmental Research (CEFTE), Sri Ramachandra Institute of Higher Education and Research (SRIHER), for providing all necessary infrastructure, facilities and resources leading to the successful completion of the study. The authors would like to express their gratitude to the study team members, especially Mr R. Dilip, for blind observation and the CEFTE Team for their support and technical assistance.
Authors’ Contribution
Dr Shonam Tamrakar: Writing, visualisation, validation, investigation, methodology, formal analysis, conceptualisation.
Dr Veeraraghavan Gayathri: Writing, visualisation, validation, review and editing, supervision, resources, investigation and conceptualisation.
Dr Natesan Pazhanivel: Methodology and data curation.
Dr Suvarna Jyothi Kantipudi: Data curation.
Availability of Data and Materials
The data sets of the study will be available from the corresponding author on reasonable request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This preclinical study was funded by Sri Ramachandra Institute of Higher Education and Research (SRIHER), Chennai, India, under GATE ‘Young Faculty Research Grant 2022–2023’.
Patient Consent
Not applicable.
Statement of Ethics
The experimental protocol and all associated processes were endorsed by the IAEC (Institutional Animal Ethics Committee) (Approval number: IAEC/70/SRIHER/846/2023) of SRIHER, Porur, Tamil Nadu, India. Every effort was made to ensure animal well-being and minimal stress. All experimental procedures were carried out at CEFTE, a CCSEA (The Committee for Control and Supervision of Experiments on Animals) (Registration No. 189/PO/ReRcBi/S/2000/CCSEA) and adhering strictly to the CCSEA and ARRIVE guidelines.